
Abstract
This study investigates the distribution and conservation status of plant species in lentic wetlands of Mediterranean islands. Field data on vascular macrophytes were collected from the online national databases of the “Conservation of the island wetlands of the Mediterranean Basin” (MedIsWet). A comprehensive inventory of wetlands was conducted, and information on the main anthropogenic impacts and conservation priorities was compiled. In addition to all vascular hygro- and hydrophytes, we retained information on those species typically adapted to environments with intermittent or temporary wetness. A significant knowledge gap regarding the distribution and conservation of plant species in Mediterranean wetlands was found. A lower endemism rate was observed particularly in coastal wetlands. However, approximately 25% of species, mainly with relatively wide distribution, were identified as endangered or with unknown conservation status. Including all macrophytes adapted to seasonal wetness is crucial when considering wetland conservation efforts. The research emphasised the importance of considering diverse wetland types, including seasonal and permanent, natural and artificial, for effective plant conservation. Artificial wetlands emerged as potential habitats with considerable biodiversity conservation value. This study provides a comprehensive inventory of wetlands and valuable insights into the distribution, ecology, and conservation relevance of aquatic macrophytes in Mediterranean islands. The research enhances our understanding of biogeographic patterns and processes, offers critical information for the management and conservation of Mediterranean island wetlands, and presents a replicable approach that can be applied to other wetland contexts.
Similar content being viewed by others


Introduction
The Mediterranean Basin is a mega biodiversity hotspot (Cañadas et al. 2014), and aquatic plants are an important, specific element in maintaining this status (Chappuis et al. 2012; Rodríguez-Merino et al. 2019).
Although generally assumed to have broad world distributional ranges (Murphy et al. 2019), aquatic plants are often listed in the Annexes of the Habitats Directive (92/43/EEC) and in the ‘Top 50 Mediterranean Island Plants’ to be urgently conserved (Pasta et al. 2017). Apart from the several ecosystem services provided, wetlands support a disproportionate amount of global diversity of vertebrates, invertebrates, and plants (Balian et al. 2008; Chambers et al. 2008), which are mainly threatened by habitat and vegetation degradation due to land reclamation and intensive use, and the introduction of exotic plants (Basset et al. 2006; Mayoral et al. 2018; Fois et al. 2021). Consequently, numerous studies on Mediterranean biodiversity have been developed in wetlands, even though they are mostly limited to specific types, such as coastal and riverine environments (Cuena-Lombraña et al. 2021; Barone et al. 2022). Several authors have instead pointed out the urgency of considering other types of wetlands that are home to rare and/or threatened plants, such as temporary ponds (Lumbreras et al. 2016; Bagella 2023), springs (Fernández-Martínez et al. 2023) or water dripping crevices (Sciandrello et al. 2023). Some artificial wetlands are also emerging as interesting environments for biodiversity conservation (e.g. De Martis et al. 2016; Briggs et al. 2019).
Aquatic plants and, in particular, macrophytes (i.e., large enough to see with the naked eye) are generally defined as “aquatic photosynthetic organisms that actively grow permanently or periodically submerged below, floating on, or up through the water surface” (Murphy et al. 2019). If permanent inland waterbodies self-evidently offer potential macrophyte habitat, temporary and ephemeral waterbodies are more open to discussion regarding their status as macrophyte habitats. Murphy et al. (2019) include macrophytes of temporary and ephemeral systems that can tolerate periodic drought, but only if they show physiological requirement for partial or total inundation during their life-cycle, such as for initiating propagule production or germination. Examples in this sense are common in the Mediterranean floras including, for instance, several species among the genera Isoëtes, Ranunculus or Juncus (Lanfranco et al. 2016; Sciandrello et al. 2016; Bolpagni et al. 2018). Despite their mentioned importance, updated and reliable data on the presence and distribution of aquatic macrophytes in the Mediterranean is fragmented. In Italy, a recent comprehensive checklist has been presented (Bolpagni et al. 2018), but it was lacking from recent field reports, and a comprehensive distributional and ecological comparison among different wetland environments. Other studies were focusing on specific sites or problems, such as the increasing invasiveness that is threatening wetland ecosystems (Desfayes 2008; Mayoral et al. 2018; Troia et al. 2020). To our knowledge, no study has collected recent field data on the occurrence of aquatic macrophytes and compared them among different Mediterranean islands, allowing the investigation on their distribution, ecology and conservation relevance.
In this paper, we retrieved all information on vascular macrophytes from the online databases of the “Conservation of the island wetlands of the Mediterranean Basin” (MedIsWet) MAVA project. In particular, we focused on the French, Italian and Maltese islands, especially Corsica, Sardinia, Sicily, Malta and other surrounding minor territories, with the aim to: (1) compile a list of vascular plants recorded in a representative set of lentic waters across multiple Mediterranean islands; (2) analyse the main geographic, environmental and anthropogenic factors influencing the composition of macrophytes in lentic wetlands; (3) define and compare their different life and chorological forms, and their conservation status across various wetland types and (4) identify, for each wetland type, the most significant plant species in terms of their representativeness and conservation relevance.
Methods
Study regions: the MedIsWet national databases
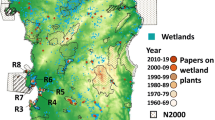
We considered the wetlands of the main W-Mediterranean islands: Sicily (surface area of 25,426 km²), Sardinia (23,821 km²), Corsica (8,679 km²), Malta (246 km2) and 11 satellite minor islands. They all are surrounded by the Mediterranean; more precisely, Sardinia and Corsica are situated between the Iberian and Italian peninsulas, Sicily is at the extreme of the Italian peninsula and Malta is located in the Central Mediterranean, 290 km off the coasts of northern Africa and 96 km from Sicily (Fig. 1). All the investigated islands are considered as ‘continental’ or ‘chersogenous’, being originated by the split from mainland (Greuter 2001). The relatively recent palaeogeographical history of Sardinia and Corsica, and of Sicily and Malta, are parallel, as they were alternately connected during the last eustatic cycles (ca. 5 Ma to 20 Ka) and thus have many floristic elements in common (Bonanno 2013; Lanfranco et al. 2013; Fois et al. 2022b). Shared, or phylogenetically related, species between the four investigated areas are also frequent due to less recent connections facilitated by the regression of the sea during the so-called ‘Messinian salinity crisis’ (6 − 5 Ma; Bocquet 1978). Repeated gene flows between many species shared by the Mediterranean islands are also possible through long-distance dispersal events, mainly mediated by migratory birds and sea currents (e.g. Kadereit et al. 2005; Martínez-López et al. 2020; Fois et al. 2022c).
In Sicily and Malta, most of the lentic wetlands are represented by small artificial farm ponds; some of them bearing a high cultural, landscape and biodiversity value (Panzeca et al. 2021; Tavilla et al. 2023a). In Sardinia and Corsica, more than half of wetlands are natural, which make them even more critical for biodiversity and conservation value. Accordingly, nine Ramsar sites have been designated in Sardinia, five in Corsica, three in Sicily and two in Malta (https://rsis.ramsar.org). The designation of these sites was mainly based on the presence and nesting of several aquatic birds, and they encompass large coastal areas; the only exceptions were the two inland and temporary wetlands of “Tourbière de Moltifao” and “Mares temporaires de Tre Padule de Suartone” in Corsica. However, numerous plants, which are mostly characteristic of small, often unprotected ponds or even rock pools, have been repeatedly cited as one of the most striking conservation gaps in conservation of the Mediterranean flora (e.g. Bagella et al. 2016; Minissale and Sciandrello 2016; Lanfranco et al. 2016, 2020).
Abiotic and biotic information on wetlands of these islands is available from the following MedIsWet national databases (accessed in May 2023): France: https://franceiswet.fr; Malta: https://www.maltawetlands.org; Italy: https://italiaiswet.it. The three datasets can be exported and can be freely used under the Attribution-Share Alike 4.0 International licensing (CC BY-SA 4.0); they were structured in an identical and compatible manner to allow them to be merged. As regards the cartographic inventory available from these datasets, it was created in Google Earth Pro (ver. 7.3.6.9345) by identifying and digitising all lentic wetlands visible on high-resolution satellite images with a surface ≥ 0.1 ha. In case of uncertainty regarding the dimension of a wetland, the largest potential polygon was defined according to seasonal fluctuations, inundation and vegetation cover that was observed in the available Google Earth imagery time series (Perennou et al. 2018a; Fois et al. 2021). According to such inventory, wetlands are very representative environments in all the study sites. The largest number of wetlands was recorded for Sicily and its surrounding islets with 11,410 lentic wetlands, followed by Sardinia (2,558), Corsica (512), and Malta (91). Of these 14,571 lentic wetlands, 1,281 sites were inventoried in the field as part of the MedIsWet project for the Italian, French and Maltese islands between 2018 and 2023. The sample sites were randomised in a stratified manner, trying to proportionally cover artificial/natural and coastal/inland environments (Fig. 1). Field data on these wetlands encompass several aspects, including the presence/absence of species and other environmental and anthropogenic factors, which will be briefly described in the following sections. For this study, we considered all 1,281 surveyed sites. Most of the data used for the subsequent analyses come from the databases listed above and are reported in the supplementary material (Supplementary material 1).

Violin plots of the 1,281 target wetlands by a, inland (778 sites) and coastal (503), and b, by permanent (864 sites) and seasonal (417) wetlands. Jittered points show individual conservation status scores by artificial (581 sites) and natural (700) wetlands. Point colours in the map c, are for natural (green) and artificial (violet) wetlands
Aquatic plant data and selection criteria
All the floristic data were retrieved from the MedIsWet national databases, which were compiled following the Rapid Assessment protocol (Tomàs-Vives 2008). Briefly, the protocol does not include permanent plots for systematic and repeated visits but only at least one field monitoring that attempted to cover all the accessible areas within the wetland delineations. Consequently, the floristic data were used more cautiously as simple records of occurrence and not to infer species abundances at each investigated site. The inventorying protocol considered all vascular plants found within wetland boundaries, including several terrestrial ones, which are nevertheless also common along wetland banks and saturated soils. We have unified the taxonomy and nomenclature of all vascular plant taxa at species and subspecies level (henceforth, species) across the three datasets according to the Euro + Med PlantBase (http://europlusmed.org). To extract aquatic plants from the initial list reported in each database, we followed the checklist of Italian aquatic plants of Bolpagni et al. (2018). For those species absent in that list, we applied the same method based on: (1) the biological form as reported by Pignatti et al. (2017), selecting all the “hydrophytes” sensu Raunkiær (1934); and (2) the Ellenberg ecological indicator for “humidity” (U) ≥ 8. According to Baattrup-Pedersen et al. (2005) all species with an Ellenberg’s humidity value U ≥ 10 should be considered as aquatic plants, being ecologically strictly related to “aquatic” habitats with permanently saturated substrates and therefore influenced by periodical submersion and/or by constant saturation of colonised sediments. Bolpagni et al. (2018) have also included some species with Ellenberg indicator values equal to 9 (e.g., Montia, Elatine, and Juncus genera), since new data on their ecology have permitted confirming their “aquatic life strategy”. According to Bolpagni et al. (2020), we also included species with U = 8, since they are likely regionally strictly exclusive to ephemeral wetlands, thus more cautiously to be considered as “wet and aquatic plants”. We used averaged moisture/humidity values reported for Italy (Guarino and La Rosa 2019) and France (Julve 2015), also updated and resumed in Tichý et al. (2023).
Geographic, environmental and anthropogenic factors
To investigate the regional arrangement of aquatic plants, a series of factors were used as explanatory variables, focusing on geographic, environmental conditions and anthropogenic factors (Table 1). Descriptive data include both abiotic and biotic information from cited literature and from original field surveys. To the information available from the MedIsWet national databases, we also included climatic information retrieved from Chakraborty et al. (2021) and the elevation (Jarvis et al. 2008). The latter values were extracted by averaging the centroids of the pixels within each wetland boundary.
Factors of plant species composition
To determine how the species composition relates to our set of abiotic variables, we used non-metric multidimensional scaling (NMDS). For this purpose, species found only once or twice and sites with less than three species were omitted from the data set beforehand to enhance the accuracy of statistical analyses and to reduce statistical noise (Gathof et al. 2022). Consequently, from an initial set of 1,281 field-inventoried sites, we restricted the analysis to a final set of 516 wetlands (Table S1 - Supplementary material 1).
First, cluster analysis was performed to classify the plots according to their floristic similarity. NMDS using Bray–Curtis pairwise distance on species presence–absence data was performed using the ‘metaMDS’ function in the vegan package of R (Oksanen et al. 2007). A minimum stress of 0.19 was achieved after 20 iterations when the model converged indicating that the data could be adequately represented by the first three ordination axes (k = 3). Then, we incorporated the explanatory variables (Table 1) into the NMDS analysis with the ‘envfit’ function using a permutation approach (999 permutations), the function automatically treats all categorical variables as factors and the rest as vectors (Table 1). In this procedure, fitted vectors depict how environmental factors correlate with the NMDS ordination (Cosentino et al. 2013). Fitted vectors are calculated using a linear model in which the value of an environmental factor is the dependent variable, and the NMDS axes are the independent variables. The direction of each vector in the ordination indicates the most rapid rate of change of an environmental factor, and the length of each vector is proportional to model fit, R2. A p value was calculated for each environmental factor using 10,000 Monte Carlo permutations. To reduce multicollinearity and to select the most insightful vectors, the “corrplot” package (Wei et al. 2017) was used to calculate Spearman’s correlation coefficient (r) and, among high correlated vectors (|r|>= 0.7), we retained those with the highest R2 obtained through ‘envfit’. Analysis of similarities (ANOSIM; Clarke 1993) with Bray–Curtis distance and 999 permutations was used to test for significant differences in species composition among wetland’s main factors.
Variation of aquatic macrophytes features
We assessed how aquatic macrophytes were distributed across the following features: (1) main plant life-forms; (2) chorology; and (3) conservation status. These three characteristics were first retrieved in R environment using TR8 package (Bocci 2015) and then manually integrated by literature data. Specifically, Corsican (Jeanmonod and Gamisans 2007), Italian (Pignatti et al. 2017–2019) and Maltese (Weber and Kendzior 2006) floras were consulted for life forms described by Raunkiær (1934) (i.e., therophyte, hydrophyte, hygrophyte, geophyte, hemicryptophyte, chamaephyte and phanerophyte) and chorological forms, which were simplified in: (Sub)cosmopolitan (distributed in more than one biogeographic realm), Palearctic, Mediterranean, Endemic (to only one up to all the target islands) and Introduced. Conservation status assessments were considered at global and regional level, accounting for assessments reported in the official IUCN portal (https://www.iucnredlist.org) as well as national red lists of Italy (Conti et al. 2005; Rossi et al. 2013, 2020), France (Largier 2019) and Malta (Schembri and Sultana 1989). The information has been simplified as follows: (1) Endangered at global/regional level: all species under Critically endangered (CR), Endangered (EN) and Vulnerable (VU) IUCN categories at the two respective geographical scales; (2) Minor risk: all species under Near Threatened (NT) and Least Concern (LC) IUCN categories; and (3) Unknown: all Data Deficient (DD) and not yet assessed, both at global and at local scale. The conservation status of introduced plants was not recorded.
Plant features were compiled for a subset of species. Apart from the above-mentioned issue regarding introduced species, we discarded those for which we were unable to determine their distribution preference among the following preferences: natural coastal wetlands, natural inland wetlands, artificial wetlands, ubiquitous. Species were assigned as typical or characteristic to one of these categories if they had at least 85% occurrence in one of them or, for ubiquitous ones, at least 10% occurrence in all types. To determine the extent of the relationship between a taxon and its wetland category, we employed the test statistic ‘IndVal.g’ implemented in the multipatt function of the R package ‘indicspecies’ (version 1.7.13; De Cáceres et al. 2010). This statistic yields an ‘Indval’ score ranging from 0 (indicating no association) to 1 (representing the highest level of association). The statistical significance (p < 0.05) of the value of each taxa was assessed by a permutation procedure that compared the observed test statistic with a distribution obtained by permuting the data (De Cáceres and Legendre 2009). All packages were run in R software ver. 4.2.2 (R Core Team, Vienna, Austria).
Results
Factors of plant species composition
Species composition differed mainly between coastal and inland wetlands (ANOSIM, R = 0.265, p = 0.001), secondly between natural and artificial (ANOSIM, R = 0.220, p = 0.001), among islands’ origin (ANOSIM, R = 0.173, p = 0.001) and, lastly, between permanent and seasonal wetlands (ANOSIM, R = 0.098, p = 0.001).
In the NMDS analysis for species composition, all variables included except Surface, showed significant relationships with aquatic plant composition (Table S1 – Supplementary material 2). However, Spearman’s correlation coefficients revealed a high correlation (|r|>= 0.7) among variables (Fig. S1 – Supplementary material 2). As a result, only Conservation status, Mean winter temperature (Tave_wt) and Mean winter precipitation (PPT_wt) were retained as the main explanatory vectors. Based on R2-values, the two climatic variables had the strongest relationship with species composition while less effect was attributable to the wetland conservation status (Table 2). With regard to the latter attribute, it is worth noting that the conservation status of natural inland wetlands is higher (i.e. better) than coastal ones (p < 0.01; Fig. S2 - Supplementary material 2).
Within groupings, both coastal and inland, and artificial and natural wetlands, differentiate along NMDS axis 1 by climatic variables (Tave_wt and PPT_wt) and along the NMDS axis 2 by the Conservation status (Fig. 2a,b).

Non-metric multidimensional scaling (NMDS) plot for axes 1 and 2, with ellipses and colours depicting artificial and natural a, and inland and coastal b, wetlands
Variation of species composition among wetland types
Of the 295 aquatic plants that were recorded in the study areas (Table S2 - Supplementary material 1), we analysed features for 215 species that were attributable to only one of the following types: ubiquitous, inland natural, coastal natural and artificial (Table S3 - Supplementary material 1); 38% of them were ubiquitous, i.e. found in all the following types: (1) coastal natural, (2) inland natural, and (3) artificial wetlands (Fig. 3). The percentage of species shared between artificial and only one natural category was small (3–4%), while most species living in artificial wetlands are ubiquitous. Among natural types, only 9% of species are shared between inland and coastal wetlands, while a higher percentage (18% and 28%, respectively) is exclusive to one of those two categories. For artificial wetlands, only three indicator species were found, while more indicator species were retained for coastal and inland lentic wetlands.

Venn diagram indicating the degree of aquatic plant species overlap among inland and coastal natural wetlands and artificial ones. Indicator species with Indval > 0.3 and p < 0.05 were reported for each wetland category. Asterisks indicate the p-values after 999 permutation tests implemented by the multipatt function of the R package ‘indicspecies’: (*) p < 0.05; (**); p < 0.01; (***); p < 0.001
Generally, plant features were equally distributed among wetland preferences, with some exceptions (Fig. 4). As regards the chorology, no endemics were typically found in artificial wetlands, while the introduced category is dominant among coastal wetlands. The highest number of endemic plants were detected in inland wetlands. Among life forms, chamaephytes are exclusive to coastal natural wetlands whereas phanerophytes are found in coastal natural wetlands or are ubiquitous. Hydrophytes and therophytes are the most important life forms in artificial wetlands but are well represented also in other types of wetlands. Plants typically present in artificial wetlands are at minor risk of extinction or not yet assessed. Instead, species endangered at global level are mainly located in inland wetlands and in a lesser extent are ubiquitous. As already mentioned, ubiquitous plants are the majority, and this is also the trend among different plant features.

Composition of aquatic vascular macrophytes among the three different plant features
Discussion
Plants recorded in aquatic environments
Compared to the around 4,080 specific and subspecific plant taxa reported in the MedIsWet databases (Fois et al. 2022a), only 7% have been identified as ‘wet and aquatic vascular plants’ (i.e. 295 species). This number is, however, higher than the 270 species reported for the whole of Italy (Bolpagni et al. 2018). The main reason is that we included all species with U = 8: this is a debated issue, which can be argued by considering the local drought conditions that characterise Mediterranean islands and, more generally, southern Mediterranean territories. If plants with U = 8 are not strictly ecologically dependent on wet habitats on a global or even national scale (see, for example, climatic variability in Italy), the same species are instead refugees in such wet conditions at the southern edges of their distribution. For example, many Lythrum species are largely found in the grasslands of temperate Europe, while they often are strictly linked to the temporarily inundated soils of the Mediterranean study areas (Brullo et al. 2022; Tavilla et al. 2023a). Ensuring the conservation of these environments would contribute to the preservation of the current genetic and distributional range of these plants, which, at the moment, are not considered as aquatic on a global scale, but may be more strictly aquatic in the future, when more extensive drought conditions are expected (Essa et al. 2023).
Only seventeen species were found as endemics (Table S3 - Supplementary material 1). This is in line with what has been reported, especially for the Mediterranean Basin (Hobohm and Bruchmann 2011; Cuena-Lombraña et al. 2021). At global ecozone scale, macrophyte endemism is pronounced, but with a significant difference between the richest (Neotropics) and poorest (Palaearctic) (Murphy et al. 2019). Among different environments, the number of endemics in wetlands is also relatively low compared to endemics of rocky habitats and screes, grasslands, scrubs and heaths, or forests. It was assumed that this is due to the young age of wetlands and other intrinsic and extrinsic factors that characterise aquatic plants, such as the generally high dispersal capacity or the tendency to develop vegetative reproduction systems (Hobohm and Bruchmann 2011; Cuena-Lombraña et al. 2021). A low number of introduced species was found, although an expectable increase is warring in view of the high economic and ecological damage that some of them have already proven to cause, as in the case of the invasive alien species of Union Concern Hydrocotyle ranunculoides or Pontederia crassipes (Brundu 2015; Troia et al. 2020; Cambria et al. 2023). Our data suggest that, apart from artificial lentic wetlands, these potential pests are likely preferring natural coastal ones, probably due to warmer temperatures that are more similar to their native conditions. These results are in accordance with recent papers (e.g., Taylor et al. 2021; Martínez-Megías and Rico 2022), that include the proliferation of invasive alien species as one of the major threats to the biodiversity decline of Mediterranean coastal wetlands, which is increasingly facilitated by the expansion of urban areas (Perennou et al. 2018b). None of the species found prevalently in artificial wetlands were endangered, while most of them have therophytic or hydrophytic life forms. This pattern might be related to the higher colonisation ability of such life forms of recently disturbed environments (Sciandrello et al. 2015), while trees and small shrubs, which are likely preferring natural conditions, might be considered as indicators of wetland conservation status. The latter suggestion is in line with the results obtained in the “Bestina” riverside area in Tuscany (Fiaschi et al. 2023), where trees and shrubs were evaluated as the most ‘conservative’ species according to an expert approach. The same authors thus suggested them as indicators of good environmental quality. In contrast to our results, hydrophytes were found in the “Bestina” riverside to be the best indicators of good environmental quality, whereas in our case they were equally found in both disturbed and undisturbed wetlands and therefore poorly effective in indicating the naturalness and conservation status of different wetlands. This confirms again that the information provided by life forms, which is often used for environmental assessments, should be complemented with other features and cautiously adapted to each specific context. It is also worth of mention that our results were based on only lentic waterbodies, thus not considering patterns along rivers and streams. For instance, the endemic Mentha requienii susp. requienii has been recorded for this study only in lentic artificial waters, although it was more commonly reported for mountain streams and waterfalls.
The identified indicator species for each wetland category are confirming the above discussed general trends of chorological and life forms. As expected, the few indicator species for artificial wetlands were plants with a large ecological valence and geographical distribution. Differently, among natural wetlands - especially for inland ones - there were cases of narrowly distributed and/or endangered species among indicators, such as the locally endangered Ranunculus ophioglossifolius or Isoëtes histrix for inland waters or Triglochin bulbosa subsp. barrelieri for coastal ones. In accordance with other authors (e.g. Lukács et al. 2013; Angiolini et al. 2017b), this underlines once again that the conservation value and naturalness of wetlands can be indicated by the presence of some - but not all - threatened species.
Factors of species composition
The main factors of species composition were partly expected in accordance with previous observations. Differences between the plant communities of coastal and inland wetlands have been largely explained by various limiting factors, such as water and soil salinity or other climate-related ones (e.g. Angiolini et al. 2017a; Lanfranco et al. 2020). Anthropogenic disturbance is another important predicted condition influencing plant communities by facilitating, for instance, most alien life-forms and both native and alien therophytes (Marini et al. 2012). However, these factors were found to be more important than the geographical separation between islands and between seasonal and permanent wetlands. Geographical isolation supported by the island biogeography theory is considered one of the most universally accepted hypotheses in explaining species distribution and composition in both oceanic and Mediterranean insular contexts (Fois et al. 2020; Schrader 2021; Médail 2022). However, aquatic plants are often able to overcome the physical barrier of the sea due to their greater ability to disperse - even by long-distance means - such as ornithochory or thalassochory (Mahanand and Behera 2019; Arjona et al. 2020; Ruzzier et al. 2021; Fois et al. 2022c). The fact that hydroperiod was found to be less important than other factors was surprising, considering the large body of supporting evidence (e.g., Fernández-Zamudio et al. 2018; Bagella et al. 2018). First, our research was not type-specific, but included almost equally all wetland categories (there were 216 seasonal wetlands out of the 516 sites used in the NMDS analysis; Fig. 1). Secondly, specific studies on seasonal wetlands require different monitoring approaches, involving repeated field visits encompassing all seasons. Furthermore, the existence of empirical evidence has substantiated the presence of aquatic species typically associated with permanently flooded soils and dry grassland species, within seasonally flooded environments. Consequently, the reliability of discerning these species based solely on their presence or absence becomes compromised. Thus, it is imperative to employ alternative statistical methodologies that incorporate considerations of temporal dynamics and frequency of occurrence to ensure precise identification of indicator species in these particular contexts (Bagella et al. 2009; Minissale et al. 2017).
As expected, precipitation and temperatures are among the main drivers of aquatic species composition. This can be reflected both directly and less directly by the differentiation between wetland categories (i.e., coastal vs. inland and natural vs. artificial), as it can be assumed that artificial dams and agricultural ponds were placed under conditions of higher water availability and fertility. In contrast, the conservation status of wetlands was found to be weakly correlated with the climate-related variables, indicating that wetlands in different climatic conditions can be in a similar conservation status. Both results have conservation implications related to climate change. First, the species composition is related to climatic features, indicating that its change might drive species composition. Moreover, as discussed above, it is likely that alien species currently prefer the coastal warmer conditions, confirming that rising temperatures could favour their expansion to inlands (Fois et al. 2020; Lozano et al. 2023). Another conservation consideration is that, despite the coastal prevalence of protected wetlands, the status of inland wetlands is equally variable and therefore deserving of similar attention. This is exacerbated when considering that several species, including those at risk, are not only roughly shared between natural coastal and inland wetlands, but are also present in artificial ones. As suggested by other authors (e.g., Panzeca et al. 2021), artificial wetland habitats could be the subject of a monitoring program as potential introduction sites for alien propagules but also suitable areas for rare and/or endangered species. Moreover, the conservation status of a large part of species typically found in coastal and artificial wetlands is unknown. This is possibly because of their wider distribution range and presenting difficulties in their assessment, confirming the need to fill gaps in global conservation status assessments of widely distributed species (Mounce et al. 2018; Fanfarillo et al. 2020; Fois et al. 2022b).
Management and conservation implications
Our data filled gaps over a relatively large area and set of environments through a relatively small effort, quantifiable in about two/three years of fieldwork by eight people (two per region/island) and about as many for data processing, refining and sharing. Similar efforts can be made to respond to the same surveys in other areas or for other environments in the same territories. Filling these knowledge gaps can substantially help conservationists to implement more effective actions aimed at the most imperative species and/or habitat types.
The presence of several threatened - at least locally - but widely distributed plants, confirms the need to manage wetland conservation on a larger scale than in other, more isolated but less fragmented contexts of similar conservation importance, such as mountain cliffs or true small islands (Schöpke et al. 2019). At a larger scale, international agreements and action plans, such as the Ramsar Convention on Wetlands, the European Water Framework Directive (2000/60/EC) or, in general, all international collaborations between experts are crucial. At a smaller scale, wetlands can be considered not as isolated bodies, but as integrated systems that could be geographically delimited within each river basin sector. At the watershed scale, the study of plant compositions and how they vary in space and time can be a crucial indicator of ecosystem functioning, which, however, needs to be complemented by further investigations into other equally important components, such as pollution, non-vascular plants, zoological communities, and, most importantly, how they all influence each other (Fernandez et al. 2014; Palmas et al. 2022). Recognising that it may take an infinite amount of resources to fulfil such knowledge, research, and this one in particular, has highlighted which and how they are most likely to need special attention. In particular, our analysis has identified, among biological forms, that therophytes and hydrophytes are most likely to be considered as potential invasives, while trees and shrubs are those to be promoted among native vascular species. This latter finding confirms the importance of considering the potential dramatic changes in riparian and aquatic biota caused by clear-cutting practices around wetlands (Angiolini et al. 2023). Our analyses confirmed that coastal wetlands are an important source of propagule pressure, but also highlighted the importance of monitoring artificial ones in this regard. Also artificial wetlands harbour endangered plants, which gives them a double importance for monitoring perspectives and a controversial role in terms of management (De Martis et al. 2016; Briggs et al. 2019; Panzeca et al. 2021).
With global warming, wetlands will continue to play an even more important role as refugia for many species, some of which are already facing critical conditions. Our results confirmed that climatic factors, namely precipitation and temperature, are central for the plant composition of Mediterranean wetlands. The focus on aquatic plants could be extended to those that are not considered as such globally, like the plants considered here with U = 8, which live mainly in ephemeral wetlands. Alien plants are instead likely to be favoured by global warming, as they were found to prefer warmer conditions, which may be more widely present in the future. Special monitoring and, when possible, eradication programmes could begin in coastal areas where species of Union Concern are already present, but more attention to occasional or potential new arrivals could help prevent the considerable costs of delayed actions (Lozano et al. 2023).
Conclusions
This study provides the first comprehensive overview of the floristic composition of the wetlands of the central Mediterranean islands, offering insights into what is known and neglected to date. The relatively large number of 295 recorded vascular species includes a questionable list of species not unanimously considered as aquatic plants. However, these species are ecologically highly dependent on wet conditions, especially in the seasonally dry environments that characterise the Mediterranean islands, and the preservation of wetlands is therefore crucial for their survival over their current entire distributional range.
As previously suggested, the information provided by life forms and chorotypes can be used to determine ecological value, although generalisations in this sense must be interpreted with caution, as many other aspects might influence this trend. Among other information, our data confirm the low endemicity of wetland floras, which is validated by the poor influence of island origin as a factor in species composition. Although natural inland wetlands had already been denoted as crucial areas for biodiversity conservation, we highlighted the presence of several endangered macrophytes also in artificial wetlands. More generally, the conservation value and naturalness of wetlands can be indicated by the presence of some - but not all - threatened species. As regards invasive alien species, these were more frequently recorded in both artificial and coastal natural wetlands, suggesting a need for special attention on these environments for their control and possible expansion facilitated by global warming and urbanisation.
Further investigations are needed to provide a more complete overview of the distribution, ecology, and interactions between wetland species. Amongst other shortcomings, the data provided here need further field investigations, particularly repeated in different seasons, to overcome limitations in examining the factors that determine species abundances. Furthermore, running waters are still neglected and could be included in the same dataset in the future. However, this study certainly represents a starting point for more informed wetland conservation in the future. The data presented here, and all other information reported in the MedIsWet national databases is freely available, precisely with the aim of laying the foundations for a more inclusive and constructive network of wetland conservationists.
References
Angiolini C, Nucci A, Landi M, Bacchetta G (2017a) What drives riparian plant taxa and assemblages. Mediterranean Rivers? Aquat Sci 79:371–384. https://doi.org/10.1007/s00027-016-0503-2
Angiolini C, Viciani D, Bonari G, Lastrucci L (2017b) Habitat conservation prioritization: a floristic approach applied to a Mediterranean wetland network. Plant Biosyst 151:598–612. https://doi.org/10.1080/11263504.2016.1187678
Angiolini C, de Simone L, Fiaschi T, Cifaldi GP, Maccherini S, Fanfarillo E (2023) Detecting the imprints of past clear-cutting on riparian forest plant communities along a Mediterranean river. River Res Appl 39:1616–1628. https://doi.org/10.1002/rra.4152
Arjona Y, Fernández-López J, Navascués M, Alvarez N, Nogales M, Vargas P (2020) Linking seascape with landscape genetics: Oceanic currents favour colonization across the Galápagos Islands by a coastal plant. J Biogeogr 47:2622–2633. https://doi.org/10.1111/jbi.13967
Baattrup-Pedersen A, Friberg N, Larsen SE, Riss T (2005) The influence of channelization on riparian plant assemblages. Freshw Biol 50:1248–1261. https://doi.org/10.1111/j.1365-2427.2005.01383.x
Bagella S (2023) Which perspectives for Mediterranean temporary ponds in the European Union in the third millennium? Biodivers Conserv 32:839–849. https://doi.org/10.1007/s10531-023-02549-3
Bagella S, Caria MC, Farris E, Filigheddu R (2009) Spatial-time variability and conservation relevance of plant communities in Mediterranean temporary wet habitats: a case study in Sardinia (Italy). Plant Biosyst 143:435–442. https://doi.org/10.1080/11263500903187068
Bagella S, Gascón S, Filigheddu R, Cogoni A, Boix D (2016) Mediterranean temporary ponds: new challenges from a neglected habitat. Hydrobiol 782:1–10. https://doi.org/10.1007/s10750-016-2962-9
Bagella S, Caria MC, Beccarisi L, Zuccarello V (2018) Ecological responses of selected vascular plants to water chemistry parameters in habitat types 3120, 3130 and 3170* (Habitat Directive 92/43/EEC). Plant Biosyst 152:1338–1345. https://doi.org/10.1080/11263504.2018.1461697
Balian EV, Segers H, Lévèque C, Martens K (2008) The freshwater animal diversity assessment: an overview of the results. Hydrobiol 595:627–637. https://doi.org/10.1007/978-1-4020-8259-7_61
Barone G, Cirlincione F, Di Gristina E, Domina G, Gianguzzi L, Mirabile G et al (2022) An analysis of botanical studies of vascular plants from Italian wetlands. Ital Bot 14:45–60. https://doi.org/10.3897/italianbotanist.14.95072
Basset A, Sabetta L, Fonnesu A, Mouillot D, Do Chi T, Viaroli P et al (2006) Typology in Mediterranean transitional waters: new challenges and perspectives. Aquat Conserv: Mar Freshw 16:441–455. https://doi.org/10.1002/aqc.767
Bocci G (2015) TR8: an R package for easily retrieving plant species traits. Methods Ecol Evol 6:347–350. https://doi.org/10.1111/2041-210X.12327
Bocquet G (1978) The Messinian model: a new outlook for the floristics and systematics of the Mediterranean area. Candollea 33:269–287
Bolpagni R, Laini A, Stanzani C, Chiarucci A (2018) Aquatic plant diversity in Italy: distribution, drivers and strategic conservation actions. Front Plant Sci 9:116. https://doi.org/10.3389/fpls.2018.00116
Bolpagni R, Laini A, Buldrini F, Ziccardi G, Soana E, Pezzi G et al (2020) Habitat morphology and connectivity better predict hydrophyte and wetland plant richness than land-use intensity in overexploited watersheds: evidence from the Po plain (northern Italy). Landsc Ecol 35:1827–1839. https://doi.org/10.1007/s10980-020-01060-2
Bonanno G (2013) Adaptive management as a tool to improve the conservation of endemic floras: the case of Sicily, Malta and their satellite islands. Biodivers Conserv 22:1317–1354. https://doi.org/10.1007/s10531-013-0473-9
Briggs A, Pryke JS, Samways MJ, Conlong DE (2019) Macrophytes promote aquatic insect conservation in artificial ponds. Aquat Conserv: Mar Freshw 29:1190–1201. https://doi.org/10.1002/aqc.3157
Brullo S, Brullo C, Sciandrello S, Tavilla G, Cambria S, Tomaselli V, Ilardi V, Giusso del Galdo G, Minissale P (2022) The Plant communities of the Class Isoëto-Nanojuncetea in Sicily. Plants 11:1214. https://doi.org/10.3390/plants11091214
Brundu G (2015) Plant invaders in European and Mediterranean inland waters: profiles, distribution, and threats. Hydrobiol 746:61–79. https://doi.org/10.1007/s10750-014-1910-9
Cambria S, Azzaro D, Caldarella O, Aleo M, Bazan G, Guarino R, Torre G, Cristaudo AE, Ilardi V, La Rosa A et al (2023) New Data on native and alien vascular flora of Sicily (Italy): New findings and updates. Plants 12:1743. https://doi.org/10.3390/plants12091743
Cañadas EM, Fenu G, Peñas J, Lorite J, Mattana E, Bacchetta G (2014) Hotspots within hotspots: endemic plant richness, environmental drivers, and implications for conservation. Biol Conserv 170:282–291. https://doi.org/10.1016/j.biocon.2013.12.007
Chakraborty D, Dobor L, Zolles A, Hlásny T, Schueler S (2021) High-resolution gridded climate data for Europe based on bias‐corrected EURO‐CORDEX: the ECLIPS dataset. Geosci. Data J 8:121–131. https://doi.org/10.1002/gdj3.110
Chambers PA, Lacoul P, Murphy KJ, Thomaz SM (2008) Global diversity of aquatic macrophytes in freshwater. Hydrobiologia 595:9–26. https://doi.org/10.1007/978-1-4020-8259-7_2
Chappuis E, Ballesteros E, Garcia E (2012) Distribution and richness of aquatic plants across Europe and Mediterranean countries: patterns, environmental driving factors and comparison with total plant richness. J Veg Sci 23:985–997. https://doi.org/10.1111/j.1654-1103.2012.01417.x
Clarke KR (1993) Non-parametric multivariate analyses of changes in community structure. Aust J Ecol 18:117–143. https://doi.org/10.1111/j.1442-9993.1993.tb00438.x
Conti F, Abbate G, Alessandrini A, Blasi C (2005) An annotated checklist of the Italian vascular Flora. Palombi Editori, Roma, Italia
Cosentino BJ, Schooley RL, Bestelmeyer BT, Coffman JM (2013) Response of lizard community structure to desert grassland restoration mediated by a keystone rodent. Biodivers Conserv 22:921–935. https://doi.org/10.1007/s10531-013-0459-7
Cuena-Lombraña A, Fois M, Cogoni A, Bacchetta G (2021) Where we come from and where to go: six decades of botanical studies in the Mediterranean wetlands, with Sardinia (Italy) as a case study. Wetlands 41:69. https://doi.org/10.1007/s13157-021-01464-z
De Cáceres M, Legendre P (2009) Associations between species and groups of sites: indices and statistical inference. Ecol 90:3566–3574. https://doi.org/10.1890/08-1823.1
De Cáceres M, Legendre P, Moretti M (2010) Improving indicator species analysis by combining groups of sites. Oikos 119:1674–1684. https://doi.org/10.1111/j.1600-0706.2010.18334.x
De Martis G, Mulas B, Malavasi V, Marignani M (2016) Can artificial ecosystems enhance local biodiversity? The case of a constructed wetland in a Mediterranean urban context. Environ Manag 57:1088–1097. https://doi.org/10.1007/s00267-016-0668-4
Desfayes M (2008) Flore Vasculaire herbacée Des Eaux Douces et des milieux humides de la Sardaigne. Flora Mediterr 18:247–331
Essa YH, Hirschi M, Thiery W, El-Kenawy AM, Yang C (2023) Drought characteristics in Mediterranean under future climate change. npj Clim Atmos Sci 6:133. https://doi.org/10.1038/s41612-023-00458-4
Fanfarillo E, Latini M, Iberite M, Bonari G, Nicolella G, Rosati L et al (2020) The segetal flora of winter cereals and allied crops in Italy: species inventory with chorological, structural and ecological features. Plant Biosyst 154:935–946. https://doi.org/10.1080/11263504.2020.1739164
Fernandez S, Bouleau G, Treyer S (2014) Bringing politics back into water planning scenarios in Europe. J Hydrol 518:17–27. https://doi.org/10.1016/j.jhydrol.2014.01.010
Fernández-Martínez M, Barquín J, Bonada N, Cantonati M, Churro C, Corbera J et al (2023) Mediterranean springs: Keystone ecosystems and biodiversity refugia threatened by global change. Glob Chang Biol 30:e16997. https://doi.org/10.1111/gcb.16997
Fernández-Zamudio R, García-Murillo P, Díaz-Paniagua C (2018) Effect of the filling season on aquatic plants in Mediterranean temporary ponds. J Plant Ecol 11:502–510. https://doi.org/10.1093/jpe/rtx026
Fiaschi T, Fanfarillo E, Maccherini S, Bacaro G, Bonari G, Foggi B et al (2023) Effectiveness of different metrics of floristic quality assessment: the simpler, the better? Ecol Indic 149:110151. https://doi.org/10.1016/j.ecolind.2023.110151
Fois M, Podda L, Médail F, Bacchetta G (2020) Endemic and alien vascular plant diversity in the small Mediterranean islands of Sardinia: drivers and implications for their conservation. Biol Conserv 244:108519. https://doi.org/10.1016/j.biocon.2020.108519
Fois M, Cuena-Lombraña A, Bacchetta G (2021) Knowledge gaps and challenges for conservation of Mediterranean wetlands: evidence from a comprehensive inventory and literature analysis for Sardinia. Aquat Conserv 31:2621–2631. https://doi.org/10.1002/aqc.3659
Fois M, Cuena-Lombraña A, Araç N, Artufel M, Atak E, Attard V, Bacchetta G et al (2022a) The Mediterranean Island wetlands (MedIsWet) inventory: strengths and shortfalls of the currently available floristic data. Flora Mediterr 32:339–349. https://doi.org/10.7320/FlMedit32.339
Fois M, Farris E, Calvia G, Campus G, Fenu G, Porceddu M, Bacchetta G (2022b) The endemic vascular flora of Sardinia: a dynamic checklist with an overview of biogeography and conservation status. Plants 11:601. https://doi.org/10.3390/plants11050601
Fois M, Cuena-Lombraña A, Zucca C, Nissardi S, Bacchetta G (2022c) Investigating plant–bird co-occurrence patterns in Mediterranean wetlands: can they reveal signals of Ecosystem Connectivity? Diversity. 14:253. https://doi.org/10.3390/d14040253
Gathof AK, Grossmann AJ, Herrmann J, Buchholz S (2022) Who can pass the urban filter? A multi-taxon approach to disentangle pollinator trait–environmental relationships. Oecologia 199:165–179. https://doi.org/10.1007/s00442-022-05174-z
Greuter W (2001) Diversity of Mediterranean island floras. Bocconea 13:55–64
Guarino R, La Rosa M (2019) Flora d’Italia Digitale. In: Pignatti S, Guarino R, La Rosa M (eds) Flora d’Italia, 2nd edn. Edagricole, Milan
Guarino R, Domina G, Pignatti S (2012) Ellenberg’s Indicator values for the Flora of Italy – first update: Pteridophyta, Gymnospermae and Monocotyledoneae. Flora Mediterr 22:197–209. https://doi.org/10.7320/FlMedit22.197
Hobohm C, Bruchmann I (2011) Are there endemic vascular plants in wet habitats of Europe? Transylv Rev Syst Ecol Res 12:1–14
Jarvis A, Reuter HI, Nelson A, Guevara E (2008) Hole-filled SRTM for the globe Version 4, available from the CGIAR-CSI SRTM 90m Database. http://srtm.csi.cgiar.org. Accessed 23 February 2023
Jeanmonod D, Gamisans J (2007) Flora Corsica. Edisud, Aix-en-Provence, France
Julve P (2015) Baseflor. Index botanique, écologique et chorologique de la flore de France (Baseflor. Botaniccological and chorological index of the flora of France). Available at: http://philippe.julve.pagesperso-orange.fr/catminat.htm. Accessed 12 May 2022
Kadereit JW, Arafeh R, Somogyi G, Westberg E (2005) Terrestrial growth and marine dispersal? Comparative phylogeography of five coastal plant species at a European scale. Taxon 54:861–876. https://doi.org/10.2307/25065567
Lanfranco S, Lanfranco E, Westermeier R, Zammit MA, Mifsud MA, Xiberras J (2013) The vascular flora of the Maltese Islands. 2nd Botanical Conference. In: Pons EC, Clarisó IE, Casademont MC, Arguimbau PF (eds), Proceedings and abstracts Islands and plants: preservation and understanding of flora on Mediterranean islands. Consell Insular de Menorca: Colleció Recerca Menorca, Menorca, pp 261–268
Lanfranco S, Grima C, Galea L (2016) A preliminary appraisal of phylogenetic patterns as a tool for long-term monitoring of plant communities in temporary freshwater rockpools. Hydrobiologia 782:201–209. https://doi.org/10.1007/s10750-016-2829-0
Lanfranco S, Bellia AF, Cuschieri P (2020) Influence of morphometric and geographic factors on plant community composition of temporary freshwater rockpools in the Maltese islands. Inland Waters 10:468–479. https://doi.org/10.1080/20442041.2019.1704152
Largier G (2019) La Liste rouge de la flore vasculaire de France métropolitaine. Le J De Botanique 87:21–22. https://www.persee.fr/doc/jobot_1280-8202_2019_num_87_1_1917 Accessed 12 September 2022
Lozano V, Di Febbraro M, Brundu G, Carranza ML, Alessandrini A, Ardenghi NMG et al (2023) Plant invasion risk inside and outside protected areas: Propagule pressure, abiotic and biotic factors definitively matter. Sci Total Environ 162993. https://doi.org/10.1016/j.scitotenv.2023.162993
Lukács BA, Sramkó G, Molnár A (2013) Plant diversity and conservation value of continental temporary pools. Biol Conserv 158:393–400. https://doi.org/10.1016/j.biocon.2012.08.024
Lumbreras A, Marques JT, Belo AF, Cristo M, Fernandes M, Galioto D et al (2016) Assessing the conservation status of Mediterranean temporary ponds using biodiversity: a new tool for practitioners. Hydrobiologia 782:187–199. https://doi.org/10.1007/s10750-016-2697-7
Mahanand S, Behera MD (2019) Understanding the Indian mainland–island biogeography through plant dispersal mechanism. Biodivers Conserv 28:2063–2084. https://doi.org/10.1007/s10531-018-01685-5
Marini L, Battisti A, Bona E, Federici G, Martini F, Pautasso M, Hulme PE (2012) Alien and native plant life-forms respond differently to human and climate pressures. Glob Ecol Biogeogr 21:534–544. https://doi.org/10.1111/j.1466-8238.2011.00702.x
Martínez-López V, García C, Zapata V, Robledano F, De la Rúa P (2020) Intercontinental long‐distance seed dispersal across the Mediterranean Basin explains population genetic structure of a bird‐dispersed shrub. Mol Ecol 29:1408–1420. https://doi.org/10.1111/mec.15413
Martínez-Megías C, Rico A (2022) Biodiversity impacts by multiple anthropogenic stressors in Mediterranean coastal wetlands. Sci Total Environ 818:151712. https://doi.org/10.1016/j.scitotenv.2021.151712
Mayoral O, Mascia F, Podda L, Laguna E, Fraga P, Rita J, Frigau L, Bacchetta G (2018) Alien plant diversity in Mediterranean wetlands: a comparative study within Valencian, Balearic and Sardinian floras. Not Bot Horti Agrobot Cluj-Napoca 46:317–326. https://doi.org/10.15835/nbha46210470
Médail F (2022) Plant biogeography and vegetation patterns of the Mediterranean islands. Bot Rev 88:63–129. https://doi.org/10.1007/s12229-021-09245-3
Minissale P, Sciandrello S (2016) Ecological features affect patterns of plant communities in Mediterranean temporary rock pools. Plant Biosyst 150:171–179. https://doi.org/10.1080/11263504.2014.986248
Minissale P, Molina JA, Sciandrello S (2017) Pilularia Minuta Durieu (Marsileaceae) discovered in south-eastern-Sicily: new insights on its ecology, distribution and conservation status. Bot Lett 164:197–208. https://doi.org/10.1080/23818107.2017.1357051
Mounce R, Rivers M, Sharrock S, Smith P, Brockington S (2018) Comparing and contrasting threat assessments of plant species at the global and sub-global level. Biodivers Conserv 27:907–930. https://doi.org/10.1007/s10531-017-1472-z
Murphy K, Efremov A, Davidson TA, Molina-Navarro E, Fidanza K, Betiol TCC et al (2019) World distribution, diversity and endemism of aquatic macrophytes. Aquat Bot 158:103127. https://doi.org/10.1016/j.aquabot.2019.06.006
Oksanen J, Kindt R, Legendre P, O’Hara B, Simpson GL, Solymons P, Stevens MHH, Wagner H (2007) The vegan package. Community Ecol Package 10:631–637. https://github.com/vegandevs/vegan
Palmas F, Cau A, Podda C, Musu A, Serra M, Pusceddu A, Sabatini A (2022) Rivers of waste: anthropogenic litter in intermittent sardinian rivers, Italy (Central Mediterranean). Environ Poll 302:119073. https://doi.org/10.1016/j.envpol.2022.119073
Panzeca P, Troia A, Madonia P (2021) Aquatic macrophytes occurrence in Mediterranean farm ponds: preliminary investigations in North-Western Sicily (Italy). Plants 10:1292. https://doi.org/10.3390/plants10071292
Pasta S, Perez-Graber A, Fazan L, de Montmollin B (2017) The Top 50 Mediterranean Island Plants UPDATE 2017. IUCN/SSC/Mediterranean Plant Specialist Group, Neuchatel. https://top50.iucn-mpsg.org/. Accessed 13 May 2018
Perennou C, Guelmami A, Paganini M, Philipson P, Poulin B, Strauch A et al (2018a) Mapping Mediterranean wetlands with remote sensing: a good-looking map is not always a good map. Adv Ecol Res 58:243–277. https://doi.org/10.1016/bs.aecr.2017.12.002
Perennou C, Gaget E, Galewski T, Geijzendorffer I, Guelmami A (2018b) Evolution of wetlands in Mediterranean region. In: Zribi M, Brocca L, Tramblay Y, Molle F (eds) Water resources in the Mediterranean Region. Elsevier Inc, Radarweg, pp 297–320
Pignatti S, Guarino R, La Rosa M (2017–2019) Flora d’Italia, 2nd ed.; Edagricole, Bologna
Raunkiaer C (1934) The Life Forms of Plants and statistical plant geography. Clarendon Press, Oxford
Rodríguez-Merino A, Fernández‐Zamudio R, García‐Murillo P (2019) Identifying areas of aquatic plant richness in a Mediterranean hotspot to improve the conservation of freshwater ecosystems. Aquat Conserv: Mar Freshw 29:589–602. https://doi.org/10.1002/aqc.3088
Rossi G, Montagnani C, Gargano D, Peruzzi L, Abeli T, Ravera S, Cogoni A, Fenu G, Magrini S, Gennai M et al (2013) Lista Rossa della Flora Italiana. 1. Policy Species e altre specie minacciate. Comitato Italiano IUCN e Ministero dell’Ambiente e della Tutela del Territorio e del Mare, Roma. http://www.iucn.it/pdf/Comitato_IUCN_Lista_Rossa_della_flora_italiana_policy_species.pdf. Accessed 25 June 2022
Rossi G, Orsenigo S, Gargano D, Montagnani C, Fenu G, Peruzzi L, Abeli T, Alessandrini A, Bacchetta G, Bartolucci F et al (2020) Lista Rossa Della Flora Italiana. 2. Specie endemiche e altre specie minacciate. Comitato Italiano IUCN e Ministero dell’Ambiente e della Tutela del Territorio e del Mare, Roma
Ruzzier E, Forbicioni L, Gentili R, Tormen N, Dondina O, Orioli V, Bani L (2021) From island biogeography to conservation: a multi-taxon and multi-taxonomic rank approach in the tuscan archipelago. Land 10:486. https://doi.org/10.3390/land10050486
Schembri PJ, Sultana J (1989) Red data book for the Maltese islands. Department of Information, Malta
Schöpke B, Heinze J, Pätzig M, Heinken T (2019) Do dispersal traits of wetland plant species explain tolerance against isolation effects in naturally fragmented habitats? Plant Ecol 220:801–815. https://doi.org/10.1007/s11258-019-00955-8
Schrader J, Wright IJ, Kreft H, Westoby M (2021) A roadmap to plant functional island biogeography. Biol Rev 96:2851–2870. https://doi.org/10.1111/brv.12782
Sciandrello S, Guarino R, Minissale P, Spampinato G (2015) The endemic vascular flora of Peloritani Mountains (NE Sicily): plant functional traits and phytogeographical relationships in the most isolated and fragmentary micro-plate of the Alpine orogeny. Plant Biosyst 149:838–854. https://doi.org/10.1080/11263504.2014.908978
Sciandrello S, Privitera M, Puglisi M, Minissale P (2016) Diversity and spatial patterns of plant communities in volcanic temporary ponds of Sicily (Italy). Biologia 71:793–803. https://doi.org/10.1515/biolog-2016-0096
Sciandrello S, Cambria S, del Galdo GG, Minissale P, Puglisi M, Tavilla G, Tomaselli V (2023) Ecological features and conservation of Urtica Rupestris Guss. (Urticaceae) a narrow endemic species of Sicily. Plants 12:164. https://doi.org/10.3390/plants12010164
Tavilla G, Minissale P, Camilleri L, Lanfranco S (2023) Addition of two species to the Maltese flora: Lythrum tribracteatum Salzm. Ex Spreng. (Lythraceae) and Poa Maroccana Nannf. (Poaceae). Check List 19:743–751. https://doi.org/10.15560/19.5.743
Tavilla G, Lamoliere A, Gabarretta J, Attard V, Henwood J, Stevens DT et al (2023a) Climate Change and Wetland ecosystems: the effects on Halophilous Vegetation Diversity in Il-Ballut ta’Marsaxlokk Natura 2000 site. (Malta) Land 12:1679. https://doi.org/10.3390/land12091679
Taylor NG, Grillas P, Al Hreisha H, Balkız Ö, Borie M, Boutron O et al (2021) The future for Mediterranean wetlands: 50 key issues and 50 important conservation research questions. Reg Environ Change 21:1–17. https://doi.org/10.1007/s10113-020-01743-1
Tichý L, Axmanová I, Dengler J, Guarino R, Jansen F, Midolo G et al (2023) Ellenberg-type indicator values for European vascular plant species. J Veg Sci e13168. https://doi.org/10.1111/jvs.13168
Tomàs-Vives P (2008) Inventory, assessment and monitoring of Mediterranean wetlands: The pan-Mediterranean wetland inventory module. TdV, MedWet, Arles. https://medwet.org/codde/wetlandinvetory.html. Accessed 25 April 2016
Troia A, Ilardi V, Oddo E (2020) Monitoring of alien aquatic plants in the inland waters of Sicily (Italy). Webbia 75:77–83. https://doi.org/10.36253/jopt-8414
Weber HC, Kendzior B (2006) Flora of the Maltese Islands. Afield Guide. Margraf Publishers, Weikersheim
Wei T, Simko V, Levy M, Xie Y, Jin Y, Zemla J (2017) Package ‘corrplot’. Statistician 56, e24
Acknowledgements
The Authors thank MAVA Foundation for having supported the database development and promoting the MedIsWet network.
Funding
This study was carried out within the “e.INS – Ecosystem of Innovation for Next Generation Sardinia” funded by the Italian Ministry of University and Research under the Next-Generation EU Programme (National Recovery and Resilience Plan – PNRR, M4C2, INVESTMENT 1.5 – DD 1056 of 23/06/2022, ECS00000038). This manuscript reflects only the authors’ views and opinions, neither the European Union nor the European Commission can be considered responsible for them.
Open access funding provided by Università degli Studi di Cagliari within the CRUI-CARE Agreement.
Author information
Authors and Affiliations
Contributions
M.F. prepared and analysed the data, wrote the manuscript with contributions from all co-authors and designed the figures and tables. M.F., A.C.L. and G.B. conceived the original idea. M.F., A.C.L., M.A., V.A., S.C., K.F., M.M., P.M., A.P., S.S., E.T. and G.B. provided the data, corrected the used species’ database. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Communicated by Daniel Sanchez Mata.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Fois, M., Cuena-Lombraña, A., Artufel, M. et al. Plant distribution and conservation in mediterranean islands’ lentic wetlands: there is more than aquatic species. Biodivers Conserv 33, 1373–1392 (2024). https://doi.org/10.1007/s10531-024-02803-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-024-02803-2