
Abstract
Climatic niche shifts occur when species occupy different climates in the introduced range than in their native range. Climatic niche shifts are known to occur across a range of taxa, however we do not currently understand whether climatic niche shifts can consistently be predicted across multiple introduced ranges. Using three congeneric weed species, we investigate whether climatic niche shifts in one introduced range are consistent in other ranges where the species has been introduced. We compared the climatic conditions occupied by Rumex conglomeratus, R. crispus, and R. obtusifolius between their native range (Eurasia) and three different introduced ranges (North America, Australia, New Zealand). We considered metrics of niche overlap, expansion, unfilling, pioneering, and similarity to determine whether climatic niche shifts were consistent across ranges and congeners. We found that the presence and direction of climatic niche shifts was inconsistent between introduced ranges for each species. Within an introduced range, however, niche shifts were qualitatively similar among species. North America and New Zealand experienced diverging niche expansion into drier and wetter climates respectively, whilst the niche was conserved in Australia. This work highlights how unique characteristics of an introduced range and local introduction history can drive different niche shifts, and that comparisons between only the native and one introduced range may misrepresent a species’ capacity for niche shifts. However, predictions of climatic niche shifts could be improved by comparing related species in the introduced range rather than relying on the occupied environments of the native range.
Similar content being viewed by others


Introduction
Attempts to predict the establishment of plants introduced to new regions have commonly assumed species inhabit similar environments across the globe, usually termed climatic niche conservatism (Bradley et al. 2012; Bradley et al. 2010a, b; Bradley et al. 2010a, b; Liu et al. 2020). A species’ realised climatic niche is the set of climates a species currently occupies and in which it experiences stable or positive population growth (Atwater et al. 2018; Holt 2009; Hutchinson 1957), and niche conservatism occurs when a species inhabits that same niche in both the native and introduced range (Liu et al. 2020; Petitpierre et al. 2012). However, studies have revealed shifts in the climatic niche between introduced and native ranges across many taxa and environments (Atwater & Barney 2021; Atwater et al. 2018; Bujan et al. 2020; Christina et al. 2019; Comte et al. 2017; Liu et al. 2016; Silva et al. 2016; Tingley et al. 2014). Niche shifts can occur when species occupy climates in the introduced range that are either available but unoccupied in the native range (expansion), unavailable in the native range (pioneering), or by failing to occupy climates in the introduced range that are occupied in the native range (unfilling; Atwater et al. 2018; Guisan et al. 2014). A better understanding of niche shifts is essential to predict the establishment and future range dynamics of introduced species under ongoing spread and global climate change.
While it is well-documented that climatic niche shifts can occur between the native range and an introduced range, the question remains whether such shifts are predictable and consistent in both direction and magnitude across multiple introduced ranges for a given species. Hundreds of species across different taxa and continents have been compared to date as part of the ongoing debate around the frequency of niche shifts (Atwater et al. 2018; Atwater & Barney 2021; Liu et al. 2020; Petitpierre et al. 2012; Webber et al. 2012), however these studies provide little insight into whether information on niche shifts is transferable between regions or species. To date, most studies have focussed on pairwise comparisons of plant distributions between the native and one introduced range (Early & Sax 2014; Fernández & Hamilton 2015; Gallagher et al. 2010; Randin et al. 2006) and comparisons across studies are challenging, since studies have used different underlying variables to define the environmental space (Liu et al. 2020). Whilst these comparisons provide useful information on how often niche shifts occur, approaches that consider several introduced ranges are required to determine if niche shifts are consistent within a species across introduced ranges. Currently the transferability of plant species distributions between different introduced ranges has been little explored (Datta et al. 2019; Petitpierre et al. 2012), however it has been assessed for some animal species (Hill et al. 2017; Pili et al. 2020; Silva et al. 2016). Understanding whether niche shifts are consistent between ranges will provide insight into how species adapt to these new climates, what environmental factors limit species distributions and predictions of future invasion risk.
When comparing niche shifts across several introduced ranges there are a number of potential outcomes (Fig. 1). Firstly, a species may consistently shift into certain climates across all observed ranges (Fig. 1A). Consistent niche shifts across ranges may be expected in species where the native range does not fully encapsulate their fundamental environmental tolerances. This would suggest that the species is either pre-adapted to, or able to rapidly adapt to, a specific subset of climates. Silva et al. (2016) showed that independent invasions of a dung beetle, Onthophagus taurus, into different introduced regions all experienced niche shifts towards more humid climates than those occupied in the native range. Alternatively, we may see idiosyncratic patterns in the direction of niche shifts, with the species shifting in different directions across ranges (Fig. 1B), or experiencing niche shifts in some ranges but niche conservatism in others (Fig. 1C). This could indicate that non-climatic factors in the introduced range are facilitating a niche shift, or that a niche shift represents an escape from non-climatic factors which restrict the species distribution in some ranges (Bulleri et al. 2016). For example gorse, Ulex europaeus, occupies warmer areas in Australia and South America than in its native range, yet cooler areas in North America, but the mechanisms behind these niche shifts are unknown (Christina et al. 2019). Inconsistent niche shifts across ranges could also be explained due to differences in genetic diversity, where founder effects result in phenotypic differences between populations (Flanagan et al. 2021; Liao et al. 2020; Ribas et al. 2018). Additionally, niche shifts may be inconsistent in magnitude, where niche shifts in two independent ranges could both be towards similar climates but more extreme in one introduced region than the other. Variations in magnitude of a niche shift would amplify the difficulty in predicting the climatic thresholds a species may be able to tolerate, and subsequently which areas are susceptible to establishment.

Possible niche shift scenarios between regions. Axes represent two different example environmental gradients across which a niche shift could be observed. Areas occupied in a species’ native range (N) are shown by the green shaded circle; areas occupied in two different introduced ranges (I1, and I2) are shown by the purple and orange circles respectively. A Niche shifts are consistent across independent ranges. B Niche shifts are inconsistent across independent ranges. C Niche shifts occur in some ranges, whilst the niche is conserved in others. D Two niche dynamics that can lead to niche shifts are shown. Niche expansion (blue) refers to areas of analogue, or overlapping, climate space between the native and introduced ranges that are only occupied in the introduced range. Niche unfilling (red) refers to areas of analogue climate space that are only occupied in the native range. Dashed circles denote available climate space for the native range (green) and introduced range (purple)
As well as understanding how niche shifts occur it is also important to know whether they occur under analogue or non-analogue climates. Analogue climates are climates available to a species in both the native and introduced ranges, whereas non-analogue climates are present in only one range (Fig. 1D). Liu et al. (2020), as well as other earlier work (Petitpierre et al. 2012; Tingley et al. 2016), chose not to investigate niche dynamics under non-analogue climates because shifts due to rapid evolution cannot be distinguished from shifts due to an underestimation of the fundamental niche. Whilst it is important to distinguish between these two types of niche shifts, shifts into non-analogue climates still represent realised niche shifts that can affect the accuracy of environmental niche models (Atwater et al. 2018; D’Amen & Azzurro 2020; Pili et al. 2020; Webber et al. 2012) and the impact of invasions. When considering niche shifts across several introduced ranges, shifts into non-analogue climates may become important as each range will have novel combinations of climates. Determining whether species experience shifts into non-analogue climates that are similar across their ranges is an important step in determining the likelihood of shifts into non-analogue climates.
It is currently not well understood whether closely related species show consistent niche shifts. Previous studies of niche shifts in multiple ranges have mostly looked at a single species (Christina et al. 2019; Datta et al. 2019; Silva et al. 2016; Tingley et al. 2014), but comparisons of closely related species would provide new insights into whether they experience consistent niche shifts across ranges. If closely related species share a similar climatic niche and shift niches consistently, we may conclude that common factors, such as climate availability, in the introduced range may be a larger driver of niche shifts than individual species attributes. If this is the case then data from related species could be used to supplement our knowledge of the species niche, and aid in predicting shifts (Smith et al. 2019). We could then use the presence of a known species to make strong inferences about where a related species could spread.
Here we assess the consistency of climatic niche shifts across multiple introduced ranges for three closely related, globally invasive, herbaceous species. Combining climatic information across both ranges and species allows us to make previously unseen direct comparisons of niche occupancy in shared environmental space to ask:
-
1.
Do species shift their niches in the same direction across different introduced ranges?
-
2.
Are climatic niche shifts consistent across closely related species?
Where species undergo climatic niche shifts, we expect to find shifts into similar climates in all introduced ranges (Fig. 1A) unless the species’ distribution in the introduced range is constrained by non-climatic factors. Similarly, we expect that such shifts would be consistent across closely related species, allowing information from the distribution of one species to help inform the likelihood of other species undergoing climatic niche shifts. With this analysis we hope to shed light on when and where niche shifts occur, and whether the direction of niche shifts can be forecast.
Methods
Study area and species
We studied three common dock species: Rumex obtusifolius L., R. crispus L., and R. conglomeratus Murray. These species are all ruderals, typically colonising open, disturbed environments associated with human activity, including pasture (Cavers & Harper 1964, 1966; Grime et al. 2007; Holm et al. 1997; Lousley & Kent 1981). Rumex spp. are short lived and tend to reproduce annually if conditions are favourable but are capable of biennial or perennial life histories (Cavers & Harper 1964; Grime et al. 2007). These congeners all prefer open environments with minimal shading, nitrogen rich soils, and can tolerate either weakly acidic or basic soils. They are capable of hybridising with one another (Cavers & Harper 1964; Holm et al. 1979; Lousley & Kent 1981) and share broad environmental tolerances in both their native and introduced ranges (Grime et al. 2007; Hill et al. 1999; Lousley & Kent 1981). Despite this, small differences in environmental tolerances are observed between species, with R. obtusifolius in particular able to tolerate extremely nutrient rich or polluted soils and waterways (Grime et al. 2007). Rumex conglomeratus has a smaller global range size and is restricted to more mild temperatures and wetter habitats, such as along waterways (Hill et al. 1999; Lousley & Kent 1981). In comparison, R. obtusifolius and R. crispus can tolerate cooler, drier climates, with their native distributions extending to the Arctic Circle (Hultén, 1950).
All three species are of Eurasian origin yet over the last five hundred years have been introduced and established across the globe as agricultural seed contaminants (Table 1; Figure S.1; Holm et al. 1979; Holm et al. 1997; Vibrans 1998). Furthermore, occurrence data for these species are numerous in both the native and several introduced ranges, making them ideal candidates for assessing global niche shifts. Despite the wide variety of ecoregions Rumex spp. inhabit (Olson et al. 2001) they generally occupy similar habitats across the globe, indicating that any possible niche shifts would likely be driven by climate rather than habitat limitations (Fig. 2; Cavers & Harper 1964; Grime et al. 2007; Holm et al. 1997; Lousley & Kent 1981). Their long invasion history suggests they are likely to have reached climatic equilibrium in their introduced ranges, and that sufficient generations have passed for adaptations to new climates to develop (Table 1; Vibrans 1998). As such, Rumex spp. have had ample opportunities for niche shifts to occur. Rumex spp. were not deliberately introduced for agricultural or horticultural purposes and have not been subjected to artificial selection which may affect their invasiveness or climatic tolerances (Kitajima et al. 2006). Therefore, any niche shifts observed are likely due to natural processes.

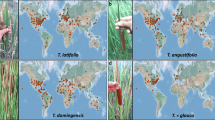
Distribution of Rumex conglomeratus (top), Rumex crispus (middle) and Rumex obtusifolius (bottom) records included in our study, plotted as black points at 50% opacity. Records span temperate and tropical zones. The areas used to determine background climates are displayed by shaded minimum convex polygons and are the same for each species to allow direct comparison: Native range–Teal; Western North America–Magenta; Eastern Australia–Orange; New Zealand–Green. New Zealand is also displayed in the inset of each distribution map for better visualization
We modelled these species’ niches across the native range, predominantly in Europe, and three regions where the species are recorded as naturalised in the Global Invasive Species Database (Invasive Species Specialist Group 2019) as well as regional floras: western North America (USDA & NRCS 2019), south-eastern Australia (Atlas of Living Australia 2019), and New Zealand (New Zealand Plant Conservation Network 2019a). These regions were selected because they have an abundance of occurrence records for all three species, a wide variety of climates, and a long history since the species were first introduced (Table 1). Additional information on the species distribution ranges is provided in the Supplementary Material (Figure S1).
Defining the native range
We define the native range for each of these species as spanning Europe, the Middle East, and Northern Africa (Fig. 2). Multiple databases, including the Global Biodiversity Information Facility (GBIF; GBIF.org) the Atlas Florae Europoaea (AFE 1979), the Flora of Japan (FOJ; Flora of Japan 2019), Calflora (Calflora 2019), Centre for Agriculture and Bioscience International (CABI 2019), U.S. Germplasm Resources Information Network (USDA 2019), and the Global Weed Compendium (Randall 2017), were consulted to determine where these species were classified as native (Table S1). Areas of continuous species occurrence contiguous with these regions were likewise considered native, unless otherwise stated as a known introduction, as these occurrences were considered likely to be naturally distributed from the same range. A literature search (Table S2) was conducted to determine whether historical records indicated known introductions. Table S1 displays a full list of the countries considered in this study and whether we classified these Rumex species as native or introduced in that country.
There is no consensus on whether Japan is part of the native or introduced range for R. obtusifolius and R. crispus. Some sources include eastern Asia and Japan as part of the native range for R. obtusifolius and R. crispus (Flora of Japan 2019; USDA 2019), while other sources consider the species as introduced (Grossrieder & Keary 2004; Makuchi & Sakai 1984; Miyagi et al. 2010; Nishida 2002). Additionally, the available data for these species in Asia show a large geographic distance between the closest European and Asian populations suggesting that they are likely to be genetically isolated, or that there are large gaps in sampling. As a result, our analyses did not include Japan as part of the native range for R. obtusifolius and R. crispus. However, we repeated the analyses with Japan included as part of the native range of R. obtusifolius and R. crispus to ensure the robustness of our results, which did not lead to significant changes in our results or conclusions (Table S5). Rumex conglomeratus is not considered native to eastern Asia by any of the sources we assessed.
Data collection
Species’ occurrence data
Occurrence records were collected for the three Rumex spp. from: GBIF (GBIF.org 2020), AFE, the Atlas of Living Australia (ALA), the Early Detection and Distribution Mapping System (EDDMapS; University of Georgia 2019), Calflora, records georeferenced from targeted journals (Table S2; Table S3), and our own surveys in the UK and NZ. Due to the underreporting of Rumex spp. distribution records in New Zealand, we examined New Zealand journals that commonly publish floristic inventories, using the search term “Rumex” and checked all results for occurrence records. Records were georeferenced at the highest possible resolution using Google Maps (google.com/maps) and details are provided in Table S4. Occurrence records were cleaned by removing records with missing or inaccurate coordinates and records with coordinate uncertainties of over 10 km. Records were separated into the native range and three introduced ranges and thinned by applying a 2.5 arc minute (~ 5 km at the equator) grid over the occurrence points, and selecting one random point per grid cell using the R package GSIF (Hengl et al. 2014). Thinning the occurrence records reduces geographic sampling bias and removes duplicate data.
Climate data
To improve predictability and interpretability of our models we used six bioclimatic variables commonly used to best representant constraints on plant ecology (Alexander 2013; Dullinger et al. 2017; Fourcade et al. 2018; Keller et al. 2009; Root et al. 2003): temperature seasonality (BIO4), maximum temperature of the warmest month (BIO5), minimum temperature of the coldest month (BIO6), precipitation seasonality (BIO15), precipitation of the wettest quarter (BIO16), and precipitation of the driest quarter (BIO17). Both temperature (Benvenuti et al. 2001; Cavers & Harper 1964, 1966) and precipitation, through soil moisture (Cavers & Harper 1964), are important in determining plant species distributions at local scales and using fewer, more biologically relevant variables makes results more interpretable. These six variables were obtained from the WorldClim dataset (Booth et al. 2014; Fick & Hijmans 2017; worldclim.org) at 2.5 arc minute (~ 5 km at the equator) resolution.
Niche analysis
In order to assess climatic niche shifts in the introduced ranges we utilised the environmental principal component analysis (PCA-env) approach proposed by Broennimann et al. (2012), and later improved by Petitpierre et al. (2012) and Guisan et al. (2014) which standardises the environment across ranges. This method corrects for sampling biases by applying a kernel density smoother to estimate the density of occurrences and is considered the “gold standard” for assessing niche conservatism (Liu et al. 2020). We adapted the standard approach by extracting climatic data from minimum convex polygons (MCPs) fitted around the combined occurrence records for all three species for each region, as opposed to extracting data for each species individually. This allowed us to compare niches and niche shifts for all three species within the same climatic boundaries. Following methods developed by Silva et al. (2016), a buffer zone of 1 decimal degree (~ 111 km at the equator) was added around species’ presences and MCPs were fitted around this area for each region (Figure S1). In order to reduce the number of variables to two, which the PCA-env approach requires, a principal component analysis (PCA) was performed on the climates available to Rumex spp. and we used values of the PCA axes at the species’ known occurrence points to calculate the occupied climate conditions for each species in each range. Following guidelines from Guisan et al. (2014) and amended by Silva et al. (2016), pairwise comparisons were performed between all ranges.
In order to determine whether climatic niche shifts occurred, we compared the available environmental conditions of each of the three introduced ranges and the native range, following metrics suggested by Guisan et al. (2014). We followed a framework based on centroid shift, overlap, unfilling, and expansion (COUE; Liu et al. 2020) however, given the known issues related to centroid shift methods (Godsoe 2010; Guisan et al. 2014) we characterised shifts based only on overlap (D), unfilling (U), and expansion (E). We also assessed niche shifts into non-analogue space by measuring niche pioneering (P), the proportion of occupied environmental space in the introduced range which is only present in the introduced range (Atwater et al. 2018; Guisan et al. 2014; Petitpierre et al. 2012; Webber et al. 2012). Niche overlap in each comparison was calculated using Schoener’s D (Broennimann et al. 2012; Schoener 1970; Warren et al. 2008), a metric which varies from 0, indicating the greatest possible distance between predicted occurrences in each range (no niche overlap), to 1, indicating no differences (complete niche overlap). Niche similarity was calculated to determine whether niches in the native and introduced ranges are more or less similar than expected by chance given the available climates (Aguirre-Gutiérrez et al. 2015; Warren et al. 2008, 2010). Significant niche similarity indicates the niches are more similar than expected by chance, whereas non-significant niche similarity indicates a niche shift. The PCA output was also used to determine the direction, and hence consistency, of the shifts in climate space across regions for each species. The same methods were then applied to measure the degree of niche overlap and similarity between species within each introduced range. All metrics were calculated using the ecospat package (Di Cola et al. 2017) in the statistical software R version 3.5.1 (R Core Team 2013).
Results
Principal component analysis
The first two principal components explained 77.4% of the variation in the original six climate variables (44.7% and 33.0% for PC1 and PC2 respectively) and therefore adequately represented the available environmental space. Increasing values of PC1 corresponded to colder areas with more seasonal variation in their temperatures, and less seasonal variation in precipitation. Increasing values of PC2 indicated areas with higher precipitation and less seasonality in their temperatures (Fig. 3).

A Contribution of climate variables to the first two axes of the principal component analysis. B Direction of variables with respect to the first two principal components. Direction of arrows indicates increasing values of that variable. The first two principal components represent the environmental space used for further niche analysis. Some variable names are shortened for simplicity: Minimum Precipitation = Precipitation of the driest quarter (BIO17), Maximum Precipitation = Precipitation of the wettest quarter (BIO16), Minimum Temperature = Minimum temperature of the coldest month (BIO6), Maximum Temperature = Maximum temperature of the warmest month (BIO5)
Niche shifts are inconsistent across regions
Climatic niche shifts were idiosyncratic between introduced regions for all species, with one region characterised by niche conservatism (Fig. 4; Australia) and two regions characterised by large niche shifts (Fig. 4; New Zealand and North America).

Comparisons of niche overlap in environmental space. Each panel shows a comparison between the native range (Teal) and one of the introduced ranges (New Zealand–Green; North America–Pink; Australia–Orange). The climate space occupied by the species is shown in solid lines and the total available climate space of the respective range is shown with dashed lines. Comparing down columns shows differences between regions whereas comparing across rows shows differences between species in the same region. Increasing values of PC1 broadly correspond to cooler, more variable temperatures, and less seasonal precipitation. Increasing values of PC2 broadly correspond to increased precipitation, and more stable temperatures. Variable correlations with PC1 and PC2 can be seen in full in Fig. 3
Australia is the only region in which Rumex spp. largely conserved their climatic niche (Fig. 4), and it was the region with the highest level of niche overlap with the native range (0.20 ≤ D ≤ 0.34; Table 2). Furthermore, Australia is the only region that had niche similarity values that were significantly similar to the native range for all three species (p = 0.01; Table 2). Rumex spp. in Australia experienced low levels of niche expansion, in both analogue and non-analogue climates (0.01 ≤ E ≤ 0.07; 0.01 ≤ P ≤ 0.06), and high levels of niche unfilling (0.39 ≤ U ≤ 0.60; Table 2). Niche unfilling in Australia occurred in wet areas with highly seasonal temperatures and drier areas with less seasonal temperatures (both higher and lower values of PC2; Fig. 4).
In New Zealand all three species expanded into warmer, wetter areas with more variable precipitation (lower values of PC1 and higher values of PC2; Fig. 4). The climatic niche shifts in NZ were primarily into non-analogue climate space (0.30 ≤ P ≤ 0.37), however niche expansion into analogue climate was also prevalent (0.24 ≤ E ≤ 0.33; Table 2; Fig. 4). New Zealand was the only introduced range where the similarity of the occupied niche in analogue climates to the native range was only of borderline statistical significance in all three species (0.05 < P < 0.10; Table 2). Niche overlap and unfilling between the occupied climates in NZ and the native range were low (D ≤ 0.16; U ≤ 0.07; Table 2).
In North America the climatic niche expanded predominantly into warmer, drier climates (lower values of PC1 and PC2), but also into cooler climates with greater seasonality in temperature (higher values of PC1; Fig. 4). Climatic niche shifts in North America were observed across both analogue and non-analogue climate space (0.40 ≤ E ≤ 0.69; 0.32 ≤ P ≤ 0.57; Table 2; Fig. 4). The niche in North America was significantly similar to the niche in the native range for both R. crispus and R. conglomeratus but only of borderline significance for R. obtusifolius. However, low levels of niche overlap (D = 0.15 & D = 0.17 respectively) suggest the species have largely expanded into non-analogue climates (Table 2). Two species, R. obtusifolius and R. conglomeratus, showed high levels of niche unfilling in North America (U = 0.52 & U = 0.91 respectively) where available wetter, cooler areas remained unoccupied in the introduced range (high values of both PC1 and PC2; Fig. 4).
Congeners display similar niche shift directions but low overlap
Both the direction and presence of climatic niche shifts were consistent across Rumex species (Fig. 4). In all but one instance niche overlap between species within a range was higher than overlap between the native range and the introduced ranges (Table 2; Table 3). The only exception was the overlap between R. crispus and R. conglomeratus in North America, where overlap between niche space in North America and the native range was higher for both species than the overlap between species in North America. Nevertheless, niche overlap between species in each introduced range was lower than between species in the native range (Table 3).
Comparisons of niche overlap among the three Rumex spp. show little consistency across introduced regions (Table 3). In New Zealand, Rumex spp. were significantly similar to one another (P < 0.05) and displayed high levels of niche overlap (0.59 ≤ D ≤ 0.76). In North America, R. obtusifolius occupied significantly similar niche space to both R. crispus and R. conglomeratus (P < 0.05) with moderate values of niche overlap (0.391 ≤ D ≤ 0.448), whilst R. crispus and R. conglomeratus occupied dissimilar niche space (P = 0.42). In contrast, occupied niche space in Australia was only significantly similar between R. conglomeratus and R. crispus (P < 0.04), despite moderate values of niche overlap for all comparisons (0.442 ≤ D ≤ 0.630).
Discussion
By comparing climatic niches between multiple introduced ranges and species, we found that independent introductions of Rumex spp. have resulted in different niche shifts across geographically distinct ranges into both analogue and non-analogue climates. We have shown that even superficially similar invasions by closely-related species can lead to examples of every niche change proposed by Guisan et al. (2014). We found evidence that the direction of climatic niche shifts was similar between close relatives, although the resulting occupied climates did not always overlap. Our work suggests that the dynamics driving niche shifts differ between ranges and that information from closely related species may offer greater insight into likely niche occupancy in the introduced range than assumptions of niche conservatism.
Our results highlight how niche shifts can vary dramatically from one range to another. This lack of consistency in niche shifts between ranges suggests that one cannot easily extrapolate the likelihood of a climatic niche shift from one range to another range and that whether species conserve their niche depends on both the species and the range considered. Work on Ulex europaeus, similarly found inconsistency in niche shifts across introduced ranges (Christina et al. 2019). This limits the utility of approaches that classify species in absolute terms as either undergoing niche shifts or conserving their niche (Atwater et al. 2018; Bates & Bertelsmeier 2021; Early & Sax 2014; Liu et al. 2020; Petitpierre et al. 2012) or which consider all geographically distinct introduced ranges together, as this may obscure differences between ranges and lead to ambiguous results (e.g. Guo et al. 2013). In order to determine how frequently niche shifts differ across regions and taxa future studies must standardise the environmental space between multiple introduced ranges as suggested by Liu et al. (2020). However, as more ranges or species are included in the standardised environmental space the environmental space is likely to become less representative for any one range or species, particularly if there are large discrepancies in the size of their respective native ranges. As such, these methods would be unsuitable for simultaneously comparing large numbers of taxa drawn from different native ranges as other studies have done (Atwater & Barney 2021; Atwater et al. 2018; Early & Sax 2014; Liu et al. 2020; Petitpierre et al. 2012).
In both North America and New Zealand, niche pioneering into non-analogue climates was a large contributor to overall niche shifts (Fig. 4), consistent with other studies (Atwater et al. 2018; D’Amen & Azzurro 2020; Pili et al. 2020; Webber et al. 2012). Ignoring non-analogue space, as some authors have done (e.g. Kolanowska & Konowalik 2014; Liu et al. 2020; Petitpierre et al. 2012), is likely to produce environmental niche models that under-predict species distributions. Given the aim of quantifying niche conservatism is to estimate the validity of transferring environmental niche models (Guisan et al. 2012; Liu et al. 2020; Petitpierre et al. 2012), information on niche shifts in non-analogue climate space should be included to prevent the under-prediction of species distributions (Atwater et al. 2018; Webber et al. 2012). Furthermore, while Liu et al. (2020) concluded that the extinction risk of introduced terrestrial plants may be higher than other taxa as they experience lower niche expansion, we demonstrate that accounting for non-analogue climates indicates Rumex spp. can tolerate warmer, cooler, wetter, or drier climates than those found in the native range (Fig. 4). This not only reduces the risk of extinction, but also increases the risk that invasive species will persist and spread.
Given that our results show Rumex spp. experienced dramatic climatic niche shifts in some ranges whilst conserving their niche in others, it begs the question what is driving these niche shifts. If Rumex spp. are pre-adapted to all of the climates they occupy in each range we would expect those climates to be occupied in every range. Alternatively, it is possible that Rumex spp. have rapidly adapted to tolerate the climates available in both North America and New Zealand (Bates & Bertelsmeier 2021; Boheemen et al. 2019; Clements & Jones 2021; Hulme & Barrett 2013; Luo et al. 2019), though Rumex spp. conserved their niche in Australia. Given that some unoccupied climates in Australia are occupied in North America, it is possible that a genetic bottleneck in Australia limited opportunities for pre-adaptation. Alternatively, factors other than climate may limit or facilitate Rumex spp. distributions in their introduced ranges. Whilst climate typically determines species’ distributions at broad scales (Bello et al. 2013), recent work has highlighted the importance of non-climatic factors, such as human disturbance, in introduced plant species distributions (Essl et al. 2020; Kołodziejek & Patykowski 2015; Mairal et al. 2021; Pysek et al. 2010; Redpath & Rapson 2015; Seebens et al. 2018). Biotic interactions can both restrict (Alexander et al. 2018; Bello et al. 2013; Godsoe et al. 2018) and facilitate (Stephan et al. 2021) changes in niche shifts, but this is less likely to influence Rumex species as they do not rely upon pollinators, seed dispersers, or mycorrhiza (Cavers & Harper 1964; Grime et al. 2007). Although they have escaped specialised herbivores in New Zealand this has only led to a shift in performance in R. crispus (Costan et al. 2022). Changes in Rumex species’ occupied niche space are more likely due to the prevalence of anthropogenic land use (e.g. disturbed roadsides, irrigated land or pasture margins) which can facilitate niche shifts into climates that otherwise would be unsuitable (Dainese et al. 2017; Koide et al. 2017). Understanding whether non-climatic factors or species traits (MacLean & Beissinger 2017) affect the consistency of climatic niche shifts may help predict whether consistent niche shifts would be observed for different ranges or species.
Compared to a recent synthesis by Liu et al. (2020), our values of niche expansion for Rumex spp. are larger than average for invasive plants whereas our values for niche unfilling vary more between ranges. Rumex species in New Zealand have occupied almost all the available analogue climate space and have experienced a large niche expansion towards warmer, wetter climates. In contrast, Rumex spp. in North America have expanded into a wide variety of climates, but their large values of unfilling suggest that they have not reached climatic equilibrium or have been under sampled. Rumex spp. are often not recorded officially at the species level, in part due to the difficulty of identifying them when not in flower (Baskin & Baskin 1978; Cavers & Harper 1964; Grime et al. 2007; Holm et al. 1977, 1997), creating taxonomic bias (Troudet et al. 2017). Additional occurrence records from introduced ranges would likely increase the magnitude of shifts observed or reduce the incidence of niche unfilling, depending on where the records occurred in climate space, and therefore would not substantially affect our conclusions. Furthermore, niche shifts in climate space observed through ordination based analyses, such as ours, do not always correlate with large geographic range shifts (Atwater & Barney 2021; Guo et al. 2013; Liu et al. 2020).
Our results indicate that qualitative similarities between congeneric species niche shifts could be used to predict whether a species is likely to experience climatic niche shifts when introduced to new ranges. Niche overlap between species within a range was almost always greater than niche overlap between a species’ introduced range and its native range. This suggests that environmental niche models (ENMs) predicting species niche shifts would be more accurate if based on congener data rather than assuming niche conservatism. Nevertheless, these ENMs would miss important niche divergence between species after invasions, as we found there was greater overlap between species in the native range than in any introduced range (Table 3). This suggests that while these species occupy similar climates in the native range, either pre-adaptation or rapid adaptations to new environments can cause even similar species to occupy different environments post invasion. Whilst this could be due to differences in intraspecific genetic diversity, it could also reflect differing non-climatic environmental preferences such as in soil nutrient richness, pH, and moisture (Cavers & Harper 1964; Hill et al. 1999; Lousley & Kent 1981). These results suggest that pooling data between related species (Smith et al. 2019; but see: Mota-Vargas & Rojas-Soto 2016) or across introduced ranges is unlikely to completely predict species resulting distributions after a climatic niche shift occurs, but may be more accurate than only including species data from the native range. However, the degree to which these results are generalisable to other species requires further investigation. Despite a large species pool, Atwater et al. (2018) were unable to disentangle the effects of species traits (growth form, life span, and cultivation status) on the occurrence of niche shifts. This suggests that niche shifts could be highly context dependent (Catford et al. 2022). They also found that herbaceous species, e.g. Rumex, generally conserve their niche more often than woody species. If future work identifies that similar niche shifts are observed between congeners of other taxa, such as woody species, it would indicate that congeners are potentially a reliable resource for assessing the risk of species niche shifts.
Conclusions
We examined climatic niche shifts in three congeners across three introduced ranges and demonstrated that not only do the species occupy different niches in the introduced range than in the native range, but that the direction and magnitude of these niche shifts differs between ranges. Though there were qualitative similarities within a range between congeners, the degree of niche overlap was generally moderate or low. This indicates that the processes that drive niche shifts, including pre-adaptation to climates outside the native range, biotic interactions, founder effects or genetic admixture, and rapid evolution are not consistent across invasions even within a species. Understanding which processes are driving these shifts and how they are influenced by their environmental context will improve our ability to predict and manage invasions, including through more accurate modelling of species’ expected distributions. In the meantime, data from closely related species may provide some qualitative indications of likely niche shifts, which can be used to guide monitoring efforts, and supplement individual species distribution models.
Availability of data and material
The datasets generated and analysed during the current study are available in the Figshare repositories: https://doi.org/10.6084/m9.figshare.17148254 & https://doi.org/10.6084/m9.figshare.17148317
Code availability
The code generated during the current study is available upon reasonable request.
Change history
13 October 2022
Missing Open Access funding information has been added in the Funding Note
References
AFE (1979) Atlas Florae Europaeae. Distribution of vascular plants in Europe. 4. Polygonaceae. In Jalas J & Suominen J (eds), Atlas Florae Europaeae, The Committee for Mapping the Flora of Europe & Societas Biologica Fennica Vanamo, p. maps 384–478
Aguirre-Gutiérrez J, Serna-Chavez HM, Villalobos-Arambula AR, Pérez de la Rosa JA, Raes N (2015) Similar but not equivalent: ecological niche comparison across closely-related Mexican white pines. Divers Distrib 21(3):245–257. https://doi.org/10.1111/ddi.12268
Alexander JM (2013) Evolution under changing climates: climatic niche stasis despite rapid evolution in a non-native plant. Proc R Soc B Biol Sci. https://doi.org/10.1098/rspb.2013.1446
Alexander JM, Diez JM, Usinowicz J, Hart SP (2018) Species’ distributions as a coexistence problem: a response to Godsoe et al. In Trends in Ecology and Evolution. vol 33, issue 3, Elsevier Ltd, pp 144–145. https://doi.org/10.1016/j.tree.2018.01.001
Atlas of Living Australia. (2019). https://www.ala.org.au/
Atwater DZ, Ervine C, Barney JN (2018) Climatic niche shifts are common in introduced plants. Nat Ecol Evolut 2:34–43. https://doi.org/10.1038/s41559-017-0396-z
Atwater DZ, Barney JN (2021) Climatic niche shifts in 815 introduced plant species affect their predicted distributions. Glob Ecol Biogeogr. https://doi.org/10.1111/geb.13342
Australia’s Virtual Herbarium (2019) MEL 0610742A. Council of Heads of Australasian Herbaria. http://avh.ala.org.au/
Baskin JM, Baskin CC (1978) A contribution to the germination ecology of Rumex crispus L. Bull Torrey Bot Club 105(4):278. https://doi.org/10.2307/2484920
Bates OK, Bertelsmeier C (2021) Climatic niche shifts in introduced species. Curr Biol 31(19):R1252–R1266. https://doi.org/10.1016/J.CUB.2021.08.035
Benvenuti S, Macchia M, Miele S (2001) Light, temperature and burial depth effects on Rumex obtusifolius seed germination and emergence. Weed Res 41(2):177–186. https://doi.org/10.1046/j.1365-3180.2001.00230.x
Boheemen LA, Atwater DZ, Hodgins KA (2019) Rapid and repeated local adaptation to climate in an invasive plant. New Phytol 222(1):614–627. https://doi.org/10.1111/nph.15564
Booth TH, Nix HA, Busby JR, Hutchinson MF (2014) Bioclim: the first species distribution modelling package, its early applications and relevance to most current MaxEnt studies. Divers Distrib. https://doi.org/10.1111/ddi.12144
Bradley BA, Blumenthal DM, Wilcove DS, Ziska LH (2010a) Predicting plant invasions in an era of global change. Trends Ecol Evol 25(5):310–318. https://doi.org/10.1016/J.TREE.2009.12.003
Bradley BA, Wilcove DS, Oppenheimer M (2010b) Climate change increases risk of plant invasion in the Eastern United States. Biol Invasions 12(6):1855–1872. https://doi.org/10.1007/s10530-009-9597-y
Bradley BA, Blumenthal DM, Early R, Grosholz ED, Lawler JJ, Miller LP, Sorte CJ, D’Antonio CM, Diez JM, Dukes JS, Ibanez I, Olden JD (2012) Global change, global trade, and the next wave of plant invasions. Front Ecol Environ 10(1):20–28. https://doi.org/10.1890/110145
Broennimann O, Fitzpatrick MC, Pearman PB, Petitpierre B, Pellissier L, Yoccoz NG, Thuiller W, Fortin MJ, Randin C, Zimmermann NE, Graham CH, Guisan A (2012) Measuring ecological niche overlap from occurrence and spatial environmental data. Glob Ecol Biogeogr. https://doi.org/10.1111/j.1466-8238.2011.00698.x
Bujan J, Charavel E, Bates OK, Gippet J, Darras H, Lebas C, Bertelsmeier C (2020) Increased acclimation ability accompanies a thermal niche shift of a recent invasion. J Anim Ecol 1365–2656:13381. https://doi.org/10.1111/1365-2656.13381
Bulleri F, Bruno JF, Silliman BR, Stachowicz JJ (2016) Facilitation and the niche: implications for coexistence, range shifts and ecosystem functioning. Funct Ecol 30(1):70–78. https://doi.org/10.1111/1365-2435.12528
CABI (2019) Invasive Species Compendium. CAB International. www.cabi.org/isc
Calflora (2019) Search for Plants. https://calflora.org//
Catford JA, Wilson JRU, Pyšek P, Hulme PE, Duncan RP (2022) Addressing context dependence in ecology. Trends Ecol Evol 37(2):158–170. https://doi.org/10.1016/J.TREE.2021.09.007
Cavers PB, Harper JL (1964) Rumex Obtusifolius L. and R. Crispus L. J Ecol 52(325):737–766
Cavers PB, Harper JL (1966) Germination Polymorphism in Rumex Crispus and Rumex Obtusifolius. J Ecol 54(2):367. https://doi.org/10.2307/2257955
Christina M, Limbada F, Atlan A (2019) Climatic niche shift of an invasive shrub (gorse, Ulex europaeus): a world scale comparison in native and introduced regions. https://hal.archives-ouvertes.fr/hal-02146154/
Clements DR, Jones VL (2021) Rapid evolution of invasive weeds under climate change: present evidence and future research needs. Front Agron. https://doi.org/10.3389/FAGRO.2021.664034
Comte L, Cucherousset J, Olden JD (2017) Global test of Eltonian niche conservatism of nonnative freshwater fish species between their native and introduced ranges. Ecography 40(3):384–392. https://doi.org/10.1111/ecog.02007
Costan C-A, Godsoe WK, Bufford JL, Marris JWM, Hulme PE (2022) Can the enemy release hypothesis explain the success of Rumex (Polygonaceae) species in an introduced range? Biol Invasions 2022:1–13. https://doi.org/10.1007/S10530-022-02810-W
D’Amen M, Azzurro E (2020) Integrating univariate niche dynamics in species distribution models: a step forward for marine research on biological invasions. J Biogeogr 47(3):686–697. https://doi.org/10.1111/JBI.13761
Dainese M, Aikio S, Hulme PE, Bertolli A, Prosser F, Marini L (2017) Human disturbance and upward expansion of plants in a warming climate. Nat Clim Change 7(8):577–580. https://doi.org/10.1038/nclimate3337
Darwin C, Keynes RD (1835) Charles Darwin’s Beagle diary. Cambridge University Press, Cambridge
Datta A, Schweiger O, Kühn I (2019) Niche expansion of the invasive plant species Ageratina adenophora despite evolutionary constraints. J Biogeogr 46(7):1306–1315. https://doi.org/10.1111/jbi.13579
de Bello F, Lavorel S, Lavergne S, Albert CH, Boulangeat I, Mazel F, Thuiller W (2013) Hierarchical effects of environmental filters on the functional structure of plant communities: a case study in the French Alps. Ecography 36(3):393–402. https://doi.org/10.1111/j.1600-0587.2012.07438.x
Di Cola V, Broennimann O, Petitpierre B, Breiner FT, D’Amen M, Randin C, Engler R, Pottier J, Pio D, Dubuis A, Pellissier L, Mateo RG, Hordijk W, Salamin N, Guisan A (2017) ecospat: an R package to support spatial analyses and modeling of species niches and distributions. Ecography 40(6):774–787. https://doi.org/10.1111/ecog.02671
Dullinger I, Wessely J, Bossdorf O, Dawson W, Essl F, Gattringer A, Klonner G, Kreft H, Kuttner M, Moser D, Pergl J, Pyšek P, Thuiller W, van Kleunen M, Weigelt P, Winter M, Dullinger S (2017) Climate change will increase the naturalization risk from garden plants in Europe. Glob Ecol Biogeogr 26(1):43–53. https://doi.org/10.1111/geb.12512
Early R, Sax DF (2014) Climatic niche shifts between species’ native and naturalized ranges raise concern for ecological forecasts during invasions and climate change. Glob Ecol Biogeogr 23(12):1356–1365. https://doi.org/10.1111/geb.12208
Essl F, Lenzner B, Bacher S, Bailey S, Capinha C, Daehler C, Dullinger S, Genovesi P, Hui C, Hulme PE, Jeschke JM, Katsanevakis S, Kühn I, Leung B, Liebhold A, Liu C, MacIsaac HJ, Meyerson LA, Nuñez MA, Roura-Pascual N (2020) Drivers of future alien species impacts: an expert-based assessment. Glob Change Biol 26(9):4880–4893. https://doi.org/10.1111/gcb.15199
Fernández M, Hamilton H (2015) Ecological niche transferability using invasive species as a case study. PLoS ONE 10(3):e0119891. https://doi.org/10.1371/journal.pone.0119891
Fick SE, Hijmans R (2017) Worldclim 2: New 1-km spatial resolution climate surfaces for global land areas. Int J Climatol 37(12):4302–4315
Flanagan BA, Krueger-Hadfield SA, Murren CJ, Nice CC, Strand AE, Sotka EE (2021) Founder effects shape linkage disequilibrium and genomic diversity of a partially clonal invader. Mol Ecol 30(9):1962–1978. https://doi.org/10.1111/MEC.15854
Flora of Japan (2019) Database of Japanese Flora. https://herbaria.plants.ox.ac.uk/bol/florajapan/Explore
Fourcade Y, Besnard AG, Secondi J (2018) Paintings predict the distribution of species, or the challenge of selecting environmental predictors and evaluation statistics. Glob Ecol Biogeogr 27(2):245–256. https://doi.org/10.1111/geb.12684
Gall L (2019) Occurrence dataset. Botany Division, Yale Peabody Museum. Yale University Peabody Museum. https://doi.org/10.15468/hrztgn
Gallagher RV, Beaumont LJ, Hughes L, Leishman MR (2010) Evidence for climatic niche and biome shifts between native and novel ranges in plant species introduced to Australia. J Ecol 98(4):790–799. https://doi.org/10.1111/j.1365-2745.2010.01677.x
GBIF.org. (22 March 2020) GBIF Occurrence Download. https://doi.org/10.15468/dl.go0jrh
Godsoe W (2010) Regional variation exaggerates ecological divergence in niche models. Syst Biol 59(3):298–306. https://doi.org/10.1093/sysbio/syq005
Godsoe W, Jankowski J, Holt RD, Gravel D (2018) Which coexistence mechanisms should biogeographers quantify? A reply to Alexander et al. Trends Ecol Evol 33:145–147. https://doi.org/10.1016/j.tree.2018.01.001
Grant S, Niezgoda C (2019) Occurrence dataset. Field museum of natural history (Botany) seed plant collection. Version 11.10. Field Museum. https://doi.org/10.15468/nxnqzf
Grime JP, Hodgson JG, Hunt R (2007) Comparative plant ecology: a functional approach to common British species, 2nd edn. Castlepoint Press, Bruce
Grossrieder M, Keary IP (2004) The potential for the biological control of Rumex obtusifolius and Rumex crispus using insects in organic farming, with particular reference to Switzerland. Biocontrol News Inf 25(3):65–79
Guisan A, Petitpierre B, Broennimann O, Kueffer C, Randin C, Daehler C (2012) Response to comment on “climatic niche shifts are rare among terrestrial plant invaders.” Science 338(6104):193–193. https://doi.org/10.1126/SCIENCE.1226051
Guisan A, Petitpierre B, Broennimann O, Daehler C, Kueffer C (2014) Unifying niche shift studies: insights from biological invasions. Trends Ecol Evol 29(5):260–269. https://doi.org/10.1016/J.TREE.2014.02.009
Guo W-Y, Lambertini C, Li X-Z, Meyerson LA, Brix H (2013) Invasion of Old World Phragmites australis in the New World: precipitation and temperature patterns combined with human influences redesign the invasive niche. Glob Change Biol. https://doi.org/10.1111/gcb.12295
Hengl T, Kempen B, Heuvelink GBM, Malone B (2014) GSIF: global soil information facilities. (R version: 0.4–1.)
Hill MP, Gallardo B, Terblanche JS (2017) A global assessment of climatic niche shifts and human influence in insect invasions. Glob Ecol Biogeogr 26(6):679–689. https://doi.org/10.1111/geb.12578
Hill MO, Mountford JO, Roy DB, Bunce RGH (1999) Ellenberg’s indicator values for British plants. ECOFACT technical annex. (Vol. 2). Institute of Terrestrial Ecology
Holm LG, Doll J, Holm E, Pancho JV, Herberger JP (1997) World weeds: natural histories and distribution. Wiley, Hoboken
Holm LG, Plucknett DL, Pancho JV, Herberger JP (1977) The World’s worst weeds: distribution and biology. The University Press of Hawaii, Honolulu
Holm L, Pancho JV, Herberger JP, Plucknett DL (1979) A geographical atlas of world weeds. John Wiley and Sons. https://www.cabi.org/isc/abstract/19802330214
Holt RD (2009) Bringing the Hutchinsonian niche into the 21st century: Ecological and evolutionary perspectives. Proc Natl Acad Sci. https://doi.org/10.1073/pnas.0905137106
Hulme PE, Barrett SCH (2013) Integrating trait- and niche-based approaches to assess contemporary evolution in alien plant species. J Ecol 101(1):68–77. https://doi.org/10.1111/1365-2745.12009
Hultén E (1950) Atlas of the distribution of vascular plants in North-West Europe. Esselte AB
Hutchinson GE (1957) Concluding remarks cold spring harbour symposia on quantitative biology. Symp Quant Biol 22:415–427
Invasive Species Specialist Group (2019) Global Invasive Species Database. http://issg.org/database/welcome/
Keller SR, Sowell DR, Neiman M, Wolfe LM, Taylor DR (2009) Adaptation and colonization history affect the evolution of clines in two introduced species. New Phytol 183(3):678–690. https://doi.org/10.1111/J.1469-8137.2009.02892.X
Kitajima K, Fox AM, Sato T, Nagamatsu D (2006) Cultivar selection prior to introduction may increase invasiveness: evidence from Ardisia crenata. Biol Invasions 8(7):1471–1482. https://doi.org/10.1007/s10530-005-5839-9
Koide D, Yoshida K, Daehler CC, Mueller-Dombois D (2017) An upward elevation shift of native and non-native vascular plants over 40 years on the island of Hawai’i. J Veg Sci 28(5):939–950. https://doi.org/10.1111/JVS.12549
Kolanowska M, Konowalik K (2014) Niche conservatism and future changes in the potential area coverage of Arundina graminifolia, an invasive orchid species from southeast Asia. Biotropica 46(2):157–165. https://doi.org/10.1111/BTP.12089
Kołodziejek J, Patykowski J (2015) Effect of environmental factors on germination and emergence of invasive Rumex confertus in central Europe. Sci World J 2015:170176. https://doi.org/10.1155/2015/170176
Liao ZY, Scheepens JF, Li QM, Wang WB, Feng YL, Zheng YL (2020) Founder effects, post-introduction evolution and phenotypic plasticity contribute to invasion success of a genetically impoverished invader. Oecologia 192(1):105–118. https://doi.org/10.1007/S00442-019-04566-Y/TABLES/3
Liu X, Petitpierre B, Broennimann O, Li X, Guisan A, Li Y (2016) Realized climatic niches are conserved along maximum temperatures among herpetofaunal invaders. J Biogeogr 44(1):111–121. https://doi.org/10.1111/jbi.12808
Liu C, Wolter C, Xian W, Jeschke JM (2020) Most invasive species largely conserve their climatic niche. Proc Natl Acad Sci 117(38):23643–23651. https://doi.org/10.1073/PNAS.2004289117
Lousley JE, Kent DH (1981) Docks and knotweeds of the British Isles. Botanical Society of the British Isles, Durham
Luo X, Xu X, Zheng Y, Guo H, Hu S (2019) The role of phenotypic plasticity and rapid adaptation in determining invasion success of Plantago virginica. Biol Invasions 21(8):2679–2692. https://doi.org/10.1007/s10530-019-02004-x
MacLean SA, Beissinger SR, Sarah MacLean CA, Chancellor B (2017) Species’ traits as predictors of range shifts under contemporary climate change: a review and meta-analysis. Glob Change Biol 23(10):4094–4105. https://doi.org/10.1111/GCB.13736
Mairal M, Chown SL, Shaw J, Chala D, Chau JH, Hui C, Kalwij JM, Münzbergová Z, Jansen van Vuuren B, Le Roux JJ (2021) Human activity strongly influences genetic dynamics of the most widespread invasive plant in the sub-Antarctic. Mol Ecol 00:1–17. https://doi.org/10.1111/MEC.16045
Makuchi T, Sakai H (1984) Seedling survival and flowering of Rumex obtusifolius L.* in various habitats. Weed Research (Japan), 29
Miyagi A, Takahara K, Takahashi H, Kawai-Yamada M, Uchimiya H (2010) Targeted metabolomics in an intrusive weed, Rumex obtusifolius L., grown under different environmental conditions reveals alterations of organ related metabolite pathway. Metabolomics 6(4):497–510. https://doi.org/10.1007/s11306-010-0220-0
Mota-Vargas C, Rojas-Soto OR (2016) Taxonomy and ecological niche modeling: Implications for the conservation of wood partridges (genus Dendrortyx). J Nat Conserv 29:1–13. https://doi.org/10.1016/j.jnc.2015.10.003
New Zealand Plant Conservation Network (2019a) New Zealand’s Flora. http://www.nzpcn.org.nz/page.aspx?flora
New Zealand Plant Conservation Network (2019b) Rumex conglomeratus. http://www.nzpcn.org.nz/flora_details.aspx?ID=2937
New Zealand Plant Conservation Network (2019c) Rumex crispus. http://www.nzpcn.org.nz/flora_details.aspx?ID=2948
Nishida T (2002) Alien plants invasion of forage crop fields and artificial grasslands-distribution and routes of invasions. Grassl Sci 48(2):168–172
Olson DM, Dinerstein E, Wikramanayake ED, Burgess ND, Powell GVN, Underwood EC, D’amico JA, Itoua I, Strand HE, Morrison JC, Loucks CJ, Allnutt TF, Ricketts TH, Kura Y, Lamoreux JF, Wettengel WW, Hedao P, Kassem KR (2001) Terrestrial ecoregions of the world: a new map of life on EarthA new global map of terrestrial ecoregions provides an innovative tool for conserving biodiversity. Bioscience 51(11):933–938. https://doi.org/10.1641/0006-3568(2001)051[0933:teotwa]2.0.co;2
Petitpierre B, Kueffer C, Broennimann O, Randin C, Daehler C, Guisan A (2012) Climatic niche shifts are rare among terrestrial plant invaders. Science (new York, N.y.) 335(6074):1344–1348. https://doi.org/10.1126/science.1215933
Pili AN, Tingley R, Sy EY, Diesmos MLL, Diesmos AC (2020) Niche shifts and environmental non-equilibrium undermine the usefulness of ecological niche models for invasion risk assessments. Sci Rep 10(1):1–18. https://doi.org/10.1038/s41598-020-64568-2
Pysek P, Jarosík V, Hulme PE, Kühn I, Wild J, Arianoutsou M, Bacher S, Chiron F, Didziulis V, Essl F, Genovesi P, Gherardi F, Hejda M, Kark S, Lambdon PW, Desprez-Loustau M-L, Nentwig W, Pergl J, Poboljsaj K, Winter M (2010) Disentangling the role of environmental and human pressures on biological invasions across Europe. Proc Natl Acad Sci USA 107(27):12157–12162. https://doi.org/10.1073/pnas.1002314107
R Core Team (2013) R: A language and environment for statistical computing. R Foundation for Statistical Computing. http://www.r-project.org/
Randall RP (2017) A global compendium of weeds In: Randall RP (ed), 3rd edn. CABI
Randin CF, Dirnböck T, Dullinger S, Zimmermann NE, Zappa M, Guisan A (2006) Are niche-based species distribution models transferable in space? J Biogeogr 33(10):1689–1703. https://doi.org/10.1111/j.1365-2699.2006.01466.x
Redpath D, Rapson G (2015) An extreme flood event initiates a decade of stand collapse in Beilschmiedia tawa forest, Turakina Valley, Rangitikei, New Zealand. NZ J Bot 53(1):38–59. https://doi.org/10.1080/0028825X.2014.1000934
Ribas LG, de Cássia-Silva C, Petsch DK, Silveira MJ, Lima-Ribeiro MS (2018) The potential invasiveness of an aquatic macrophyte reflects founder effects from native niche. Biol Invasions 20(11):3347–3355. https://doi.org/10.1007/S10530-018-1780-6/FIGURES/2
Root TL, Price JT, Hall KR, Schneider SH, Rosenzweig C, Pounds JA (2003) Fingerprints of global warming on wild animals and plants. Nature 421(6918):57–60. https://doi.org/10.1038/nature01333
Schoener TW (1970) Nonsynchronous spatial overlap of lizards in patchy habitats. Ecology 51(3):408–418. https://doi.org/10.2307/1935376
Seebens H, Blackburn TM, Dyer EE, Genovesi P, Hulme PE, Jeschke JM, Pagad S, Pyšek P, van Kleunen M, Winter M, Ansong M, Arianoutsou M, Bacher S, Blasius B, Brockerhoff EG, Brundu G, Capinha C, Causton CE, Celesti-Grapow L, Essl F (2018) Global rise in emerging alien species results from increased accessibility of new source pools. Proc Natl Acad Sci USA 115(10):E2264–E2273. https://doi.org/10.1073/pnas.1719429115
Silva DP, Vilela B, Buzatto BA, Moczek AP, Hortal J (2016) Contextualized niche shifts upon independent invasions by the dung beetle Onthophagus taurus. Biol Invasions 18(11):3137–3148. https://doi.org/10.1007/s10530-016-1204-4
Smith AB, Godsoe W, Rodríguez-Sánchez F, Wang H-H, Warren D (2019) Niche estimation above and below the species level. Trends Ecol Evol 34(3):260–273. https://doi.org/10.1016/J.TREE.2018.10.012
Stephan P, Mora BB, Alexander JM (2021) Positive species interactions shape species’ range limits. Oikos 130(10):1611–1625. https://doi.org/10.1111/OIK.08146
Tingley R, Vallinoto M, Sequeira F, Kearney MR (2014) Realized niche shift during a global biological invasion. PNAS 111(28):10233–10238. https://doi.org/10.1073/pnas.1405766111
Tingley R, Thompson MB, Hartley S, Chapple DG (2016) Patterns of niche filling and expansion across the invaded ranges of an Australian lizard. Ecography 39(3):270–280. https://doi.org/10.1111/ECOG.01576
Troudet J, Grandcolas P, Blin A, Vignes-Lebbe R, Legendre F (2017) Taxonomic bias in biodiversity data and societal preferences. Sci Rep 7(1):9132. https://doi.org/10.1038/s41598-017-09084-6
University of Georgia (2019) EDDMapS species distribution maps. Centre for Invasive Species and Ecosystem Health. https://www.eddmaps.org/distribution/
USDA (2019) Taxon: Rumex crispus L. GRIN. https://npgsweb.ars-grin.gov/gringlobal/taxon/taxonomydetail?id=32530
USDA, & NRCS (2019) The PLANTS Database. National Plant Data Team, Greensboro, NC 27401–4901 USA. http://plants.usda.gov
Vibrans H (1998) Native maize field weed communities in south-central Mexico. Weed Res 38(2):153–166. https://doi.org/10.1046/j.1365-3180.1998.00082.x
Victorian Biodiversity Atlas (2019a) 2447402.00. The State of Victoria, Department of Environment, Land, Water & Planning
Victorian Biodiversity Atlas (2019b) 2448526.00. The State of Victoria, Department of Environment, Land, Water & Planning
Warren DL, Glor RE, Turelli M (2008) Environmental niche equivalency versus conservatism: quantitative approaches to niche evolution. Evolution 62(11):2868–2883. https://doi.org/10.1111/j.1558-5646.2008.00482.x
Warren DL, Glor RE, Turelli M (2010) ENMTools: a toolbox for comparative studies of environmental niche models. Ecography 33(3):607–611. https://doi.org/10.1111/j.1600-0587.2009.06142.x
Webber BL, Le Maitre DC, Kriticos DJ (2012) Comment on Climatic niche shifts are rare among terrestrial plant invaders, Science (New York, NY), 338(6104), 193; author reply 193. https://doi.org/10.1126/science.1225980
Acknowledgements
This project was funded by a Tertiary Education Commission grant to the Bio-Protection Research Centre as a Centre of Research Excellence. We thank the following people and organizations who helped us locate and survey additional populations: Botanical Society of the British Isles, Scottish Natural Heritage and Historic Environment Scotland, Christchurch and Invercargill City Councils, Taane Johnsen & Colin Ferguson, Andrei Costan, and numerous farm owners and managers around New Zealand. We are grateful to Prof. Jake Alexander, Prof. Stephen Hartley, and three anonymous reviewers, whose comments substantially improved this work.
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions. This project was supported by the New Zealand Tertiary Education Commission CoRE grant to the Bio-Protection Research Centre.
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design. Philip Hulme obtained the research funding. Data collection and analysis were performed by Thomas Carlin and Jennifer Bufford. The first draft of the manuscript was written by Thomas Carlin and all authors participated in revisions and approved the final manuscript.
Corresponding author
Ethics declarations
Conflicts of interest
The authors have no conflicts of interest to declare that are relevant to the content of this article.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Carlin, T.F., Bufford, J.L., Hulme, P.E. et al. Global assessment of three Rumex species reveals inconsistent climatic niche shifts across multiple introduced ranges. Biol Invasions 25, 79–96 (2023). https://doi.org/10.1007/s10530-022-02893-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-022-02893-5