
Abstract
Feline coronavirus (FCoV) has been classified into two biotypes: avirulent feline coronavirus (feline enteric coronavirus: FECV) and virulent feline coronavirus (feline infectious peritonitis virus: FIPV). In FIPV infection, antibody-dependent enhancement (ADE) has been reported and was shown to be associated with severe clinical disease. On the other hand, the potential role of ADE in FECV infection has not been examined. In this study, using laboratory strains of serotype II FIPV WSU 79-1146 (FIPV 79-1146) and serotype II FECV WSU 79-1683 (FECV 79-1683), we investigated the relationship between ADE and induction of inflammatory cytokines, which are pathogenesis-related factors, for each strain. As with ADE of FIPV 79-1146 infection, a monoclonal antibody against the spike protein of FCoV (mAb 6-4-2) enhanced FECV 79-1683 replication in U937 cells and primary feline monocytes. However, the ADE activity of FECV 79-1683 was lower than that of FIPV 79-1146. Moreover, mRNA levels of inflammatory cytokines (TNF-α, IL-1β, and IL-6) significantly increased with ADE of FIPV 79-1146 infection in primary feline monocytes, but FECV 79-1683 did not demonstrate an increase in these levels. In conclusion, infection of monocytes by FECV was enhanced by antibodies, but the efficiency of infection was lower than that of FIPV.
Similar content being viewed by others

Introduction
Feline coronavirus (FCoV) is an enveloped positive-strand RNA virus belonging to the family Coronaviridae, subfamily Coronavirinae, and genus Alphacoronavirus [7]. There are two serotypes of FCoV: FCoV has been classified into types I and II based on the amino acid sequence of its spike (S) protein [11, 18]. Separate from these serotypes, FCoV has been classified into two biotypes: weakly pathogenic feline enteric coronavirus (FECV; avirulent FCoV) and strongly pathogenic feline infectious peritonitis virus (FIPV; virulent FCoV) [20]. FIP is a lethal, immune-mediated infectious disease of members of the family Felidae, and there has been no established therapy for treatment. It has been suggested that FIPV is a mutant of FECV, i.e., it is considered that after infecting a cat, the open reading frame (ORF)2 gene and ORF3c gene of FECV partially mutates in the cat’s body and becomes virulent FCoV, which is FIPV [21]. The presence of FCoVs with a broad pathogenic spectrum in the field has also been proposed as another hypothesis [2]. There are serotype I and II FECV and FIPV in FCoV. Serotype II FCoVs are known to utilize feline aminopeptidase N (fAPN) as a virus receptor [6, 8].
There are 11 ORFs in the FCoV genome, and of these, ORF2, ORF4, ORF5, and ORF6 encode structural proteins [5]. The S protein, encoded by ORF2, plays an essential role in cell entry, being involved in binding to the FCoV-specific receptor on the host cell surface and fusion with the cell membrane [1]. The S protein also plays an important role in antibody-dependent enhancement (ADE) of FIPV infection [9, 19, 34]. ADE is a phenomenon in which binding to an antibody makes host cell entry easier for the virus. We previously reported that monoclonal antibodies (mAbs) to the S protein mediated ADE of FIPV infection [9]. When ADE of FIPV infection occurs, virus production in macrophages increases, and production of inflammatory cytokines, such as tumor necrosis factor alpha (TNF-α), interleukin (IL)-1β, and IL-6, is enhanced [27, 30, 31]. Therefore, ADE is closely associated with the severity of FIPV infection. However, no studies on the difference in pathogenicity between FIPV and FECV with regard to ADE of infection have been reported.
Since serotype I FCoV infection is dominant in the field [10, 14, 15, 25], it is desirable to use serotype I FCoV in studies on FIP. However, isolation of serotype I FECV using cultured cell lines has not been successful, and it is difficult to study ADE of infection in vitro. Moreover, an mAb inducing ADE activity of serotype I FCoV has not yet been prepared.
In this study, using laboratory strains of serotype II FIPV WSU 79-1146 (FIPV 79-1146) and serotype II FECV WSU 79-1683 (FECV 79-1683), we investigated the relationship between ADE and induction of inflammatory cytokines, which are pathogenesis-related factors, in each strain.
Materials and methods
Cell cultures and viruses
Felis catus whole fetus (fcwf)-4 cells (kindly supplied by Dr. M. C. Horzinek of the State University of Utrecht) were grown in Eagle’s minimum essential medium containing 50% L-15 medium, 5% fetal calf serum (FCS), 100 U of penicillin per mL, and 100 μg of streptomycin per mL. Cells of the human monocyte cell line U937 were cultured in RPMI 1640 medium containing 10% FCS and antibiotics. Primary feline monocytes were maintained in RPMI 1640 growth medium supplemented with 10% FCS, 100 U of penicillin per mL, 100 μg of streptomycin per mL, and 50 μM 2-mercaptoethanol. FIPV 79-1146 was kindly provided by Dr. M. C. Horzinek. FECV 79-1683 was kindly supplied by Dr. A. J. McKeirnan of Washington State University. These viruses were grown in fcwf-4 cells at 37 °C with 5% CO2.
Antibodies
mAb 6-4-2 (IgG2a) used in the present study recognizes the S protein of serotype II FCoV [9]. It has been reported that mAb 6-4-2 exhibits neutralizing activity in fcwf-4 and CrFK cells but enhancing activity in primary feline monocytes and macrophages depending on the reaction conditions. The mAb 6-4-2 was used at a dilution of 10 except in the experiment shown in Fig. 1A. mAb R-G-4 (recognizing fAPN; IgG1) and IgG1 mAb control (recognizing feline interferon-gamma) prepared by our laboratory [8] were used.

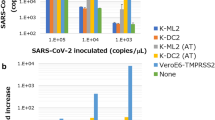
ADE of FECV infection in U937 cells and feline monocytes. (A) U937 cells were infected with FCoV in the presence or absence of mAb 6-4-2. The results are shown as the mean ± SE (n = 5). (B) Feline monocytes were infected with FCoV in the presence or absence of mAb 6-4-2. The results are shown as the mean ± SE (n = 10). Black bar, FIPV 79-1146; white bar, FECV 79-1683; N.D., not detected
Inoculation of U937 cells with FCoV
FIPV 79-1146 or FECV 79-1683 (1 × 106 TCID50 for both) reacted with mAb 6-4-2 at 4 °C for 1 h was added to the culture and adsorbed to the U937 cells (2 × 105 cells) in tubes at 37 °C with 5% CO2 for 3 h. The cells were washed three times with PBS and cultured in tubes at 37 °C with 5% CO2 for 48 h, and the supernatants were collected. The virus titer in the culture supernatant (TCID50) was determined by the method of Reed and Muench [22] with fcwf-4 cells.
Inoculation of primary feline monocytes with FCoV
Primary feline monocytes were isolated from specific-pathogen-free (SPF) cats as described previously by Dewerchin et al. [4]. The blood sampling in SPF cats was performed in accordance with the Guidelines for Animal Experiments of Kitasato University (approval number, 16-086). FIPV 79-1146 or FECV 79-1683 (1 × 104 TCID50 for both) reacted with mAb 6-4-2 at 4 °C for 1 h was added to the culture and adsorbed to the primary feline monocytes (2 × 105 cells) in 24-well multi-plates at 37 °C with 5% CO2 for 1 h. The cells were washed three times with PBS and cultured in 24-well multi-plates at 37 °C with 5% CO2 for 48 h, and the supernatants and cells were collected. The virus titer in the culture supernatant (TCID50) was determined by the method of Reed and Muench [22] with fcwf-4 cells. Cells were used to measure the mRNA expression levels of inflammatory cytokines.
Effects of anti-fAPN mAb in primary feline monocytes infected with FCoV
Primary feline monocytes (2 × 105 cells) were cultured in medium containing mAb R-G-4 or IgG1 mAb control in 24-well multi-plates at 4 °C for 1 hour. After washing with PBS, FIPV 79-1146 or FECV 79-1683 (1 x 104 TCID50 for both) with or without pretreatment with mAb 6-4-2 was added to the culture and allowed to adsorb to the cells at 37 °C with 5% CO2 for 1 h in the presence of mAbs. After washing with PBS, the cells were cultured in the medium, and the culture supernatants were collected after 48 h. The virus titer in the culture supernatant (TCID50) was determined by the method of Reed and Muench [22] with fcwf-4 cells.
Effects of lysosomotropic agents on the virus in primary feline monocytes with FCoV
Primary feline monocytes (2 × 105 cells) were cultured in medium containing chloroquine (Wako Pure Chemical Industries, Japan) or ammonium chloride (Wako Pure Chemical Industries, Japan) in 24-well multi-plates at 37 °C with 5% CO2 for 1 h. After washing with PBS, FIPV 79-1146 or FECV 79-1683 (1 × 104 TCID50 for both) with or without pretreatment with mAb 6-4-2 was added to the culture and allowed to adsorb to the cells at 37 °C with 5% CO2 for 1 h in the presence of chloroquine or ammonium chloride. After washing with PBS, the cells were cultured in the medium containing chloroquine or ammonium chloride, and the culture supernatants were collected after 48 h. The virus titer in the culture supernatant (TCID50) was determined by the method of Reed and Muench [22] with fcwf-4 cells. To evaluate the cytotoxic effects of chloroquine and ammonium chloride in primary feline monocytes, cell viability was measured by the WST-8 assay as described before [29].
RNA isolation and cDNA preparation
RNA isolation and cDNA preparation were performed employing the method of Takano et al. [29].
Determination of levels of feline GAPDH mRNA, TNF-α mRNA, IL-1β mRNA, and IL-6 mRNA expression
cDNA was amplified by PCR using specific primers for feline glyceraldehyde 3-phosphate dehydrogenase (GAPDH) mRNA, TNF-α mRNA, IL-1β mRNA, and IL-6 mRNA. The primer sequences are shown in Table 1. PCR was performed using the method of Takano et al. [32]. The band density was quantified under appropriate UV exposure by video densitometry using Image J software (NIH, USA). TNF-α mRNA, IL-1β mRNA, and IL-6 mRNA were semi-quantitatively analyzed in terms of the relative density value to that of the mRNA for the housekeeping gene GAPDH.
Statistical analysis
Data from two groups were analyzed by Student’s t-test, and multiple groups were analyzed by one-way ANOVA.
Results
ADE of FECV infection in U937 cells and primary feline monocytes
As U937 cells do not express fAPN, FIPV alone does not infect U937 cells, but it enters FIPV via the U937 cell-surface Fc receptor when anti-FCoV S antibodies are present [12, 31]. therefore, U937 cells were used as a prediction model to determine the mAb 6-4-2 concentrations required for ADE of FECV infection in vitro (Fig. 1A). FCoVs did not infect U937 cells in the absence of mAb 6-4-2. Several dilutions of mAb 6-4-2 were incubated with the virus. The titer of FIPV 79-1146 was increased in the culture supernatant at several dilutions of mAb 6-4-2 (1:1, 7.7 × 102 ± 3.9 × 102 TCID50/mL; 1:10, 1.4 × 103 ± 6.2 × 102 TCID50/mL; 1:100, 8.3 × 102 ± 8.2 × 101 TCID50/mL). In contrast to FIPV 79-1146, the titer of FECV 79-1683 did not increase at any dilution except 10−1 (8.3 × 100 ± 1.6 × 100 TCID50/mL), i.e. maximum ADE activity of FECV 79-1683 infection occurred at an mAb dilution of 1:10. We investigated the ADE activity of FECV infection in primary feline monocytes (Fig. 1B). mAb 6-4-2 (dilution of 1:10) enhanced the infection of monocytes with FIPV 79-1146; the median increase was 200-fold for the infection by virus alone (virus alone, 2.1 × 103 ± 7.2 × 102 TCID50/mL; virus and mAb 6-4-2, 6.1 × 105 ± 1.6 × 105 TCID50/mL). On the other hand, mAb 6-4-2 (dilution of 1:10) slightly increased the infection of cells with FECV 79-1683; the median increase was tenfold (virus alone, 1.3 × 102 ± 5.9 × 101 TCID50/mL; virus and mAb 6-4-2, 1.5 × 103 ± 9.9 × 102 TCID50/mL), i.e., the ADE activity of FECV 79-1683 was 10 to 20-fold lower than that of FIPV 79-1146.
Involvement of the virus receptor fAPN in ADE of FECV infection in primary feline monocytes
The involvement of virus receptor fAPN in ADE of FECV infection in primary feline monocytes was investigated (Fig. 2). When monocytes were allowed to react with mAb R-G-4 beforehand and then inoculated with FECV 79-1683 alone, the virus titer decreased significantly (virus alone, 1.5 × 102 ± 5.2 × 101 TCID50/mL; virus and mAb R-G-4, < 100 TCID50/mL). However, when monocytes were allowed to react with mAb R-G-4 beforehand and then inoculated with a mixture of FECV 79-1683 and mAb 6-4-2, the virus titer did not decrease, as in cells without mAb R-G-4 treatment (virus and mAb 6-4-2, 1.6 × 103 ± 1.0 × 103 TCID50/mL; virus, mAb 6-4-2, and mAb R-G-4, 5.6 × 102 ± 3.6 × 102 TCID50/mL). Nearly identical results were obtained as in the case of FIPV 79-1146 (Fig. 2).

Involvement of virus receptors (fAPN) in ADE in feline monocytes. The results are shown as the mean ± SE (n = 10). Black bar, FIPV 79-1146, white bar, FECV 79-1683; N.D, not detected; N.S., not significant
Effects of lysosomotropic agents on ADE of FECV infection in primary feline monocytes
The ADE activity of FIPV is inhibited by the action of chloroquine and ammonium chloride because the endosomal pH rises and inhibits the fusion of FIPV with the endosome membrane in cells treated with chloroquine and ammonium chloride. Inhibition of FECV replication by treatment with chloroquine and ammonium chloride has been reported. However, the influence of chloroquine and ammonium chloride on ADE activity of FECV has not been reported. Thus, we investigated whether the ADE activity of FECV is inhibited by chloroquine and ammonium chloride. Primary feline monocytes were treated with chloroquine or ammonium chloride, and the influence on viral replication after FECV infection was investigated. The viability of primary feline monocytes after treatment with chloroquine and ammonium chloride was 98.7 ± 1.6 % (mean ± SD) and 101.2 ± 3.6 % (mean ± SD), respectively. The virus titer was significantly higher in the culture supernatant of monocytes inoculated with a mixture of FECV 79-1683 and mAb 6-4-2 than in monocytes cultured with the virus alone (virus alone, 1.8 × 102 ± 4.8 × 101 TCID50/mL; virus and mAb 6-4-2, 1.5 × 103 ± 4.9 × 102 TCID50/mL). However, treatment with 20 μM chloroquine or 5 mM ammonium chloride significantly decreased the virus titer in the culture supernatant of cells infected with virus alone or a mixture of virus and mAb (< 0 TCID50/mL, in all cases of treatment with lysosomotropic agents). Similar results were obtained with FIPV 79-1146 (Fig. 3).

Effects of lysosomotropic agents on ADE in feline monocytes. The results are shown as the mean ± SE (n = 10). Black bar, FIPV 79-1146; white bar, FECV 79-1683; N.D., not detected
Effect of ADE of FECV infection on inflammatory cytokine mRNA levels in primary feline monocytes
Inflammatory cytokine production is enhanced when primary feline monocytes are inoculated with FIPV in the presence of anti-FIPV S antibodies. However, it is not clear whether inflammatory cytokine production increases with ADE of FECV infection. Thus, we investigated the inflammatory cytokine mRNA levels in primary feline monocytes inoculated with a mixture of FECV 79-1683 and mAb 6-4-2 (Fig. 4). The TNF-α, IL-1β, and IL-6 mRNA levels were significantly increased in monocytes inoculated with a mixture of FIPV 79-1146 and mAb 6-4-2. However, these mRNA levels were not increased in monocytes inoculated with a mixture of FECV 79-1683 and mAb 6-4-2.

Inflammatory cytokine mRNA expression levels in ADE in feline monocytes. The results are shown as the mean ± SE (n = 10). Black bar, FIPV 79-1146; white bar, FECV 79-1683; N.S., not significant
Discussion
FECV infects primary feline monocytes and macrophages, but the infectivity of FECV for these cells is lower than that of FIPV [4, 24]. This difference has been suggested to be correlated with the pathogenicity of these viruses [24]. FECV is assumed to cause almost no clinical symptoms because it does not readily infect primary feline monocytes or macrophages and it replicates only in local regions (intestinal epithelium) [3]. In contrast, FIPV is considered to cause FIP because it readily infects primary feline monocytes and macrophages and circulates throughout the body. However, the conserved FCoV 7b gene is also detected in regions other than the intestine in some cases of FECV infection [13, 16], and the reason for this has not yet been elucidated. Monocyte and macrophage infection by FIPV is potentiated in the presence of antibodies. It was assumed that the efficiency of infection of monocytes and macrophages by FECV increases in the presence of antibodies, as is observed with FIPV. If this assumption is correct, the mechanism of systemic circulation of enterocyte-tropic FECV may be explained. Thus, we investigated whether FECV has ADE activity.
A human histiocytic lymphoma cell line, U937, was used to measure ADE of FIPV infection. As U937 cells do not express fAPN, FIPV alone does not infect U937 cells, but it enters U937 cells via the cell-surface Fc receptor when anti-FCoV S antibodies are present [12, 31]. Furthermore, as no feline monocyte/macrophage cell line has been established, U937 cells are useful as a model to predict ADE activity of FCoV. In this study, FECV exhibited ADE activity in U937 cells, but the activity level was lower than that of FIPV. FIPV exhibited ADE activity in the presence of mAb 6-4-2 at several concentrations, but FECV was only active at a tenfold dilution of mAb 6-4-2, i.e., the monocyte infection efficiency of FIPV was enhanced in the presence of the antibody, but the increase in the efficiency of FECV infection in the presence of the antibody was only slight. We also investigated the difference in the ADE activity of FIPV and FECV in primary feline monocytes. The experiment was performed at an antibody concentration maximizing ADE activity of FECV based on the experimental results with U937 cells. ADE activity in primary feline monocytes was confirmed for both FIPV and FECV, but, similar to the results of the experiment with U937 cells, the ADE activity level of FECV was lower than that of FIPV. Specifically, the mean ADE activity level of FECV was approximately 1/20 of that of FIPV. FIPV-infected primary feline monocytes and macrophages expressing ADE release excess cytokines, such as TNF-α and IL-6, as virus production increases [27, 28 30]. These cytokines cause a reduction in the number of lymphocytes and neutrophil activation, aggravating the FIP pathology. Therefore, high-level ADE activity is related to clinical aggravation to a serious state in FIPV-infected cats. Therefore, the low ADE activity of FECV may reflect the weak pathogenicity of FECV.
In primary feline monocytes, the ADE activity level of FECV was lower than that of FIPV. FIPV does not require the virus receptor to enter cells to induce ADE [31], but FECV is assumed to require both Fc and virus receptors; therefore, FECV enters cells through an inefficient mechanism of ADE compared with FIPV. To demonstrate the necessity of fAPN for ADE activity of FECV, we investigated whether anti-fAPN mAb inhibits ADE activity of FECV in primary feline monocytes. Contrary to our assumption, it was confirmed that FECV, like FIPV, does not use fAPN for ADE, suggesting that the difference in replicability in ADE between FIPV and FECV occurs after entering cells. The non-fAPN-mediated cell entry mechanism in ADE of type II FCoV infection is unclear. It has been reported that several viruses do not utilize their virus receptors when they enter cells using antibodies [26], but the mechanism has not been elucidated. It has been reported that viruses possessing an envelope, such as FCoV, can fuse their envelope with the cell membrane without mediation by the virus receptor, depending on the conditions [35]. For the non-fAPN-mediated cell entry mechanism of type II FCoV, further investigation is necessary.
Regan et al. [23] reported a study on the cell entry mechanism of FECV in which entry was dependent on the endosomal pH. However, the cell entry mechanism of FECV in ADE has not been clarified. We investigated its dependence on pH. In chloroquine- or ammonium-chloride-treated primary feline monocytes, FECV replication was strongly inhibited regardless of the presence or absence of mAb 6-4-2, suggesting that cell entry of FECV depends on the endosomal pH, regardless of the presence or absence of ADE.
ADE is associated with the severity of FIPV infection. There is a strong association between clinical symptoms of FIP and inflammatory cytokine levels released by primary feline monocytes and macrophages with ADE [27, 28, 30]. The TNF-α, IL-1β, and IL-6 mRNA levels significantly increased with ADE of FIPV infection, whereas no increase was noted in any of these despite ADE of FECV infection being observed. Several studies on the relationship between the amount of inflammatory cytokine production in virus-infected primary feline monocytes and macrophages and the severity of the disease have been reported. For example, in equine arteritis virus (EAV), virulent EAV significantly induces TNF-α production in macrophages compared with avirulent EAV, and a similar finding has been observed with equine infectious anemia virus [17]. Therefore, there may be a correlation between FCoV pathogenesis and the level of inflammatory cytokine production.
Only a few studies on ADE of serotype I FCoV infection have been reported. We previously reported that serotype I FIPV challenge hastened FIP development in cats carrying anti-serotype I FIPV antibodies [33]. In that study, ADE was observed not only in serotype II CoV infection but also serotype I FCoV infection. However, no in vitro study on the mechanism of ADE of serotype I FCoV infection has been performed because no mAb with ADE activity has been prepared against serotype I FCoV. Preparation of an anti-serotype I FCoV mAbs with ADE activity and similar experiments using it are anticipated.
We confirmed ADE in both FIPV and FECV infections. We also confirmed the difference in ADE between FECV and FIPV infections by basic analysis. The ADE activity of FECV was lower than that of FIPV, suggesting that ADE activity is involved in the difference in FCoV pathogenicity. It is necessary to closely analyze the ADE mechanism of FECV infection using cellular imaging and flow cytometry techniques. Furthermore, it is desirable to investigate whether there is a difference in ADE activity between FECV and FIPV similar to that observed in the present study in Fc-receptor-expressing cells (B cells and neutrophils) other than monocytes and macrophages. We hope that the present study helps further the understanding of ADE of FCoV infection and that it will help to elucidate ADE of serotype I FCoV infection.
References
Belouzard S, Millet JK, Licitra BN, Whittaker GR (2012) Mechanisms of coronavirus cell entry mediated by the viral spike protein. Viruses 4:1011–1033
Brown MA, Troyer JL, Pecon-Slattery J, Roelke ME, O’Brien SJ (2009) Genetics and pathogenesis of feline infectious peritonitis virus. Emerg Infect Dis 15:1445–1452
Desmarets LM, Theuns S, Olyslaegers DA, Dedeurwaerder A, Vermeulen BL, Roukaerts ID, Nauwynck HJ (2013) Establishment of feline intestinal epithelial cell cultures for the propagation and study of feline enteric coronaviruses. Vet Res 44:71
Dewerchin HL, Cornelissen E, Nauwynck HJ (2005) Replication of feline coronaviruses in peripheral blood monocytes. Arch Virol 150:2483–2500
Dye C, Siddell SG (2005) Genomic RNA sequence of Feline coronavirus strain FIPV WSU-79/1146. J Gen Virol 86:2249–2253
Dye C, Temperton N, Siddell SG (2007) Type I feline coronavirus spike glycoprotein fails to recognize aminopeptidase N as a functional receptor on feline cell lines. J Gen Virol 88:1753–1760
de Groot RJ, Baker SC, Baric R, Enjuanes L, Gorbalenya AE, Holmes KV, Perlman S, Poon L, Rottier PJM, Talbot PJ, Woo PCY, Ziebuhr J (2011) Family Coronaviridae. In: King AMQ, Adams MJ, Carstens EB, Lefkowitz EJ (eds) Virus taxonomy, ninth report of the International Committee on Taxonomy of Viruses. Elsevier Academic Press, San Diego, pp 806–828
Hohdatsu T, Izumiya Y, Yokoyama Y, Kida K, Koyama H (1998) Differences in virus receptor for type I and type II feline infectious peritonitis virus. Arch Virol 143:839–850
Hohdatsu T, Nakamura M, Ishizuka Y, Yamada H, Koyama H (1991) A study on the mechanism of antibody-dependent enhancement of feline infectious peritonitis virus infection in feline macrophages by monoclonal antibodies. Arch Virol 120:207–217
Hohdatsu T, Okada S, Ishizuka Y, Yamada H, Koyama H (1992) The prevalence of types I and II feline coronavirus infections in cats. J Vet Med Sci 54:557–562
Hohdatsu T, Sasamoto T, Okada S, Koyama H (1991) Antigenic analysis of feline coronaviruses with monoclonal antibodies (MAbs): preparation of MAbs which discriminate between FIPV strain 79-1146 and FECV strain 79-1683. Vet Microbiol 28:13–24
Hohdatsu T, Yamada M, Tominaga R, Makino K, Kida K, Koyama H (1998) Antibody-dependent enhancement of feline infectious peritonitis virus infection in feline alveolar macrophages and human monocyte cell line U937 by serum of cats experimentally or naturally infected with feline coronavirus. J Vet Med Sci 60:49–55
Kipar A, Meli ML, Baptiste KE, Bowker LJ, Lutz H (2010) Sites of feline coronavirus persistence in healthy cats. J Gen Virol 91:1698–1707
Kummrow M, Meli ML, Haessig M, Goenczi E, Poland A, Pedersen NC, Hofmann-Lehmann R, Lutz H (2005) Feline coronavirus serotypes 1 and 2: seroprevalence and association with disease in Switzerland. Clin Diagn Lab Immunol 12:1209–1215
Lin CN, Su BL, Wang CH, Hsieh MW, Chueh TJ, Chueh LL (2009) Genetic diversity and correlation with feline infectious peritonitis of feline coronavirus type I and II: a 5-year study in Taiwan. Vet Microbiol 136:233–239
Meli M, Kipar A, Müller C, Jenal K, Gönczi E, Borel N, Lutz H (2004) High viral loads despite absence of clinical and pathological findings in cats experimentally infected with feline coronavirus (FCoV) type I and in naturally FCoV-infected cats. J Feline Med Surg 6:69–81
Moore BD, Balasuriya UB, Watson JL, Bosio CM, MacKay RJ, MacLachlan NJ (2003) Virulent and avirulent strains of equine arteritis virus induce different quantities of TNF-α and other proinflammatory cytokines in alveolar and blood-derived equine macrophages. Virology 314:662–670
Motokawa K, Hohdatsu T, Hashimoto H, Koyama H (1996) Comparison of the amino acid sequence and phylogenetic analysis of the peplomer, integral membrane and nucleocapsid proteins of feline, canine and porcine coronaviruses. Microbiol Immunol 40:425–433
Olsen CW, Corapi WV, Ngichabe CK, Baines JD, Scott FW (1992) Monoclonal antibodies to the spike protein of feline infectious peritonitis virus mediate antibody-dependent enhancement of infection of feline macrophages. J Virol 66:956–965
Pedersen NC (2009) A review of feline infectious peritonitis virus infection: 1963–2008. J Feline Med Surg 11:225–258
Pedersen NC (2014) An update on feline infectious peritonitis: Virology and immunopathogenesis. Vet J 201:123–132
Reed LJ, Muench H (1938) A simple method of estimating fifty per cent endpoints. Am J Hyg 27:493–497
Regan AD, Shraybman R, Cohen RD, Whittaker GR (2008) Differential role for low pH and cathepsin-mediated cleavage of the viral spike protein during entry of serotype II feline coronaviruses. Vet Microbiol 132:235–248
Rottier PJM, Nakamura K, Schellen P, Volders H, Haijema BJ (2005) Acquisition of macrophage tropism during the pathogenesis of feline infectious peritonitis is determined by mutations in the feline coronavirus spike protein. J Virol 79:14122–14130
Shiba N, Maeda K, Kato H, Mochizuki M, Iwata H (2007) Differentiation of feline coronavirus type I and II infections by virus neutralization test. Vet Microbiol 124:348–352
Takada A, Kawaoka Y (2003) Antibody-dependent enhancement of viral infection: molecular mechanisms and in vivo implications. Rev Med Virol 13:387–398
Takano T, Azuma N, Hashida Y, Satoh R, Hohdatsu T (2009) B-cell activation in cats with feline infectious peritonitis (FIP) by FIP-virus-induced B-cell differentiation/survival factors. Arch Virol 154:27–35
Takano T, Azuma N, Satoh M, Toda A, Hashida Y, Satoh R, Hohdatsu T (2009) Neutrophil survival factors (TNF-alpha, GM-CSF, and G-CSF) produced by macrophages in cats infected with feline infectious peritonitis virus contribute to the pathogenesis of granulomatous lesions. Arch Virol 154:775–781
Takano T, Hohdatsu T, Hashida Y, Kaneko Y, Tanabe M, Koyama H (2007) A “possible” involvement of TNF-alpha in apoptosis induction in peripheral blood lymphocytes of cats with feline infectious peritonitis. Vet Microbiol 119:121–131
Takano T, Hohdatsu T, Toda A, Tanabe M, Koyama H (2007) TNF-alpha, produced by feline infectious peritonitis virus (FIPV)-infected macrophages, upregulates expression of type II FIPV receptor feline aminopeptidase N in feline macrophages. Virology 364:64–72
Takano T, Katada Y, Moritoh S, Ogasawara M, Satoh K, Satoh R, Hohdatsu T (2008) Analysis of the mechanism of antibody-dependent enhancement of feline infectious peritonitis virus infection: aminopeptidase N is not important and a process of acidification of the endosome is necessary. J Gen Virol 89:1025–1029
Takano T, Katoh Y, Doki T, Hohdatsu T (2013) Effect of chloroquine on feline infectious peritonitis virus infection in vitro and in vivo. Antivir Res 99:100–107
Takano T, Kawakami C, Yamada S, Satoh R, Hohdatsu T (2008) Antibody-dependent enhancement occurs upon re-infection with the identical serotype virus in feline infectious peritonitis virus infection. J Vet Med Sci 70:1315–1321
Vennema H, De Groot RJ, Harbour DA, Dalderup M, Gruffydd-Jones T, Horzinek MC, Spaan WJ (1990) Early death after feline infectious peritonitis virus challenge due to recombinant vaccinia virus immunization. J Virol 64:1407–1409
Watanabe R, Matsuyama S, Taguchi F (2006) Receptor-independent infection of murine coronavirus: analysis by spinoculation. J Virol 80:4901–4908
Acknowledgements
This work was supported by JSPS KAKENHI (Grant-in-Aid for Scientific Research (B)) Grant Number JP16H05039.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Ethical approval
All applicable international, national, and institutional guidelines for the care and use of animals were followed. The blood sampling in SPF cats was performed in accordance with the Guidelines for Animal Experiments of Kitasato University (approval number 16-086).
Rights and permissions
About this article

Cite this article
Takano, T., Nakaguchi, M., Doki, T. et al. Antibody-dependent enhancement of serotype II feline enteric coronavirus infection in primary feline monocytes. Arch Virol 162, 3339–3345 (2017). https://doi.org/10.1007/s00705-017-3489-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00705-017-3489-8