
Abstract
UV photolysis has been recommended as an alternative pretreatment method for the elimination of antibacterial activity of antibiotics against the indicator strain, but the pretreated antibiotic intermediates might not lose their potential to induce antibiotic resistance genes (ARGs) proliferation during subsequent biotreatment processes. The presence of florfenicol (FLO) in wastewater seriously inhibits the metabolic performance of anaerobic sludge microorganisms, especially the positive correlation between UV irradiation doses and ATP content, while it did not significantly affect the organics utilization ability and protein biosynthetic process of aerobic microorganisms. After sufficient UV pretreatment, the relative abundances of floR from genomic or plasmid DNA in subsequent aerobic and anaerobic biotreatment processes both decreased by two orders of magnitude, maintained at the level of the groups without FLO selective pressure. Meanwhile, the abundances of floR under anaerobic condition were always lower than that under aerobic condition, suggesting that anaerobic biotreatment systems might be more suitable for the effective control of target ARGs. The higher abundance of floR in plasmid DNA than in genome also indicated that the potential transmission risk of mobile ARGs should not be ignored. In addition, the relative abundance of intI1 was positively correlated with floR in its corresponding genomic or plasmid DNA (p < 0.05), which also increased the potential horizontal transfer risk of target ARGs. This study provides new insights into the effect of preferential UV photolysis as a pretreatment method for the enhancement of metabolic performance and source control of target ARGs in subsequent biotreatment processes.
Key points
• Sufficient UV photolytic pretreatment efficiently controlled the abundance of floR
• A synchronous decrease in abundance of intI1 reduced the risk of horizontal transfer
• An appreciable abundance of floR in plasmid DNA was a potential source of total ARGs
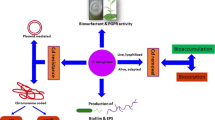
Graphical abstract

Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
As emerging contaminants, antibiotics have become a global threat to human health (Li et al. 2018; Van Boeckel et al. 2015), and their environmental risk mainly originates from the selective pressure of antibacterial activity on the surrounding environmental microorganisms (Boonsaner and Hawker 2013; Zhang et al. 2015; Zhou et al. 2013). Wastewater treatment plants (WWTPs), as the receiving terminals of wastewater originating from medical treatment, animal production and pharmaceutical industries, are the most important hotspot for the evolution and transmission of antibiotic resistance bacteria and genes (ARB and ARGs, respectively) (Deng et al. 2018; Luo et al. 2014; Oberoi et al. 2019). Therefore, effective reduction and control of the risk of antibiotic resistance in WWTPs is an important link in preventing the evolution of bacterial resistance.
The risk of antibiotic resistance is closely related to the specific chemical structure of antibiotics. Florfenicol (FLO) is currently the most widely used ingredient in animal and aquaculture production owning to its strong antibacterial effect and excellent biological safety (Zhang et al. 2015). However, due to its fatal disadvantages of the stable structure and difficult biodegradation, FLO seriously inhibits biological activity and aggravates the evolution, enrichment and dissemination of ARGs in sludge (Deng et al. 2012; Grandclement et al. 2017; Hao et al. 2019; Liang et al. 2019; Lu et al. 2014; Meng et al. 2019). Thus, it is necessary to preferentially eliminate the antibacterial activity of the target antibiotic using abiotic pretreatment technology. Advanced oxidation processes are presently among the most widely reported methods for antibiotic removal (Haider et al. 2019; Homem and Santos 2011; Kong et al. 2015; Yang et al. 2019). However, owing to the non-targeted functional mechanism of oxidative reactive species, the coexisting organic matters will competitively consume most of the reactive species, resulting in high treatment costs with a low removal efficiency of the target antibiotic (Benitez et al. 2011; Yi et al. 2016). In particular, the corresponding antibacterial activity cannot be effectively eliminated during the oxidative process. Therefore, the selective elimination of the antibacterial activity of target antibiotics in multicomponent wastewater has become an important research topic.
UV photolysis has been recommended as a new alternative method for wastewater treatment against a wide range of environmental pollutants, especially antibiotic contaminants (Ou et al. 2016; Song et al. 2016). Our previous study also showed that UV photolysis can efficiently eliminate the antibacterial activity of frequently consumed antibiotics, such as FLO, cefalexin, tetracycline, sulfadiazine and ofloxacin (Ding et al. 2020). However, the elimination of antibacterial activity only decreased its inhibitory effect on a single indicator strain, which did not mean that the pretreated antibiotic contaminants lost their potential to induce ARG proliferation in sludge microorganisms during the subsequent biotreatment. Therefore, additional efforts are needed to evaluate the abundance level of the ARGs associated with the target antibiotic and integron in the subsequent biotreatment with UV photolytic pretreatment. Meanwhile, how the abundance of corresponding ARGs varies with antibiotic selective pressure during the biotreatment process is also a key issue that needs to be determined and verified, which will be achieved by the corresponding control of UV irradiation dose during the pretreatment process. In addition, there is still a lack of comprehensive understanding of the correlation between the fate of ARGs and different subsequent wastewater biotreatment processes (Tong et al. 2019).
The present study aimed to investigate the effect of FLO selective pressure reduction by UV254 irradiation pretreatment on the enhancement of the metabolic performance of sludge microorganisms and the source control of characteristic ARGs in subsequent aerobic and anaerobic biotreatment processes. The specific objectives were to (i) demonstrate the effect of different UV254 irradiation dose pretreatments on the elimination performance of the antibacterial activity of FLO and metabolic activity of sludge microorganisms in aerobic and anaerobic bioreactors, (ii) investigate the effects of UV254 irradiation pretreatment on the fate of floR and intI1 and the alteration of sludge characteristics during the change of antibiotic selective pressure and (iii) reveal the linkage between variations in ARG abundance and the elimination of antibiotic selective pressure during the variation of UV irradiation doses in the pretreatment process. The results of this study provide a potential feasibility of UV photolysis as a pretreatment method for the effective treatment of antibiotic-containing wastewater and source control of corresponding ARGs in subsequently biological treatment processes.
Materials and methods
UV photolytic pretreatment combined with biotreatment experiment
UV photolytic pretreatment coupled with subsequent biological treatment process was the synthetic antibiotic-containing wastewater pretreated with different doses of UV irradiation and then used as the influent of aerobic or anaerobic bioreactors for subsequent biotreatment processes. The UV photolytic pretreatment experiment was conducted in a 200 mL cylindrical glass reactor, and the details of the reactor construction and operating instructions were provided in the Supporting Information (SI). FLO acted as the target antibiotic in the synthetic antibiotic-containing wastewater, with an initial concentration of 50 mg L−1 based on the upper limit level detected in the surrounding environments of its production or consumption industries (Periyasamy et al. 2022). During the pretreatment phase, FLO-containing wastewater was continuously irradiated by a low-pressure Hg UV lamp for 1, 2 and 3 h, which corresponded to doses of 20.16, 40.32 and 60.48 J cm−2, respectively. For the subsequent biotreatment phase, 100 mL of pretreated wastewater was transferred into a 200 mL brown glass bottle with 50 mL of aerobic or anaerobic sludge (MLSS = 10 g L−1) (Fig. S1b, c). Synthetic wastewater without FLO and FLO-containing wastewater without UV irradiation were considered as the influents of the negative and positive groups, respectively. Three parallel bottles were set up for each pretreatment wastewater sample (Table S1). After 8 weeks of operation, the influent of the part bioreactors was adjusted to verify the effect of variation in antibiotic selective pressure on ARG abundance, and the detailed information was shown in Table S1. The influent and effluent were sampled weekly, whereas the biomass samples were collected biweekly for subsequent measurements.
Water chemistry
Chemical oxygen demand (COD) was measured by a digestion instrument and UV-Vis spectrophotometer (HACH, DRB200 and DR6000, USA) according to the manufacturer’s instructions. Volatile fatty acids (VFAs) were measured by a high-performance liquid chromatograph (HPLC, DGU 20A3R, Shimadzu, Japan) fitted with an Aminex HPX-87H Ion Exclusion Column (7.8 × 300 mm, Bio-Rad, USA). The mobile phase was 0.27‰ (v/v) H2SO4, and the flow rate was 0.6 mL min−1. CH4 in biogas was collected in a gas bag and quantified by a gas chromatograph (Agilent 7892, USA) equipped with a thermal conductivity detector and a 1.5 m stainless-steel column. FLO was quantified by an HPLC fitted with a reversed-phase C18 analytical column (5 μm d, 4.6 × 250 mm, Waters, Ireland) maintained at 35 °C. The mobile phases consisted of methanol and water (50: 50, v/v) in 0.8 mL min−1, and the detection wavelength was 223 nm (Dong et al. 2023). The limit of detection was 0.1 mg L−1 for FLO quantification. The recovery rate for FLO was in the range 97.5 to 102.8%.
Microbial activity and protein analysis
The supernatant of the biotreatment bottle was collected for adenosine triphosphate (ATP) analysis, while the lower sludge was collected for protein analysis. ATP was measured using the BacTiter-GloTM Microbial Cell Viability Assay (G8231, Promega Corporation, Dübendorf, CH) and a GloMax 20/20 Luminometer (Turner BioSystems, Sunnyvale, CA, USA) following the optimized protocol described by Hammes et al (Hammes et al. 2010), which was detailed in the SI. A conversion factor of 1.75 × 10−10 nmol ATP per cell was used to calculate the bacterial cell number (Zhu et al. 2019). The collected sludge was used for protein extraction and measurement using the Bacterial Protein Extraction Kit and the Modified BCA Protein Assay Kit (Sangon Biotech Co., Ltd., Shanghai, China), according to the manufacturer’s instructions. All ATP and protein measurements were performed in triplicate, and the detection procedures were performed under sterile conditions.
DNA extraction and ARGs quantification by qPCR
For genomic and plasmid DNA extraction, each biomass sample was collected and separated into two parts before centrifugation for 5 min at 9 000× g. Genomic and plasmid DNA were then extracted from the precipitates using the FastDNA@ Spin Kit for Soil (MP Biomedicals, CA, USA) and TIANpure Mini Plasmid Kit (TIANGEN Biotech Co., Ltd., Beijing, China), respectively, following the manufacturer’s instructions. The DNA concentration was measured by a microspectrophotometer (Nanodrop 2000, Thermo Scientific, USA). All the extracted DNA was preserved at −20 °C for further analysis. Quantitative PCR (qPCR) was performed to determine the abundances of 16S rRNA, floR and intI1 genes using a QuantStudio 6 Flex System (Thermo, USA). A series of tenfold dilutions of plasmid standards ranging from 4.7 × 103 to 4.7 × 107 gene copies μL−1 were conducted to generate the target gene standard curves. Details on the qPCR measurements, including the primer sequences and standard curves, were presented in the SI (Table S2 and S3).
Statistical analysis
Statistical significance and Pearson’s correlation were determined at a 95% confidence level by SPSS 17.0. To determine the significance of changes in ARG abundance at different UV irradiation doses, a two-tailed unpaired Student’s t test was performed between all sampling data. Correlation analysis was performed between all ARGs, UV irradiation doses, biomass and antibiotic concentrations during the entire operational period. Prior to data analysis, the relative abundance of the target gene was determined by normalizing the target gene copies against its 16S rRNA amount.
Results
Biological treatment performance of FLO-containing wastewater pretreated with UV photolysis
The removal rates of FLO in synthetic wastewater were close to 100% in the 2 and 3 h UV pretreatment groups (Fig. S2). As for the 1 h UV group, the average residual FLO concentration was approximately 1.4 mg L−1, which was still well above the threshold concentration (0.3 mg L−1) for the elimination of FLO antibacterial activity reported in our previous study (Ding et al. 2020). Then, the unpretreated and pretreated wastewaters containing the residual FLO parent compound or its photoproducts were fed into the subsequently corresponding bioreactors as their influent. During the first operation phase, the influent concentration of FLO in adequate UV irradiation pretreatment groups (2 and 3 h UV) was below 0.1 mg L−1, while the effluent concentrations of FLO after aerobic and anaerobic biological process with an HRT of 7 days from the 1 h UV group were in the ranges of 0.5–1.7 mg L−1 and 0.1–1.3 mg L−1, respectively, which still provided an appropriate FLO selective pressure for the sludge bacteria in both aerobic and anaerobic bioreactors. Another group under a FLO selective pressure was the 0 h UV (without UV irradiation) group, whose FLO concentration decreased from 38.5–60.6 mg L−1 in the influent to 23.3–50.2 mg L−1 in the effluent (Fig. S3b, g). Meanwhile, the relatively low removal rate of FLO in a single biological process indicated its low biodegradability in both aerobic and anaerobic bacteria and further proved the technical necessity of FLO pretreatment by UV photolysis.
During the following validation phase, the influent of the original bioreactors was adjusted by extending or shortening the UV irradiation dose. Although low biological removal rates of FLO were also found in the bioreactors with unpretreated wastewater, a relatively higher removal rate was observed in these bioreactors acclimated by the relatively low FLO selective pressure during the prior phase, e.g. 1, 2 and 3 h UV groups. During the biotreatment of the wastewater containing 1.53 ± 0.30 mg L−1 residual FLO which was irradiated by UV light for 1 h, the FLO removal rates increased with the improvement of UV irradiation dose in the FLO-containing wastewater, which was regarded as the influent acclimating the original bioreactors. Indeed, a similar sequence of FLO removal rates was observed during the biotreatment of the influent irradiated by UV light for 2 h. Therefore, early domestication of low concentration FLO pollution was helpful in improving the degradability of subsequent treatments with higher concentrations of FLO. Moreover, an interesting phenomenon was that the removal rate of FLO under aerobic condition was higher than that under anaerobic condition with the same pretreated influent, which suggested that the aerobic biotreatment process was more conducive to FLO removal, but had a higher potential risk of ARG proliferation due to its rapid adaptation to the antibiotic selective pressure.
Because of the negligible COD removal in a single UV photolytic process, the organic matter in the synthetic wastewater was entirely consumed by sludge microorganisms. Thus, the synthetic wastewater after UV photolytic pretreatment and the bioreactor process were regarded as the influent and effluent for the biological treatment process, respectively. Except for the positive control (0 h UV), the COD removal rates in the aerobic bioreactors were generally higher than 95% during the first operation process (Phase I), resulting in an effluent COD of 10–20 mg L−1 (Fig. 1a, c, d, e), while the residual effluent COD in the group without UV pretreatment was mainly composed of the FLO parent compound (Fig. 1b). After changing the UV irradiation doses in the influent of the original bioreactors in Phase II (the next 8 weeks), the COD removal rates in aerobic bioreactors (CK–0 h, 1 h–0 h UV, 2 h–0 h UV and 3 h–0 h UV) with unpretreated wastewater dropped to 85–90%, which was the same as that of the 0 h–1 h UV group. Meanwhile, the COD removal rates in the other treatment groups continued to remain between 95 and 100%.

Performance of COD removal in aerobic (a, b, c, d, e) and anaerobic (f, g, h, i, j) bioreactors fed with pretreated influent with different UV irradiation doses. CK represented the wastewater sample without the antibiotic. 0 h, 1 h, 2 h and 3 h denoted the wastewater samples subjected to UV irradiation for 0, 1, 2 and 3 h, respectively. After 63 days of operation, the pretreatment process of the influent of the original three parallel bioreactors under each pretreatment condition was adjusted, which included two bioreactors fed with changed influent and one bioreactor fed with unchanged influent
In contrast, the COD removal rates in the anaerobic bioreactors were significantly lower than those in the aerobic bioreactors (p < 0.05) (Fig. 1f, g, h, i, j). In Phase I, apart from the negative control (100%), the COD removal rates in the anaerobic bioreactors were as low as 29–36%. In Phase II, after the 2# parallel bioreactor of other pretreatment groups was fed with the unpretreated wastewater, the COD removal rates generally decreased to approximately 10%, even lower than that of the bioreactors (UV 0 h), which were always fed with unpretreated FLO wastewater. The abnormally high COD removal performance in 0 h UV group might have resulted from the evolution of the higher tolerance capacity of microorganisms under long-term selective pressure from FLO. In addition, the COD removal rates of other bioreactors fed with FLO wastewater pretreated with 1 or 2 h of UV irradiation did not change significantly in Phase II. For the bioreactors fed with wastewater pretreated with 3 h of UV irradiation, the COD removal rates in the CK–3 h and 3 h UV groups were more than 90%, which might be due to the adaptation of sludge microorganisms to selective pressure from low concentrations of FLO. Moreover, the higher residual COD in the anaerobic bioreactors was mainly composed of acetic acid and propionic acid, while isovaleric acid was also detected in the bioreactors fed with unpretreated FLO wastewater, and its concentration was remarkably higher than that of the other two VFAs (data not shown). However, the presence of a high concentration of FLO would inhibit the activity of methanogens in sludge (Zarei-Baygi et al. 2019), and the average volume of biogas and CH4 production increased steadily with increasing UV irradiation doses received by the influent of the anaerobic bioreactors in Phase I (p ˂ 0.05, Fig. S4). Meanwhile, the proportion of CH4 in the biogas was also positively correlated with the UV irradiation dose received by the influent of the anaerobic bioreactors (Table S4). This indicated that the inhibitory effect of FLO on methanogenic activity in the anaerobic digestion system had a concentration effect. A similar phenomenon was also observed in the biogas production of different treatment groups in Phase II. Obviously, the cumulative biogas production of the bioreactors with the wastewater without UV pretreatment as their influent decreased significantly, especially the volume of biogas production in CK–0 h was less than a third of that in the other CK group (Fig. S4). The above results further verified that the preferential reduction of FLO antibacterial activity by UV photolytic pretreatment contributed to the improvement of methanogenic and metabolic performance of sludge microorganisms in subsequent anaerobic biotreatment systems.
Metabolism and activity of sludge microorganisms fed with the wastewater after UV photolytic pretreatment
FLO is a highly potent inhibitor of bacterial protein biosynthesis, and its antibacterial mechanism is mainly to interfere with the normal function of peptidyltransferase by reversible binding to the 50S ribosomal subunit of the 70S ribosome in prokaryotes, thus blocking the elongation of new peptide chains and inhibiting the synthesis of proteins (Schwarz et al. 2004). Therefore, the protein biosynthetic process of sludge microorganisms might be inhibited accordingly under severe environmental selective pressure from high concentration FLO. Meanwhile, since all physiological and metabolic activities of microorganisms are based on the participation of enzymes in physical and chemical processes, the environmental stress of FLO might also affect the biological activity of sludge microorganisms. To study the effect of UV photolytic pretreatment to eliminate the antibacterial activity of FLO on the growth and activity of sludge microorganisms in subsequent biological processes, the sludge samples were collected after 8 weeks of operation under different UV treatment doses, and the contents of ATP and protein were detected.
In the aerobic bioreactor, the ATP content of the blank control group without the FLO selective pressure was 2.39 ± 0.35 × 10−14 cell L−1, while the ATP contents of the experimental groups fed with FLO wastewater were 0.42–3.07 × 10−13 cell L−1, accounting for only 1.8–12.8% of the blank control group (Fig. 2a). This indicated that the presence of FLO indeed inhibited the activity of sludge microorganisms in the biological system, causing the number of active microorganisms to decline by more than one order of magnitude. Meanwhile, it was worth noting that the treatment group with the lowest ATP content was not the 0 h UV group fed with the highest FLO concentration but rather the 3 h UV group with the highest UV photolytic pretreatment dose (4.21 ± 1.63 × 10−12 cell L−1). The above results showed that the simple elimination of FLO antibacterial activity by photolytic pretreatment was not sufficient to completely minimize its inhibitory effect on microbial metabolism in the sludge system. Therefore, it was also necessary to consider the inhibition or destruction of microorganisms by photolytic intermediates of coexisting substances in the wastewater and some oxidized reactive species (•OH, 1O2, 3DOM*) produced by long-term UV photolysis (Zhang et al. 2019b). In the anaerobic system, the ATP content of the blank control group was as high as 2.64 ± 0.06 × 10−14 cell L−1, which was 2 times higher than that in the 0 h UV treatment group. In addition, the ATP content in the other pretreatment groups increased with an increase in the UV irradiation dose (r = 0.969, p < 0.05). The ATP content from the 3 h UV treatment group was as high as 2.82 ± 0.06 × 10−14 cell L−1, returning to the initial level similar to that of the blank control group, which indicated that an adequate dose of UV photolytic pretreatment could effectively reduce the inhibitory effect of FLO on anaerobic microorganisms and restore the ATP activity to normal metabolic level. The above results showed that the ATP content in the anaerobic system was significantly higher than that in the aerobic system during the same period, which further indicated that FLO was more inclined to inhibit the activity of aerobic microorganisms than that of anaerobic microorganisms.

ATP (a) and protein (b) contents in aerobic and anaerobic bioreactors fed with pretreated influent with different UV irradiation doses during Phase I
Owing to the potential role of FLO in inhibiting protein biosynthesis, the proteins of the sludge microorganisms were extracted and detected. In the aerobic bioreactors, except for 0 h UV treatment group, there was no significant difference in the protein content of the sludge between the blank control group and the other three treatment groups. In particular, the protein content of the 0 h UV treatment group was as high as 594.11 ± 27.38 mg g−1 VSS−1, which was 1.2 times than that of the corresponding blank control group. Based on the above hypothesis, it seems that FLO did not inhibit the protein biosynthetic process of aerobic sludge microorganisms. However, the protein content of sludge in the anaerobic bioreactors decreased with an increase in the UV pretreatment dose (r = 0.955, p < 0.05), which was not consistent with the hypothesis that FLO would have an inhibitory effect on protein biosynthesis by microorganisms. Because the sludge biological system is a complex ecological community with many kinds of microorganisms, the microbial species under aerobic and anaerobic conditions will evolve quite differently under the selective pressure from the presence of FLO (Tao et al. 2012). In fact, FLO could not inhibit the protein biosynthetic process of every microorganism, resulting in the protein contents in the aerobic and anaerobic bioreactors having significantly different trends among the different UV pretreatment groups. In other words, the presence of FLO played exactly an opposite directional screening role in changing the abundance of the respective sensitive microflora under aerobic and anaerobic conditions. Therefore, the corresponding sensitive strains selected as the model microorganisms could be used to conduct a verification experiment of FLO selective pressure inhibition on the protein biosynthetic process of microorganisms in a pure culture environment. In conclusion, the selective pressure of FLO significantly decreased the ATP content of microorganisms in the sludge biological system, and the impact on aerobic microorganisms was more remarkable than that on anaerobic microorganisms, while the correlation between the increased dose of UV photolysis and the change in ATP content was not as remarkable as that in the anaerobic sludge system. Meanwhile, the significant negative correlation between the protein content and UV irradiation dose under anaerobic condition further confirmed the targeted inhibitory effect of the FLO selective pressure on the protein biosynthetic process of anaerobic microorganisms.
Source control of relative abundance of floR in bioreactors after UV photolytic pretreatment
To further confirm the variation in the relative abundance of key ARGs under different UV pretreatment doses, we performed qPCR analysis on the selected ARGs in sludge samples from the two different operation phases. floR, an important representative of the chloramphenicol resistance gene located in both plasmid and genomic DNA, is sensitive to environmental pressure fluctuations from FLO and encodes a drug efflux pump (Braibant et al. 2005; Tao et al. 2012). After 18 weeks of operation, several significant differences were observed in the relative abundance of floR among the bioreactors fed with pretreated influent after different UV irradiation doses (Fig. 3). In the genome, the highest relative abundances of floR in the positive control (0 h UV, i.e. without UV pretreatment) in the aerobic and anaerobic bioreactors occurred at the end of Phase I, which were as high as 4.22 ± 0.34 × 10−1 copies/16S rRNA copy and 2.16 ± 0.12 × 10−1 copies/16S rRNA copy (Fig. 3a, b), and they were 311 and 263 times higher than that of the negative control (CK, i.e. wastewater without antibiotics), respectively. Although the relative abundances of floR in the 1 h UV pretreatment groups were remarkably decreased, especially under aerobic condition, accounting for only 4.0–56.9% of those in the positive controls, the abundance values were still 4–50 times higher than those of the negative controls. When the pretreatment doses for the elimination of FLO antibacterial activity were increased, the relative abundance levels of floR in the 2 and 3 h UV pretreatment groups were close to that of the CK group (10−3 copies/16S rRNA gene copy), which indicated that 2 h or longer UV irradiation during the pretreatment process could lead to two orders of magnitude decrease in the relative abundance of floR. The above results showed that UV photolytic pretreatment could effectively control the proliferation of target ARG by eliminating the selective pressure from the corresponding antibiotic in both aerobic and anaerobic bioreactors.

Relative abundance of floR located in genome (a, b) and plasmid (c, d) in aerobic (a, c) and anaerobic (b, d) bioreactors fed with pretreated influent after different UV irradiation doses. The relative abundances of floR were determined using qPCR by normalizing their copies against copies of the 16S rRNA gene (log 10 transformed)
After 4 weeks of operation in Phase II, the 2# parallel bioreactors of different treatment groups were changed to feed with the unpretreated wastewater as the influent, and the relative abundances of floR in these bioreactors increased significantly (p < 0.05), reaching 10−1 copies/16S rRNA copy. In particular, the relative abundance of floR in the CK–0 h and UV 3 h–0 h groups under anaerobic condition were increased to 1.55 ± 0.48 × 10−1 copies/16S rRNA copy and 1.49 ± 0.18 × 10−1 copies/16S rRNA copy, respectively, which were significantly higher than those in the aerobic bioreactors under the same influent conditions during the same period (p < 0.05). For the bioreactors acclimating to the high concentration FLO selective pressure (UV 0 h) in Phase I, although the FLO selection pressure was eliminated by increasing the UV irradiation dose in Phase II, the relative abundance of floR still could not be reduced to the level of the negative control or 2 and 3 h UV treatment groups, and was stable between 0.45 and 1.07 × 10−1 copies/16S rRNA copy, which was slightly lower than the abundance level of floR in the other bioreactors fed with the wastewater without UV irradiation. The above results for Phase II indicated that reducing the selective pressure from the influent of FLO-containing wastewater through UV photolytic pretreatment in the bioreactor containing a relatively high abundance level of ARGs was unhelpful for the elimination of ARGs abundance and transmission risk. This further verified the importance and necessity of UV photolysis pretreatment to preferentially eliminate the antibacterial activity of antibiotic wastewater for the subsequent biological treatment system to source control the ARG pollution risk. In addition, the relative abundances of floR in the different bioreactors under anaerobic condition were always lower than that under aerobic condition during the two operation stages, which also suggested that anaerobic biotreatment systems might be more suitable for the treatment of antibiotic wastewater than aerobic biotreatment systems, in terms of the effective control of the ARG transmission risk (Ding et al. 2021).
Similar to the distribution in genomic DNA, the relative abundance of floR in plasmid DNA in both aerobic and anaerobic bioreactors decreased significantly with an increase in the UV irradiation dose (p < 0.05). In particular, the relative abundance of floR in the 0 h UV treatment group under aerobic condition was as high as 5.96 ± 0.25 × 10−1 copies/16S rRNA copy, which was higher than that in the anaerobic bioreactors (p < 0.05, Fig. 3c, d). This result was consistent with the distribution of floR abundance in genomic DNA, which might have been caused by the more vigorous metabolism and efficient intergenerational proliferation of ARB in aerobic activated sludge, thus leading to the enhanced amplification of ARGs and plasmids carried by them (Hou et al. 2019). In addition, the relative abundance of floR in plasmid DNA in the aerobic or anaerobic bioreactors was generally higher than that in genomic DNA under the same condition. This also indicated that there were abundant ARGs in plasmid DNA, which were frequently overlooked. Therefore, it is essential to develop an effective pretreatment method for potential extracellular ARGs to control the potential risk of ARG proliferation and transmission (Chang et al. 2017; Zhang et al. 2018). After 8 weeks of operation in Phase II, the relative abundance of floR in the aerobic bioreactor dropped from the highest level of nearly 6.00 × 10−1 copies/16S rRNA copy to 3.00 × 10−1 copies/16S rRNA copy, which was at the same abundance level as that under anaerobic condition. In addition, the relative abundance of floR in 2# parallel bioreactors of different treatment groups significantly increased after the treatment with high concentrations of FLO wastewater (p < 0.05), reaching the abundance level of genomic DNA under the same conditions (10−1 copies/16S rRNA copy). Meanwhile, the relative abundances of floR in plasmid DNA under anaerobic conditions were 2.06–2.69 × 10−1 copies/16S rRNA copy, which were higher than that in the aerobic reactor at the same time (p < 0.05). For both the aerobic and anaerobic bioreactors, the relative abundances of floR in genomic and plasmid DNA were positively correlated with their corresponding FLO concentrations; that is, sufficient UV irradiation pretreatment could effectively control the relative abundances of floR in genomic and plasmid DNA during the subsequent biotreatment process, which could be maintained at a similar abundance level as the negative control group. In conclusion, the effective control of floR abundance in genomic and plasmid DNA further verified the wide applicability of UV photolytic pretreatment to antibiotic-containing wastewater.
Source control of relative abundance of intI1 in bioreactors after UV photolytic pretreatment
Integrons have been investigated in wastewater treatment systems because of their potential role in the horizontal transfer of ARGs among bacterial species and in the evolution of multidrug resistance in environmental pathogenic bacteria (An et al. 2022; Gao et al. 2017). In particular, class 1 integrons, which contain the intI1 gene, have been frequently detected at a relatively high abundance compared to other types of integrons in wastewater treatment systems (Garner et al. 2018; Quintela-Baluja et al. 2021; Ren et al. 2018). For the genomic DNA from the aerobic sludge in Phase I, the relative abundances of intI1 in 1, 2 or 3 h UV pretreatment groups were close to the abundance level of its corresponding negative control group (mostly below 1.0 × 10−1 copies/16S rRNA copy). However, the abundances of intI1 in the anaerobic bioreactors under the same experimental condition increased to more than 1.0 × 10−1 copies/16S rRNA copy, which were significantly higher than that in aerobic bioreactors (p < 0.05), as well as higher than that in the negative control group under anaerobic condition (Fig. 4a, b). In contrast, the relative abundance of intI1 in the positive control group increased to 6.75 ± 0.19 × 10−1 and 5.12 ± 0.33 × 10−1 copies/16S rRNA copy under aerobic and anaerobic conditions, respectively, which were 7.1 and 6.4 times higher than that of the negative control group after 8 weeks of biological treatment. Different from the distribution of floR under the same conditions, the relative abundances of intI1 showed no significant difference among the different pretreatment groups, except for the 0 h UV experimental group, which also indicated that only 1 h of UV irradiation could effectively control the proliferation of integrons. In addition, the distribution and relative abundance of intI1 derived from aerobic microbial plasmid DNA in the above five treatment groups were consistent with that of genomic DNA, with the highest abundance reaching 7.10 ± 0.21 × 10−1 copies/16S rRNA copy, which was 11 times higher than that in the negative control group under the same influent condition during the same period (Fig. 4c, d). However, under anaerobic condition, the selective pressure from the high concentration FLO did not actually induce a significant increase in the abundance of intI1 in plasmid DNA, and the highest abundance was only 2.63 ± 0.20 × 10−1 copies/16S rRNA copy, which was 3 times higher than that of the corresponding negative control group, while, the relative abundances of intI1 in the other treatment groups were maintained at the same level as that of the negative control group. The above results indicated that the pretreatment of FLO wastewater with 1 h UV irradiation dose could effectively limit the induction performance for the proliferation of intI1 in biological processes, especially under aerobic condition, and its reduction effect was more significant than that under anaerobic condition. Moreover, the abundance of intI1 in plasmid DNA under aerobic condition was at the same level as that in genomic DNA, which was a non-negligible source of intI1.

Relative abundance of intI1 located in genome (a, b) and plasmid (c, d) in aerobic (a, c) and anaerobic (b, d) bioreactors fed with pretreated influent after different UV irradiation doses. The relative abundances of intI1 were determined using qPCR by normalizing their copies against copies of the 16S rRNA gene (log 10 transformed)
In Phase II, the relative abundances of intI1 in genomic DNA from other treatment groups fed with unpretreated wastewater also increased significantly; in particular, the abundance of the 2# parallel bioreactor in the negative control group was as high as 6.14 ± 0.33 × 10−1 copies/16S rRNA copy, which was the same level as the abundance of the 0 h UV group (Fig. 4a). Meanwhile, the relative abundance of intI1 in plasmid DNA from the 0 h UV treatment group increased steadily with operation time, and the highest abundance reached 1.42 ± 0.36 copies/16S rRNA copy, which was nearly twice that in genomic DNA under the same condition. In addition, the relative abundances of intI1 in the 0 h−1 h UV and 0 h–2 h UV aerobic bioreactors actually increased to 1.00 copies/16S rRNA copy after increasing the UV pretreatment dose in the influent to eliminate the FLO selective pressure (Fig. 4a). This result indicated that the simple elimination of environmental antibiotic selective pressure was not sufficient to control or reduce the abundance of intI1 in the aerobic bioreactors which already had contained high abundance levels of the corresponding ARGs. For the other treatment groups that changed the influent with unpretreated wastewater in Phase II, the relative abundances of intI1 also increased significantly, which were higher than that in genomic DNA under the same condition. In addition, the relative abundances of intI1 in the anaerobic bioreactors during the second phase were positively related to the FLO selective pressure on its surrounding environment; that is, the abundance of intI1 in the bioreactors fed with unpretreated wastewater as the influent was generally maintained at a higher level. For plasmid DNA under the same condition, the high abundance of intI1 was mainly concentrated in the treatment groups with high concentration FLO selective pressure or those that experienced high FLO selective pressure in Phase II. In conclusion, the higher abundance of intI1 in plasmid DNA during aerobic biological treatment might enhance the potential for horizontal transfer of ARGs through more frequent interactions of genetic materials with ARB (Hu et al. 2017). Compared with the unpretreated groups under high FLO selective pressure, the relative abundances of intI1 in the pretreatment groups were significantly decreased under aerobic or anaerobic conditions, which indicated that UV pretreatment could not only effectively reduce the abundance of corresponding ARGs, but also effectively control the abundance level of intI1, providing long-term stability at the same level as the blank control group. It was worth noting that, compared with the abundance of floR, the initial abundance of intI1 in the biological system was relatively high, which indicated that the role of UV pretreatment in reducing environmental selective pressure seems to be particularly important.
Discussion
To explore the possible dose–response relationship between the UV irradiation dose and representative ARGs and integrons, the correlation between the metabolic activity or the abundance levels of target ARGs and the UV irradiation dose was analyzed in both aerobic and anaerobic biotreatment processes after 8 weeks of operation (Phase I). Firstly, the relative abundances of floR in genomic DNA from the aerobic bioreactors without UV pretreatment were always higher than that under anaerobic condition, while the abundances of floR decreased remarkably with increasing UV pretreatment dose in both the aerobic and anaerobic bioreactors (r = −0.746, p < 0.01 and r = −0.852, p < 0.01, Table 1), resulting in the same abundance level. For the same distribution as floR in genomic DNA, its relative abundance in plasmid DNA also had a significantly negative correlation with the pretreatment dose of UV irradiation (r = −0.692, p < 0.01 and r = −0.778, p < 0.01, Table 1). The above significant correlations indicated that the presence of environmental selective pressure from FLO would exponentially increase the abundance of corresponding characteristic ARGs (floR) in wastewater treatment systems, whereas sufficient UV pretreatment would successfully reduce the relative abundance of floR in both genomic and plasmid DNA in subsequent aerobic and anaerobic biotreatment processes. In addition, as a major class of integrons, the relative abundance of intI1 in genomic or plasmid DNA was also negatively correlated with its UV pretreatment dose in the antibiotic-containing wastewater (r > 0.6, p < 0.01, Table 1). This suggested that the preferential UV pretreatment could effectively control the abundance of intI1 in the subsequent biotreatment systems, which would reduce the horizontal transfer risk of corresponding ARGs (Shin et al. 2022; Zheng et al. 2020). Meanwhile, the relative abundance of intI1 was positively correlated with its corresponding floR in both genomic and plasmid DNA (r > 0.9, p < 0.05, Table S5), which might exacerbate the potential risk of the horizontal transfer of floR. In addition, the high abundance of intI1 might also enhance the horizontal transfer process of other types of ARGs among bacteria by capturing and recombining exogenous gene fragments, which could aggravate the environmental pollution problem of ARGs (Ren et al. 2018; Zhang et al. 2019a). The above results further verified that UV photolytic pretreatment played an important role in source controlling the abundance level of intI1 in subsequent biological treatment systems by eliminating the FLO environmental selection pressure. Furthermore, the reduction in the FLO selective pressure after a significant proliferation of intI1 could not promptly reduce its corresponding abundance level, which had already remained relatively high level in the earlier phase. This result emphasized the urgency and necessity of reasonable and effective UV photolytic pretreatment to eliminate antibiotic antibacterial activity before the subsequent biological treatment of antibiotic wastewater.
This study provided comprehensive insights into the impact of UV photolysis as a pretreatment method on the enhancement of metabolic performance and the source control of corresponding ARGs and integrons in subsequent aerobic and anaerobic biotreatment processes fed with pretreated FLO containing wastewater. The presence of FLO in wastewater inhibited the activity of microorganisms in anaerobic sludge. In contrast, the environmental pressure of FLO did not significantly affect the utilization of organic matters by aerobic microorganisms, including the relatively high removal rate of COD (95%) for FLO-containing wastewater and the no remarkable difference in the protein biosynthetic process among the different doses of UV pretreatment. A sufficient dose of UV irradiation pretreatment could effectively reduce the relative abundance of the representative FLO resistance gene (floR) and the frequently occurred integron (intI1) in both genomic and plasmid DNA in the subsequent aerobic and anaerobic biotreatment processes, which were closed to those without antibiotic contamination. Meanwhile, the relative abundance of floR under anaerobic condition was generally lower than that under aerobic condition, and the abundance of floR in plasmid DNA was higher than that in genomic DNA under the same conditions during the two operational stages. In addition, the significantly positive correlation between the relative abundance of floR and its corresponding intI1 in genomic or plasmid DNA increased the potential risk of horizontal transfer of target ARGs. Although there are methodological limitations of extracting and analyzing genomic and plasmid DNA separately, the distribution trend of the target ARGs among the different pretreatment groups was not affected theoretically. Specifically, the abundances of floR and intI1 in UV pretreatment groups were significantly lower than that of the positive control, both in the genomic and plasmid DNA. In order to more accurately obtain the distribution and abundance of target genes on different genetic elements, molecular biological methods, such as southern hybridization, are needed to determine the genetic location of target ARGs or the correlation between floR and intI1. These results demonstrated that UV photolysis could be adopted as an efficient and promising pretreatment method for the enhancement of biotreatment systems and source control of characteristic ARGs in subsequent aerobic and anaerobic biotreatment processes.
Data availability
All data generated or analyzed during this study are included in this published article and its supplementary information files.
References
An XL, Abass OK, Zhao CX, Xu MR, Pan T, Pu Q, Liao H, Li H, Zhu YG, Su JQ (2022) Nanopore sequencing analysis of integron gene cassettes in sewages and soils. Sci Total Environ 817:152766. https://doi.org/10.1016/j.scitotenv.2021.152766
Benitez FJ, Acero JL, Real FJ, Roldan G, Casas F (2011) Comparison of different chemical oxidation treatments for the removal of selected pharmaceuticals in water matrices. Chem Eng J 168(3):1149–1156. https://doi.org/10.1016/j.cej.2011.02.001
Boonsaner M, Hawker DW (2013) Evaluation of food chain transfer of the antibiotic oxytetracycline and human risk assessment. Chemosphere 93(6):1009–1014. https://doi.org/10.1016/j.chemosphere.2013.05.070
Braibant M, Chevalier J, Chaslus-Dancla E, Pages JM, Cloeckaert A (2005) Structural and functional study of the phenicol-specific efflux pump FloR belonging to the major facilitator superfamily. Antimicrob Agents Chemother 49(7):2965–2971. https://doi.org/10.1128/AAC.49.7.2965-2971.2005
Chang PH, Juhrend B, Olson TM, Marrs CF, Wigginton KR (2017) Degradation of extracellular antibiotic resistance genes with UV254 treatment. Environ Sci Technol 51(11):6185–6192. https://doi.org/10.1021/acs.est.7b01120
Deng Y, Li B, Zhang T (2018) Bacteria that make a meal of sulfonamide antibiotics: blind spots and emerging opportunities. Environ Sci Technol 52(7):3854–3868. https://doi.org/10.1021/acs.est.7b06026
Deng Y, Zhang Y, Gao Y, Li D, Liu R, Liu M, Zhang H, Hu B, Yu T, Yang M (2012) Microbial community compositional analysis for series reactors treating high level antibiotic wastewater. Environ Sci Technol 46(2):795–801. https://doi.org/10.1021/es2025998
Ding Y, Jiang W, Liang B, Han J, Cheng H, Haider MR, Wang H, Liu W, Liu S, Wang A (2020) UV photolysis as an efficient pretreatment method for antibiotics decomposition and their antibacterial activity elimination. J Hazard Mater 392:122321. https://doi.org/10.1016/j.jhazmat.2020.122321
Ding Y, Liang B, Jiang W, Han J, Guadie A, Yun H, Cheng H, Yang R, Liu SJ, Wang A, Ren N (2021) Effect of preferential UV photolysis on the source control of antibiotic resistome during subsequent biological treatment systems. J Hazard Mater 414:125484. https://doi.org/10.1016/j.jhazmat.2021.125484
Dong S, Ding Y, Feng H, Xu J, Han J, Jiang W, Xia Y, Wang A (2023) Source preventing mechanism of florfenicol resistance risk in water by VUV/UV/sulfite advanced reduction pretreatment. Water Res 235:119876. https://doi.org/10.1016/j.watres.2023.119876
Gao P, Gu C, Wei X, Li X, Chen H, Jia H, Liu Z, Xue G, Ma C (2017) The role of zero valent iron on the fate of tetracycline resistance genes and class 1 integrons during thermophilic anaerobic co-digestion of waste sludge and kitchen waste. Water Res 111:92–99. https://doi.org/10.1016/j.watres.2016.12.047
Garner E, Chen C, Xia K, Bowers J, Engelthaler DM, McLain J, Edwards MA, Pruden A (2018) Metagenomic characterization of antibiotic resistance genes in full-scale reclaimed water distribution systems and corresponding potable systems. Environ Sci Technol 52(11):6113–6125. https://doi.org/10.1021/acs.est.7b05419
Grandclement C, Seyssiecq I, Piram A, Wong-Wah-Chung P, Vanot G, Tiliacos N, Roche N, Doumenq P (2017) From the conventional biological wastewater treatment to hybrid processes, the evaluation of organic micropollutant removal: A review. Water Res 111:297–317. https://doi.org/10.1016/j.watres.2017.01.005
Haider MR, Jiang WL, Han JL, Sharif HMA, Ding YC, Cheng HY, Wang AJ (2019) In-situ electrode fabrication from polyaniline derived N-doped carbon nanofibers for metal-free electro-Fenton degradation of organic contaminants. Appl Catal B-Environ 256. https://doi.org/10.1016/j.apcatb.2019.117774
Hammes F, Goldschmidt F, Vital M, Wang Y, Egli T (2010) Measurement and interpretation of microbial adenosine tri-phosphate (ATP) in aquatic environments. Water Res 44(13):3915–3923. https://doi.org/10.1016/j.watres.2010.04.015
Hao L, Okano K, Zhang C, Zhang Z, Lei Z, Feng C, Utsumi M, Ihara I, Maseda H, Shimizu K (2019) Effects of levofloxacin exposure on sequencing batch reactor (SBR) behavior and microbial community changes. Sci Total Environ 672:227–238. https://doi.org/10.1016/j.scitotenv.2019.03.272
Homem V, Santos L (2011) Degradation and removal methods of antibiotics from aqueous matrices—a review. J Environ Manage 92(10):2304–2347. https://doi.org/10.1016/j.jenvman.2011.05.023
Hou J, Chen Z, Gao J, Xie Y, Li L, Qin S, Wang Q, Mao D, Luo Y (2019) Simultaneous removal of antibiotics and antibiotic resistance genes from pharmaceutical wastewater using the combinations of up-flow anaerobic sludge bed, anoxic-oxic tank, and advanced oxidation technologies. Water Res 159:511–520. https://doi.org/10.1016/j.watres.2019.05.034
Hu HW, Wang JT, Li J, Shi XZ, Ma YB, Chen D, He JZ (2017) Long-term nickel contamination increases the occurrence of antibiotic resistance genes in agricultural soils. Environ Sci Technol 51(2):790–800. https://doi.org/10.1021/acs.est.6b03383
Kong D, Liang B, Yun H, Cheng H, Ma J, Cui M, Wang A, Ren N (2015) Cathodic degradation of antibiotics: characterization and pathway analysis. Water Res 72:281–292. https://doi.org/10.1016/j.watres.2015.01.025
Li S, Shi W, Liu W, Li H, Zhang W, Hu J, Ke Y, Sun W, Ni J (2018) A duodecennial national synthesis of antibiotics in China’s major rivers and seas (2005-2016). Sci Total Environ 615:906–917. https://doi.org/10.1016/j.scitotenv.2017.09.328
Liang B, Ma J, Cai W, Li Z, Liu W, Qi M, Zhao Y, Ma X, Deng Y, Wang A, Zhou J (2019) Response of chloramphenicol-reducing biocathode resistome to continuous electrical stimulation. Water Res 148:398–406. https://doi.org/10.1016/j.watres.2018.10.073
Lu X, Zhen G, Liu Y, Hojo T, Estrada AL, Li YY (2014) Long-term effect of the antibiotic cefalexin on methane production during waste activated sludge anaerobic digestion. Bioresour Technol 169:644–651. https://doi.org/10.1016/j.biortech.2014.07.056
Luo Y, Guo W, Ngo HH, Nghiem LD, Hai FI, Zhang J, Liang S, Wang XC (2014) A review on the occurrence of micropollutants in the aquatic environment and their fate and removal during wastewater treatment. Sci Total Environ 473-474:619–641. https://doi.org/10.1016/j.scitotenv.2013.12.065
Meng Y, Sheng B, Meng F (2019) Changes in nitrogen removal and microbiota of anammox biofilm reactors under tetracycline stress at environmentally and industrially relevant concentrations. Sci Total Environ 668:379–388. https://doi.org/10.1016/j.scitotenv.2019.02.389
Oberoi AS, Jia Y, Zhang H, Khanal SK, Lu H (2019) Insights into the fate and removal of antibiotics in engineered biological treatment systems: a critical review. Environ Sci Technol 53(13):7234–7264. https://doi.org/10.1021/acs.est.9b01131
Ou HS, Ye JS, Ma S, Wei CH, Gao NY, He JZ (2016) Degradation of ciprofloxacin by UV and UV/H2O2 via multiple-wavelength ultraviolet light-emitting diodes: effectiveness, intermediates and antibacterial activity. Chem Eng J 289:391–401. https://doi.org/10.1016/j.cej.2016.01.006
Periyasamy S, Lin X, Ganiyu SO, Kamaraj SK, Thiam A, Liu D (2022) Insight into BDD electrochemical oxidation of florfenicol in water: kinetics, reaction mechanism, and toxicity. Chemosphere 288(Pt 1):132433. https://doi.org/10.1016/j.chemosphere.2021.132433
Quintela-Baluja M, Frigon D, Abouelnaga M, Jobling K, Romalde JL, Gomez Lopez M, Graham DW (2021) Dynamics of integron structures across a wastewater network—implications to resistance gene transfer. Water Res 206:117720. https://doi.org/10.1016/j.watres.2021.117720
Ren S, Boo C, Guo N, Wang S, Elimelech M, Wang Y (2018) Photocatalytic reactive ultrafiltration membrane for removal of antibiotic resistant bacteria and antibiotic resistance genes from wastewater effluent. Environ Sci Technol 52(15):8666–8673. https://doi.org/10.1021/acs.est.8b01888
Schwarz S, Kehrenberg C, Doublet B, Cloeckaert A (2004) Molecular basis of bacterial resistance to chloramphenicol and florfenicol. FEMS Microbiol Rev 28(5):519–542. https://doi.org/10.1016/j.femsre.2004.04.001
Shin J, Choi S, Park CM, Wang J, Kim YM (2022) Reduction of antibiotic resistome in influent of a wastewater treatment plant (WWTP) via a chemically enhanced primary treatment (CEPT) process. Chemosphere 286(Pt 1):131569. https://doi.org/10.1016/j.chemosphere.2021.131569
Song K, Mohseni M, Taghipour F (2016) Application of ultraviolet light-emitting diodes (UV-LEDs) for water disinfection: a review. Water Res 94:341–349. https://doi.org/10.1016/j.watres.2016.03.003
Tao W, Lee MH, Wu J, Kim NH, Kim JC, Chung E, Hwang EC, Lee SW (2012) Inactivation of chloramphenicol and florfenicol by a novel chloramphenicol hydrolase. Appl Environ Microbiol 78(17):6295–6301. https://doi.org/10.1128/AEM.01154-12
Tong J, Tang A, Wang H, Liu X, Huang Z, Wang Z, Zhang J, Wei Y, Su Y, Zhang Y (2019) Microbial community evolution and fate of antibiotic resistance genes along six different full-scale municipal wastewater treatment processes. Bioresour Technol 272:489–500. https://doi.org/10.1016/j.biortech.2018.10.079
Van Boeckel TP, Brower C, Gilbert M, Grenfell BT, Levin SA, Robinson TP, Teillant A, Laxminarayan R (2015) Global trends in antimicrobial use in food animals. Proc Natl Acad Sci U S A 112(18):5649–5654. https://doi.org/10.1073/pnas.1503141112
Yang L, Chen Z, Cui D, Luo X, Liang B, Yang L, Liu T, Wang A, Luo S (2019) Ultrafine palladium nanoparticles supported on 3D self-supported Ni foam for cathodic dechlorination of florfenicol. Chem Eng J 359:894–901. https://doi.org/10.1016/j.cej.2018.11.099
Yi Q, Gao Y, Zhang H, Zhang H, Zhang Y, Yang M (2016) Establishment of a pretreatment method for tetracycline production wastewater using enhanced hydrolysis. Chem Eng J 300:139–145. https://doi.org/10.1016/j.cej.2016.04.120
Zarei-Baygi A, Harb M, Wang P, Stadler LB, Smith AL (2019) Evaluating antibiotic resistance gene correlations with antibiotic exposure conditions in anaerobic membrane bioreactors. Environ Sci Technol 53(7):3599–3609. https://doi.org/10.1021/acs.est.9b00798
Zhang MQ, Yuan L, Li ZH, Zhang HC, Sheng GP (2019a) Tetracycline exposure shifted microbial communities and enriched antibiotic resistance genes in the aerobic granular sludge. Environ Int 130:104902. https://doi.org/10.1016/j.envint.2019.06.012
Zhang QQ, Ying GG, Pan CG, Liu YS, Zhao JL (2015) Comprehensive evaluation of antibiotics emission and fate in the river basins of China: source analysis, multimedia modeling, and linkage to bacterial resistance. Environ Sci Technol 49(11):6772–6782. https://doi.org/10.1021/acs.est.5b00729
Zhang X, Li J, Fan WY, Yao MC, Yuan L, Sheng GP (2019b) Enhanced photodegradation of extracellular antibiotic resistance genes by dissolved organic matter photosensitization. Environ Sci Technol 53(18):10732–10740. https://doi.org/10.1021/acs.est.9b03096
Zhang Y, Li A, Dai T, Li F, Xie H, Chen L, Wen D (2018) Cell-free DNA: a neglected source for antibiotic resistance genes spreading from WWTPs. Environ Sci Technol 52(1):248–257
Zheng W, Huyan J, Tian Z, Zhang Y, Wen X (2020) Clinical class 1 integron-integrase gene—a promising indicator to monitor the abundance and elimination of antibiotic resistance genes in an urban wastewater treatment plant. Environ Int 135:105372. https://doi.org/10.1016/j.envint.2019.105372
Zhou LJ, Ying GG, Liu S, Zhang RQ, Lai HJ, Chen ZF, Pan CG (2013) Excretion masses and environmental occurrence of antibiotics in typical swine and dairy cattle farms in China. Sci Total Environ 444:183–195. https://doi.org/10.1016/j.scitotenv.2012.11.087
Zhu TT, Cheng HY, Yang LH, Su SG, Wang HC, Wang SS, Wang AJ (2019) Coupled sulfur and iron(II) carbonate-driven autotrophic denitrification for significantly enhanced nitrate removal. Environ Sci Technol 53(3):1545–1554. https://doi.org/10.1021/acs.est.8b06865
Funding
This work was supported by the China Postdoctoral Science Foundation (No. 2022M722809), the Zhejiang Provincial Postdoctoral Research Program (ZJ2022070), the Zhejiang Provincial Ten Thousand Plan (No. 2021R52055) and the Key Research and Development Project of Shandong Province (No. 2020CXGC011202).
Author information
Authors and Affiliations
Contributions
YC D, HJ F and JL H conceived, designed research and wrote the manuscript. YC D conducted the experiments. WL J and SJ D analyzed data. HY C, MZ W and AJ W reviewed and edited the manuscript. All authors read and approved the manuscript.
Corresponding author
Ethics declarations
Ethical approval
This article does not contain any studies with human participants or animals performed by any of the authors.
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
ESM 1
(PDF 395 kb)
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Ding, Y., Feng, H., Han, J. et al. Effect of UV pretreatment on the source control of floR during subsequent biotreatment of florfenicol wastewater. Appl Microbiol Biotechnol 108, 120 (2024). https://doi.org/10.1007/s00253-023-12826-4
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00253-023-12826-4