
Abstract
Seedling size-related functional attributes of Mediterranean forest species are critical traits to be considered in nursery practices aimed at improving the survival and performance of seedlings exposed to summer drought in dryland reforestation projects. We looked at how nursery light regimes and nitrogen fertilization affected the survival and performance of Pinus pinaster Ait. under contrasting post-planting watering regimes. Seedlings grown in the nursery for 13 months were given low or high nitrogen doses and exposed to three light regimes (full sun, medium light and low light). After outplanting, half of the seedlings were watered fortnightly during the hottest months of the first summer and the other half were left unwatered. Seedling size-related variables (shoot height, basal diameter, stem volume and sturdiness quotient) were measured at the beginning and at the end of the study, and summer drought survival was checked three times. Our results showed that fertilization and the nursery light regime influenced initial seedling quality and transplant stress. Post-planting first summer watering also increased seedling survival probability. The medium-light and especially the full-sunlight regimes in the nursery decreased the probability of seedling mortality after outplanting. However, a low-light regime in the nursery negatively affected plant traits and increased the probability of post-planting seedling mortality. In comparison with low-N seedlings, high-N fertilization increased post-planting stress (evaluated in the change of sturdiness quotient after one field growth season), but did not affect survival.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Forest and landscape restoration seeks to recover ecological functionality and ecosystem services, which play an important economic role and maximize human well-being worldwide (Lavorel 2013; César et al. 2021). In places with seasonal drought, successful forest restoration by planting seedlings to artificially regenerate managed stands (hereafter plantations) is heavily dependent on early cultivation practices (Trubat et al. 2010; Shi et al. 2019) and how seedling attributes develop in imposed nursery conditions (Villar-Salvador et al. 2004, 2013; Trubat et al. 2010, 2011; Cortina et al. 2013; Grossnickle and McDonald 2018a). Successful forest restoration requires seedlings with optimal growth potential (Grossnickle and McDonalds 2018b). Restoration success can be improved by producing in-nursery phenotypes with morphological drought-avoidance traits through controlled light and fertilization regimes (Oliet et al. 2009; Trubat et al. 2010; Rodríguez-García and Bravo 2013; Merine et al. 2015; Kildisheva et al. 2017; Shi et al. 2019). Examples of such traits include high root collar diameter, low shoot height and sturdiness quotient and large stem and root volume (Ovalle et al. 2016), which are also key attributes in post-planting response (Oliet et al. 2009). For example, maximum plant height is related to light acquisition, competitive vigour, reproductive investment and dispersal distance (Ostertag et al. 2015). Seedling stature or relative position in the forest can also influence species interactions, growth, and survival (Rodríguez-García et al. 2011a, b) at different trophic levels (Lavorel 2013; Ostertag et al. 2015; Andivia et al. 2021).
Knowledge of how nursery conditions affect post-planting treatments to promote seedling survival and performance in dry continental Mediterranean forests it is still scarce and more research is needed. Some studies indicate that nursery-cultured N-rich seedlings grow larger and better, with higher survival and greater competitive ability in the field after outplanting (Puértolas et al. 2003; Villar-Salvador et al. 2004, 2012, 2013; Fernández et al. 2007; Oliet et al. 2009, 2013; Cuesta et al. 2010a, b). However, the impact of nursery culturing techniques on the subsequent performance of outplanted seedlings could be site-specific (Luis et al. 2009; Cortina et al. 2013; Ovalle et al. 2016; Oliet et al. 2021) during the early establishment stage. Post-planting first-year summer irrigation generally has a positive effect on seedling performance and survival (Ruano et al. 2009, 2015; Pardos et al. 2015). Relevant research also emphasizes the importance of irradiance aspects such as photoperiod, PFD (photon flux density) and the amount of incident light per day used to cultivate tree species in the nursery (Bazzaz and Wayne 1994; Luo et al. 2021). For example, seedlings grown in shade have a higher sturdiness quotient, greater imbalance in the shoot/root ratio (Luis et al. 2009) and suffer photoinhibition when transferred to high light conditions (Puértolas et al. 2009). However, if seedlings can be grown with shade in the nursery without impairing their quality, a great amount of water would be saved (see Puértolas et al. 2009).
Maritime pine (Pinus pinaster Ait) is a Mediterranean forest species of great ecological and economic importance. It has been planted around the world because it provides a wide array of ecosystem services (Ruano et al. 2015). In this study, we examined Maritime pine seedling stock characteristics to determine how nursery light, fertilization and post-planting first-year summer irrigation affected seedling survival and performance in the field. We hypothesized that (1) using nutrient-rich soil would produce larger seedlings, but differences in field survival would depend on nursery light conditions; (2) there would be differences in survival and growth among full-sunlight, medium-light and low-light nutrient-rich seedlings; (3) field survival and growth would be higher in watered post-planting seedlings.
Material and methods
Seedling production and experimental design in the nursery
Maritime pine seedlings from the Cuéllar provenance (41° 10′ N–4° 30′ W) in Segovia (Castile-Leon, central Spain) were used in this study (Alía et al. 1996, 2009). The seedlings were grown from commercial seeds (harvest year 2003) received in early January 2006 from the Centro Nacional de Mejora Forestal (INIA, National Centre for Forest Improvement, Madrid, Spain) in a nursery at the University of Valladolid in Palencia (central Spain, 42° 01′ N–4° 32′ W, 739 m.a.s.l.) under continental Mediterranean climate conditions (annual precipitation and average temperature of 450 mm and 12 °C, respectively). They were cultivated along with seedlings from nine other provenances, which were used for a study about Maritime pine seedling plasticity (Rodríguez-García and Bravo 2013).
First, the seeds were stratified for 6 weeks (from 17 January 2006 to 28 February 2006) in aluminium trays with moist sterile sand (Fig. S1A) in a cold room at 5 °C (www.miteco.gob.es). After stratification, the sand was removed by washing the seedlings with abundant sterile water. Then, the seed coats were sterilized by immersing the seeds for 10 min in a 20% solution of Domestos (a commercial soapy bleach composed of sodium hypochlorite with 4.3% active chlorine solution along with non-ionic and cationic surfactants, soap and perfume). They were washed again, kept in sterile water for 48 h, then sown in 12 cm diameter Petri dishes (20 seeds/dish) on two layers of sterile filter paper saturated with demineralized water. The dishes were placed in a germination chamber (Fig. S1B) on 3 March 2006 in a randomized design under controlled photoperiod (14 h light/10 h dark), temperature (21 °C day/17 °C night) and relative humidity (60% day and night) conditions. The seeds were examined every three days and water was added to keep the filter paper saturated. After two weeks, all the seeds needed for the trial had germinated (the radicle could be seen, Fig. S1C).
Following germination and initial radicle growth (1–3 cm long), the seedlings were transferred to Arnabat® 48 cavity forest trays (308 cm3, Fig. S1D and S1E). from 17 March to 1 April 2006. The seedlings planted in the 24 central cavities of the trays were used for the experiment while seedlings planted in the border cavities were excluded (as in Rodríguez-García and Bravo 2013). Each cavity was 18 cm2 with vertical, anti-spiralling ribs along the inside walls. The trays were filled with a 3:1 (v:v) peat-vermiculite mixture, to which 3.5 kg/m3 of Plantacote® NPK (14-8-15) slow release fertilizer (SRF) was added following commercial dose indications for normal nutrition of conifer cultures remaining longer than 6 months in trays. The range suggested by the commercial brand varied between 3.0 and 7 kg/m3. The formula has a stated equivalent nutrient release period of 8–9 months at 21 °C (as in Rodríguez-García and Bravo 2013).
The forest trays were kept inside glazed, periodically ventilated greenhouse installations for two months to prevent frost damage (which is quite common in Palencia), then taken out to a plot of land next to the greenhouse in early June 2006 (Fig. S1F). In a three-block, split-plot experimental design, a light gradient consisting of two extremes and one intermediate level was randomly established per block (Fig. S2). The light plots were oriented north–south in three 15 m long parallel lines or replicate blocks. The distance between the blocks was 2 m. Each block had three main plots: a high-light (HL) plot exposed to full sun irradiance with an average photosynthetic photon flux density (PPFD) of 337.1 µmol m2 s−1, a medium-light plot (ML, average PPFD of 120.6 µmol m2 s1), and a low-light plot (LL, average PPFD of 30.0 µmol m2 s−1). The ML and LL plots were established by constructing shade houses consisting of wire tents (2.60 m2 × 1.90 m high) covered with one (ML) or two (LL) layers of black raffia mesh. The shade houses reduced sun irradiance by 64% and 91% for ML and LL, respectively. PPFD was measured in each light plot with cross-calibrated radiation sensors (Li-190SA PAR Quantum Sensors; Li-COR Biosciences, USA) connected to a data logger that measured conditions every 10 min and gathered the data every hour. PPFD measurements were taken over an entire 24 h period on 1 or 2 sunny days in June, July, August and September, 2006 (Rodríguez-García and Bravo 2013).
Two forest trays from the Cuéllar provenance were placed in each light plot. Then, a two-level nutrient treatment (low N and high N) was implemented in which half the trays in each light plot were randomly assigned and administered a low dose of nitrogen, while the other half received a higher dose. A total of 432 Cuéllar seedlings were used (24 usable seedlings per tray × 3-level light treatment × 2-level nutrient treatment × 3 blocks). The amount of SRF added to the substrate when the trays were filled was established as the low-N control treatment. It corresponds to standard use of slow-release fertilizer for conifer cultures with medium nutrient needs that would remain longer than 6 months in trays. On average, each cavity received 222 mg N, 127 mg P and 238 mg K. The high-N (2× the low-N dose) treatment was obtained by adding N to the control substrate in a concentrated NH4NO3 solution (32%;1.3 g/mL) that was divided into 18 equal weekly doses (early June to mid-November 2006) and administered to the other half of the trays. Thus, the total N administered was 222 mg per low-N seedling and 444 mg per high-N seedling. Seedlings were kept well-watered, and the ML, LL and full-sunlight plots had similar average relative air humidity (around 69%) and air temperature (15 °C) throughout the experiment (see Rodríguez-García and Bravo 2013 for a detailed description).
Outplanting and field site conditions
In May 2007, we randomly selected 120 seedlings (20 seedlings per combination of nursery light x fertilization; 7 plants per combination were selected from two of the blocks and 6 from the third one) and transplanted them in an experimental field site located in Cuéllar (41° 22′ N, 4° 29′ W; 749 m.a.s.l.). Previous analyses (Rodríguez-García and Bravo 2013) indicated no significant effects of the block on plant growth in the nursery, so this factor was not included in our study. The site was a flat, natural P. pinaster stand with sandy, acidic, Entisol soil (Rodríguez-García et al. 2010) located in a clear-cut area (harvested in 2004) without regeneration at the time of plantation. The continental Mediterranean climate features drought in summer (June to September), frost in winter, and rainfall mainly in spring and late autumn. Mean annual precipitation is 610 mm, and mean annual temperature is 11.2 °C (Ruano et al. 2015; data from Sistema de Clasificación Bioclimática Mundial (1996–2009)). The floristic community is mainly composed of grassland species with small patches of low shrub species and isolated Pinus pinea trees scattered around the site.
The plantation surface covered 0.48 ha (240 m × 20 m). Seedlings were planted manually by making planting holes (0.20 m wide × 0.45 m deep) distributed 2 m × 2 m apart (a density of 2500 seedlings/ha) to prevent interspecific competition for soil water (Ovalle et al. 2016). Ten rows of 12 seedlings each were planted at the experimental site on 1 May 2007 (Fig. S3). The Pinus pinaster plantation density in our study is typical for stands in Castile-Leon, where reforestation planting density does not fall below 2000 plants/ha. The main objective of this approach is to dominate existing shrubs as much as possible or quickly cover completely bare soils that need protection (Serrada et al. 2008).
Post-planting first summer irrigation and water availability
Watering treatments began in June 2007 and ended in mid-September 2007. The experimental unit and sampling unit were each individual seedling. Half of the experimental units (60 seedlings; three levels of nursery light x two levels of nursery nitrogen × 10 replicates) were randomly assigned the summer watering (SW) treatment while the other half (60 seedlings) received no summer watering (NSW). The homogeneous conditions of the study site (sandy soil, flat terrain, not stony, same herbaceous vegetation and no tree canopy) allowed us to assign the treatments without blocking. SW seedlings were watered seven times, with 2 l of water every two weeks (Ruano et al. 2009) during the period mentioned. Each watering event was equivalent to a rainfall of 8 l/m2 and each seedling received a total of 56 l/m2. The total amount of rainfall in the period from 1 June to 15 September 2007 was 104 mm (Fig. 1). So, the cumulative summer water availability was 160 mm for the SW seedlings (irrigation plus rainfall), and 104 mm for the NSW seedlings (rainfall only). Climate data was obtained from the nearest meteorological station (SG01-Gomezserracín; Segovia), which is located 20 km from the experimental site and managed by the Instituto Tecnológico Agrario de Castilla y León (Agricultural Technological Institute of Castile-Leon) (www.inforiego.org).

(Source: Instituto Tecnológico Agrario de Castilla y León, Gómezserracín station, Segovia, Spain)
Monthly mean, minimum and maximum air temperature, and total precipitation at the trial site during the 2007–2008 growing season
Data collection and variables assessed
Seedling stock (morphological) attributes
At the time of transplant (2007), seedling root collar diameter (hereafter basal diameter; d, cm) and shoot height (h, cm) were measured. We calculated the sturdiness quotient (h/d) and stem volume (v) using the formula for a cone \((\frac{\pi {d}^{2}}{12}x h\), cm3).
Planting and watering response: growth and survival
One growing season after plantation, that is, 8 months after the first-summer watering treatment, and before the plants faced their second summer in the field, the seedlings were measured for shoot height and groundline stem diameter a second time and evaluated for short-term mortality (January 2008). The sturdiness quotient, stem volume and relative growth (RG) in height, basal diameter and stem volume were calculated as the height, diameter and stem volume increment observed in one growing season, using the formula RG = (X1−X0)/(X0), where X1 and X0 are the final and initial variables, respectively. The variables analysed (RG, X1 and X0) were subjected to a natural logarithm transformation to make relationships linear (Lenssen et al. 2003). After this second measurement, we checked the seedlings for medium-term mortality after three years (June 2010) and long-term mortality after ten years (February 2017) to determine plantation success.
Statistical analysis
First, we carried out a descriptive analysis and verified compliance with normality, homogeneity of variance, and linearity assumptions for all seedling size-related variables using Shapiro–Wilk (1965) and Levene (1960) tests. Prior to analysis, all variables were transformed to their natural logarithms because they did not meet normality and homoscedasticity assumptions. We examined the linear relationships between all variables for nursery and field experiments by means of Pearson correlation coefficients (r). Relationships were considered significant when P < 0.05. Coefficients r ≤ 0.35 were considered to represent low or weak correlations; those r from 0.36 to 0.67 represented modest or moderate correlations, those r from 0.68 to 1.0 indicated strong or high correlations and r coefficients ≥ 0.90 denoted very high correlations (Taylor 1990).
Second, differences in seedling morphological attributes before planting and one growing season after outplanting were assessed using General Linear Models with a split-plot design (Eq. 1).
where, \({\tau }_{i}\), \({\beta }_{j}\) and \({\left(\tau \beta \right)}_{ij}\) represent the whole plot and \({\gamma }_{k}\), \({\left(\tau \gamma \right)}_{ik},{ \left(\beta \gamma \right)}_{jk}\) and \({\left(\tau \beta \gamma \right)}_{ijk}\) represent the split plot. Here, \({\tau }_{i}\), \({\beta }_{j}\) and \({\gamma }_{k}\), are block effect, factor A (light) and factor B (fertilization) effect, respectively. The sums of squares for the factors are computed as in the three-way analysis of variance without replication. In the first case, we tested the light and fertilization effects. The main effects of light were tested against the block x light interaction term due to the split-plot nursery design, but block showed no significant effects. In the second General Linear Model, we tested the effects of light, fertilization and watering on morphology.
Planting stress occurs when seedlings are not fully coupled into the hydrologic cycle (Grossnickle and Ivetic 2022). The Transplant Stress Index, TSI, (South and Zwolinski 1997) can be used to estimate the intensity and length of planting stress (or transplant shock due to adjustment to a new environment). With time, the seedling establishes new roots and the direct effect of transplanting is gradually reduced. At some point in time, the seedling adjusts fully and subsequent growth is the same as that of a younger, non-transplanted seedling of equivalent size (South and Zwolinski 1997). TSI is defined as the slope of the linear relationship between the study variable at the beginning of the growth period and the increment of that variable. Negative or zero slopes indicate that seedlings have suffered a certain amount of post-planting stress.
In this study, the slope (regression coefficient) of the relationships between height, basal diameter, sturdiness quotient and stem volume at planting and those same variables after the first growth period (1 year) in the field—the Transplant Stress Index (TSI)—were checked with an analysis of covariance (ANCOVA). Height growth and basal diameter growth were used as dependent variables, the effect of light, nitrogen and post-planting watering as categorical factors (levels of treatment), and initial height and initial basal diameter as the independent variables (covariates) in the analyses. As block effect was negligible compared with treatments’ effects, it was not considered in the analyses. The aim was to separate the effect of seedling height and basal diameter at the beginning of the growth period from the effects of light, nitrogen and post-planting levels of treatment. As the levels of treatments (sets) were much related to initial height and basal diameter, ANCOVA adjustment removed the partial set effect due to initial height and basal diameter from the overall effects. Reciprocally, this allowed us to determine the relationships between initial height and basal diameter and growth in these variables by adjusting for the set effect. The common regression coefficient obtained gives a TSI value for each nursery light, nursery fertilization and post-planting watering level. This procedure helps to correct the effects of combining different sets when calculating TSI. This TSI value is called adjusted TSI in the text (TSIadj) (see Puertolas et al. 2003). The model (Eq. 2) was:
where \({Y}_{ij}=\) growth of seedling j from set i, \(\mu \) = growth overall mean, \({\tau }_{i}\) = effect of set I on growth, \(\beta \) = common regression coefficient (TSIadj), \({x}_{ij}=\) initial height/basal diameter mean of set i, \({X}_{..}\)= initial height/basal diameter overall mean, \({\varepsilon }_{ij}=\) error term.
Finally, we examined the relationship between post-planting survival probability, nursery culturing effects, field irrigation treatment effects and seedling stock (morphological) attributes (h, d, h/t and v) using Generalized Linear Models (GLZ; McCullagh and Nelder 1989). The ‘survival of first summer conditions’ response variable (Eq. 3) was fitted to a binomial distribution with the Logit link function (logistic regression, Eq. 4). The response variable can take only one of two possible values, denoted by 0 and 1. Therefore:
Eq. (3) express the probabilities of ‘failure’ and ‘success’, respectively, while the logistic regression model can be written symbolically as Eq. 4,
where \(\alpha , \beta , \gamma \) refer to categorical factors and \(\delta \) refer to continuous predictors. Categorical factors included light environment, N fertilization and post-planting watering. Seedling stock attributes were considered as continuous independent predictors. Two models were tested; one included seedling stock attributes as continuous independent predictors and the other did not. The final model was chosen based on its goodness of fit with Akaike information criterion (AIC, Johnson and Omland 2004) and statistical stability, as evaluated by ratios of deviance to respective degrees of freedom. After assessing the role of nursery culturing practices and watering in first-summer survival, we evaluated the persistence of significant effects on short- and long-term survival using GLZ models, following the same procedures. All analyses were carried out using Statistica 6.0 and SPSS 15.0 software.
Results
Effects of nursery fertilization and light on seedling stock attributes prior to field planting
At the end of the nursery cultivation period, 13 months after sowing, we saw that seedling morphological attributes differed significantly according to light and fertilization treatments (Table 1). The clearest effects were observed for low-light, high-N conditions and for medium-light conditions (Table 2). In general, greater shade was correlated with increased shoot height and sturdiness quotient but with decreased basal diameter and stem volume. Meanwhile, the effect of fertilization on seedling nursery attributes varied according to the light regime in the nursery (Tables 1 and 2). Fertilization in high light did not alter seedling shoot height (mean height of 30 cm, with a 95% confidence interval [CI] of 27.4–32.2 cm) and medium-light conditions (mean height of 38 cm; CI of 35.4–40.6 cm). In contrast, high-N fertilization significantly reduced seedling shoot height by 22% (30 cm) compared to both low-N, low-light seedlings, and medium-light seedlings (Table 2). The shoot height of the nutrient-rich low-light seedlings was similar (without significant differences) to that of seedlings grown in full sunlight (Table 2).
Basal diameter did not vary between high-N and low-N seedlings (Table 1 and Table 2) that received full sunlight and medium light (0.45 cm and 0.42 cm, respectively). In low-light conditions, however, basal diameter in high-N seedlings decreased by 15% (0.29 cm) compared to low-N seedlings (0.34 cm). High-N fertilization reduced the sturdiness quotient, but it was only significant for seedlings grown in medium-light conditions (Table 2). In contrast, seedling stem volume increased (although P > 0.05) with high-N fertilization in full-sunlight and medium-light conditions (by around 0.3 and 0.4 cm3, respectively). Despite this, stem volume was significantly smaller—around 42% (0.7 cm3) smaller—in seedlings grown in high-N, low-light conditions.
Total shoot height and the groundline basal diameter had a significant but low correlation (r = 0.20; P = 0.026), while the correlation of total shoot height with the sturdiness quotient (r = 0.57; P = 0.000) and stem volume (r = 0.53; P = 0.000) was moderately significant. A moderate negative correlation between basal diameter and the sturdiness quotient (r =−0.66; P = 0.000) was observed, along with a positive and strong correlation between basal diameter and stem volume (r = 0.90; P = 0.000). We also observed a weak correlation between high sturdiness quotient and low stem volume (r =−0.32; P = 0.000).
Evaluation of seedling functional traits one year after planting
One growth season after outplanting (May 2008), we observed a significant effect of post-planting first-summer watering on shoot height only (Table 3 and Fig. S4). However, light availability in the nursery significantly affected all seedling morphological attributes. Shoot height and the sturdiness quotient increased with shade, while basal diameter and stem volume followed the reverse pattern. Shoot height in seedlings grown in medium and low light was 15% higher than those grown in full sunlight, but no significant differences were observed between ML and HL, and between ML and LL (Fig. 2a). Consistently, the sturdiness quotient was 35% and 37% higher (index of 57 and 58, respectively) among seedlings grown in medium and low nursery light environments, when compared to full-sunlight seedlings (index of 42) (Fig. 2b). A moderate reduction of 12% (Fig. 3a) in the basal diameter of medium-light seedlings (0.69 cm) was observed, along with a sharper reduction of 24% for seedlings grown in low-light nursery conditions (0.59 cm), compared to seedlings that received full sunlight (0.78 cm). Seedling stem volume (Fig. 3b) was also appreciably lower in seedlings grown under shade in the nursery. Medium light decreased stem volume by around 7% (4.89 cm3) with respect to full-sunlight seedlings (5.25 cm3), but no significant differences were observed. Meanwhile, stem volume was significantly lower, (39% or 3.19 cm3) among seedlings grown in low-light versus full-sunlight nursery conditions. The basal diameter was significantly increased (7%) with high-N fertilization compared to low-N seedlings. The seedling sturdiness quotient (49) was significantly reduced (11%) with high-N fertilization (Table 3 and Fig. S5) compared to low-N seedlings (55). This was due to a 16% increase in basal diameter (average of 0.71 cm) in the absence of any notable effects of high-N fertilization on shoot height.

Effects of nursery light on Pinus pinaster a seedling shoot height (h1, cm) and b sturdiness quotient (h1/d1) one field-growth season after outplanting in a dry area. Letters indicate significant differences (P < 0.05)

Effects of nursery light on Pinus pinaster a seedling basal diameter (d1, cm) and b stem volume (v1, cm3) one field-growth season after outplanting in a dry area. Letters indicate significant differences (P < 0.05)
One year after planting in the field, significant correlations were still observed among all morphological attributes except shoot height and basal diameter. The sturdiness quotient correlated positively and highly with shoot height (r = 0.65; P = 0.000) and negatively with basal diameter (r =−0.63; P = 0.000). Meanwhile, stem volume correlated significantly and very highly with basal diameter (r = 0.89; P = 0.000), but negatively and weakly with the sturdiness quotient (r =−0.25; P = 0.016).
ANCOVA analyses and the regression coefficients of the linear regression models for pre-transplant size and post-transplant annual relative growth indicated transplant stress (Tables 4 and 5). The initial height and a low-light regime in the nursery significantly and negatively (negative ß) affected annual relative growth in height. The initial basal diameter at planting also negatively affected annual relative growth in basal diameter. However, a full-sunlight regime at the nursery positively affected relative growth in basal diameter. In contrast, initial seedling stem volume did not affect relative growth in volume (Table 5), which was not affected by the nursery treatments and watering in the field. Analysis of the sturdiness quotient showed a significant and negative coefficient (Table 5). In other words, a higher index at the time of transplant did not contribute to a higher quotient one year later. However, low-N fertilization and medium-light conditions in the nursery significantly and positively influenced the relative increment in seedling sturdiness after transplanting.
Field survival
Short term (1 year after transplanting)
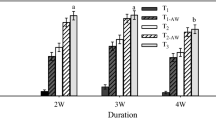
One year after planting, field survival among seedlings that had been watered in the summer of the first year after planting was 23% higher (Fig. 4) than for unwatered seedlings (88% and 65%, respectively). Regarding nursery treatments and the effect of the initial seedling stock attributes, we found the following. The first GLZ model (Table 6), which included seedling morphological variables (h0, d0, h0/t0 and v0) as continuous predictors and watering, fertilization, and light treatments as categorical predictors, showed no significant effects of nursery light and fertilization. However, no summer watering (NSW) after planting significantly increased the probability of mortality during the first summer (estimate = 0.959; Wald statistic = 9.927; P = 0.000). In the second GLZ model, which did not include seedling morphological attributes (Table 6), the effect of no watering remained significant (AIC = 117.46; estimate = 0.779; Wald statistic = 9.149; P = 0.002). However, the model revealed differences in field survival between seedlings grown in nursery with full sunlight and the other light treatments. The probability of mortality among seedlings grown in full sunlight was significantly lower (estimate =−0.942; Wald statistic = 5.442; P = 0.019) than that of seedlings grown in medium and low light. Then, we tested separately for the effect of medium- and low-light regimes on survival and consistently observed that a medium-light regime in the nursery decreased the probability of mortality in the field (estimate =−0.541; Wald statistic = 4.019; P = 0.045). However, a low-light regime in the nursery was associated with increased field mortality (estimate = 1.063; Wald statistic = 9.591; P = 0.002).

Survival of Pinus pinaster evaluated 1, 3 and 10 years after outplanting (1 June 2007) for effects of nursery fertilization and light regime, and field watering after transplanting. Initially, n = 10 seedlings (replicates) per treatment combination of light x fertilization x watering. HL = high light (average PPFD of 337.1 µmol m2 s−1); ML = medium light (average PPFD of 120.6 µmol m2 s−1); LL = low light (average PPFD of 30.0 µmol m2 s−1); low N = 222 mg/seedling; high N = 444 mg/seedling; SW = summer watering (160 mm); NSW = no summer watering (104 mm). Numbers within squares indicate the percentage of seedling survival
Medium-term (3 years) and long-term (10 years) field survival
As shown in Fig. 4, survival had decreased only slightly by 2010. Fertilization in the nursery and the post-planting watering treatment had no significant effects on seedling survival three years after outplanting. However, the full-sunlight treatment in the nursery still significantly decreased the probability of mortality in the field (estimate =−0.957; Wald statistic = 10.20; P = 0.001). Seven years after the second evaluation (2017), the survival rate was almost identical to that of the previous evaluation in all seedling groups (Fig. 4). As predicted, a significant effect of nursery light on field survival was still observable. Full sunlight in the nursery decreased the probability of seedling mortality in the field (estimate =−0.924; Wald statistic = 10.11; P = 0.001).
Discussion
Nursery regimes and seedling stock attributes
The results obtained in this study support the hypothesis that nursery light regime plays a greater part than nursery N fertilization in the performance and morphology of Pinus pinaster seedlings. The effect of fertilization was also important but varied according to the light regime, because light is needed for the expression of N effects on P. pinaster seedling growth (Rodríguez-García and Bravo 2013) and because expression of light-dependent plasticity depends on nutrient availability (Portsmuth and Niinemets 2007). Although high-N fertilization tended to increase shoot height and basal diameter, there were no significant differences between high-N and low-N seedlings within each of the light regimes. In contrast, the high-N fertilization treatment significantly reduced the sturdiness quotient attained by seedlings under medium-light conditions. Under the low-light regime, high-N fertilization also significantly reduced seedling shoot height to levels comparable to plants grown in full-sunlight conditions and seedling stem volume to almost half of the values observed in low-N plants.
This greater response to fertilization in shade environments has been documented for rapid-growth pioneer species (Monnier et al. 2013). It suggests that Maritime pine seedlings may perform better in full sunlight while also exhibiting a degree of shade tolerance and phenotypic plasticity in morphological aboveground traits during the early establishment stage (Sánchez-Gómez et al. 2006; Rodríguez-García et al. 2011a, b; Corcuera et al. 2011) that favour survival through acclimatation (Collet et al. 2006; Monnier et al. 2013). During this stage, such traits may not be especially responsive to soil fertility (Valdecantos et al. 2006) under high or medium light availability and would only respond under very shady conditions and/or to very high nutrient availability in deep shade (e.g. in very dense regenerations after fire). This was observed in medium-light and especially in low-light seedlings with high-N fertilization, which reduced aboveground investment and seedling elongation as shade-avoidance strategies. It implies that under canopy cover, high-N fertilization may be a tool for avoiding biomechanically weak phenotypes with poor light interception efficiency (Pearcy et al. 2005; Sánchez-Gómez et al. 2006).
On fertile sites, stem growth is essential as it allows individual trees to intercept light and avoid overtopping from neighbours, including shrubs (King 2003). Our results show that with higher nutrient availability in shade, P. pinaster shade tolerance mechanisms would not signal seedlings to increase height as a shade avoidance strategy, and competition for nutrients would become a higher priority than competition for light. A high nutrient supply generates early root growth and a positive carbon balance that helps sustain the demand for light to maintain aboveground growth biomass (Luis et al. 2009; Cuesta et al. 2010a, b; Villar-Salvador et al. 2012; Ovalle et al. 2016).
Regarding seedling size, the maximum height and minimum root collar diameter of our seedlings at the end of the nursery period were higher (Table 2) than the market requirements for one-year-old Pinus pinaster seedlings (30 cm and 2 mm, respectively) (Ministerio de la Presidencia 2003). The treatments applied in the study probably enhanced the development of these attributes.
Post-transplant stress and seedling performance
Higher seedling size at planting time negatively affected subsequent first-year growth. After adjusting for initial seedling size, the magnitude of the transplanting stress index (TSIadj), the nursery light and fertilization effects and the effect of post-planting watering varied depending on the seedling attribute. The TSIadj values observed in P. pinaster in this study were high, and dramatically higher than the TSIadj values observed for P. halepensis (Puértolas et al. 2003; Oliet et al. 2009). For Maritime pine, this indicates an initially low degree of adjustment to the site (Oliet et al. 2002) and that differences in seedling size are very important (Rodríguez-García et al. 2011b) in relation to initial growth after transplanting. For instance, the first-year relative growth in height and root basal diameter in the field were controlled by nursery light. A full-sunlight regime in the nursery reduced transplant stress in the field and is likely connected to the lower shoot height and sturdiness quotient of those seedlings, as well as a more equal shoot/root ratio (Luis et al. 2009), higher needle biomass (Rodríguez-García and Bravo 2013), higher photosynthesis efficiency (Bazzaz and Wayne 1994) and higher basal diameter (Collet et al. 2006). Some studies point out that the probability of seedling survival at the end of each year increases with higher basal diameter (Villar-Salvador et al. 2012; Kabrick et al. 2015 and references within). This demonstrates a relationship between easily measurable aboveground seedling morphological variables and new root elongation (Kabrick et al. 2015) after transplanting to overcome transplant shock (Oliet et al. 2002; Villar-Salvador et al. 2012). The sturdiness quotient of seedlings grown in the nursery with medium light and low N had a positive coefficient: they grew taller but thinner than the rest of seedlings. These results show great variability in size attributes and growth responses that were dependent on the nursery regimes. They also point to potential post-planting interactions (e.g., competition and vegetation dominance) between seedling size and site characteristics (drought and soil fertility, canopy light) (Oliet et al. 2009) that may influence the provision and functionality of restored forest and ecosystem services.
Effects of nursery regimes and post-transplant summer watering on seedling survival
The results obtained in this study support the hypothesis that the nursery light regime combined with higher summer water availability in the field play a greater part than nursery N fertilization in the survival of transplanted Pinus pinaster seedlings, because high-N fertilization in the nursery did not directly affect post-planting seedling survival. Prior research has documented the significant effects of in-nursery fertilization on the size of seedling stock, but not on seedling survival in the field after outplanting (see Trubat et al. 2010 and Ovalle et al. 2016). This may be linked to interactions of the fertilization regime with culturing characteristics (Villar-Salvador et al. 2012), species strategy, site conditions (local climate, soil heterogeneity) and planting conditions (Cortina et al. 2013). It highlights the need to match nursery fertilization regimes with the site characteristics (Oliet et al. 2021) and the specific objectives (e.g., wood production, resin production, soil conservation) of the plantation that receives the seedlings. As expected, post-planting first-year summer irrigation reduced the probability of seedling mortality in the field. Pardos et al. (2015) have also documented how first-year summer irrigation ensured seedling survival and physiological performance in outplanted Pinus pinea seedlings. Similarly, Ruano et al. (2009) reported that field watering after outplanting greatly influenced the growth and biomass allocation of naturally regenerated P. pinaster seedlings.
In our study, full-sunlight in the nursery decreased outplanted seedling mortality much more than medium light, which had no effect on outplanted seedling mortality. However, seedlings grown in low-light nursery conditions had a higher probability of field mortality after transplanting. Increased sturdiness in the low-light stock seedlings, along with the resulting imbalance created by their smaller initial basal diameter and stem volume, may have been negative for water stress avoidance and field survival. The increased field survival of seedlings cultured with full sunlight available in the nursery may be linked to photosynthesis, which supports early root growth (Villar-Salvador et al. 2012) and a more balanced aboveground morphological structure. Other studies about the interaction of light and water on water-use efficiency in seedlings of Mediterranean species such as cork oak (Quercus suber L.) show that seedlings receiving higher irradiance maintain a higher rate of carbon uptake at a particular stomatal conductance. This suggests that shaded seedlings have a lower water-use efficiency that is unrelated to water availability (Aranda et al. 2007). Species are often constrained by multiple stress factors operating simultaneously, which is an area that needs much more research. The good results of our combined nursery study and field experiment can provide valuable insights for nursery and forest managers.
Conclusions
The effect of fertilization on seedling aboveground morphological attributes after one growing season in the nursery became more evident (significant) with increased nursery shade. Nevertheless, fertilization in the nursery did not condition field survival. Field survival one growing season after outplanting was significantly higher for seedlings cultured in full-sunlight nursery conditions and for those that received first-year summer irrigation following outplanting. Medium-light conditions in the nursery did not increase field mortality but low-light conditions did, probably because those seedlings had very low basal diameter and stem volume, and higher shoot height and sturdiness than seedlings cultured in full sunlight and medium light. These results can be used by nursery managers to develop and produce high-quality stock that increases reforestation success. They can also inform forest managers in their efforts to assess, increase or sustain forest functionality for Maritime pine and other Mediterranean pine reforestation projects.
References
Alía R, Martín S, De Miguel J, Galera RM, Agúndez D, Gordo J, Salvador L, Catalán G, Gil L (1996) Regiones de procedencia de Pinus pinaster Aiton. Área de Selvicultura y Mejora. INIA-CESEFOR, Madrid
Alía R, Garcia del Barrio JM, Iglesias S et al (2009) Regiones de procedencia de especies forestales de España. Organismo Autónomo Paques Nacionales, Madrid
Andivia E, Villar-Salvador P, Oliet JA et al (2021) Climate and species stress resistance modulate the higher survival of large seedlings in forest restorations worldwide. Ecol Appl 31:1–11. https://doi.org/10.1002/eap.2394
Aranda I, Pardos M, Puértolas J et al (2007) Water-use efficiency in cork oak (Quercus suber) is modified by the interaction of water and light availabilities. Tree Physiol 27:671–677. https://doi.org/10.1093/treephys/27.5.671
Bazzaz FA, Wayne PM (1994) Coping with environmental heterogeneity: the physiological ecology of tree seedling regeneration across the gap-understory continuum. In: Caldwell MM, Pearcy RW (eds) Exploitation of environmental heterogeneity by plants: ecophysiological processes above- and belowground. Academic Press, Cambridge
César RG, Belei L, Badari CG et al (2021) Forest and landscape restoration: a review emphasizing principles, concepts, and practices. Land 10:1–22. https://doi.org/10.3390/land10010028
Collet C, Chenost C (2006) Using competition and light estimates to predict diameter and height growth of naturally regenerated beech seedlings growing under changing canopy conditions. Forestry 49:489–502. https://doi.org/10.1093/forestry/cpl033
Cortina J, Vilagrosa A, Trubat R (2013) The role of nutrients for improving seedling quality in drylands. New for 44:719–732. https://doi.org/10.1007/s11056-013-9379-3
Corcuera L, Cochard H, Gil-Pelegrin E, Notivol E (2011) Phenotypic plasticity in mesic populations of Pinus pinaster improves resistance to xylem embolism (P50) under severe drought. Trees 25:1033–1042. https://doi.org/10.1007/s00468-011-0578-2
Cuesta B, Vega J, Villar-Salvador P, Rey-Benayas JM (2010a) Root growth dynamics of Aleppo pine (Pinus halepensis Mill.) seedlings in relation to shoot elongation, plant size and tissue nitrogen concentration. Trees Struct Funct 24:899–908. https://doi.org/10.1007/s00468-010-0459-0
Cuesta B, Villar-Salvador P, Puértolas J et al (2010b) Why do large, nitrogen rich seedlings better resist stressful transplanting conditions? A physiological analysis in two functionally contrasting Mediterranean forest species. For Ecol Manag 260:71–78. https://doi.org/10.1016/j.foreco.2010.04.002
Fernández M, Marcos C, Tapias R et al (2007) Nursery fertilisation affects the frost-tolerance and plant quality of Eucalyptus globulus Labill. cuttings. Ann for Sci 64:865–873. https://doi.org/10.1051/forest:2007071
Grossnickle SC, McDonald JE (2018a) Seedling quality: history, application, and plant attributes. Forests 9:283. https://doi.org/10.3390/f9050283
Grossnickle SC, McDonald JE (2018b) Why seedlings grow: influence of plant attributes. New for 49:1–3. https://doi.org/10.1007/s11056-017-9606-4
Grossnickle SC, Ivetic V (2022) Root system development and field establishment: effect of seedling quality. New for 53:1021–1067. https://doi.org/10.1007/s11056-022-09916-y
Johnson JB, Omland KS (2004) Model selection in ecology and evolution. Trends Ecol Evol 19:101–108. https://doi.org/10.1016/j.tree.2003.10.013
Kabrick JM, Knapp BO, Dey DC, Larsen DR (2015) Effect of initial seedling size, understory competition, and overstory density on the survival and growth of Pinus echinate seedlings underplanted in hardwood forests for restoration. New for 46:897–918. https://doi.org/10.1007/s11056-015-9487-3
Kildisheva OA, Aghai MM, Bouazza K, Davis AS (2017) Improving restoration success through research-driven initiatives: case studies targeting Pinus pinea reforestation stock development in Lebanon. Plant Ecol 218:39–53. https://doi.org/10.1007/s11258-016-0632-7
King DA (2003) Allocation of above-ground growth is related to light in temperate deciduous saplings. Funct Ecol 17:482–488. https://doi.org/10.1046/j.1365-2435.2003.00759.x
Lavorel S (2013) Plant functional effects on ecosystem services. J Ecol 101:4–8. https://doi.org/10.1111/1365-2745.12031
Lenssen JPM, Menting FBJ, Van der Putten WH (2003) Plant responses to simultaneous stress of waterlogging and shade: amplified or hierarchical effects? New Phytol 157:281–290. https://doi.org/10.1046/j.1469-8137.2003.00666.x
Levene H (1960) Robust tests for the equality of variances. In: Olkin J (ed) Contributions to probability and statistics. Stanford University Press, Palo Alto
Luis VC, Puértolas J, Climent J et al (2009) Nursery fertilization enhances survival and physiological status in Canary Island pine (Pinus canariensis) seedlings planted in a semiarid environment. Eur J for Res 128:221–229. https://doi.org/10.1007/s10342-009-0257-7
Luo N, Villar-Salvador P, Li G, Wang J (2021) The dark side of nursery photoperiod reduction on summer plantation performance of a temperate conifer: high winter mortality mediated by reduced seedling carbohydrate and nitrogen storage. For Ecol Manag 491:119171. https://doi.org/10.1016/j.foreco.2021.119171
McCullagh P, Nelder JA (1989) Generalized linear models. Great Britain at the University Press, Cambridge
Merine AK, Rodríguez-García E, Alía R et al (2015) Effects of water stress and substrate fertility on the early growth of Acacia senegal and Acacia seyal from Ethiopian Savanna woodlands. Trees- Struct Funct 29:593–604. https://doi.org/10.1007/s00468-014-1138-3
Ministerio de la Presidencia (2003) Real Decreto 289/2003, de 7 de marzo, sobre comercialización de los materiales forestales de reproducción
Monier Y, Bousquet-Mélou A, Vila B, Prévosto B, Fernandez C (2013) How nutrient availability influences acclimatation to shade of two (pionner and late-successional) Mediterranean tree species? Eur J for Res 132:325–333. https://doi.org/10.1007/s10342-012-0677-7
Oliet J, Planelles R, López-Arias M, Artero F (2002) Soil water content and water relations in planted and naturally regenerated Pinus halepensis Mill. seedlings during the first year in semiarid conditions. New for 23:31–44. https://doi.org/10.1023/A:1015668815037
Oliet JA, Planelles R, Artero F et al (2009) Field performance of Pinus halepensis planted in Mediterranean arid conditions: relative influence of seedling morphology and mineral nutrition. New for 37:313–331. https://doi.org/10.1007/s11056-008-9126-3
Oliet JA, Puértolas J, Planelles R, Jacobs DF (2013) Nutrient loading of forest tree seedlings to promote stress resistance and field performance: a Mediterranean perspective. New for 44:649–669. https://doi.org/10.1007/s11056-013-9382-8
Oliet JA, Puértolas J, Valenzuela P, Vázquez de Castro A (2021) Light transmissivity of tree shelters interacts with site environment and species ecophysiology to determine outplanting performance in Mediterranean climates. Land 10:753. https://doi.org/10.3390/land10070753
Ostertag R, Warman L, Cordell S, Vitousek PM (2015) Using plant functional traits to restore Hawaiian rainforest. J Appl Ecol 52:805–809. https://doi.org/10.1111/1365-2664.12413
Ovalle JF, Arellano EC, Oliet JA et al (2016) Linking nursery nutritional status and water availability post-planting under intense summer drought: the case of a south american mediterranean tree species. Iforest 9:758–765. https://doi.org/10.3832/ifor1905-009
Pardos M, Calama R, Mayoral C et al (2015) Addressing post-transplant summer water stress in Pinus pinea and Quercus ilex seedlings. Iforest 8:348–358. https://doi.org/10.3832/ifor1256-007
Pearcy RW, Muraoka H, Valladares F (2005) Crown architecture in sun and shade environments: assessing function and trade-offs with a three-dimensional simulation model. New Phytol 166:791–800. https://doi.org/10.1111/j.1469-8137.2005.01328.x
Portsmuth A, Niinemets U (2007) Structural and physiological plasticity in response to light and nutrients in five temperate deciduous woody species of contrasting shade tolerance. Funct Ecol 21:61–77. https://doi.org/10.1111/j.1365-2435.2006.01208.x
Puértolas J, Benito LF, Peñuelas JL (2009) Effects of nursery shading on seedling quality and post-planting performance in two Mediterranean species with contrasting shade tolerance. New for 38:295–308. https://doi.org/10.1007/s11056-009-9148-5
Puértolas J, Gil L, Pardos JA (2003) Effects of nutritional status and seedling size on field performance of Pinus halepensis planted on former arable land in the Mediterranean basin. Forestry 76:159–168. https://doi.org/10.1093/forestry/76.2.159
Rodríguez-García E, Bravo F (2013) Plasticity in Pinus pinaster populations of diverse origins: Comparative seedling responses to light and Nitrogen availability. For Ecol Manag 307:196–205. https://doi.org/10.1016/j.foreco.2013.06.046
Rodríguez-García E, Bravo F, Spies TA (2011a) Effects of overstorey canopy, plant-plant interactions and soil properties on Mediterranean Maritime pine seedling dynamics. For Ecol Manag 262:244–251. https://doi.org/10.1016/j.foreco.2011.03.029
Rodríguez-García E, Juez L, Bravo F (2010) Environmental influences on post-harvest natural regeneration of Pinus pinaster Ait. in Mediterranean forest stands submitted to the seed-tree selection method. Eur J for Res 129:1119–1128. https://doi.org/10.1007/s13595-011-0078-y
Rodríguez-García E, Ordóñez C, Bravo F (2011b) Effects of shrub and canopy cover on the relative growth rate of Pinus pinaster Ait. seedlings of different sizes. Ann for Sci 68:337–346. https://doi.org/10.1007/s13595-011-0039-5
Ruano I, Manso R, Fortin M, Bravo F (2015) Extreme climate conditions limit seed availability to successfully attain natural regeneration of Pinus pinaster in sandy areas of central Spain. Can J for Res 45:1795–1802. https://doi.org/10.1139/cjfr-2015-0257
Ruano I, Pando V, Bravo F (2009) How do light and water influence Pinus pinaster Ait. germination and early seedling development? For Ecol Manag 258:2647–2653. https://doi.org/10.1016/j.foreco.2009.09.027
Sánchez-Gómez D, Valladares F, Zavala MA (2006) Performance of seedlings of Mediterranean woody species under experimental gradients of irradiance and water availability: trade-offs and evidence for niche differentiation. New Phytol 170:795–806. https://doi.org/10.1111/j.1469-8137.2006.01711.x
Serrada R, Montero G, Reque JA (2008) Compendio de selvicultura aplicada en España. Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria, y Ministerio de Educación y Ciencia. Madrid
Shapiro SS, Wilk MB (1965) An analysis of variance test for normality (complete samples). Biometrika 52:591–611. https://doi.org/10.2307/2333709
Shi W, Grossnickle SC, Li G et al (2019) Fertilization and irrigation regimes influence on seedling attributes and field performance of Pinus tabuliformis Carr. Forestry 92:97–107. https://doi.org/10.1093/forestry/cpy035
South DB, Zwolinski JB (1997) Transplant Stress Index: a proposed method of quantifying planting check. New for 13:311–324. https://doi.org/10.1023/a:1006546627342
Trubat R, Cortina J, Vilagrosa A (2010) Nursery fertilization affects seedling traits but not field performance in Quercus suber L. J Arid Environ 74:491–497. https://doi.org/10.1016/j.jaridenv.2009.10.007
Trubat R, Cortina J, Vilagrosa A (2011) Nutrient deprivation improves field performance of woody seedlings in a degraded semi-arid shrubland. Ecol Eng 37:1164–1173. https://doi.org/10.1016/j.ecoleng.2011.02.015
Valdecantos A, Cortina J, Vallejo VR (2006) Nutrient status and field performance of tree seedlings planted in Mediterranean degraded areas. Ann for Sci 63:249–256. https://doi.org/10.1051/forest
Villar-Salvador P, Peñuelas JL, Nicolás-Peragón JL et al (2013) Is nitrogen fertilization in the nursery a suitable tool for enhancing the performance of Mediterranean oak plantations? New for 44:733–751. https://doi.org/10.1007/s11056-013-9374-8
Villar-Salvador P, Planelles R, Enríquez E, Rubira JP (2004) Nursery cultivation regimes, plant functional attributes, and field performance relationships in the Mediterranean oak Quercus ilex L. For Ecol Manag 196:257–266. https://doi.org/10.1016/j.foreco.2004.02.061
Villar-Salvador P, Puértolas J, Cuesta B et al (2012) Increase in size and nitrogen concentration enhances seedling survival in Mediterranean plantations. Insights from an ecophysiological conceptual model of plant survival. New for 43:755–770. https://doi.org/10.1007/s11056-012-9328-6
Acknowledgements
The authors are grateful to the staff who assisted in the nursery and in the field, and to the regional government for making the land available for the experiment. The authors gratefully acknowledge the Spanish Ministry of Science and Education, the MEDFOR research program, and the Project INTERNATIONALISATION OF EXCELLENCE RESEARCH STRUCTURES (CL-EI-2021-05) funded by the Junta de Castilla y león, and linked to the Project "Escalera de la Excelencia -Instituto iuFOR" (CLU -2019-01) of the University of Valladolid and co-financed by the European Regional Development Fund (ERDF "Europe drives our growth"). The authors thank the Fundação para a Ciência e a Tecnologia (FCT, Portugal) and the European Commission for their support to M. A. Al Pavel through a doctoral grant (SFRH/PD/BD/135442/2017) and an exceptional doctoral grant (COVID/BD/152046/2021).
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature. This work was financially supported by the Junta de Castilla y León (University of Valladolid project ‘CLU-2019-01-iuFOR Institute Unit of Excellence’), co-financed by the European Union (ERDF ‘Europe drives our growth’). This research was also funded by an Erasmus Mundus International Master’s course in MEDFOR (Mediterranean Forestry and Natural Resources Management, Reference 520137-1-2011–1-PT-ERA MUNDUS-EMMC) which sponsored Muha Abdullah Al Pavel, who was awarded a full Erasmus Mundus Partner Country Scholarship.
Author information
Authors and Affiliations
Contributions
MAAP: field plantation, field data collection; exploratory statistical analyses, early draft writing and editing, and final version review. FB: conceptualization; project administration; methodology, reviewing and editing. CO: methodology; assistance in the nursery experiment; field plantation and data collection; reviewing and editing. ER-G: nursery experiment; final data analysis; writing, reviewing and editing of final version.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Al Pavel, M.A., Bravo, F., Ordóñez, C. et al. Linking the effects of nursery light and fertilization and post-transplant first summer irrigation on the field survival and growth of Pinus pinaster seedlings. New Forests 55, 251–271 (2024). https://doi.org/10.1007/s11056-023-09973-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11056-023-09973-x