
Abstract
How much do different groups of humans differ genetically? Starting from a thought experiment, this chapter shows that we differ less than we might think—and that, in a sense, we are all Africans. I draw on several key findings: that compared to many other species, Homo sapiens has relatively little genomic variation; that of all continental regions, African genomic variation is the richest and most encompassing; that population-level genetic variation decreases with distance from Africa; and that most genomic variation is found within local human populations (with only about 7% or so found across continental groups). This is not to deny that there are—small but somewhat systematic—group-level differences, but it is also important to recognize that these should not matter ethically or politically.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others


1 A Thought Experiment
Parts of this book chapter were adapted from chapters 6 and 9 of Winther (2022a).
A few months after your intergalactic expedition left Earth, you arrive in a far-off world.Footnote 1 Departing your ship, you come upon some of the planet’s inhabitants. They all seem to look the same. They are all more-or-less one meter tall and have the same muscular body and rounded facial features. You know that people sometimes say that other groups of people “all look the same,” but in this case it seems nearly true: finding a distinguishing mark on any one of them is a challenge.
“Greetings, traveler,” you hear through a universal translation device held up to the mouth of a single inhabitant who has emerged to meet you on the rock where you stand. “Please be at ease.” You are told that every person on the planet looks the same; your greeter is no exception. You learn that centuries ago there was a Great Tragedy brought on by ethnic conflict including a calamitous civil war. Committed to preventing further destruction, the survivors agreed that for the rest of time they would engage in random breeding to avoid ethnic strife.
Call this Planet Unity.
Now, let’s imagine a different scenario, perhaps a little more familiar: Imagine the Galápagos Islands, the natural experimental laboratory where Darwin studied evolution, with their wide variety and number of finch and tortoise species. Let’s in our minds populate these islands—or any analogous archipelago—with identical small populations of early humans. Let’s then add a few more dozen islands that are larger, mutually unreachable, and have distinct environments: Some are relatively cool and rainy, while others are hot and arid. What happens after tens of millions of years of evolution? Will the populations become more alike, similar to Planet Unity? Just the opposite, of course: Unlike on Planet Unity, the populations on these islands have come to be quite different from each other—not just in body and behavior, but in culture, as well.
Call this Galápagos-Writ-Large.
Now the real question: are modern humans more like the inhabitants of Unity or Galápagos-Writ-Large? Where do we fall between these extremes?
When we look around us on the street or in a Zoom meeting, we can easily see differences in people’s eyes, skin tone, height, and the rest. Identical twins might be a partial exception! This isn’t just a point about individuals: some groups of people seem to share physical traits in common. And this would seem to be an argument for humans fitting the Galápagos-Writ-Large scenario.
I’m going to use this chapter to suggest the opposite: the Planet Unity scenario is a much better description of our situation. To help convince you, we’ll review our best genomic evidence for the (relative) unity of the human species.
It was only in the middle of the twentieth century—long after Darwin’s death—that scientists started unlocking the mysteries of genetic inheritance at the molecular level. The environment is an essential part of making you what you are, but the other side of the matter is that molecule called DNA, which encodes our genes.
A reasonable starting point for examining human diversity, then, is to ask how genes differ, if at all, between individuals and groups. How similar or dissimilar genetically is anybody from anybody else, really? Let’s have a look.
The genomics of recent decades has had some surprising findings. We have learned that, compared to many other species, Homo sapiens has relatively little genomic variation; that of all continental regions, African genomic variation is the richest and most encompassing; that population-level genetic variation decreases with distance from Africa; and that most genomic variation is found within local human populations (with only about 7% or so found across continental groups).
What does this mean? As I will show, it means we are all Africans. But groups also differ, and these small but somewhat systematic group-level differences explode with highly charged and controversial political and ethical debates and implications.
2 Relatively Little Homo Sapiens Intraspecies Genomic Variation
Among species for which we have good data, Homo sapiens has rather low average nucleotide variation. At roughly 999 base pairs out of 1000 (on average), all members of Homo sapiens are basically identical.Footnote 2 Given our total, haploid genome size of roughly 3 billion nucleotides, two individuals will typically and on average differ at approximately 3 million nucleotides—only a difference of about 0.1%. Our closest cousins are on either side of us in this measure, but not too far: bonobos differ by 0.076%, chimpanzees by 0.134%, and gorillas by 0.158%.Footnote 3 By contrast, the most classic model organism for genetic studies, Drosophila fruit flies, differ from each other by 1% on average, or about 10 times our nucleotide diversity. And maize has even more nucleotide diversity than Drosophila.Footnote 4
In short, there is, according to our best genomic data, a single overarching human race, at least as compared to less unified species.
3 Out of Africa Migrations
Even so, we do differ in small ways, both at the individual level and the group level. This is what makes human evolutionary genomics so fraught and so interesting. Are there alleles associated with cancer, running speed, or intelligence, and might they be distributed unevenly across human populations? If so, what are the consequences for medicine, social policy, and our self-understanding?
It is crucial to recognize that our basic evolutionary pattern is Out of Africa migration. You may know the story: our species originated in Africa with a few small and highly related populations. Several times, groups left Africa, grew in size and range, and underwent evolution by natural selection, random genetic drift, and mutation. (Some migration back to Africa also occurred.) In this way, different human populations spanned the world, occasionally interbreeding, especially in the last tens of thousands of years.Footnote 5
3.1 Of All Continental Regions, African Genomic Variation is the Richest and Most Encompassing
African human populations are the most genetically variable in the world. African populations have roughly double the nucleotide diversity of other populations. In other words, two people whose recent ancestors are of African origin differ on average by about 1:900 nucleotides (0.11%), whereas two people whose recent ancestors are of European origin differ on average by only approximately 1:1600 (0.063%).Footnote 6 Another study found that Mandinka peoples from Mali, Guinea, and the Ivory Coast and the San peoples of South Africa harbored nucleotide diversities of 0.12% and 0.126%, respectively, while Han Chinese and Basque peoples had, respectively, 0.081% and 0.087%.Footnote 7 According to Rosenberg (2011), Africa has approximately half of the world’s “private alleles”—that is, alleles at a locus unique to a geographic region.Footnote 8 Many more pieces of evidence indicate that genomic variation is generally much higher in Africa, in part due to the long periods of time of human population differentiation across the African continent. Given our Out of Africa picture, none of this is surprising.
3.2 Genomic Heterozygosity of Populations Decreases with Increasing Distance from Africa, Along Human Migration Routes
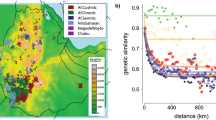
Early humans had to pass through northeastern Africa to leave it, and following this path leads to interesting insights. Genetic variation in human populations reduces in proportion to its distance from Africa, as measured along historical human migration routes. This means that the further apart two populations are along these lines, the more they differ. Here we can talk about human migration as well as genetic migration, or the process of gene exchange when populations mix. Genetic migration also tends to diminish along human migration routes as humans move farther from Africa.Footnote 9

(Source Fig. 4a, p. 15946, Support from the relationship of genetic and geographic distance in human populations for a serial founder effect originating in Africa by Ramachandran et al. (2005); https://doi.org/10.1073/pnas.0507611102 © (2005) National Academy of Sciences, U.S.A. Reprinted with permission from PNAS)
Heterozygosity to distance from Eastern Africa global pattern. The amount of heterozygosity of each of approximately 40 worldwide populations diminishes as a function of their respective distance, along (approximate) migration routes, from Addis Ababa.
The Out of Africa migration model also aligns with the data on the decline in genomic variation as you move away from our evolutionary cradle, providing very strong evidence for a general Out-of-Africa scenario of human evolution (Fig. 2.1). This occurs because when a species migrates, relatively few individuals typically move and survive. Even fewer make it when difficult barriers such as mountains are crossed. This means two things. First, due to random genetic drift (random changes in allele frequencies across generations), small populations tend to lose genetic variation over generations (more so the smaller they are). Second, migrating natural populations experience genetic bottlenecks or a “founder effect” whenever a small group inhabits new areas.Footnote 10 Each time this happens, the founder population represents only some of the genetic variation of its parent population. These factors combine to lower the amount of genomic variation, especially outside of Africa, the home of our originary genetic variation. Interdisciplinarity and acceptance of multiple models and methods will also help paint the full picture here. For instance, archaeological and paleontological evidence can deepen and broaden our analysis.
Africa is indisputably the capital of Planet Unity—as well as the most diverse part of that planet. This is not to deny, of course, that we can see features of Galápagos-Writ-Large as a consequence of human migration and local adaptation to distinct environments.
3.3 Two Groundbreaking Episodes
In 1972, biologist Richard Lewontin studied the distribution of alleles across individuals from numerous groups around the globe.Footnote 11 He used genes coding for blood proteins to compare genetically different individuals within a population to individuals from other populations, both within and across continental regions. Here is his list of seven continental regions, into which he pooled populations that had been sampled and whose allele distributions (or allele frequencies) had been identified: Africans, Europeans, Asians, Southeast Asians, Indigenous Americans, Oceanians, Australian Aborigines.
Lewontin found something astonishing: genetically, speaking, you are almost as different from someone chosen at random from your group as you are from a random person from anywhere on the globe. Our familiar racial categories are not backed up by the genetics. The human species has very little genetic variation, and it is mostly between individuals within groups—not between groups.
Nevertheless, 40 years later, a group of researchers led by Noah Rosenberg showed that even these small between-group differences provide enough information to correctly assign someone to a racial group based on knowing about 20–30 of her genes.Footnote 12
What are we to make of this mix of results? Lewontin’s work seems to suggest that races and other large human populations aren’t, genetically speaking, “really real.” But then how could it be possible to assign you to a race or ethnicity using just a small sample of your genes? This is not an idle question. Biologists and doctors—not to mention big pharma and insurance companies—have large stakes in understanding this puzzle.
The full answer as to why this mix of findings can co-exist would take more detail than available here. What I have elsewhere called Lewontin’s “variance partitioning” and Rosenberg’s “clustering analysis” are two sides of the same coin, mathematically speaking (Winther 2014). The twofold truth is this: human populations are similar in allele frequencies at many genes—but whenever we aggregate information about small differences across two populations, we can use this information to cluster and classify individuals into populations. Knowing an individual’s multilocus genotype provides just this information. Although some refer to “Lewontin’s Fallacy,” I see no fallacy here: Lewontin and Rosenberg were interested in two different sets of questions and methods.
Recall our thought experiment: Planet Unity and Galápagos-Writ-Large are extremes on a spectrum. Yet, in general and on average, our species fits the Planet Unity model surprisingly well. This makes sense in light of Lewontin’s results. And, in at least some places on Earth—especially big multicultural cities—our children are becoming even more Unity-like, both genetically and culturally. We see Galápagos-Writ-Large properties when we zoom in to a finer grain to explore specific genes or small populations. Admittedly, for some genes there are very large frequency differences in different parts of the world. This shows the validity of Rosenberg and colleagues’ results.
4 Is Race Biologically Real?
Some claim that our best science indicates that human races are biologically real entities (e.g., Robin Andreasen, Charles Murray, Neven Sesardić, Quayshawn SpencerFootnote 13). According to this view of racial realism, racial groups are clearly genetically differentiated. I believe that our best genomics obviously undercuts such a view. A subset of racial realists—call them “hereditarians”—argue further that the existence of biological populations explains and justifies certain social inequalities (e.g., Herrnstein and Murray 1995; Murray 2020). For them, multiple social and political inequalities, both within and between nations, are due largely to hereditary differences in the (average) innate abilities of races (as this concept is usually conceived)—intelligence, for instance. This view gives us a Galápagos-Writ-Large picture of biological race. Many hereditarians stand on the political right (e.g., conservatives, libertarians).
Compare the racial antirealism view developed in the wake of Lewontin’s (1972) paper “The Apportionment of Human Genetic Diversity” (e.g., Joshua Glasgow, Adam Hochman, Naomi ZackFootnote 14). For the racial anti-realist, races are social fictions without a biological basis. Such a view is widespread throughout the social sciences,Footnote 15 and is widely held by those who identify with the academic left. It can be thought of as the liberal consensus on race. In my view, this position goes too far and is somewhat disingenuous—it simply ignores some data.
This controversy is riddled with burden of proof arguments: both sides like to argue that the burden lies with the other side. The left wants the right to show strong genetic evidence for specific traits, while the right wants the left to prove that no genetic explanation for between-group differences can be given.
I would suggest that our global genome leaves it open to interpretation whether we can say that racial groups exist or not. The story is complex. Interpretation differs depending on the trait—blood proteins, skin color, behaviors, etc. At any rate, our best genomic studies in the last decade have shown that racial groups aren’t very strongly identifiable and don’t explain very much.Footnote 16 Moreover, the Out of Africa paradigm is basically correct. Ample genomic evidence shows, beyond a doubt, that Africa is the capital of our Planet Unity (albeit the most internally diverse part). Even so, while we are all quite similar, there may be real heritable differences that could, potentially, be politically controversial, especially if they inspire public policy or medical interventions.
We should also bear in mind that the story of race, however we conceive this fraught concept and category, is not, for the most part, a genetic story. Cultural narratives, questions of identity and belonging, are central in our attempts to understand all human groups and populations, racial or otherwise. As we will see below, whether or how race is biologically real is a separate issue from whether or how it might be socially real.
5 Philosophy and Post-Racial Futures
I believe philosophical analysis of genomics of race shows that race must be characterized in both realist (social race) and antirealist (biological race) terms. (And in conventionalist terms for genomic race.) It all depends on which “race” you’re talking about. Mills (1998) provides a useful taxonomy of views, ranging from the denial of races’ existence (Glasgow 2009) to the belief in deep biological racial “essences.” To me, Mills’ “objective constructivism” seems the most plausible: races are socially real and are shaped by ongoing social practices. Ásta (2018) provides a similar “conferralist” framework for analyzing the social construction of race and other categories such as sex and gender. For Ásta, race is defined by a social property, namely, “social status consisting in constraints on and enablements to the individual’s behavior in a context” (2018, 2). Finally, psychiatrist and philosopher Frantz Fanon illustrates the reality and consequences of social race (e.g., structural racism), in the European colonial context.Footnote 17
Crucially, the social construction process of race is historical and contingent.Footnote 18 This means that racial categories could always be otherwise. In When Maps Become the World, I defended a practice of imagining “What if…?”:
In posing this most capacious question, philosophy opens up a space for memories, feelings, hopes, and imagination. When we ask “What if …?,” we swap one set of assumptions for another and follow the world-making consequences of each, whether in the future or in potential existence more generally. Perhaps this is a kind of future-oriented pluralistic ontologizing. … What if social relations were structured with institutions, values, and behaviors dramatically different from those in place here, today? (Winther 2020, p. 253)
Let us here briefly address one “what if…?” possibility, that of post-racial societal futures.
A post-racial future is one in which perceived membership in a racial category is no longer associated with differential access to social goods, including dignity, trust, and opportunity. For such a future to exist, we must rip up what Charles Mills calls the racial contract. According to Mills, the seemingly neutral social contract theory tradition—including such philosophical luminaries as Thomas Hobbes, John Locke, Jean-Jacques Rousseau, Immanuel Kant, and John Rawls—has effectively posited a social contract between just the people who count, which is to say white people. Plausibly, he suggests that “White supremacy is the unnamed political system that has made the modern world what it is today.” A post-racial future, then, requires struggles for recognition, power, land, and a decent standard of living—for an expanded social contract that includes all.Footnote 19
Post-racial societies seem possible, and perhaps desirable. They are possible if, per some of the above discussions, each of us is willing to do the hard, critical work of simultaneously feeling a sense of belonging to our respective groups and to all of us, and to all of life. In so doing, we must also work hard not to reify and naturalize social expectations and prejudice. The result of the work need not be societies in which people no longer “see” race as a social organizing principle. For instance, social race, as I have defined it, may continue to have some relevance, but not for oppressive or negatively discriminatory purposes—perhaps only to celebrate differences, as some like to say, in culture, music, or food. Whether such a society, in the absence of stereotypical racial judgments, would be more inclined to reward people based on any number of individual qualities, such as work ethic or charm, is a matter for further “what if…?” discussion.Footnote 20
Allow me to put my point starkly: genomics simply shouldn’t matter to politics.Footnote 21 Regardless of genomic results and what they could or could not tell us, we must continue to work towards equality and dignity—also for non-human life—premised on ethics and law instead of on scientific information. Normative equality simply does not depend upon factual equality. Abstracting away from genomics, I encourage us to strengthen moral and legal frameworks to guarantee equality and protection to all.
6 Coda: Who Are We?
Who am I? It no longer makes sense for us to ask this question. Here, on this pale simulacrum of Planet Unity, we learn more about ourselves as individuals when we ask about the collective: Who are we? Who were we? Where did we come from—from what places and what ancestors? Whence humanity?
These questions include another as their flipside: Who are we not? Homo sapiens—that collection of us oddball apes—is not Homo erectus or Homo heidelbergensis, let alone Australopithecus anamensis. Nor are we Neanderthals or Denisovans, although here there is much more overlap. There are morphological and (postulated) behavioral differences between all hominin species and sub-species, including differences in average brain size, presence or absence of air sacks, and the production of particular forms of tools and weapons.
Neanderthals and Denisovans both likely emerged out of Africa. Neanderthals also evolved in Europe and western Asia, and Denisovans called eastern and southern Asia—and later Oceania—their home. There are reasonably good African Homo fossils before 400,000 years and after 260,000 years ago. In between lies a gap. Europe, meanwhile, has a solid fossil record for Neanderthal throughout this period (Hublin 2013).
These two subspecies or sister species of Homo sapiens elicit much interest. After all, most humans outside of Africa today have a few percentage of Neanderthal DNA, some of which is adaptive, for example, in fighting viruses.Footnote 22 And Oceanian peoples such as Papuans and Australian aborigines often have more than 5% Denisovan DNA.Footnote 23 Recent research finds Neanderthal DNA even in contemporary African individuals, suggesting that there were “Back to Africa” migrations, perhaps even by European Homo sapiens, themselves carrying Neanderthal DNA, over the last 20,000 years or so.Footnote 24
Still, we have no evidence of Neanderthal or Denisovan mitochondrial DNA or Y-chromosomal haplotypes in any modern human population. Perhaps male fetuses with Neanderthal fathers and human mothers were inviable.Footnote 25 I leave it to the interested reader to draw out various mating combinations, in order to see that both Neanderthal mitochondrial DNA and Y-chromosomes can be lost in two generations, while autosomal DNA easily introgresses into the human genealogy.Footnote 26
While we have a few answers, many more remain: The history of Homo cannot but raise questions about how to untangle our DNA, “whose” DNA it is, and why Neanderthals went extinct approximately 40,000 years ago.Footnote 27 Taking a wider view, we can ask which species—and places—gave rise to our nearest contemporary cousins: bonobos, chimpanzees, and gorillas (Fig. 2.2). Such questions bring us closer to our ancestors, inviting a sense of connection and belonging across the branches of the tree of life. Our best genomics blurs all kinds of putative boundaries, showing the deep and broad connection of all life.

Our cousins: Bonobos. Bonobos (Pan paniscus) are known for their empathy, matriarchal society, relative peacefulness, and rampant sexual activity across a range of relations (e.g., de Waal 2013, 2019). Bonobo mothers frequently permit other group members to handle their infants; 96% of these interactions were positive in one study (Klaree et al. 2018). (Illustrated by Daphné Damoiseau-Malraux. © 2020 Rasmus Grønfeldt Winther)
Notes
- 1.
I first presented this thought experiment in Winther (2018). This work was completed in 2015.
- 2.
- 3.
Yu et al. (2004).
- 4.
Brown et al. (2004).
- 5.
- 6.
Yu et al. (2002).
- 7.
- 8.
Rosenberg (2011), p. 667.
- 9.
- 10.
- 11.
- 12.
- 13.
- 14.
- 15.
- 16.
- 17.
- 18.
- 19.
- 20.
A starting point for such a discussion might be Sandel (2020).
- 21.
I develop this point in detail in Rasmus Grønfeldt Winther. 2022a. Our Genes: A Philosophical Introduction to Human Evolutionary Genomics. Cambridge University Press.
- 22.
E.g., Enard and Petrov (2018). The divergence between Neanderthals and modern humans, Homo sapiens can be estimated. Mendez et al. (2016) use Y-chromosome DNA from a Neanderthal to estimate divergence time to be around 588,000 years ago; Langergraber et al. (2012) find the temporal range for the split to be 400–800,000 years ago; Gómez-Robles (2019) uses fossil teeth, not genetics, to place the split at no less than 800,000 years ago.
- 23.
Many central, south, and east Asian populations, as well as Indigenous Americans, have small amounts of Denisovan DNA (Jacobs et al. 2019). Typically, because of chromosomal recombination over many generations, the regions of the Neanderthal genome interspersed in our genome are less than a hundred kilobases long (Sankararaman et al. 2014). For further reviews and citations, see also Nielsen et al. (2017), Bergström et al. (2020), and Winther (2022a), especially chapter two.
- 24.
Chen et al. (2020).
- 25.
See, e.g., Mendez et al. (2016), p. 732. Or maybe any such haplotype lineages in Homo sapiens died out due to selection or chance (See Prüfer et al. 2014; Nielsen et al. 2017, https://humanorigins.si.edu/evidence/genetics/ancient-dna-and-neanderthals/interbreeding).
- 26.
Of course, it is possible that we have yet to discover a new Neanderthal Y-chromosome or mitochondrial DNA haplotype from archaic remains, perhaps present somewhere in a human population, either already sampled or not. (On the converse point of human introgression into the Neanderthal lineage, see, e.g., Raff 2017).
- 27.
References
Alcoff LM (2015) The future of whiteness. Polity, Cambridge, UK
Andreasen RO (2000) Race: biological reality or social construct? Philos Sci 67:S653–S666
Andreasen RO (2007) Biological conceptions of race. In: Matthen M, Stephens C (eds) Philosophy of biology. Elsevier, Amsterdam, pp 455–481
Ásta (2018) Categories we live by: the construction of sex, gender, race, and other social categories. Oxford University Press, New York
Auton A, et al. (2015) (The 1000 Genomes Project Consortium.). A global reference for human genetic variation. Nature 526(7571): 68–74. https://doi.org/10.1038/nature15393
Barbujani G, Magagni A, Minch E, Cavalli-Sforza LL (1997) An apportionment of human DNA diversity. Proc Natl Acad Sci USA 94:4516–4519
Bergström A, McCarthy SA, Hui R, Almarri MA, Ayub Q, et al. (2020) Insights into human genetic variation and population history from 929 diverse genomes. Science 367(6484):eaay5012. https://doi.org/10.1126/science.aay5012
Biddanda A, Rice DP, Novembre J (2020) A variant-centric perspective on geographic patterns of human allele frequency variation. eLife 9:e60107. https://doi.org/10.7554/eLife.60107
Brown GR, Gill GP, Kuntz RJ, Langley CH, Neale DB (2004) Nucleotide diversity and linkage disequilibrium in loblolly pine. Proc Natl Acad Sci USA 101:15255–15260
Campbell MC, Tishkoff SA (2008) African genetic diversity: implications for human demographic history, modern human origins, and complex disease mapping. Anny Rev Genomics Hum Genet 9:403–433
Chen L, Wolf AB, Wenqing F, Li L, Akey, JM (2020) Identifying and interpreting apparent Neanderthal ancestry in African individuals. Cell 180(4):677–687
Coulthard GS (2014) Red skin, white masks: rejecting the colonial politics of recognition. University of Minnesota Press, Minneapolis, MN
de Waal F (2013) The Bonobo and the atheist: in search of humanism among the primates. WW Norton, New York
de Waal F (2019) Mama’s last hug: animal emotions and what they tell us about ourselves. WW Norton, New York
Enard D, Petrov DA (2018) Evidence that RNA viruses drove adaptive introgression between Neanderthals and modern humans. Cell 175(2):360–371. https://doi.org/10.1016/j.cell.2018.08.034
Fanon F. (1952/2008) Black skin, white masks. (original: peau noire, masques blancs.) Grove Press, New York
Fanon F. (1961/2004) The wretched of the Earth. (original: les damnés de la terre.) Grove Press, New York
Fujimura JH, Bolnick DA, Rajagopalan R, Kaufman JS, Lewontin RC, et al. (2014) Clines without classes: how to make sense of human variation. Sociol Theory 32(3):208–227
Galeano E (1973) Open veins of Latin America: five centuries of the pillage of a continent. Monthly Review Press, New York
Glasgow J (2009) A theory of race. Routledge, New York
Gómez-Robles A (2019) Dental evolutionary rates and its implications for the Neanderthal–modern human divergence. Sci Adv 5(5):eaaw1268. https://doi.org/10.1126/sciadv.aaw1268
Hacking I (1999) The social construction of what? Harvard University Press, Cambridge, MA
Happe KE (2013) The material gene: gender, race, and heredity after the human genome project. New York University Press, New York
Hartl DL, Clark AG (1989) Principles of population genetics. Sinauer Associates, Sunderland, MA
Herrnstein RJ, Murray C (1995) The bell curve: intelligence and class structure in American life. The Free Press, New York
Hochman A (2013) Against the new racial naturalism. J Phil 110(6):331–351
Hochman A (2016) Race: deflate or pop? Stud Hist Philos Sci C 57:60–68
Hublin J-J (2013) The middle pleistocene record. On the origin of Neandertals, modern humans and others. In: Begun D (ed) A companion to paleoanthropology. Wiley-Blackwell, pp 517–537
Jacobs GS, Hudjashov G, Saag L, Kusuma P, Darusallam, CC, et al. (2019) Multiple deeply divergent denisovan ancestries in Papuans. Cell 177(4):1010–1021
Kaplan JM, Winther RG (2014) Realism, antirealism, and conventionalism about race. Philos Sci 81:1039–1052
Kendi IX (2019) How to be an antiracist. One World, New York
Klaree B, Frances W, Colin B, Audra M, Josh S (2018) Infant handling in bonobos (Pan paniscus): exploring functional hypotheses and the relationship to oxytocin. Physiol Behav 193(Pt A):154–166
Langergraber KE, Prüfer K, Rowney C, Boesch C, Crockford C, et al. (2012) Generation times in wild chimpanzees and gorillas suggest earlier divergence times in great ape and human evolution. Proc Nat Acad Sci (USA) 109(39):15716–15721. https://doi.org/10.1073/pnas.1211740109
Lawson Handley LJ, Manica A, Goudet J, Balloux F (2007) Going the distance: human population genetics in a clinal world. Trends Genet 23(9):432–439
Lewontin RC (1972) The apportionment of human diversity. In: Dobzhansky T, Hecht MK, Steere WC (eds) Evolutionary biology, vol. 6. Springer, New York, NY, pp 381–398
Li W-H, Sadler LA (1991) Low nucleotide diversity in man. Genetics 129:513–523
Mendez FL, Poznik GD, Castellano S, Bustamante CD (2016) The divergence of Neandertal and modern human Y chromosomes. Am J Hum Genet 98(4):728–734. https://doi.org/10.1016/j.ajhg.2016.02.023
Mills CW (1997) The racial contract. Cornell University Press, Ithaca, NY
Mills CW (1998) But what are you really? The metaphysics of race. In: Mills CW (ed) Blackness visible: essays on philosophy and race. Cornell University Press, Ithaca, NY, pp 41–66
Murray C (2020) Human diversity: the biology of gender, race, and class. Twelve, New York
Nielsen R, Akey JM, Jakobsson M, Pritchard JK, Tishkoff S, Willerslev E (2017) Tracing the peopling of the world through genomics. Nature 541(7637):302–310. https://doi.org/10.1038/nature21347
Prüfer K, Racimo F, Patterson N, Jay F, Sankararaman S, et al. (2014) The complete genome sequence of a Neanderthal from the Altai mountains. Nature 505:43–49
Raff J (2017) Did human women contribute to Neanderthal genomes over 200,000 years ago? The Guardian. Website. https://www.theguardian.com/science/2017/jul/18/did-human-women-contribute-to-neanderthal-genomes-over-200000-years-ago. Accessed June 2020
Ramachandran S, Deshpande O, Roseman CC, Rosenberg NA, Feldman MW, et al. (2005) Support from the relationship of genetic and geographic distance in human populations for a serial founder effect originating in Africa. Proc Natl Acad Sci USA 102(44):15942–15947
Reardon J (2005) Race to the finish: identity and governance in an age of genomics. Princeton University Press, Princeton, NJ
Rosenberg NA (2011) A population-genetic perspective on the similarities and differences among worldwide human populations. Hum Biol 83:659–684
Rosenberg NA, Pritchard JK, Weber JL, Cann HM, Kidd KK, et al. (2002) Genetic structure of human populations. Science 298(5602):2381–2385
Rosenberg NA (2018) Variance-partitioning and classification in human population genetics. In: Winther RG (ed) Phylogenetic inference, selection theory, and history of science: selected papers of AWF Edwards with commentaries. Cambridge University Press, Cambridge, pp 399–403
Sandel MJ (2020) The tyranny of merit: what’s become of the common good? Farrar, Straus and Giroux, New York
Sankararaman S, Mallick S, Dannemann M, Prüfer K, Kelso J, et al. (2014) The genomic landscape of Neanderthal ancestry in present-day humans. Nature 507:354–357. https://doi.org/10.1038/nature12961
Serre D, Pääbo S (2004) Evidence for gradients of human genetic diversity within and among continents. Genome Res 14(9):1679–1685
Sesardić N (2010) Race: a social destruction of a biological concept. Biol Philos 25(2):143–162
Sesardić N (2013) Confusions about race: a new installment. Stud Hist Philos Sci C 44:287–293
Spencer Q (2013) Biological theory and the metaphysics of race: a reply to Kaplan and Winther. Biol Theory 8:114–120
Spencer Q (2014) A radical solution to the race problem. Philos Sci 81:1025–1038
Spencer Q (2015) Philosophy of race meets population genetics. Stud Hist Philos Sci C 52:46–55
Spencer Q (2019) How to be a biological racial Realist. In: Glasgow J, Haslanger S, Jeffers C, Spencer Q (eds) What is race? Oxford University Press, New York, pp 73–110
TallBear K (2013) Native American DNA: tribal belonging and the false promise of genetic science. MN, University of Minnesota Press, Minneapolis
Templeton AR (1997) Out of Africa? What do genes tell us? Curr Opin Geneti Dev 7:841–847
Templeton AR (1999) Human races: a genetic and evolutionary perspective. Am Anthropol 100(3):632–650
Templeton AR (2002) Out of Africa again and again. Nature 416:45–51
Vaesen K, Scherjon F, Hemerik L, Verpoorte A (2019) Inbreeding, Allee effects and stochasticity might be sufficient to account for Neanderthal extinction. PLoS ONE 14(11):e0225117. https://doi.org/10.1371/journal.pone.0225117
Wall JD, Cox MP, Mendez FL, Woerner A, Severson T, et al. (2008) A novel DNA sequence database for analyzing human demographic history. Genome Res 18:1354–1361
Winther RG (2014) The genetic reification of ‘race’? A story of two mathematical methods. Crit Philos Race 2:204–223
Winther RG (2018) Race and biology. In: Taylor PC, Alcoff L, Anderson L (eds) The Routledge companion to the philosophy of race. Routledge, New York, pp 305–320
Winther RG (2019) A beginner’s guide to the new population genomics of Homo sapiens: origins, race, and medicine. Harvard Rev Philos 25:1–18
Winther RG (2020) When maps become the world. University of Chicago Press, Chicago, IL
Winther RG (2022a) Our genes: a philosophical perspective on human evolutionary genomics. Cambridge University Press, Cambridge
Winther RG (2022b) Lewontin (1972). In: Lorusso L, Winther RG (eds) Remapping race in a global context. Routledge, London, pp 9–47
Yu N, Chen F-C, Ota S, Jorde LB, Pamilo P, et al. (2002) Larger genetic differences within Africans than between Africans and Eurasians. Genetics 161:269–274
Yu N, Jensen-Seaman MI, Chemnick L, Ryder O, Li W-H (2004) Nucleotide diversity in Gorillas. Genetics 166:1375–1383
Zack N (2002) Philosophy of science and race. Routledge, New York
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Open Access This chapter is licensed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license and indicate if changes were made.
The images or other third party material in this chapter are included in the chapter's Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the chapter's Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
Copyright information
© 2022 The Author(s)
About this chapter

Cite this chapter
Winther, R.G. (2022). Race and Human Genomic Variation. In: Swartz, B., Mishler, B.D. (eds) Speciesism in Biology and Culture. Springer, Cham. https://doi.org/10.1007/978-3-030-99031-2_2
Download citation
DOI: https://doi.org/10.1007/978-3-030-99031-2_2
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-99029-9
Online ISBN: 978-3-030-99031-2
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)