
Abstract
In addition to cis-cleavage activity that recognizes and cleaves nucleic acid sequences, a trans-cleavage activity that indiscriminately and non-specifically cleaves single-stranded DNA or RNA has been discovered in some Cas proteins, including Cas12a and Cas13a. Various detection methods using this activity have been widely reported. Herein, we describe a new highly efficient DNA reporter (5′-TTATT-CCCCC-3′; TTATT-5C) that outperformed the existing AT-rich DNA reporter (5′-TTATT-3′) used in most Cas12a-based target nucleic detection assays. By systematically investigating the effect of DNA reporter length and sequence on the trans-cleavage activity of Cas12a, we achieved up to a 100-fold increase in fluorescence signal intensity derived from the trans-cleavage activity of Cas12a compared to that achieved using the existing AT-rich DNA reporter. The new DNA reporter was also applied, along with the existing AT-rich DNA reporter, for the detection of the Salmonella enterotoxin (stn) gene. Importantly, both detection speed and limit were significantly enhanced with the new DNA reporter. In addition, polymerase chain reaction (PCR) was adopted to the CRISR/Cas-Based system of the new DNA reporter, thereby confirming its practical applicability. The high-efficiency DNA reporter described herein can pave the way for further improving the trans-cleavage activity of other Cas proteins, as well as the sensitivity of CRISPR/Cas-Based systems.
Similar content being viewed by others

1 Introduction
Nucleic acid testing, one of the molecular diagnostic systems is essential for the detection of various microorganisms including bacteria, archaea, and viruses as well as cancer-associated biomarkers. The recent advent of SARS-CoV-2 and its variants have threatened humanity, incurring a high social cost and emphasized the importance of nucleic acid testing. Great research efforts have been devoted to detecting nucleic acids with high sensitivity and selectivity that could contribute to the effective control and prevention of viral transmission [1,2,3].
Notably, the clustered and regularly interspaced short palindromic repeats (CRISPR)/CRISPR-associated (Cas) protein that originated as a bacterial and archaeal defense mechanism has received special attention [4, 5]. Several Cas proteins exist due to the coevolution of host cells and foreign entities [6], among which Cas9 has attracted considerable attention for genome editing owing its ability to cleave specific DNA sequences [7, 8]. In principle, Cas9 binds to a complementary DNA sequence with the help of specificity determinant RNA molecules (named guide RNA), which then cleaves the complementary DNA sequence via its nucleolytic activity termed cis-cleavage activity [8, 9]. Recently, some Cas proteins, such as Cas12a and Cas13a, were found to have not only cis-cleavage activity but also trans-cleavage activity [10]. Trans-cleavage is a reaction that indiscriminately cuts a non-target, single-stranded DNA or RNA sequence, and is only activated after cis-cleavage [11]. For example, Cas12a is transformed into a non-specific endonuclease that indiscriminately cleaves the nearby single-stranded DNA sequences after the cis-cleavage onto single- or double-stranded target DNA sequence (named DNA activator) [12]. Similarly, Cas13a that completes the cis-cleavage onto a single-stranded RNA (named RNA activator) is transformed into a non-specific endonuclease capable of indiscriminately cleaving nearby single-stranded RNA sequences [13]. This intrinsic activity of Cas12a or Cas13a in combination with various nucleic acid amplification techniques has enabled the development of reliable nucleic acid detection systems, such as an one-hour low-cost multipurpose highly efficient system (HOLMES) [14], DNA Endonuclease Targeted CRISPR Trans Reporter (DETECTR) [10, 15] and Specific High-sensitivity Enzymatic Reporter unlocking (SHERLOCK) [16, 17]. As compared to traditional ones relying on nucleic acid amplification only [18,19,20], CRISPR/Cas-based diagnostic systems exhibit better sensitivity and selectivity due to the highly efficient catalytic activity of Cas proteins.
In general, a fluorescence resonance energy transfer (FRET)-based reporter, which comprises short DNA or RNA oligonucleotides labeled with a fluorophore and quencher, is used to monitor the trans-cleavage activity of Cas proteins [11, 21, 22]. A negligible fluorescence signal is initially observed due to the close proximity of the fluorophore and quencher, but a significantly increased fluorescence signal is generated after a FRET-based reporter is indiscriminately cleaved by Cas proteins [21, 22]. To date, many studies have been reported to improve the trans-cleavage activity of Cas proteins because it is closely related to the detection sensitivity. A representative example is that of the study by Nguyen et al., which showed that a 7-mer DNA extension at the 3′-end of the crRNA increased the trans-cleavage activity of Cas12a by approximately fivefold than that of the CRISPR RNA (crRNA) without the DNA extension [11]. In addition, the effects of salt concentration [23] and activator DNA with different modifications (such as phosphorothioate linkage [24]) on the Cas12a trans-cleavage activity have also been investigated.
In this study, we optimized the nucleotide sequence and length of the DNA reporter to maximize the trans-cleavage activity of Cas12a. After a thorough screening process, a high-efficiency DNA reporter (5′-TTATT-CCCCC-3′; TTATT-5C) that exceeded the performance of the existing AT-rich DNA reporter (5′-TTATT-3′) [25, 26] was discovered and successfully applied to the detection of the Salmonella enterotoxin (stn) gene. Consequently, the new DNA reporter described here was able to enhance the detection sensitivity by approximately tenfold compared to that achieved using the existing AT-rich DNA reporter. In addition, the CRISPR/Cas system was combined with PCR to demonstrate its practical applicability. We expect that this enhanced DNA reporter can be used as a core technology to improve the sensitivity of CRISPR/Cas-based systems.
2 Materials and Methods
2.1 Reagents and Materials
The oligonucleotide sequences used in this work (Table 1) were purchased from Bionics (Seoul, Korea) and Integrated DNA Technologies (Coralville, IA, USA). HiScribe™ T7 High Yield RNA Synthesis Kit, LbaCas12a, 10 × NEBuffer 2.1, DNase I, and DNase I reaction buffer were purchased from New England Biolabs (Ipswich, MA, USA). TOPreal™ SYBR Green qPCR PreMIX was purchased from Enzynomics (Daejeon, Korea).
2.2 Preparation of crRNA
The crRNAs extended with 11-mer DNA (No. 12 in Table 1) [27] were chemically synthesized by Integrated DNA Technologies. For obtaining crRNA to detect the stn gene, nuclease-free water (5.5 μL), 10 × reaction buffer (1.5 μL), NTPs (1.5 μL of each; final concentration: 7.5 mM), 3.6 μM IVT template DNA (5.5 μL; No. 14 in Table 1), and T7 RNA Polymerase Mix (1.5 μL) were mixed. The mixture was incubated at 37 °C for 16 h and then treated with DNase I to remove the remaining IVT template DNA; nuclease-free water (70 μL), 10 × DNase I buffer (10 mM Tris–HCl, 2.5 mM MgCl2, 0.5 mM CaCl2; pH 7.6, 25 °C) (10 μL), and 2 U/μL DNase I (2 μL) were added to 20 μL of the IVT product, and the mixture was incubated at 37 °C for 15 min. Finally, crRNA was purified using the Monarch RNA Cleanup Kit (New England Biolabs). The purity and concentration of the synthesized crRNA were evaluated using a Nanodrop Spectrometer (Spectramax iD5 multi-mode microplate reader; Molecular Devices, San Jose, CA, USA) [27].
2.3 Cas12a-Catalyzed Trans-Cleavage Reaction with Various DNA Reporters
First, nuclease-free water (11.8 μL), 10 × NEBuffer™ r2.1 (500 mM NaCl, 100 mM Tris–HCl, 100 mM MgCl2, 1000 μg/mL recombinant albumin; pH 7.9, 25 °C) (2 μL), 1 μM crRNA (0.4 μL), 2 μM various DNA reporters (5 μL), and 1 μM Cas12a (0.4 μL) were mixed and incubated at 25 °C for 10 min. Next, 100 nM single-stranded DNA activator (No. 13 in Table 1) or target DNA encoding stn gene (0.4 μL; No. 15 in Table 1) was added to the mixture, which was then incubated at 37 °C for 60 min in a CFX Connect Real-Time PCR Detection System (Bio-Rad, Hercules, CA, USA). The fluorescence signals were collected once every minute. All experiments were performed in triplicate.
2.4 Bacterial Cell Culture and gDNA Isolation
All bacterial strains (Salmonella enterica, Staphylococcus aureus, Klebsiella pneumoniae, Pseudomonas aeruginosa, Enterobacter cloacae) were cultured in Luria–Bertani (LB) medium (BD, Franklin Lakes, NJ, USA) with shaking (150 rpm) at 37 °C for 24 h. After the cultures were centrifuged at 5000g for 10 min to obtain cell pellets, the gDNA was then isolated using a Total DNA Extraction S&V kit (Bionics, Seoul, Korea) according to the manufacturer’s instructions. The concentrations of gDNA were evaluated using a Nanodrop instrument (Spectramax iD5 multi-mode microplate reader, Molecular Devices, San José, CA, USA), and the gDNA was stored at − 20 °C until use [28, 29].
2.5 Detection of Target gDNA Using CRISPR/Cas System in Combination with PCR
First, the samples for PCR reactions were prepared with nuclear-free water (7 μL), 5 μM primer mix (1 μL) (No. 16, 17 in Table 1) [30], different concentrations of gDNA (2 μL), and 2X TOPreal™ SYBR Green qPCR PreMIX (10 μL), which was then incubated in a CFX Connect Real-Time PCR Detection System (Bio-Rad) under the following conditions: initial denaturation at 95 °C for 10 min; followed by 60 cycles of denaturation at 95 °C for 10 s, annealing at 60 °C for 15 s, and extension at 72 °C for 30 s. Next. the PCR reaction products (0.4 μL) were added to the pre-incubated mixture at 25 °C for 10 min composed of nuclear-free water (11.8 μL), 10 × NEB buffer™ r2.1(2 μL), 1 μM crRNA (0.4 μL), 2 μM reporter (No. 6 in Table 1) (5 μL), and 1 μM Cas12a (0.4 μL), which was finally incubated at 37 °C for 60 min using the CFX Connect Real-Time PCR Detection System (Bio-Rad). The fluorescence signals were collected once every minute. All experiments were performed in triplicate.
3 Results and Discussion
3.1 Optimization of the DNA Reporter
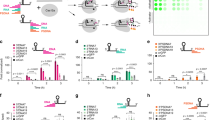
To maximize the Cas12a trans-cleavage activity, the effects of the length and sequence of the DNA reporter were investigated. First, DNA reporters of different lengths, such as TTATT-2 T, TTATT-5 T, TTATT-10 T, and TTATT-20 T (Table 1), were prepared based on the existing AT-rich DNA reporter (TTATT; 5-mer). As shown in Fig. 1A, a higher fluorescence signal was observed as the length of DNA reporters increased up to 15-mer (TTATT-10 T), which is indicative of the improved trans-cleavage activity. However, when the DNA reporter became longer (TTATT-20 T; 25-mer), the fluorescence signal was not as high as the one in 15-mer (TTATT-10 T), implying that TTATT-10 T is the most effective for the generation of the highest fluorescence signal after the Cas12a-catalyzed trans-cleavage reaction. Next, the nucleotide sequence included at the 3′-end of the existing DNA reporter (TTATT) was altered to investigate its effect on the trans-cleavage activity. It is worth noting that the nucleotide sequence at the 5′-end of the DNA reporter, where the fluorophore is placed, may have an effect on the fluorescence signal [31], making it difficult to precisely evaluate the effect of the nucleotide sequences on the trans-cleavage activity. Thus, we focused on the nucleotide sequences at the 3′-end of the DNA reporter. Specifically, DNA reporters were designed to contain an additional deoxycytidine (C), deoxyuridine (dU), deoxythymidine (T), deoxyadenosine (A), deoxyguanosine (G), or uridine (rU) at the 3′-end of the existing DNA reporter. Interestingly, different fluorescence signals were obtained depending on the nucleotide sequence, with TTATT-5C being the most effective in generating a high fluorescence signal (Fig. 1B). As expected, the trans-cleavage activity of Cas12a was not induced in the absence of single-stranded DNA activator (non-target control; NTC), which was demonstrated by the negligible fluorescence signal detected. In addition, when comparing the output of TTATT and TTATT-5rU, we observed that the signal was stronger with TTATT-5rU than with TTATT, which is very interesting considering Cas12a is known to exert its effective trans-cleavage activity on single-stranded DNA, but not on RNA [32]. Thus, we assumed that the extension of the DNA reporter even with RNA sequences can effectively enhance the trans-cleavage activity of Cas12a.

Fluorescence signals generated by the Cas12a-catalyzed trans-cleavage reaction with various DNA reporters. A Effect of the DNA reporter length. B Effect of the DNA reporter sequence. C Comparison of TTATT-5C with 10C. The fluorescence signal was calculated by subtracting the initial one from the one measured at 5 min after the Cas12a-catalyzed trans-cleavage reaction. NTC (Blue) and DNA activator (Red) represent the non-target control, and the presence of DNA activator (2 nM), respectively. Signal-to-noise ratio, which is shown above the bar graph, was calculated by dividing the fluorescence signal measured in the presence of the single-stranded DNA activator by one of the NTC. In all cases (except for 10C), the fluorescence signals of NTC were close to 0
Based on the excellent efficiency of TTATT-5C demonstrated in Fig. 1B, we conducted an experiment wherein the TTATT of TTATT-5C was changed to C (10C; Table 1) while maintaining the length of the DNA reporter. In the presence of the single-stranded DNA activator, the fluorescence signal of 10C was higher than that of TTATT-5C; however, the background fluorescence signal of 10C in the absence of single-stranded DNA activator (NTC) increased more significantly than that of TTATT-5C (Fig. 1C), revealing that the signal-to-noise ratio of TTATT-5C was approximately 9-times higher than that of 10C. Taken together, the above-described screening approach revealed TTATT-5C as a high-efficiency DNA reporter, with a 100-fold stronger fluorescence signal compared to that of the existing DNA reporter (TTATT). We assumed that the degree of Cas12a-catalyzed trans-cleavage reaction depends on the length and nucleotide sequence of the DNA reporter that can affect the interaction chance for Cas12a with the DNA reporter.
3.2 Detection of Target DNA Using CRISPR/Cas System Equipped with the New DNA Reporter
The new DNA reporter (TTATT-5C, Fig. 1) was further tested by assessing its ability to detect target DNA, which was compared to the existing DNA reporter (TTATT). As a proof-of-concept, we chose the Salmonella enterotoxin (stn) gene as the target DNA and accordingly designed crRNA for its detection. As shown in Fig. 2A, the fluorescence signals in the case of the new DNA reporter (TTATT-5C) significantly increased as the CRISPR/Cas reaction proceeded. In contrast, the fluorescence signal ineffectively increased in the case of the existing DNA reporter (TTATT) (Fig. 2B). These results suggest that the DNA reporter engineered here can significantly improve the detection performance by reducing the reaction time. In addition, the limit of detection (LOD) was estimated by performing the experiments at different concentrations of target DNA (5–500 pM). As illustrated in Fig. 2C, D, the TTATT reporter had a LOD of 200 pM, whereas the TTATT-5C reporter had a LOD of 20 pM. Consequently, the newly discovered DNA reporter enhanced the detection sensitivity approximately 10 times than that of the existing DNA reporter.

Detection of the target DNA using CRISPR/Cas system with two different DNA reporters. Time-dependent fluorescence intensities of the (A) new DNA reporter (TTATT-5C) and (B) existing DNA reporter (TTATT) during CRISPR/Cas-catalyzed trans-cleavage reaction in the presence of different concentrations of the target DNA (5–500 pM). Fluorescence signals of the (C) new DNA reporter (TTATT-5C) and (D) existing DNA reporter (TTATT) calculated by subtracting the initial one from the one measured at 10 min after the CRISPR/Cas reaction in the presence of different concentrations of the target DNA. The cut-off line (red line) was set according to the following equation: NTC (non-target control) + 3 × standard deviation (SD) of the NTC sample
3.3 Detection of Target gDNA Using CRISPR/Cas System in Combination with PCR
To examine the practical applicability of the new DNA reporter (TTATT-5C), PCR was adopted as the pre-amplification step before the CRISPR/Cas-catalyzed reaction, and the experiment was performed by targeting gDNA sequences encoding stn gene. As illustrated in Fig. 3, the first step is the PCR amplification using the DNA primers specific to stn gene, where the DNA primers are rationally designed for PCR product to contain PAM sequence (5ʹ-TTTV-3ʹ) such that Cas12a can be activated to exert the effective trans-cleavage activity [24]. The second step is the Cas12a-catalyzed reaction, where crRNA mediates Cas12a to recognize the specific sequences of PCR product, and enables the indiscriminate, trans-cleavage reactions after the cis-cleavage, thereby leading to the generation of high fluorescence signal. Since the proposed system relies on PCR amplification in combination with the new DNA reporter endowing the effective trans-cleavage activity of Cas12a, the highly sensitive detection of target nucleic acids can be achieved.

Schematic of the target DNA detection strategy using CRISPR/Cas in combination with polymerase chain reaction (PCR)
The results in Fig. 4A, B show that the fluorescence signals in the presence of target gDNA extracted from Salmonella enterica (SE) at different concentrations from 100 zM to 100 fM were clearly distinguished from the non-target control (NTC). Importantly, an LOD of 100 zM that corresponds to 1.2 copies per reaction was achieved by the proposed system using the new DNA reporter. Moreover, it was confirmed that the presence of non-target gDNA from different bacteria such as Staphylococcus aureus (SA), Klebsiella pneumoniae (KP), Pseudomonas aeruginosa (PA), Enterobacter cloacae (EC) generated the negligible, fluorescence signal, which was significantly lower than that in the presence of target gDNA from SE (Fig. 4C). Importantly, the high sensitivity and selectivity of the proposed system was clearly demonstrated by the comparison with PCR only where non-specific amplification curves were observed in the presence of non-target gDNA (Fig. S1).

Detection of target gDNA using CRISPR/Cas system in combination with PCR. A Time-dependent fluorescence intensities of the new DNA reporter (TTATT-5C) during CRISPR/Cas-catalyzed trans-cleavage reaction in the presence of different concentrations of the target gDNA (100 zM-100 fM). B Fluorescence signals of the new DNA reporter calculated by subtracting the initial one from the one measured at 2 min after the CRISPR/Cas reaction in the presence of different concentrations of the target gDNA (100 zM-100 fM). The cut-off line (red line) was set according to the following equation: NTC + 3 × SD of NTC sample. NTC and SD indicate non-target control and standard deviation, respectively. C Fluorescence signals of the new DNA reporter calculated by subtracting the initial one from the one measured at 2 min after the CRISPR/Cas reaction in the presence of various gDNAs (1 fM) (SE Salmonella enterica, SA Staphylococcus aureus, KP Klebsiella pneumoniae, PA Pseudomonas aeruginosa, EC Enterobacter cloacae)
4 Conclusion
In this study, we engineered a high-efficiency DNA reporter (TTATT-5C) that can significantly enhance the trans-cleavage activity of Cas12a. This new DNA reporter was also confirmed to improve the detection sensitivity of the CRISPR/Cas system by approximately tenfold than that of the existing TTATT reporter. In addition, the practical applicability of the new DNA reporter was demonstrated by adopting PCR as a pre-amplification step. However, more structural, and biochemical characterization of Cas12a interacting with DNA reporters is necessary to understand the mechanism for the enhancement effect. Furthermore, the application of the new reporter for other types of Cas proteins such as Cas13a and Cas14a as well as the construction of CRISPR/Cas-based diagnostic systems relying on the isothermal amplification technologies remain to be investigated in future studies. Nonetheless, the strategy used here to improve the detection performance of the CRISPR/Cas system is simple and cost-effective, as it does not require the addition of any substances or physicochemical treatment other than the extension of the DNA reporter. Hence, this new DNA reporter may be used as a core technology to improve the trans-cleavage activity of other Cas proteins, as well as for enhancing the detection performance of CRISPR/Cas-based systems.
Data availability
The data that support the findings of this study are available from the corresponding author upon reasonable request.
References
El-Said, W.A., Al-Bogami, A.S., Alshitari, W., El-Hady, D.A., Saleh, T.S., El-Mokhtar, M.A., Choi, J.W.: Electrochemical microbiosensor for detecting COVID-19 in a patient sample based on gold microcuboids pattern. Biochip J. 15, 287–295 (2021). https://doi.org/10.1007/s13206-021-00030-3
Kim, H., Huh, H.J., Park, E., Chung, D.-R., Kang, M.: Multiplex molecular point-of-care test for syndromic infectious diseases. Biochip J. 15, 14–22 (2021). https://doi.org/10.1007/s13206-021-00004-5
Yoon, T., Shin, J., Choi, H.J., Park, K.S.: Split T7 promoter-based isothermal transcription amplification for one-step fluorescence detection of SARS-CoV-2 and emerging variants. Biosens. Bioelectron. 208, 114221 (2022). https://doi.org/10.1016/J.BIOS.2022.114221
Deshpande, K., Vyas, A., Balakrishnan, A., Vyas, D.: Clustered regularly interspaced short palindromic repeats/Cas9 genetic engineering: robotic genetic surgery. Am. J. Robotic. Surg. 2, 49–52 (2015). https://doi.org/10.1166/ajrs.2015.1023
Al-Attar, S., Westra, E.R., van der Oost, J., Brouns, S.J.J.: Clustered regularly interspaced short palindromic repeats (CRISPRs): the hallmark of an ingenious antiviral defense mechanism in prokaryotes. Biol. Chem. 392, 277–289 (2011). https://doi.org/10.1515/bc.2011.042
Knott, G.J., Cress, B.F., Liu, J.-J., Thornton, B.W., Lew, R.J., Al-Shayeb, B., Rosenberg, D.J., Hammel, M., Adler, B.A., Lobba, M.J., Xu, M., Arkin, A.P., Fellmann, C., Doudna, J.A.: Structural basis for AcrVA4 inhibition of specific CRISPR-Cas12a. Elife (2019). https://doi.org/10.7554/eLife.49110
Li, H., Yang, Y., Hong, W., Huang, M., Wu, M., Zhao, X.: Applications of genome editing technology in the targeted therapy of human diseases: mechanisms, advances and prospects. Signal Transduct. Target. Ther. 5, 1 (2020). https://doi.org/10.1038/s41392-019-0089-y
Ran, F.A., Hsu, P.D., Wright, J., Agarwala, V., Scott, D.A., Zhang, F.: Genome engineering using the CRISPR-Cas9 system. Nat. Protoc. 8, 2281–2308 (2013). https://doi.org/10.1038/nprot.2013.143
Wu, X., Kriz, A.J., Sharp, P.A.: Target specificity of the CRISPR-Cas9 system. Quant. Biol. 2, 59–70 (2014). https://doi.org/10.1007/s40484-014-0030-x
Chen, J.S., Ma, E., Harrington, L.B., Da Costa, M., Tian, X., Palefsky, J.M., Doudna, J.A.: CRISPR-Cas12a target binding unleashes indiscriminate single-stranded DNase activity. Science 360, 436–439 (2018). https://doi.org/10.1126/science.aar6245
Nguyen, L.T., Smith, B.M., Jain, P.K.: Enhancement of trans-cleavage activity of Cas12a with engineered crRNA enables amplified nucleic acid detection. Nat. Commun. 11, 1–13 (2020). https://doi.org/10.1038/s41467-020-18615-1
Li, S.-Y., Cheng, Q.-X., Liu, J.-K., Nie, X.-Q., Zhao, G.-P., Wang, J.: CRISPR-Cas12a has both cis- and trans-cleavage activities on single-stranded DNA. Cell Res. 28, 491–493 (2018). https://doi.org/10.1038/s41422-018-0022-x
Meeske, A.J., Nakandakari-Higa, S., Marraffini, L.A.: Cas13-induced cellular dormancy prevents the rise of CRISPR-resistant bacteriophage. Nature 570, 241–245 (2019). https://doi.org/10.1038/s41586-019-1257-5
Li, S.-Y., Cheng, Q.-X., Wang, J.-M., Li, X.-Y., Zhang, Z.-L., Gao, S., Cao, R.-B., Zhao, G.-P., Wang, J.: CRISPR-Cas12a-assisted nucleic acid detection. Cell Discov. 4, 20 (2018). https://doi.org/10.1038/s41421-018-0028-z
Broughton, J.P., Deng, X., Yu, G., Fasching, C.L., Servellita, V., Singh, J., Miao, X., Streithorst, J.A., Granados, A., Sotomayor-Gonzalez, A., Zorn, K., Gopez, A., Hsu, E., Gu, W., Miller, S., Pan, C.-Y., Guevara, H., Wadford, D.A., Chen, J.S., Chiu, C.Y.: CRISPR-Cas12-based detection of SARS-CoV-2. Nat. Biotechnol. 38, 870–874 (2020). https://doi.org/10.1038/s41587-020-0513-4
Gootenberg, J.S., Abudayyeh, O.O., Lee, J.W., Essletzbichler, P., Dy, A.J., Joung, J., Verdine, V., Donghia, N., Daringer, N.M., Freije, C.A., Myhrvold, C., Bhattacharyya, R.P., Livny, J., Regev, A., Koonin, E.V., Hung, D.T., Sabeti, P.C., Collins, J.J., Zhang, F.: Nucleic acid detection with CRISPR-Cas13a/C2c2. Science 356, 438–442 (2017). https://doi.org/10.1126/science.aam9321
Kellner, M.J., Koob, J.G., Gootenberg, J.S., Abudayyeh, O.O., Zhang, F.: SHERLOCK: nucleic acid detection with CRISPR nucleases. Nat. Protoc. 14, 2986–3012 (2019). https://doi.org/10.1038/s41596-019-0210-2
Lee, J.S., Ahn, J.J., Kim, S.J., Yu, S.Y., Koh, E.J., Kim, S.H., Sung, H.S., Huh, J.W., Hwang, S.Y.: POCT detection of 14 respiratory viruses using multiplex RT-PCR. Biochip J. 15, 371–380 (2021). https://doi.org/10.1007/S13206-021-00037-W/FIGURES/2
Kim, J., Hwang, E.S.: Multiplexed diagnosis of four serotypes of dengue virus by real-time RT-PCR. Biochip J. 14, 421–428 (2020). https://doi.org/10.1007/S13206-020-4409-7/TABLES/5
Kim, J., Jung, C.: SF-qPCR: strand displacement-based fast quantitative polymerase chain reaction. Biochip J. 16, 41–48 (2022). https://doi.org/10.1007/S13206-021-00044-X/FIGURES/6
Bogers, J.F.M., Berghuis, N.F., Busker, R.W., van Booma, A., Paauw, A., van Leeuwen, H.C.: Bright fluorescent nucleic acid detection with CRISPR-Cas12a and poly(thymine) templated copper nanoparticles. Biol. Methods Protoc. (2021). https://doi.org/10.1093/biomethods/bpaa020
Xiong, Y., Zhang, J., Yang, Z., Mou, Q., Ma, Y., Xiong, Y., Lu, Y.: Functional DNA regulated CRISPR-Cas12a sensors for point-of-care diagnostics of non-nucleic-acid targets. J. Am. Chem. Soc. 142, 207–213 (2020). https://doi.org/10.1021/jacs.9b09211
Fuchs, R.T., Curcuru, J., Mabuchi, M., Yourik, P., Robb, G.B.: Cas12a trans-cleavage can be modulated in vitro and is active on ssDNA, dsDNA, and RNA. bioRxiv. (2019). https://doi.org/10.1101/600890
Swarts, D.C., Jinek, M.: Mechanistic Insights into the cis- and trans-acting DNase activities of Cas12a. Mol. Cell. 73, 589-600.e4 (2019). https://doi.org/10.1016/j.molcel.2018.11.021
Ding, X., Yin, K., Li, Z., Lalla, R.V., Ballesteros, E., Sfeir, M.M., Liu, C.: Ultrasensitive and visual detection of SARS-CoV-2 using all-in-one dual CRISPR-Cas12a assay. Nat. Commun. 11, 4711 (2020). https://doi.org/10.1038/s41467-020-18575-6
Zhang, M., Wang, H., Wang, H., Wang, F., Li, Z.: CRISPR/Cas12a-assisted ligation-initiated loop-mediated isothermal amplification (CAL-LAMP) for highly specific detection of microRNAs. Anal. Chem. 93, 7942–7948 (2021). https://doi.org/10.1021/acs.analchem.1c00686
Han, J., Park, J.S., Kim, S., Cha, B.S., Lee, E.S., Kim, J.H., Kim, S., Shin, J., Jang, Y., Chowdhury, P., Park, K.S.: Modulation of CRISPR/Cas12a trans-cleavage activity by various DNA-modifying enzymes. Microchem. J. 180, 107606 (2022). https://doi.org/10.1016/j.microc.2022.107606
Kim, S., Kim, J.H., Kim, S., Park, J.S., Cha, B.S., Lee, E.S., Han, J., Shin, J., Jang, Y., Park, K.S.: Loop-mediated isothermal amplification-based nucleic acid lateral flow assay for the specific and multiplex detection of genetic markers. Anal. Chim. Acta. 1205, 339781 (2022). https://doi.org/10.1016/j.aca.2022.339781
Shin, J., Yoon, T., Park, J., Park, K.S.: Sensitive and simultaneous detection of hygiene indicator bacteria using an enhanced CRISPR/Cas system in combination with a portable fluorescence detector. Sens. Actuators B Chem. 365, 131871 (2022). https://doi.org/10.1016/j.snb.2022.131871
Moore, M.M., Feist, M.D.: Real-time PCR method for Salmonella spp. targeting the stn gene. J. Appl. Microbiol. 102, 516–530 (2007). https://doi.org/10.1111/j.1365-2672.2006.03079.x
Mao, H., Luo, G., Zhan, Y., Zhang, J., Yao, S., Yu, Y.: The mechanism and regularity of quenching the effect of bases on fluorophores: The base-quenched probe method. Analyst. 143, 3292–3301 (2018). https://doi.org/10.1039/c8an00116b
Nalefski, E.A., Patel, N., Leung, P.J.Y., Islam, Z., Kooistra, R.M., Parikh, I., Marion, E., Knott, G.J., Doudna, J.A., Le Ny, A.L.M., Madan, D.: Kinetic analysis of Cas12a and Cas13a RNA-guided nucleases for development of improved CRISPR-based diagnostics. iScience 24, 102996 (2021). https://doi.org/10.1016/j.isci.2021.102996
Acknowledgements
This work was supported by a grant from the National Research Foundation of Korea (NRF), funded by the Korean government (Ministry of Science and ICT) (No. NRF-2020R1C1C1012275), by a grant (21163MFDS501) from the Ministry of Food and Drug Safety in 2021, by the LCA Experts Training Graduate Program for Knowledge-based Environmental Service from the Ministry of Environment, Korea and by Konkuk University's research support program for its faculty on sabbatical leave in 2022.
Author information
Authors and Affiliations
Contributions
SL: Conceptualization, Methodology, Validation, Formal analysis, Writing-original draft. DN: Methodology, Validation, Formal analysis. JSP: Conceptualization, Methodology, Validation, Writing-original draft. SK: Methodology, Formal analysis. ESL: Methodology, Formal analysis. BSC: Methodology, Formal analysis. KSP: Conceptualization, Methodology, Writing-review & editing, Project administration.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Lee, S., Nam, D., Park, J.S. et al. Highly Efficient DNA Reporter for CRISPR/Cas12a-Based Specific and Sensitive Biosensor. BioChip J 16, 463–470 (2022). https://doi.org/10.1007/s13206-022-00081-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13206-022-00081-0