
Abstract
Cervical cancer is the fourth most frequent cancer among women worldwide. Human papillomaviruses (HPVs) cause almost all cervical cancers in low-income countries. Three prophylactic HPV virus-like particle-based vaccines have been licensed to date, and they have all shown high efficacy and reliable safety profiles. However, isolated safety issues have resulted in a reluctance to use these vaccinations. In addition, the high prices of the vaccinations have caused the inequitable distribution of the vaccine: the prices are unaffordable for low-income countries. Meanwhile, great effort has been put into the development of therapeutic HPV vaccines, including protein/peptide-, live vector-, DNA- and cell-based vaccines. These new vaccines have considerable therapeutic potential but limited practical use. The development of immune checkpoint inhibitors and personalized immunotherapy remain challenges for future study. In this article, the current status of the licensed vaccines, therapeutic HPV vaccines and biosimilars, and new platforms for HPV vaccines, are reviewed, and safety issues related to the licensed vaccines are discussed. In addition, the prospects for HPV vaccines are considered.
Similar content being viewed by others


Introduction
Recent studies estimate that 23% of all human malignancies are caused by infectious agents, such as hepatitis B and C viruses (liver cancer), human papillomaviruses (HPVs) (cervical and anogenital cancers), and Helicobacter pylori (stomach cancer) (WHO 2003; zur Hausen 2009). Moreover HPVs are responsible for 27.9–30% of all such malignancies worldwide (zur Hausen 2009; Bravo et al. 2010). HPVs can be isolated from skin swabs and hair of normal immunocompetent individuals (Antonsson et al. 2000, 2003), and there is strong evidence that HPVs cause cancers of the cervix uteri, penis, vulva, vagina, anus, and the head and neck in both men and women (Depuydt et al. 2016). HPV is the most common sexually transmitted infectious agent, and all sexually active individuals are likely to acquire HPV at some point during their life (Dunne et al. 2007). Around 291 million women worldwide are carriers of HPV DNA, and this corresponds to a prevalence of 10.4% (16.9% in women younger than 25 years) (de Sanjose et al. 2007). Worldwide, 527,624 women are diagnosed with cervical cancer and 265,672 die from the disease annually (Bruni et al. 2016). Cervical cancer is the fourth most frequent cancer among women and the second most common cancer in women aged 15–44 years (Bruni et al. 2016). In this article, the current status of prophylactic and therapeutic HPV vaccines are reviewed, and prospects for future vaccine development are discussed.
HPV biology and malignant transformation
HPVs are small non-enveloped viruses, 50–60 nm in diameter, packaging an approximately 8 kb circular double-stranded DNA genome (Doorbar et al. 2015; Araldi et al. 2017). The capsid of HPV is composed of major capsid protein L1 and minor capsid protein L2, and structural mapping shows that the T = 7 icosahedral capsid of HPV includes 360 copies of L1 protein arranged into 72 pentamers (Schiller and Lowy 2012). The L2 protein is believed to be located in the center of the pentamers, as the L2:L1 ratio is estimated to be 1:5–1:10 (Schellenbacher et al. 2017). All papillomaviruses contain well-conserved core genes for replication (E1 and E2) and capsid formation (L1 and L2), and genes for driving the cell cycle, immune evasion and virus release (E6, E7, E5 and E4) (Doorbar et al. 2015). Assembly occurs in association with cellular histones or nuclear factors (Cerqueira et al. 2016; Kim et al. 2016). HPVs infect only epithelial cells and replication is strictly controlled by the differentiation state of the epithelial cells (Cerqueira and Schiller 2017). It is well known that the E6 and E7 proteins target p53 and pRB to promote cell proliferation and prolong cell cycle progression, and the resulting genomic instability of host cells drives malignant transformation, eventually leading to invasive cervical cancer (Araldi et al. 2017). The integration of HPV genomes into the host significantly affects the functions of both viral and host genome; it causes disruption of the E1, E2, E5 and L2 gene regions, and consequently leads to overexpression of E6 and E7 proteins (Senapati et al. 2016).
During natural HPV infection, the virus is restricted to the intraepithelial layer of the mucosa and induces only a weak immune response. The antibody response against natural HPV infection is slow and elicits low antibody titers (WHO 2014). It is disputed whether the antibody response induced by natural HPV infection can reduce the risk of re-infection (Gravitt 2011). Approximately 70–90% of HPV infections are asymptomatic and resolve spontaneously within 1–2 years (Cubie 2013).
Cervical cancers are classified into squamous cell carcinomas, adenocarcinomas and adenosquamous carcinomas, with almost 90% of cervical cancers classified as squamous cell carcinomas (Liu et al. 2001). Cervical cancer develops from a precancerous lesion, known as a cervical intraepithelial neoplasia (CIN), which is classified by histopathological grade (CIN 1, CIN 2 or CIN 3) (Senapati et al. 2016).
Prevalence of HPV
More than 100 types of HPV have been identified, and more than 40 of these are transmissible through the genital area (Watson 2005). HPVs can be grouped by their malignant properties as low-risk and high-risk types. The low-risk types include HPV6, HPV11, HPV40, HPV42, HPV43, HPV44, HPV55, and the high-risk types include HPV16, HPV18, HPV31, HPV33, HPV35, HPV39, HPV45, HPV51, HPV52, HPV56, HPV58 and HPV59 (Bouvard et al. 2009). High-risk types are found in almost all patients with of cervical cancer: HPV16 and HPV18 are responsible for 70% of all cervical cancers, and HPV31, HPV33, HPV35, HPV45, HPV52 and HPV58 account for an additional 22% worldwide (Bruni et al. 2016). HPVs are also found in over 10% of women with normal cytology (Bruni et al. 2016), indicating that HPVs are circulating in healthy individuals with high frequency. The risk of squamous cell carcinoma in the cervix is known to be about 400- and 250-times higher in women infected with HPV16 and HPV18, respectively, compared to healthy individuals (WHO 2014). In the United States, 87% of anal cancers and 63% oropharyngeal cancer are caused by HPV infections (Zandberg et al. 2013): HPV16 and HPV18 cause 90% of HPV-positive anal cancers and 96% of HPV-positive oropharyngeal cancers (Daling and Sherman 1992; Gillison 2008). HPV6 and HPV11 cause 90% of all cases of genital warts (Braaten and Laufer 2008). The prevalence of HPV types in invasive cervical cancer is, in descending order, HPV16 (55.1%), HPV18 (14.3%), HPV45 (5%), HPV33 (4.2%), HPV58 (3.9%), HPV31 (3.5%), HPV52 (3.5%), HPV35 (1.7), HPV39 (1.5%) and HPV59 (1.4%) (Bruni et al. 2016). For this reason, HPV16 and HPV18 are the main targets for vaccine development.
Almost 85% of cervical cancers occur in low-income countries; the burden of cervical cancer is high in sub-Saharan Africa, Latin America and India. In addition, it is assumed that Mongolia and China have a high burden of cervical cancer, although the disease burden is unreported and therefore uncertain (Crosbie et al. 2013; Bruni et al. 2016).
Licensed prophylactic HPV vaccines
Virus-like particle (VLP)-based prophylactic HPV vaccines were developed incrementally by many contributors (McNeil 2006). The technical innovation of the prophylactic HPV vaccine is the application of VLPs as the vaccine antigens: the HPV vaccine is the second VLP-based vaccine to be commercialized, after the hepatitis B virus vaccine. Recombinant L1 protein is a subunit of the VLP, and the recombinant L1 protein can self-assemble into VLPs (Buck et al. 2013). The repetitive epitopes displayed on the surface of VLPs mimic the properties of the native HPV virion, and are able to elicit anti-pathogen-specific neutralizing antibodies. These particulates are then recognized and processed by antigen-presenting cells (APCs) (Kim and Kim 2016).
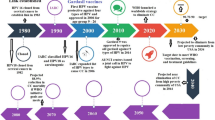
Three prophylactic VLP-based HPV vaccines have been commercialized and are currently available. Gardasil-4 (Merck Sharp & Dohme, MSD) is produced in Saccharomyces cerevisiae (S. cerevisiae) and received first approval in 2006 (Markowitz et al. 2007); Cervarix (GlaxoSmithKline, GSK) is produced in insect cells and earned first approval in 2007 (Monie et al. 2008). In 2014, the United States Food and Drug Administration (FDA) approved Gardasil-9 (MSD) which is again produced in the S. cerevisiae expression system (Joura et al. 2015; Petrosky et al. 2015). The properties and formulations of these vaccines are summarized in Table 1. Cervarix is a bivalent vaccine that contains HPV16 and HPV18 L1 VLPs to prevent cervical cancer. Gardasil-4 is a quadrivalent vaccine that contains HPV16 and HPV18 L1 VLPs to prevent cervical cancer as well as HPV6 and HPV11 L1 VLPs to prevent genital warts, and was the first quadrivalent vaccine to be licensed for the prevention of cervical cancers. Gardasil-9 is a nonavalent vaccine that contains the same four L1 VLPs as Gardisil-4 and, in addition, contains HPV31, 33, 45, 52 and 58 L1 VLPs (Corp. 2016; Harper and DeMars 2017). Recent studies show that these three commercial vaccines have similar effectiveness in preventing cervical cancer (WHO 2017).
Vaccination schedules
Gardasil-4 and Gardasil-9 are licensed for use in male and female children, adolescents and young adults aged 9–26 years old (FDA 2006, 2014a), and Cervarix is licensed for use in female children, adolescents and young adults aged 9–25 years old (FDA 2014b). The vaccines were originally licensed using a three-dose vaccination schedule. In 2006 the Advisory Committee on Immunization Practice recommended routine vaccination for preventing HPV infection at age 11 or 12 years for female adolescents, and in 2011 this was extended to include male adolescents (Meites et al. 2016). A two-dose regimen for HPV vaccination has been proposed because the dosing schedule could potentially enable the vaccination of 50% more people for the same price, compared to a three-dose regimen (Kreimer et al. 2011). In December 2016, the United State Centers for Disease Control and Prevention (CDC) recommended two-dose vaccination for male and female adolescents between the ages of 9 and 14 years, with the doses given at least 6 months apart (Meites et al. 2016). Several randomized studies of Cervarix, Gardarsil-4 and Gardasil-9 confirmed that the two-dose regimen in female adolescents aged 9–14 years was either non-inferior or inconclusive in terms of immunogenicity, compared to the three-dose regimen (Dobson et al. 2013; Puthanakit et al. 2016; Romanowski et al. 2016; Corp. 2017). The CDC recommends that adolescents and young adults who receive the vaccine between the ages of 15 and 26 years and immunocompromised individuals should continue to receive the original three-dose regimen (Meites et al. 2016).
Immune responses and efficacy
The antibody responses elicited by vaccination with Cervarix, Gardasil-4 and Gardasil-9 are significantly stronger than those elicited by natural HPV infections. Antibody titers were sustained for at least 10, 9.9 and 5 years following vaccination with Cervarix, Gardasil-4 and Gardasil-9, respectively, when female children and adolescents aged 9–15 years received the three-dose regimen (WHO 2017), although assessments are ongoing. In clinical trials, antibody titers peaked 4 weeks after the third vaccination, declined within the first year and stabilized thereafter (Mishra et al. 2015). Seropositivity rates 94 months after receiving three doses of Gardasil-4 were 94.3, 89.4, 99.5 and 88.8% total IgGs for HPV6, HPV11, HPV16 and HPV18, respectively (Ferris et al. 2014). Following vaccination with Cervarix the antibody titers were sustained for at least 8.4 years, with seropositivity for HPV16 and HPV18 at 100% (Schiller et al. 2012). A head-to-head trial comparing the immunogenicity of Cervarix and Gardasil-4 reported that Cervarix elicited 3.7- and 7.3-fold higher anti-HPV16 and 18 antibody levels, respectively, compared to Gardasil-4, in women aged 18–26 years, 7 months after the initial vaccination (Einstein et al. 2009), although the clinical relevance of this difference in antibody titer is unclear.
Gardasil-4 is reported to have 43.0% efficacy (95% confidence interval [CI] 13.0–63.2) against the development of CIN 3, irrespective of HPV type, in an intention-to-treat (ITT)-naïve analysis, and 16.4% efficacy (95% CI 0.4–30.0) in an ITT analysis (WHO 2014). Gardasil-4 conferred almost 100% protection against genital warts associated with HPV6 and HPV11 among HPV-naïve individuals (Munoz et al. 2010); the efficacy reduced 83% of all anogenital warts. Irrespective of HPV type, the efficacy of Cervarix against developing CIN 3 was 93.2% (95% CI 78.9–98.7) in the total vaccinated cohort (TVC)-naïve analysis, and 45.6% efficacy against developing CIN 3 (95% CI 28.8–58.7) in the TVC analysis (WHO 2014).
Cross protection
Phylogenetically, HPV16 is related to HPV31 and HPV33, and HPV18 is related to HPV45. These related types share considerable amino acid sequence in the major capsid protein L1 (Draper et al. 2011; Kemp et al. 2011). Cervarix and Gardasil-4 are therefore able to exert cross protection against HPV types that are not included in the vaccines; specifically cross protection against HPV31, HPV33 and HPV45 (Malagon et al. 2012). Cervarix conferred higher cross protection for HPV31, HPV33 and HPV45 than Gardasil-4 (Draper et al. 2013). Gardasil-9 was designed to provide direct protection against HPV6, HPV11, HPV16, HPV18, HPV31, HPV33 and HPV45 as well as HPV52 and HPV58, but exactly how much protection against HPV infection is provided by this nonavalent vaccine requires further study.
HPV6 and HPV11 are not phylogenetically related to HPV16 or HPV18. The PATRICIA trial, however, reported 35% efficacy of Cervarix against persistent (6-month) infections by HPV6 and HPV11 (Szarewski et al. 2013), although a study by Woestenberg and colleagues (2017) reported no cross protection by Cervarix against HPV6 and HPV11 infections, or against the development of anogenital warts.
Isolated safety issues
There have been isolated reports regarding the safety of HPV vaccines, but in all cases the safety issues described have been shown to be unrelated to HPV vaccination. In Colombia, over 300 girls experienced fainting, shortness of breath and weakness in the limbs after vaccination with Gardasil-4; the symptoms were later suggested to be caused by mass somatization disorder (hysterical disorder) (Stillo et al. 2015). In the United Kingdom, a girl died following vaccination; however, the cause of death was later revealed to be extensive tumor infiltration into the chest. Spanish health authorities withdrew tens of thousands of doses of Gardasil-4 in 2009 due to the hospitalization of two girls who received the vaccine (Mercola 2009). The Japanese government withdrew the recommendation for use of HPV vaccines until more detailed information about adverse effects became available (Morimoto et al. 2015). Meanwhile, the World Health Organization (WHO) Global Advisory Committee for Vaccine Safety indicated that policy decisions based on weak evidence of serious adverse events resulting from the use of HPV vaccines leads to the reduced use of safe and effective vaccines, which eventually causes significant harm (WHO 2015).
Autoimmune adverse events following vaccination
A recent issue that has been raised regarding the safety of HPV vaccines is adverse autoimmune events following vaccination. Several epidemiology studies reported that vaccination with Gardasil-4 is significantly related to serious autoimmune adverse events, including gastroenteritis, rheumatoid arthritis, thrombocytopenia, systemic lupus erythematosus, vasculitis, alopecia, central nervous system demyelinating conditions, ovarian damage, and irritable bowel syndrome (Soldevilla et al. 2012; Arango et al. 2015; Geier and Geier 2015, 2017; Gruber and Shoenfeld 2015). One study, however, found no evidence to support the association of Gardasil-4 with autoimmune events, including neurological and venous thromboembolic adverse events (Arnheim-Dahlstom et al. 2013). Therefore, the autoimmune events relating to the use of Gardasil-4 remain controversial and require further study.
Autoimmune events are not limited to the HPV vaccine. Numerous studies indicate that several vaccines, including vaccines against hepatitis B virus, tetanus toxoid, influenza virus, and polio vaccines, amongst others, are associated with such events (Shoenfeld and Aron-Maor 2000). One explanation for this involves ‘molecular mimicry’: antibodies elicited by vaccination attack host tissues because of structural similarity between the vaccine antigen and existing autoantigens (Waisbren 2008). As many viruses mimic the properties of host antigens to avoid immune surveillance (Lubyova and Pitha 2000), it seems inevitable that autoimmune events occur. Note, however, that the autoimmune response is also dependent upon the genetic background (Shoenfeld and Aron-Maor 2000).
Serious adverse events are very rare following vaccination with HPV vaccines, and the reasons for them are not clear. Since serious adverse events may cause a decline in participation in vaccination programs, long-term investigations should be undertaken to identify individuals who are at high risk of developing them.
The future of prophylactic HPV vaccines
L2-based vaccines
Vaccination with HPV L1 VLPs provides HPV type-specific protection (Wang and Roden 2013b) and cross protection against some types of HPV (Malagon et al. 2012). The specificity of L1 VLP-based vaccines may therefore allow infection by HPVs that are not targeted by the vaccine. In contrast, the N-terminal region of the L2 protein, aa 11–200, is highly conserved between HPV types (Schellenbacher et al. 2017), and the RG1 epitope (aa 17–36) within this region is pivotal for providing broad cross-neutralization activity (Gambhira et al. 2007). Regions aa 36–49, 69–81 and 108–120 of the L2 protein are also thought to confer cross protection (Kawana et al. 1999; Laniosz et al. 2007). The L2 protein is located inside the HPV capsid and is therefore inaccessible to the immune system (Kines et al. 2009; Wang and Roden 2013a). L2-specific antibody responses following natural HPV infections are indeed very rare: only seven of 1078 patients with high grade CINs or high grade vulvar intraepithelial neoplasia had serum that was reactive to L2 protein (Wang et al. 2015b).
Short lengths of the L2 peptide are poorly immunogenic. Therefore, several strategies have been applied to elicit robust immune responses, including displaying L2 epitopes on a carrier protein, the use of a peptide multimers, and conjugation with a fusion partner (Kanda and Kondo 2009; Jagu et al. 2011; Schellenbacher et al. 2017). A chimeric HPV16 L1 VLP vaccine displaying the HPV16 RG1 L2 epitope conferred efficient protection against vaginal challenge with mucosal pseudovirus high-risk types HPV16, HPV18, HPV45, HPV31, HPV33, HPV52, HPV58, HPV35, HPV39, HPV51, HPV59, HPV68, HPV56, HPV73, HPV26, HPV53, HPV66, and HPV34 as well as the low-risk types HPV6, HPV43, and HPV44 (Schellenbacher et al. 2013). The L2-based vaccines therefore have great potential for providing protection against a broad range of HPVs at a reduced vaccination price because of their simple format (Schiller and Muller 2015; Wang et al. 2015a; Huber et al. 2017).
The second-generation of HPV L1 VLP vaccines
The three currently licensed prophylactic HPV vaccines use insect or yeast cell expression systems and their manufacturing processes are therefore complex, which is the main reason for the high vaccine price. Numerous workers have sought to enhance the production processes for VLP-based vaccines, and considerable success has been achieved (Liew et al. 2010; Kim et al. 2014a; Thompson et al. 2016). HPV L1 VLP-based vaccines produced in S. cerevisiae, Escherichia coli (E. coli) and methylotrophic yeast species, such as Hansenula polymorpha (H. polymorpha) and Pichia pastoris (P. pastoris) are currently being studied in preclinical or clinical trials (Table 2).
EyeGene (Seoul, South Korea) have developed a bivalent vaccine containing HPV16 and HPV18 L1 VLPs produced in S. cerevisiae and administered in a vaccine formulation containing a proprietary adjuvant, CIA06 (Kim et al. 2014b). Recently, Xiamen INNOVAX Biotech (Xiamen, China) started a phase I clinical trial for Gelcolin (HPV6 and HPV11 L1 VLPs, developed for preventing genital warts) and a phase III clinical trial for Celcolin (HPV16 and HPV18 L1 VLPs for preventing cervical cancer). Both vaccines are produced in E. coli. The Serum Institute of India (Pune, India) has developed a quadrivalent vaccine that contains HPV6, HPV11, HPV16 and HPV18 using a H. polymorpha expression system, which is currently in a phase I clinical trial (Insight 2017a). Cadila Healthcare (Ahmedabad, India) are also currently running a phase I clinical trial (Insight 2017b) of their bivalent vaccine that contains HPV16 and HPV18 L1 VLPs produced in P. pastoris (Gupta et al. 2017). Shanghai Zerun Biotechnology’s bivalent vaccine (HPV16 and HPV18 L1 VLPs produced in P. pastoris) is currently in a phase III clinical trial. It is therefore expected that several biosimilar vaccines will be commercialized in the near future.
Therapeutic HPV vaccines
The main function of prophylactic vaccines is to induce humoral immunity against target pathogens, eventually leading to antibody induction and neutralization of pathogen activity (Mariani and Venuti 2010). Unlike prophylactic vaccines, the goal of therapeutic vaccines is to generate antigen-specific cytotoxic T-lymphocytes (CTLs) targeting pathogen-infected cells or cancer cells (Sayour and Mitchell 2017). The oncoproteins HPV E6 and E7 are expressed constantly during cervical carcinogenesis, and therefore several different therapeutic HPV vaccines targeting E6 and E7 have been developed (Hung et al. 2008). Processing of tumor-associated antigens by APCs, T cell recognition of the processed antigens, activation and proliferation of the T cells, and long-lasting T-cell responses are all required for effective antitumor responses (Farkona et al. 2016). Various types of therapeutic vaccines for treating CINs and cervical cancers have been developed, including protein/peptide-based vaccines, bacterial or viral vector-based vaccines, DNA-based vaccines and cell-based vaccines, with reported success in both animal studies and clinical trials (Yang et al. 2017).
Protein/peptide-based vaccines
The low immunogenicity of protein/peptide-based vaccines is insufficient to elicit successful therapeutic effects against cancerous lesions, although the vaccines are stable, safe and easy to produce (Kumar et al. 2015). These vaccines therefore require adjuvants and immunostimulating molecules (Lin et al. 2010a). A few HPV protein-based vaccines have reached the clinical trial stage and have demonstrated potential for eradicating high grade CINs (Vici et al. 2016).
Most epitopes present within the full-length HPV antigen activate the MHC class II pathway, which elicits the production of antibodies, rather than the CTL response (Su et al. 2010). Therefore, the inclusion of the MHC class I-restricted peptide from the E6 or E7 protein in the vaccine is thought to strengthen the CTL immune response (Skeate et al. 2016).
One important consideration in the development of peptide-based vaccines is the heterogeneity of the human leukocyte antigen (HLA) system between individuals, which causes different responses to epitopes used as vaccine peptides (Jin and Wang 2003). Therefore, long, overlapping peptides have been used to elicit cancer-cell-specific CTLs and therefore avoid failure caused by HLA polymorphisms (Kenter et al. 2009). The HPV E6 and E7 peptide-based vaccines show promising effects on tumor regression in an animal model (Yang et al. 2017). In clinical trials, however, the CTL responses induced by the peptide vaccines did not correlate with clinical outcomes, and tumor regression was minimal even though HPV-specific CTL responses were detected in some patients (Brinkman et al. 2007).
Live vector-based vaccines
Vector-based vaccines can be subdivided into viral and bacterial vector-based vaccines. Listeria monocytogenes and Lactobacillus spp. have been used as platforms for bacterial vector-based HPV vaccines (Sewell et al. 2004; Adachi et al. 2010), and Listeria monocytogenes is a particularly promising vector system for the delivery of HPV antigens (Cory and Chu 2014). The vaccine antigen can be expressed by fusion to the Listeria virulence factors LLO or ActA. The fused vaccine antigen is then presented through the MHC class I pathway because the virulence factors possess motif sequences for MHC class I-processing (Wood et al. 2008). Moreover, the localization of Listeria to the cytoplasm and endosomal compartment enables targeting of the vaccine antigen for both MHC class I and II presentation (Bolhassani and Zahedifard 2012). These properties of Listeria therefore facilitate the induction of both antigen-specific CTL and helper T-cell responses (Peters and Paterson 2003).
Adenoviruses, adeno-associated viruses, alphaviruses and vaccinia viruses have been used to deliver HPV E6, E7 and E2 antigens in vaccines (Hung et al. 2008). The vaccinia virus vector-based HPV vaccines have demonstrated considerable potential for causing disease regression in clinical trials (Vici et al. 2016). Ten (48%) of 21 patients with CIN2/3 had a clinical response following vaccination with the Modified Vaccinia Ankara (MVA) vector encoding HPV16 E6/E7 and IL-2 (Brun et al. 2011). The MVA vector is the preferred system for several types of therapeutic vaccines (Ura et al. 2014).
The live vector-based vaccines elicit strong immune responses against vaccine antigens because the live vectors can replicate within the body (Saxena et al. 2013). However, neutralizing antibodies against the vectors (both bacterial and viral), acquired either from vaccination or from pre-existing immunity, limit the booster effect usually achieved through repetitive vaccination (Saxena et al. 2013; Yang et al. 2017).
DNA-based vaccines
DNA-based vaccines are attractive vehicles for therapeutic HPV vaccination because they can induce both cellular and humoral immune responses, and they enable prolonged expression of the antigens (Hung et al. 2007; Lin et al. 2010b). DNA-based vaccines are relatively safe and can be injected repeatedly for booster vaccinations, unlike live vector-based vaccines, as they are non-live, non-replicating (Lin et al. 2010b) and less immunogenic than the live vectors (Klinman et al. 2000). Moreover, DNA-based vaccines are easy to produce for a low cost and are stable (Khan 2013). HPV DNA-based vaccines have shown promising therapeutic effects in clinical trials (Khallouf et al. 2014). Vaccination using DNA encoding HPV E7 with an abolished pRb binding site and Hsp70 from Mycobacterium tuberculosis, which was introduced to enhance APC uptake and MHC class I presentation, showed histologic regression in five of 12 patients with CIN 3 following prime-boost vaccination (Trimble et al. 2009; Maldonado et al. 2014). Meanwhile, suppression of the oncogenic activity of the HPV oncogenes E6 and E7 is an important consideration when designing DNA-based HPV vaccines (Kim et al. 2004; Trimble et al. 2009). An important consideration in the use of DNA-based vaccines is that the naked DNA molecules have low immunogenicity (Yosefi et al. 2014). Delivery by electroporation, gene gun, or laser, or encapsulation in microparticles were suggested to enhance antigen expression (Lin et al. 2010b). A HPV DNA vaccine, GX-188E, which was designed for electroporation-enhanced immunization and targeting HPV antigens E6 and E7 to dendritic cells, showed complete regression in seven of nine patient with CIN 3 (Kim et al. 2014c). Similarly, a HPV DNA vaccine VGX-3100, which was designed for electroporation, caused histopathological regression in 55 of 114 patient with CIN 2/3 (Trimble et al. 2015).
Several types of DNA-based vaccines are commercially available for animal use. Currently, however, no DNA-based vaccine has been approved for human use. Oncogene activation by random integration of the foreign DNA into host genome, immunologic tolerance by antigens expressed inside host body and atypical processing of the antigen produced by DNA vaccine have been indicated as drawbacks of the DNA-based vaccines (Khan 2013). Developments of safe and effective DNA vaccines are ongoing.
Cell-based vaccines
Cell-based vaccines are immune cells from patients that have been modified to have strong anti-cancer-cell activity, and they are therefore categorized as personalized immunotherapy (Janikashvili et al. 2010; Bethune and Joglekar 2017). Cell-based vaccines include dendritic cell (DC)-based and T-cell-based strategies (Palucka and Banchereau 2013; Fousek and Ahmed 2015).
Autologous DCs can be pulsed ex vivo with recombinant HPV E6 or E7 proteins or their peptides, and then returned to the patient (Ferrara et al. 2003; Santin et al. 2006; Rahma et al. 2014), where they present antigen to induce CTL proliferation. The drawbacks of DC-based vaccines are inconsistency of vaccine quality caused by variations in cell culture strategies, difficulty in obtaining large numbers of autologous DCs from the patient, low efficacy of DC transduction and the limited lifespan of autologous DCs (Skeate et al. 2016). DC-based vaccines gave no clinical responses, although HPV antigen-specific CTL responses were identified following implantation of pulsed DCs (Ferrara et al. 2003; Santin et al. 2006).
T-cell-based vaccines are based upon cancer-antigen-specific CTLs that are prepared ex vivo (Rosenberg and Restifo 2015). Autologous CTLs can be activated or genetically engineered ex vivo and delivered back into the patient (Rosenberg and Restifo 2015). Unlike DC-based vaccines, which act by stimulating CTLs in vivo, the CTL-based strategy misses out this step and instead directly affects the target cancer cells (Rosenberg et al. 2008). T-cell-based therapy is particularly promising, with fast patient response rates: tumor regression was reported in approximately 50% of patients with metastatic melanoma in two separate studies (Rosenberg et al. 2008; Kelderman et al. 2016). Many types of cancer are able to evade immune surveillance and downregulate immune functions by depleting cancer-cell-reacting CTLs (Chew et al. 2012; Quail and Joyce 2013). The advantage of ex vivo T-cell expansion is, therefore, that it enables T cells to be separated from the microenvironment of cancer (Yang et al. 2017). In a recent in phase II clinical trial, complete regression was reported in two of nine patients with metastatic cervical cancer following a single vaccination with CTLs reactive for HPV16 E6 and E7 proteins (Stevanovic et al. 2016). The processes required for adaptive cell transfer are presented in Fig. 1.

Schematic diagram of adaptive cell transfer for a patient with metastatic cervical cancer. The procedure for preparing tumor-infiltrating T cells was based on Stevanovic et al. (2016). The patient received cyclophosphamide prior to T-cell infusion. This lymphodepleting chemotherapy suppresses endogenous regulatory T cells and increases the persistence of the tumor-infiltrating T cells
Future prospects of HPV vaccines
Prophylactic vaccines
The duration of vaccine efficacy, as measured by antibody titers, is a critical consideration in the design of HPV vaccination schedules, because the main population targets for HPV vaccination are children and adolescents aged between 9 and 15 years. A cost effectiveness study indicated that current HPV vaccines will not provide protection against cervical cancer unless protection lasts for at least 15 years (Barnabas et al. 2006). Therefore, careful monitoring of the protection against cervical cancer conferred by prophylactic vaccines is required to fully evaluate the effect of vaccination. In addition, changes in vaccination schedules, routes of administration and doses of antigen for currently licensed vaccines, as well as the addition of more HPV antigens, may be required. Several second-generation HPV L1 VLP vaccines and next-generation vaccines, such as HPV L2-based vaccines and chimeric HPV VLP-based vaccines, are currently undergoing clinical trials or preclinical study (Schiller and Muller 2015). Agreement on well-established, relevant clinical endpoints and protocols is therefore a high priority.
The retail price of current HPV vaccines is over 450 USD for the full series (CDC 2017), and this high vaccination price leads to an inequitable distribution of the vaccines, as they are totally unaffordable for low-income countries. Most cervical cancers occur in low-income countries that have either no or few options for cervical cancer screening (Crosbie et al. 2013; Bruni et al. 2016). The impact of vaccination worldwide is therefore negligible, even if the vaccination rate reaches 100% in high-income countries (Harper and DeMars 2017). It is clear, therefore, that the goal must be the development of high efficacy and low price HPV vaccines.
Several factors affect price of a vaccine, such as the choice of vaccine platform, the design of upstream and downstream antigen production processes, and the sizes and durations of clinical trials. The primary endpoint has a critical role in determining the size and duration of a clinical trial. To date, CIN2 or worse (CIN2+) has been used as the clinical endpoint to assess HPV vaccination efficacy. However, the use of CIN2+ requires large and time-consuming clinical trials, and this hinders the development of new generation HPV vaccines (Lowy et al. 2015). Recently, a workshop of the International Agency for Research on Cancer and US National Cancer Institute recommended the use of persistent HPV infection, which occurs more frequently than CIN2+, as a primary endpoint for HPV vaccination trials, and suggested that immunobridging trials such as antibody titration using ELISA or pseudovirus-based neutralizing assay are sufficient for assessing immunological non-inferiority for alternative dosing schedules of licensed or biosimilar vaccines (Lowy et al. 2015). These recommendations should encourage the development of trials for new types of HPV vaccines in addition to new protocols for vaccination.
Therapeutic vaccines
Numerous trials of therapeutic HPV vaccines have advanced the development of cancer immunotherapy, as well as identified its limitation in practice. Unlike prophylactic vaccines, which provide humoral immunity for healthy individuals, therapeutic vaccines eliminate existing cancers, and therefore require an in-depth understanding of the tumor microenvironment and the immune system that controls it. Previous studies indicate that the duration of CTL responses generated by therapeutic vaccination is shorter than initially expected, and is therefore insufficient for eliminating advanced-stage cervical cancer (Skeate et al. 2016).
One of the important limitations of the function of cervical-cancer-cell-specific CTLs is the expression of immune checkpoints (Heong et al. 2017); activated CTLs upregulate CTL-associated protein 4 (CTLA-4) and programmed cell death protein 1 (PD1), which attenuate CTL activation (Buchbinder and Desai 2016). In addition, cervical cancer cells upregulate PD1 ligands on their surface, leading to the attenuation of cancer-specific CTL activation (Lyford-Pike et al. 2013). Recent trials of agents targeting immune checkpoints suggest that they show promise as therapeutic vaccines, not only for cervical cancer but also for other cancers (Postow et al. 2015; Lee et al. 2016).
Moreover, the recently developed concept of personalized immunotherapy has potential for cancer immunotherapy. Recent studies indicate that both intra- and inter-heterogeneity exist in the cancer microenvironment (Wayteck et al. 2014), and diverse somatic mutational events result in heterogeneity not only between patients but also between regions of hematological and solid cancer within an individual (Yap et al. 2012). This heterogeneity implies that full eradication of the cancer may be difficult even if immunotherapy targets multiple antigens. Moreover, cancer has several resistance mechanisms for evading immunotherapy, which are barriers to be overcome (Junttila and de Sauvage 2013). The heterogeneity of the cancer microenvironment might originate from diversity in genetic backgrounds and mutations during the evolution of the cancer (Schmitt et al. 2012); therefore, widespread use of next-generation sequencing will provide better data on which to base personalized therapy. In addition, the elucidation of new markers that predict the effect of immunotherapy will increase success (Wayteck et al. 2014).
Conclusions
Research data accumulated during the past decade indicates that the three currently licensed prophylactic HPV vaccines are highly efficacious, reliable and safe. Adverse effects related to autoimmune responses following vaccination with HPV vaccines are also reported for other types of vaccines. It is expected that consistent monitoring and research into identifying the correlation between adverse effects and genetic background will refine the vaccination protocol to avoid adverse effects not only in HPV vaccines but also in other vaccines.
The prophylactic vaccines have considerable limitation in terms of widespread use. Various types of second-generation HPV L1 VLP-based vaccines and new HPV L2-based vaccines are currently undergoing either preclinical or clinical trials. Agreement on the clinical endpoint for persistent HPV infection and endorsement of immunobridging studies will lessen the cost and duration of clinical trials and therefore facilitate the development of biosimilar and next-generation HPV vaccines of high efficacy and low price.
Trials of therapeutic HPV vaccines, including protein/peptide-, live vector-, DNA- and cell-based vaccines report that these HPV vaccines have considerable potential for safe and non-invasive treatment of cervical cancer as well as highlighting shortcomings in their practical use. One significant concern is that the cancer microenvironment and immune evasion mechanism are much more complex than previously thought. Chemotherapy with targeting of immune checkpoints has a synergistic effect on the elimination of cancer cells. Understanding the optimum conditions for immunotherapy remains a challenge, and further trials are needed to examine the practical use of therapeutic HPV vaccines.
References
Adachi K, Kawana K, Yokoyama T, Fujii T, Tomio A, Miura S, Tomio K, Kojima S, Oda K, Sewaki T, Yasugi T, Kozuma S, Taketani Y (2010) Oral immunization with a Lactobacillus casei vaccine expressing human papillomavirus (HPV) type 16 E7 is an effective strategy to induce mucosal cytotoxic lymphocytes against HPV16 E7. Vaccine 28:2810–2817
Adis Insight (2017a) Human papillomavirus vaccine recombinant quadrivalent—Serum Institute of India. Adis Insight. http://adisinsight.springer.com/drugs/800047216. Accessed 1 Aug 2017
Adis Insight (2017b) An open-label, single-treatment, single-period, single dose, clinical phase 1 study to assess the safety and tolerability of bivalent human papillomavirus (types 16 L1 & 18 L1) vaccine of M/s Cadila Healthcare Ltd., India in healthy, adult, female, human subjects. Adis Insight. http://adisinsight.springer.com/drugs/800040232. Accessed 26 Aug 2016
Antonsson A, Forslund O, Ekberg H, Sterner G, Hansson BG (2000) The ubiquity and impressive genomic diversity of human skin papillomaviruses suggest a commensalic nature of these viruses. J Virol 74:11636–11641
Antonsson A, Karanfilovska S, Lindqvist PG, Hansson BG (2003) General acquisition of human papillomavirus infections of skin occurs in early infancy. J Clin Microbiol 41:2509–2514
Araldi RP, Assaf SM, Carvalho RF, Carvalho MA, Souza JM, Magnelli RF, Modolo DG, Roperto FP, Stocco RC, Becak W (2017) Papillomaviruses: a systematic review. Genet Mol Biol 40:1–21
Arango MT, Tomljenovic L, Blank M, Shoenfeld Y (2015) HPV vaccination of Nzbxw/F1 mice. Arthritis Rheumatol 67(Supple 10):2147–2148
Arnheim-Dahlstom L, Pasternak B, Svanstrom H, Sparen P, Hviid A (2013) Autoimmune, neurological, and venous thromboembolic adverse events after immunisation of adolescent girls with quadrivalent human papillomavirus vaccine in Denmark and Sweden: cohort study. BMJ 347:f5906
Barnabas RV, Laukkanen P, Koskela P, Kontula O, Lehtinen M, Garnett GP (2006) Epidemiology of HPV 16 and cervical cancer in Finland and the potential impact of vaccination: mathematical modelling analyses. PLoS Med 3:e138
Bethune MT, Joglekar AV (2017) Personalized T cell-mediated cancer immunotherapy: progress and challenges. Curr Opin Biotechnol 48:142–152
Bolhassani A, Zahedifard F (2012) Therapeutic live vaccines as a potential anticancer strategy. Int J Cancer 131:1733–1743
Bouvard V, Baan R, Straif K, Grosse Y, Secretan B, El Ghissassi F, Benbrahim-Tallaa L, Guha N, Freeman C, Galichet L, Cogliano V, Wo WIaRCM (2009) A review of human carcinogens-part B: biological agents. Lancet Oncol 10:321–322
Braaten KP, Laufer MR (2008) Human papillomavirus (HPV), HPV-related disease, and the HPV vaccine. Rev Obstet Gynecol 1:2–10
Bravo IG, De Sanjose S, Gottschling M (2010) The clinical importance of understanding the evolution of papillomaviruses. Trends Microbiol 18:432–438
Brinkman JA, Hughes SH, Stone P, Caffrey AS, Muderspach LI, Roman LD, Weber JS, Kast WM (2007) Therapeutic vaccination for HPV induced cervical cancers. Dis Markers 23:337–351
Brun JL, Dalstein V, Leveque J, Mathevet P, Raulic P, Baldauf JJ, Scholl S, Huynh B, Douvier S, Riethmuller D, Clavel C, Birembaut P, Calenda V, Baudin M, Bory JP (2011) Regression of high-grade cervical intraepithelial neoplasia with TG4001 targeted immunotherapy. Am J Obstet Gynecol 204:169.e1–169.e8
Bruni L, Barrionuevo-Rosas L, Albero G, Serrano B, Mena M, Gómez D, Muñoz J, Bosch FX, De Sanjosé S (2016) Human papillomavirus and related diseases report. ICO information centre on HPV and cancer. http://www.hpvcentre.net/statistics/reports/XWX.pdf. Accessed 27 July 2017
Buchbinder EI, Desai A (2016) CTLA-4 and PD-1 pathways similarities, differences, and implications of their inhibition. Am J Clin Oncol-Canc 39:98–106
Buck CB, Day PM, Trus BL (2013) The papillomavirus major capsid protein L1. Virology 445:169–174
CDC (2017) Vaccines for children program (VFC) CDC vaccine price list. https://www.cdc.gov/vaccines/programs/vfc/awardees/vaccine-management/price-list/index.html. Accessed 3 July 2017
Cerqueira C, Schiller JT (2017) Papillomavirus assembly: an overview and perspectives. Virus Res 231:103–107
Cerqueira C, Pang YYS, Day PM, Thompson CD, Buck CB, Lowy DR, Schiller JT (2016) A cell-free assembly system for generating infectious human papillomavirus 16 capsids implicates a size discrimination mechanism for preferential viral genome packaging. J Virol 90:1096–1107
Chew V, Toh HC, Abastado JP (2012) Immune microenvironment in tumor progression: characteristics and challenges for therapy. J Oncol 2012:608406
Corp. MSD (2016) GARDASIL®9 [human papillomavirus 9-valent vaccine, recombinant]. Product monography. http://www.merck.ca/assets/en/pdf/products/GARDASIL_9-PM_E.pdf. Accessed 7 Dec 2016
Corp. MSD (2017) A Phase III Study of a 2-dose regimen of a multivalent human papillomavirus (HPV) vaccine (V503), administered to 9 to 14 year-olds and compared to young women, 16 to 26 years old (V503-010). ClinicalTrials.gov. https://clinicaltrials.gov/ct2/show/NCT01984697. Accessed 22 June 2017
Cory L, Chu C (2014) ADXS-HPV: a therapeutic Listeria vaccination targeting cervical cancers expressing the HPV E7 antigen. Hum Vaccine Immunother 10:3190–3195
Crosbie EJ, Einstein MH, Franceschi S, Kitchener HC (2013) Human papillomavirus and cervical cancer. Lancet 382:889–899
Cubie HA (2013) Diseases associated with human papillomavirus infection. Virology 445:21–34
Daling JR, Sherman KJ (1992) Relationship between human papillomavirus infection and tumours of anogenital sites other than the cervix. IARC Sci Publ 119:223–241
De Sanjose S, Diaz M, Castellsague X, Clifford G, Bruni L, Munoz N, Bosch FX (2007) Worldwide prevalence and genotype distribution of cervical human papillomavirus DNA in women with normal cytology: a meta-analysis. Lancet Infect Dis 7:453–459
Depuydt T, Beert J, Bosmans E, Salembier G (2016) Human papillomavirus (HPV) virion induced cancer and subfertility, two sides of the same coin. Facts Views Vis Obgyn 8:211–222
Dobson SR, Mcneil S, Dionne M, Dawar M, Ogilvie G, Krajden M, Sauvageau C, Scheifele DW, Kollmann TR, Halperin SA, Langley JM, Bettinger JA, Singer J, Money D, Miller D, Naus M, Marra F, Young E (2013) Immunogenicity of 2 doses of HPV vaccine in younger adolescents vs 3 doses in young women: a randomized clinical trial. JAMA 309:1793–1802
Doorbar J, Egawa N, Griffin H, Kranjec C, Murakami I (2015) Human papillomavirus molecular biology and disease association. Rev Med Virol 25(Suppl 1):2–23
Draper E, Bissett SL, Howell-Jones R, Edwards D, Munslow G, Soldan K, Beddows S (2011) Neutralization of non-vaccine human papillomavirus pseudoviruses from the A7 and A9 species groups by bivalent HPV vaccine sera. Vaccine 29:8585–8590
Draper E, Bissett SL, Howell-Jones R, Waight P, Soldan K, Jit M, Andrews N, Miller E, Beddows S (2013) A randomized, observer-blinded immunogenicity trial of Cervarix and Gardasil human papillomavirus vaccines in 12-15 year old girls. PLoS ONE 8:e61825
Dunne EF, Unger ER, Sternberg M, Mcquillan G, Swan DC, Patel SS, Markowitz LE (2007) Prevalence of HPV infection among females in the United States. JAMA 297:813–819
Einstein MH, Baron M, Levin MJ, Chatterjee A, Edwards RP, Zepp F, Carletti I, Dessy FJ, Trofa AF, Schuind A, Dubin G, Grp H-S (2009) Comparison of the immunogenicity and safety of Cervarix (TM) and Gardasil (R) human papillomavirus (HPV) cervical cancer vaccines in healthy women aged 18–45 years. Hum Vaccines 5:705–719
Farkona S, Diamandis EP, Blasutig IM (2016) Cancer immunotherapy: the beginning of the end of cancer? BMC Med 14:73
FDA (2006) June 8, 2006 Approval letter—human papillomavirus quadrivalent (types 6, 11, 16, 18) vaccine, recombinant. US Food and Drug Administration https://www.fda.gov/biologicsbloodvaccines/vaccines/approvedproducts/ucm111283.htm. Accessed 8 June 2006
FDA (2014a) December 10, 2014 Approval letter—GARDASIL 9. US Food and Drug Administration https://www.fda.gov/BiologicsBloodVaccines/Vaccines/ApprovedProducts/ucm426520.htm. Accessed 10 Dec 2014
FDA (2014b) July 21, 2014 Approval letter—CERVARIX. US Food and Drug Administration https://www.fda.gov/biologicsbloodvaccines/vaccines/approvedproducts/ucm406129.htm. Accessed 21 July 2014
Ferrara A, Nonn M, Sehr P, Schreckenberger C, Pawlita M, Durst M, Schneider A, Kaufmann AM (2003) Dendritic cell-based tumor vaccine for cervical cancer II: results of a clinical pilot study in 15 individual patients. J Cancer Res Clin 129:521–530
Ferris D, Samakoses R, Block SL, Lazcano-Ponce E, Restrepo JA, Reisinger KS, Mehlsen J, Chatterjee A, Iversen OE, Sings HL, Shou Q, Sausser TA, Saah A (2014) Long-term study of a quadrivalent human papillomavirus vaccine. Pediatrics 134:e657–e665
Fousek K, Ahmed N (2015) The evolution of T-cell therapies for solid malignancies. Clin Cancer Res 21:3384–3392
Gambhira R, Karanam B, Jagu S, Roberts JN, Buck CB, Bossis I, Alphs H, Culp T, Christensen ND, Roden RB (2007) A protective and broadly cross-neutralizing epitope of human papillomavirus L2. J Virol 81:13927–13931
Geier DA, Geier MR (2015) A case-control study of quadrivalent human papillomavirus vaccine-associated autoimmune adverse events. Clin Rheumatol 34:1225–1231
Geier DA, Geier MR (2017) Quadrivalent human papillomavirus vaccine and autoimmune adverse events: a case-control assessment of the vaccine adverse event reporting system (VAERS) database. Immunol Res 65:46–54
Gillison ML (2008) Human papillomavirus-related diseases: oropharynx cancers and potential implications for adolescent HPV vaccination. J Adolesc Health 43:S52–S60
Gravitt PE (2011) The known unknowns of HPV natural history. J Clin Invest 121:4593–4599
Gruber N, Shoenfeld Y (2015) A link between human papilloma virus vaccination and primary ovarian insufficiency: current analysis. Curr Opin Obstet Gynecol 27:265–270
Gupta G, Glueck R, Rishi N (2017) Physicochemical characterization and immunological properties of Pichia pastoris based HPV16L1 and 18L1 virus like particles. Biologicals 46:11–22
Harper DM, Demars LR (2017) HPV vaccines: a review of the first decade. Gynecol Oncol 146:196–204
Heong V, Ngoi N, Tan DSP (2017) Update on immune checkpoint inhibitors in gynecological cancers. J Gynecol Oncol 28:e20
Huber B, Schellenbacher C, Shafti-Keramat S, Jindra C, Christensen N, Kirnbauer R (2017) Chimeric L2-based virus-like particle (VLP) vaccines targeting cutaneous human papillomaviruses (HPV). PLoS ONE 12:e0169533
Hung CF, Monie A, Alvarez RD, Wu TC (2007) DNA vaccines for cervical cancer: from bench to bedside. Exp Mol Med 39:679–689
Hung CF, Ma B, Monie A, Tsen SW, Wu TC (2008) Therapeutic human papillomavirus vaccines: current clinical trials and future directions. Expert Opin Biol Ther 8:421–439
Jagu S, Malandro N, Kwak K, Yuan H, Schlegel R, Palmer KE, Huh WK, Campo MS, Roden RBS (2011) A multimeric L2 vaccine for prevention of animal papillomavirus infections. Virology 420:43–50
Janikashvili N, Larmonier N, Katsanis E (2010) Personalized dendritic cell-based tumor immunotherapy. Immunotherapy 2:57–68
Jin P, Wang E (2003) Polymorphism in clinical immunology: from HLA typing to immunogenetic profiling. J Transl Med 1:8
Joura EA, Giuliano AR, Iversen OE, Bouchard C, Mao C, Mehlsen J, Moreira ED Jr, Ngan Y, Petersen LK, Lazcano-Ponce E, Pitisuttithum P, Restrepo JA, Stuart G, Woelber L, Yang YC, Cuzick J, Garland SM, Huh W, Kjaer SK, Bautista OM, Chan IS, Chen J, Gesser R, Moeller E, Ritter M, Vuocolo S, Luxembourg A (2015) A 9-valent HPV vaccine against infection and intraepithelial neoplasia in women. N Engl J Med 372:711–723
Junttila MR, De Sauvage FJ (2013) Influence of tumour micro-environment heterogeneity on therapeutic response. Nature 501:346–354
Kanda T, Kondo K (2009) Development of an HPV vaccine for a broad spectrum of high-risk types. Hum Vaccines 5:43–45
Kawana K, Yoshikawa H, Taketani Y, Yoshiike K, Kanda T (1999) Common neutralization epitope in minor capsid protein L2 of human papillomavirus types 16 and 6. J Virol 73:6188–6190
Kelderman S, Heemskerk B, Fanchi L, Philips D, Toebes M, Kvistborg P, Van Buuren MM, Van Rooij N, Michels S, Germeroth L, Haanen JB, Schumacher NM (2016) Antigen-specific TIL therapy for melanoma: a flexible platform for personalized cancer immunotherapy. Eur J Immunol 46:1351–1360
Kemp TJ, Hildesheim A, Safaeian M, Dauner JG, Pan YJ, Porras C, Schiller JT, Lowy DR, Herrero R, Pinto LA (2011) HPV16/18 L1 VLP vaccine induces cross-neutralizing antibodies that may mediate cross-protection. Vaccine 29:2011–2014
Kenter GG, Welters MJP, Valentijn ARPM, Lowik MJG, Berends-Van Der Meer DMA, Vloon APG, Essahsah F, Fathers LM, Offringa R, Drijfhout JW, Wafelman AR, Oostendorp J, Fleuren GJ, Van Der Burg SH, Melief CJM (2009) Vaccination against HPV-16 oncoproteins for vulvar intraepithelial neoplasia. New Engl J Med 361:1838–1847
Khallouf H, Grabowska AK, Riemer AB (2014) Therapeutic vaccine strategies against human papillomavirus. Vaccines (Basel) 2:422–462
Khan KH (2013) DNA vaccines: roles against diseases. Germs 3:26–35
Kim HJ, Kim H-J (2016) Yeast as an expression system for producing virus-like particles: what factors do we need to consider? Lett Appl Microbiol 64:111–123
Kim JW, Hung CF, Juang J, He L, Kim TW, Armstrong DK, Pai SI, Chen PJ, Lin CT, Boyd DA, Wu TC (2004) Comparison of HPV DNA vaccines employing intracellular targeting strategies. Gene Ther 11:1011–1018
Kim HJ, Jin Y, Kim H-J (2014a) The concentration of carbon source in the medium affects the quality of virus-like particles of human papillomavirus type 16 produced in Saccharomyces cerevisiae. PLoS One 9:e94467
Kim KS, Park SA, Ko KN, Yi S, Cho YJ (2014b) Current status of human papillomavirus vaccines. Clin Exp Vaccine Res 3:168–175
Kim TJ, Jin HT, Hur SY, Yang HG, Seo YB, Hong SR, Lee CW, Kim S, Woo JW, Park KS, Hwang YY, Park J, Lee IH, Lim KT, Lee KH, Jeong MS, Surh CD, Suh YS, Park JS, Sung YC (2014c) Clearance of persistent HPV infection and cervical lesion by therapeutic DNA vaccine in CIN3 patients. Nat Commun 5:5317
Kim HJ, Kwag HL, Kim HJ (2016) Characterization of human papillomavirus type 16 pseudovirus containing histones. BMC Biotechnol 16:63
Kines RC, Thompson CD, Lowy DR, Schiller JT, Day PM (2009) The initial steps leading to papillomavirus infection occur on the basement membrane prior to cell surface binding. Proc Natl Acad Sci USA 106:20458–20463
Klinman DM, Takeshita F, Kamstrup S, Takeshita S, Ishii K, Ichino M, Yamada H (2000) DNA vaccines: capacity to induce auto-immunity and tolerance. Dev Biol (Basel) 104:45–51
Kreimer AR, Rodriguez AC, Hildesheim A, Herrero R, Porras C, Schiffman M, Gonzalez P, Solomon D, Jimenez S, Schiller JT, Lowy DR, Quint W, Sherman ME, Schussler J, Wacholder S, Grp CV (2011) Proof-of-principle evaluation of the efficacy of fewer than three doses of a bivalent HPV16/18 vaccine. J Natl Cancer I 103:1444–1451
Kumar S, Biswas M, Jose T (2015) HPV vaccine: current status and future directions. Med J Armed Forces India 71:171–177
Laniosz V, Nguyen KC, Meneses PI (2007) Bovine papillomavirus type 1 infection is mediated by SNARE syntaxin 18. J Virol 81:7435–7448
Lee SJ, Yang A, Wu TC, Hung CF (2016) Immunotherapy for human papillomavirus-associated disease and cervical cancer: review of clinical and translational research. J Gynecol Oncol 27:e51
Liew MWO, Rajendran A, Middelberg APJ (2010) Microbial production of virus-like particle vaccine protein at gram-per-litre levels. J Biotechnol 150:224–231
Lin K, Doolan K, Hung CF, Wu TC (2010a) Perspectives for preventive and therapeutic HPV vaccines. J Formos Med Assoc 109:4–24
Lin K, Roosinovich E, Ma B, Hung CF, Wu TC (2010b) Therapeutic HPV DNA vaccines. Immunol Res 47:86–112
Liu S, Semenciw R, Mao Y (2001) Cervical cancer: the increasing incidence of adenocarcinoma and adenosquamous carcinoma in younger women. CMAJ 164:1151–1152
Lowy DR, Herrero R, Hildesheim A, Endpoin INWP (2015) Primary endpoints for future prophylactic human papillomavirus vaccine trials: towards infection and immunobridging. Lancet Oncol 16:E226–E233
Lubyova B, Pitha PM (2000) Characterization of a novel human herpesvirus 8-encoded protein, vIRF-3, that shows homology to viral and cellular interferon regulatory factors. J Virol 74:8194–8201
Lyford-Pike S, Peng SW, Young GD, Taube JM, Westra WH, Akpeng B, Bruno TC, Richmon JD, Wang H, Bishop JA, Chen LP, Drake CG, Topalian SL, Pardoll DM, Pai SI (2013) Evidence for a role of the PD-1:PD-L1 pathway in immune resistance of HPV-associated head and neck squamous cell carcinoma. Cancer Res 73:1733–1741
Malagon T, Drolet M, Boily MC, Franco EL, Jit M, Brisson J, Brisson M (2012) Cross-protective efficacy of two human papillomavirus vaccines: a systematic review and meta-analysis. Lancet Infect Dis 12:781–789
Maldonado L, Teague JE, Morrow MP, Jotova I, Wu TC, Wang CG, Desmarais C, Boyer JD, Tycko B, Robins HS, Clark RA, Trimble CL (2014) Intramuscular therapeutic vaccination targeting HPV16 induces T cell responses that localize in mucosal lesions. Sci Transl Med 6:221ra13
Mariani L, Venuti A (2010) HPV vaccine: an overview of immune response, clinical protection, and new approaches for the future. J Transl Med 8:105
Markowitz LE, Dunne EF, Saraiya M, Lawson HW, Chesson H, Unger ER (2007) Quadrivalent human papillomavirus vaccine: recommendations of the advisory committee on immunization practices (ACIP). MMWR Recomm Rep 56:1–24
Mcneil C (2006) Who invented the VLP cervical cancer vaccines? J Natl Cancer Inst 98:433
Meites E, Kempe A, Markowits LE (2016) Use of a 2-dose schedule for human papillomavirus vaccination: updated recommendations of the advisory committee on immunization practices. CDC 65:1405–1408
Mercola (2009) Spain withdraws Gardasil after illnesses. http://articles.mercola.com/sites/articles/archive/2009/02/24/spain-withdraws-gardasil-after-illnesses.aspx. Accessed 24 Feb 2009
Mishra GA, Pimple SA, Shastri SS (2015) HPV vaccine: one, two, or three doses for cervical cancer prevention? Indian J Med Paediatr Oncol 36:201–206
Monie A, Hung CF, Roden R, Wu TC (2008) Cervarix: a vaccine for the prevention of HPV 16, 18-associated cervical cancer. Biologics 2:97–105
Morimoto A, Ueda Y, Egawa-Takata T, Yagi A, Terai Y, Ohmichi M, Ichimura T, Sumi T, Murata H, Kanzaki H, Nakai H, Mandai M, Yoshino K, Fujita M, Kimura T, Saito J, Sobue T, Nishikawa N, Sekine M, Enomoto T, Horikoshi Y, Takagi T (2015) Effect on HPV vaccination in Japan resulting from news report of adverse events and suspension of governmental recommendation for HPV vaccination. Int J Clin Oncol 20:549–555
Munoz N, Kjaer SK, Sigurdsson K, Iversen OE, Hernandez-Avila M, Wheeler CM, Perez G, Brown DR, Koutsky LA, Tay EH, Garcia PJ, Ault KA, Garland SM, Leodolter S, Olsson SE, Tang GWK, Ferris DG, Paavonen J, Steben M, Bosch FX, Dillner J, Huh WK, Joura EA, Kurman RJ, Majewski S, Myers ER, Villa LL, Taddeo FJ, Roberts C, Tadesse A, Bryan JT, Lupinacci LC, Giacoletti KED, Sings HL, James MK, Hesley TM, Barr E, Haupt RM (2010) Impact of human papillomavirus (HPV)-6/11/16/18 vaccine on all HPV-associated genital diseases in young women. J Natl Cancer I 102:325–339
Palucka K, Banchereau J (2013) Dendritic-cell-based therapeutic cancer vaccines. Immunity 39:38–48
Peters C, Paterson Y (2003) Enhancing the immunogenicity of bioengineered Listeria monocytogenes by passaging through live animal hosts. Vaccine 21:1187–1194
Petrosky E, Bocchini JA, Hariri S, Chesson H, Curtis CR, Saraiya M, Unger ER, Markowitz LE (2015) Use of 9-valent human papillomavirus (HPV) vaccine: updated HPV vaccination recommendations of the advisory committee on immunization practices. Morbid Mortal Wkly 64:300–304
Postow MA, Callahan MK, Wolchok JD (2015) Immune checkpoint blockade in cancer therapy. J Clin Oncol 33:1974–1982
Puthanakit T, Huang LM, Chiu CH, Tang RB, Schwarz TF, Esposito S, Frenette L, Giaquinto C, Mcneil S, Rheault P, Durando P, Horn M, Klar M, Poncelet S, De Simoni S, Friel D, De Muynck B, Suryakiran PV, Hezareh M, Descamps D, Thomas F, Struyf F (2016) Randomized open trial comparing 2-dose regimens of the human papillomavirus 16/18 AS04-adjuvanted vaccine in girls aged 9-14 years versus a 3-dose regimen in women aged 15-25 years. J Infect Dis 214:525–536
Quail DF, Joyce JA (2013) Microenvironmental regulation of tumor progression and metastasis. Nat Med 19:1423–1437
Rahma OE, Herrin VE, Ibrahim RA, Toubaji A, Bernstein S, Dakheel O, Steinberg SM, Abu Eid R, Mkrtichyan M, Berzofsky JA, Khleif SN (2014) Pre-immature dendritic cells (PIDC) pulsed with HPV16 E6 or E7 peptide are capable of eliciting specific immune response in patients with advanced cervical cancer. J Transl Med 12:353
Romanowski B, Schwarz TF, Ferguson L, Peters K, Dionne M, Behre U, Schulze K, Hillemanns P, Suryakiran P, Thomas F, Struyf F (2016) Sustained immunogenicity of the HPV-16/18 AS04-adjuvanted vaccine administered as a two-dose schedule in adolescent girls: five-year clinical data and modeling predictions from a randomized study. Hum Vaccine Immunother 12:20–29
Rosenberg SA, Restifo NP (2015) Adoptive cell transfer as personalized immunotherapy for human cancer. Science 348:62–68
Rosenberg SA, Restifo NP, Yang JC, Morgan RA, Dudley ME (2008) Adoptive cell transfer: a clinical path to effective cancer immunotherapy. Nat Rev Cancer 8:299–308
Santin AD, Bellone S, Palmieri M, Ravaggi A, Romani C, Tassi R, Roman JJ, Burnett A, Pecorelli S, Cannon MJ (2006) HPV16/18 E7-pulsed dendritic cell vaccination in cervical cancer patients with recurrent disease refractory to standard treatment modalities. Gynecol Oncol 100:469–478
Saxena M, Van TTH, Baird FJ, Coloe PJ, Smooker PM (2013) Pre-existing immunity against vaccine vectors: friend or foe? Microbiol-SGM 159:1–11
Sayour EJ, Mitchell DA (2017) Manipulation of innate and adaptive immunity through cancer vaccines. J Immunol Res 2017:3145742
Schellenbacher C, Kwak K, Fink D, Shafti-Keramat S, Huber B, Jindra C, Faust H, Dillner J, Roden RBS, Kirnbauer R (2013) Efficacy of RG1-VLP vaccination against infections with genital and cutaneous human papillomaviruses. J Invest Dermatol 133:2706–2713
Schellenbacher C, Roden RB, Kirnbauer R (2017) Developments in L2-based human papillomavirus (HPV) vaccines. Virus Res 231:166–175
Schiller JT, Lowy DR (2012) Understanding and learning from the success of prophylactic human papillomavirus vaccines. Nat Rev Microbiol 10:681–692
Schiller JT, Muller M (2015) Next generation prophylactic human papillomavirus vaccines. Lancet Oncol 16:E217–E225
Schiller JT, Castellsague X, Garland SM (2012) A review of clinical trials of human papillomavirus prophylactic vaccines. Vaccine 30(Suppl 5):F123–F138
Schmitt MW, Prindle MJ, Loeb LA (2012) Implications of genetic heterogeneity in cancer. Ann N Y Acad Sci 1267:110–116
Senapati R, Senapati NN, Dwibedi B (2016) Molecular mechanisms of HPV mediated neoplastic progression. Infect Agents Cancer 11:59
Sewell DA, Douven D, Pan ZK, Rodriguez A, Paterson Y (2004) Regression of HPV-positive tumors treated with a new Listeria monocytogenes vaccine. Arch Otolaryngol 130:92–97
Shoenfeld Y, Aron-Maor A (2000) Vaccination and autoimmunity: ‘Vaccinosis’: a dangerous liasion? J Autoimmun 14:1–10
Skeate JG, Woodham AW, Einstein MH, Da Silva DM, Kast WM (2016) Current therapeutic vaccination and immunotherapy strategies for HPV-related diseases. Hum Vaccine Immunother 12:1418–1429
Soldevilla HF, Briones SF, Navarra SV (2012) Systemic lupus erythematosus following HPV immunization or infection? Lupus 21:158–161
Stevanovic S, Draper LM, Langhan MM (2016) Complete regression of metastatic cervical cancer after treatment with human papillomavirus-targeted tumor-infiltrating T cells. J Clin Oncol 34:519
Stillo M, Carrillo Santisteve P, Lopalco PL (2015) Safety of human papillomavirus vaccines: a review. Expert Opin Drug Saf 14:697–712
Su JH, Wu AJ, Scotney E, Ma B, Monie A, Hung CF, Wu TC (2010) Immunotherapy for cervical cancer research status and clinical potential. Biodrugs 24:109–129
Szarewski A, Skinner SR, Garland SM, Romanowski B, Schwarz TF, Apter D, Chow SN, Paavonen J, Del Rosario-Raymundo MR, Teixeira JC, De Carvalho NS, Castro-Sanchez M, Castellsague X, Poppe WA, De Sutter P, Huh W, Chatterjee A, Tjalma WA, Ackerman RT, Martens M, Papp KA, Bajo-Arenas J, Harper DM, Torne A, David MP, Struyf F, Lehtinen M, Dubin G (2013) Efficacy of the HPV-16/18 AS04-adjuvanted vaccine against low-risk HPV types (PATRICIA randomized trial): an unexpected observation. J Infect Dis 208:1391–1396
Thompson CM, Aucoin MG, Kamen AA (2016) Production of virus-like particles for vaccination. Method Mol Biol 1350:299–315
Trimble CL, Peng S, Kos F, Gravitt P, Viscidi R, Sugar E, Pardoll D, Wu TC (2009) A phase I trial of a human papillomavirus DNA vaccine for HPV16+ cervical intraepithelial neoplasia 2/3. Clin Cancer Res 15:361–367
Trimble CL, Morrow MP, Kraynyak KA, Shen XF, Dallas M, Yan J, Edwards L, Parker RL, Denny L, Giffear M, Brown AS, Marcozzi-Pierce K, Shah D, Slager AM, Sylvester AJ, Khan A, Broderick KE, Juba RJ, Herring TA, Boyer J, Lee J, Sardesai NY, Weiner DB, Bagarazzi ML (2015) Safety, efficacy, and immunogenicity of VGX-3100, a therapeutic synthetic DNA vaccine targeting human papillomavirus 16 and 18 E6 and E7 proteins for cervical intraepithelial neoplasia 2/3: a randomised, double-blind, placebo-controlled phase 2b trial. Lancet 386:2078–2088
Ura T, Okuda K, Shimada M (2014) Developments in viral vector-based vaccines. Vaccines (Basel) 2:624–641
Vici P, Pizzuti L, Mariani L, Zampa G, Santini D, Di Lauro L, Gamucci T, Natoli C, Marchetti P, Barba M, Maugeri-Sacca M, Sergi D, Tomao F, Vizza E, Di Filippo S, Paolini F, Curzio G, Corrado G, Michelotti A, Sanguineti G, Giordano A, De Maria R, Venuti A (2016) Targeting immune response with therapeutic vaccines in premalignant lesions and cervical cancer: hope or reality from clinical studies. Expert Rev Vaccines 15:1327–1336
Waisbren BA (2008) Acquired autoimmunity after viral vaccination is caused by molecular mimicry and antigen complementarity in the presence of an immunologic adjuvant and specific HLA patterns. Med Hypotheses 70:346–348
Wang JW, Roden RBS (2013a) L2, the minor capsid protein of papillomavirus. Virology 445:175–186
Wang JW, Roden RBS (2013b) Virus-like particles for the prevention of human papillomavirus-associated malignancies. Expert Rev Vaccines 12:129–141
Wang DN, Li ZH, Xiao JQ, Wang JQ, Zhang L, Liu YJ, Fan F, Xin L, Wei MX, Kong ZB, Yu H, Gu Y, Zhang J, Li SW, Xia NS (2015a) Identification of broad-genotype HPV L2 neutralization site for pan-HPV vaccine development by a cross-neutralizing antibody. PLoS ONE 10:e0123944
Wang JW, Jagu S, Wu WH, Viscidi RP, Macgregor-Das A, Fogel JM, Kwak K, Daayana S, Kitchener H, Stern PL, Gravitt PE, Trimble CL, Roden RBS (2015b) Seroepidemiology of human papillomavirus 16 (HPV16) L2 and generation of L2-specific human chimeric monoclonal antibodies. Clin Vaccine Immunol 22:806–816
Watson RA (2005) Human papillomavirus: confronting the epidemic: a urologist’s perspective. Rev Urol 7:135–144
Wayteck L, Breckpot K, Demeester J, De Smedt SC, Raemdonck K (2014) A personalized view on cancer immunotherapy. Cancer Lett 352:113–125
WHO (2003) Global cancer rates could increase by 50% to 15 million by 2020. http://www.who.int/mediacentre/news/releases/2003/pr27/en/. Accessed 3 April 2003
WHO (2014) Human papillomavirus vaccines: WHO position paper, October 2014. Wkly Epidemiol Rec 89:465–492
WHO (2015) Global advisory committee on vaccine safety statement on safety of HPV vaccines. http://www.who.int/vaccine_safety/committee/GACVS_HPV_statement_17Dec2015.pdf. Accessed 17 Dec 2015
WHO (2017) Human papillomavirus vaccines: WHO position paper, May 2017. Wkly Epidemiol Rec 92:241–268
Woestenberg PJ, King AJ, Der Sande Van, MaB Donken R, Leussink S, Van Der Klis FRM, Hoebe CJPA, Bogaards JA, Van Benthem BHB, Labs MM, Serv PH (2017) No evidence for cross-protection of the HPV-16/18 vaccine against HPV-6/11 positivity in female STI clinic visitors. J Infect 74:393–400
Wood LM, Guirnalda PD, Seavey MM, Paterson Y (2008) Cancer immunotherapy using Listeria monocytogenes and listerial virulence factors. Immunol Res 42:233–245
Yang A, Farmer E, Lin J, Wu TC, Hung CF (2017) The current state of therapeutic and T cell-based vaccines against human papillomaviruses. Virus Res 231:148–165
Yap TA, Gerlinger M, Futreal PA, Pusztai L, Swanton C (2012) Intratumor heterogeneity: seeing the wood for the trees. Sci Transl Med 4:127ps10
Yosefi T, Tafvizi F, Shamsara M, Harkynejad T, Ghorashi SA, Mahravani H (2014) Improve immunogenicity of DNA vaccine against foot-and-mouth disease virus: role of intron and probably viral 3C protease as biological adjuvants. Mol Genet Microbiol 29:154–157
Zandberg DP, Bhargava R, Badin S, Cullen KJ (2013) The role of human papillomavirus in nongenital cancers. Cancer J Clin 63:57–81
Zur Hausen H (2009) The search for infectious causes of human cancers: where and why. Virology 392:1–10
Acknowledgements
The present study was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (NRF-2015R1D1A1A01057370).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declared no conflict of interest.
Rights and permissions
About this article

Cite this article
Kim, H.J., Kim, HJ. Current status and future prospects for human papillomavirus vaccines. Arch. Pharm. Res. 40, 1050–1063 (2017). https://doi.org/10.1007/s12272-017-0952-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12272-017-0952-8