Abstract
Activation of translocator protein (18 kDa) (TSPO) plays an important role to mediate rapid anxiolytic efficacy in stress response and stress-related disorders by the production of neurosteroids. However, little is known about the ligand of TSPO on the anxiety-like and depressive behaviors and the underlying mechanisms in chronic unpredictable mild stress (UCMS) mice. In the present study, a novel ligand of TSPO, ZBD-2 [N-benzyl-N-ethyl-2-(7,8-dihydro-7-benzyl-8-oxo-2-phenyl-9H-purin-9-yl) acetamide] synthesized by our laboratory, was used to evaluate the anxiolytic and antidepressant efficacy and to elucidate the underlying mechanisms. ZBD-2 (3 mg/kg) significantly attenuated anxiety-like and depressive behaviors in the UCMS mice, which was blocked by TSPO antagonist PK11195 (3 mg/kg). Treatment of ZBD-2 reversed the decrease in biogenic amines (norepinephrine, dopamine, and serotonin) in the brain region of hippocampus in the UCMS mice. The decreases in TSPO, GluN2B-containing N-methyl-d-aspartate (NMDA) receptors, GluA1, p-GluA1-Ser831, p-GluA1-Ser845, PSD-95, and GABAA-a2 were integrated with the increases of CaMKII and iNOS levels in the hippocampus of the UCMS mice. ZBD-2 significantly reversed the changes of above proteins. However, ZBD-2 or PK11195 treatment did not affect the levels of GluN2A-containing NMDA receptors and the total levels of GAD67. Our study provides strong evidences that ZBD-2 has a therapeutic effect on chronic stress-related disorders such as depression and anxiety through regulating the biogenic amine levels and the synaptic proteins in the hippocampus.
Similar content being viewed by others

Introduction
The translocator protein (18 kDa) (TSPO) is a five trans-membrane domain protein that is mainly located in the central nervous system. It is mainly located in the outer mitochondrial membrane and favors the transport of cholesterol to the inner mitochondrial membrane, ultimately promoting neurosteroid synthesis (Nothdurfter et al. 2012). The importance of neurosteroids for the regulation of emotion and therapeutic use in depression and anxiety may include GABAergic mechanisms (Zorumski et al. 2013). TSPO is not directly targeted to the GABAA receptor (Anholt et al. 1986). TSPO plays a crucial role in the synthesis of neurosteroids by promoting transport of cholesterol to the inner mitochondrial membrane, which is the rate-limiting step in neurosteroidogenesis (Nothdurfter et al. 2012). For example, TSPO regulates the synthesis of pregnenolone from cholesterol (Papadopoulos et al. 2006). The progesterone metabolite, allopregnanolone, is a potent positive allosteric modulator of the action of GABAA receptors (Belelli and Lambert 2005). TSPO density has been found to be altered in stress responses: Down-regulation has been shown in repeated or chronic stress, while up-regulation has been found in acute stress conditions (Veenman and Gavish 2006; Gavish et al. 1996). However, little is known regarding the roles and the underlying mechanisms of TSPO in unpredictable chronic mild stress-induced anxiety-like and depressive behaviors.
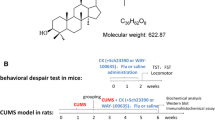
ZBD-2 is a novel ligand for TSPO synthesized in our laboratory (Wang et al. 2015). It is an analog of the XBD173 (Rupprecht et al. 2009), one of the ligands of TSPO. Our previous study reported that ZBD-2 showed high affinity for TSPO prepared from rat brain mitochondria in the nanomolar range (Ki = 0.463 nM) (Wang et al. 2015), comparable to the affinity of 3H-PK11195, the classic TSPO ligand, for TSPO (Ki = 0.657 nM) (Kita et al. 2004). Activation of TSPO by ZBD-2 attenuated chronic pain-induced anxiety-like behaviors by regulating the balance between GABAergic and glutamatergic transmission in the BLA of hindpaw CFA-injected mice (Wang et al. 2015). We also found that systemic treatment of ZBD-2 provided significant neuroprotection by inhibiting GluN2B-containing N-methyl-d-aspartate (NMDA) receptor-mediated excitotoxicity in mice subjected to middle cerebral artery occlusion in vivo (Li et al. 2015).
Hippocampus is the critical brain region in the development of the stress-related pathophysiological disorder (Dranovsky and Hen 2006). The alterations of synaptic transmission, biogenic amines, and inflammation in the central nervous system are involved in the etiology of depression (Haase and Brown 2015). A key component of serotonin 5-HT1A is abundantly expressed in the hippocampus that plays pivotal roles in fear, anxiety, stress, and cognition (Albert et al. 2014). Chronic treatment with selective serotonin reuptake inhibitors (SSRIs) stimulates hippocampal neurogenesis through the activation of 5-HT1A receptors (Murata et al. 2015). Glutamate receptors are involved in the development of emotional disorders. Increase in NMDA receptor subtypes levels has been shown to be related to impaired hippocampal-dependent task performance and depression (Vasquez et al. 2014). At the same time, chronic social stress results in decreases in mRNA of α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor (AMPAR) GluA1 subunits in CA1, and chronic SSRI treatment rescued AMPAR dysfunction in chronic stress animals (Schmidt et al. 2010; Kallarackal et al. 2013). Recently, a genetic variation in the calcium/calmodulin-dependent protein kinase II (CaMKII) pathway has been linked to antidepressant response (Li et al. 2013). These studies indicate that glutamate receptors and excitatory synaptic-related proteins are critical in the development and/or maintenance of depression. As a pro-inflammatory factor, iNOS is related to the development of depression. Its inhibitors significantly abolish the depressive-like behaviors (Khoshnoodi et al. 2015; Tomaz et al. 2014; Liu et al. 2015), suggesting the beneficial therapeutic effects by inhibiting iNOS expression. Because the monoamine, in particular serotonergic neurotransmission, has long been recognized as an important factor in the etiology of depression, and the role of hippocampus in development of emotional disorders, we focused on the hippocampus to elucidate the effects of ZBD-2.
SSRIs, tricyclic antidepressants, and benzodiazepines (e.g., diazepam) are main drugs for the treatment of both depression and anxiety disorders, but these drugs are with a great burden of adverse effects (Nelson and Philbrick 2012; Anderson et al. 2000). Here, we found that ZBD-2 has a therapeutic effect on chronic stress-related disorders such as depression and anxiety. The underlying mechanisms involve the regulation of biogenic amine levels and neurotransmitter receptors in the hippocampus.
Materials and Methods
Drugs
All chemicals were purchased from Sigma-Aldrich (St. Louis, MO, USA) unless otherwise stated. ZBD-2 was synthesized in our laboratory with the purity of 99.9 % and dissolved in the saline (0.9 % NaCl). l-Cysteine, dopamine hydrochloride (DA), serotonin (5-HT), norepinephrine (NE), γ-aminobutyric acid (GABA), 3,4-dihydroxyphenylacetic acid (DOPAC), 5-hydroxyindole-3-acetic acid (5-HIAA), and homovanillic acid (HVA) were obtained from J&K (Beijing, China). HPLC-grade methanol, acetic acid, and perchloric acid were purchased from VWR (Milan, China). Anti-GluN2A, anti-GluN2B, and anti-GAD67 were purchased from Millipore (Billerica, MA, USA). Anti-CaMKII, anti-TSPO, and anti-GABAA-a2 antibodies were purchased from Chemicon (Temecula, CA, USA). Anti-GluA1, anti-PSD-95, p-GluA1-Ser 831, and p-GluA1-Ser 845 were purchased from Abcam (Cambridge, UK). Anti-iNOS, anti-5-HT1A, and anti-5-HT2A antibodies were purchased from GenTex (Alton PkwyIrvine, CA, USA).
Animals
Male adult C57BL/6 mice (6–8 weeks old) were obtained from the Laboratory Animal Center of the Fourth Military Medical University. The animals were housed in plastic boxes in groups with food and water ad libitum in a colony room with controlled temperature (24 ± 2 °C), humidity (50–60 %), and a 12:12 h light–dark cycle. Mice were allowed to adapt to laboratory conditions for at least 1 week before the procedure. The experiments were performed during the light phase of the cycle. The Institutional Ethical Committee of the Fourth Military Medical University specifically approved this study.
Chronic Unpredictable Mild Stress (UCMS) and Drug Treatments
To establish the UCMS model, mice were exposed to various stressors for 5 weeks, and time-matched control mice did not receive any stressors. The stressors included restraint (2 h), inversion of day/night light cycle (light off and light on), cold (in a cold room at 4 °C for 1 h), 45° tilted cage (overnight), cage rotation (20 min), wet bedding (250 ml of water added into cage, overnight), no bedding (overnight), food and water deprivation (overnight), forced swimming (cold water 4 °C for 6 min), and overcrowding (overnight) (Willner et al. 1987; Koo and Duman 2008). On average, two stressors were administered per day. Non-stressed controls were handled only for cage changes and behavioral tests.
Mice were divided into four groups: control group, UCMS + vehicle, UCMS + ZBD-2, UCMS + ZBD-2 + PK11195. ZBD-2 (once daily, p.o., 3 mg/kg) was treated at the last 2 weeks (from day 21 to day 35) for the long duration of UCMS, which was longer than the time course in our previous study about chronic pain (Wang et al. 2015). The dose of ZBD-2 (3 mg/kg) was selected according to our previous research (Wang et al. 2015). PK11195 (a potent TSPO antagonist ligand, 3 mg/kg, i.p.) was treated 30 min after ZBD-2. Behavior experiments were conducted 60 min after the last administration with ZBD-2 or 30 min after the last administration with PK11195. Brain samples were harvested immediately for biochemical detections after behavioral experiments.
Open Field (OF)
OF was conducted as described in our previous report (Guo et al. 2012). The open field is a square arena (30 × 30 × 30 cm3) with clear Plexiglas walls and floor. It is placed inside an isolation chamber with dim illumination and a fan. Mice were placed in the center of the box and allowed to adjust to the environment for 10 min. Mice were videotaped using a camera fixed above the floor and analyzed with a video tracking system. The “center” field is defined as the central 15 × 15 cm2 area of the open field, one-fourth of the total area. Each subject was placed in the center of the open field, and its activity was measured for 15 min.
Elevated Plus Maze (EPM)
EPM was conducted as described in our previous report (Guo et al. 2012). The apparatus comprises two open arms (25 × 8 × 0.5 cm) and two closed arms (25 × 8 × 12 cm) that extend from a common central platform (8 × 8 cm). The apparatus is elevated to a height of 50 cm above the floor. Mice were allowed to habituate to the testing room for 2 days before the test and pretreated with gentle handling two times per day to eliminate nervousness. For each test, individual animals were placed in the center square, facing an open arm, and allowed to move freely for 5 min. Mice were videotaped using a camera fixed above the maze and analyzed with a video tracking system. The number of entries and time spent in each arm were recorded.
Forced Swimming Test (FST)
Mice were individually placed in an open cylindrical container (diameter 10 cm, height 25 cm) containing 19 cm of water at 25 ± 1 °C as previously described (Bourin et al. 2004). Mice were allowed to swim for 6 min. Each mouse was judged to be immobile when it ceased struggling and remained floating motionless in water, making only those movements necessary to keep its head above water. The duration of immobility was recorded during the last 5 min of the test by a competent observer blind to treatment.
Tail Suspension Test (TST)
TST was performed as previously described (Cates et al. 2013). Briefly, mice were individually suspended by tail with a clamp (1 cm from the tip of the end) in a box (25 × 25 × 30 cm3) with the head 5 cm from the bottom. Testing was carried out in a darkened room with minimal background noise. The duration of the test was 6 min, and immobility was determined for the last 5 min of the test. Mice were considered immobile only when they hung passively and completely motionless. The time was detected by a competent observer blind to treatment.
Measurement of Neurotransmitters
Tissue samples from the hippocampus were dissected from brain slices (300 μm) under an anatomical microscope. Biogenic amines and their metabolites (DA, 5-HT, 5-HIAA, HVA, DOPAC, NE and GABA) in the hippocampus were estimated by HPLC (Shimadzu LC-20A, Kyoto, Japan) as described in a previous report (De Benedetto et al. 2014). Stock solution (100 μg/ml) was prepared in water and stored in dark at 4 °C. Working solutions were freshly prepared by diluting the stock solution to the appropriate concentration with 0.2 M perchloric acid containing 3 mM cysteine. Standard solutions with concentration ranges as 0.031, 0.062, 0.312, 0.625, 0.938, 1.25, and 2.50 ng/μl for DA, NE, 5-HT, GABA, 5-HIAA, DOPAC, and HVA were prepared by diluting the stock solutions. Hippocampus dissection was homogenized in 200 μl of 0.2 M perchloric acid containing 3 mM cysteine. After 20 strokes, the homogenate was centrifuged (12,000×g, 10 min, 4 °C), and the resulting supernatant was left on ice for 5 min and then analyzed by HPLC. Chromatographic separation was achieved on a Microsorb-MV100-5 C18 chromatography column (250 × 4.6 mm, 5 μm). The column temperature was maintained at 35 °C, and the flow rate was 1.0 ml/min for all the experiments. The mobile phase was acetate buffer (pH 3.5, 12 mM acetic acid, 0.26 mM Na2EDTA)–methanol (86:14, v/v). The photodiode array (PDA) was monitored at excitation and emission wavelengths of 279 and 320 nm, respectively. Peaks were identified by comparing their retention time in the sample (tissue extracts) solution with that of standard solution. The supernatant was further filtered through 0.22-μm nylon filters before injecting in the HPLC injection pump. Separation was carried out at a flow rate of 0.8 ml/min.
Western Blot
Western blot analysis was performed as described previously (Wang et al. 2015). Equal amounts of protein (50 μg) from the hippocampus were separated and electro-transferred onto PDVF membranes (Invitrogen), which were probed with antibodies for CaMKII (dilution ratio 1:1000), iNOS (dilution ratio 1:1000), anti-PSD-95 (dilution ratio 1:1000), GluA1 (dilution ratio 1:500), p-GluA1-Ser 831 (dilution ratio 1:1000), and p-GluA1-Ser 845 (dilution ratio 1:1000), anti-5-HT1A (dilution ratio 1:500), anti-5-HT2A (dilution ratio 1:500), GluN2A (dilution ratio 1:500), GluN2B (dilution ratio 1:500), GABAA-a2 (dilution ratio 1:500), GAD67 (dilution ratio 1:1000), and with β-actin (dilution ratio 1:10,000) as a loading control. The membranes were incubated with horseradish peroxidase-conjugated secondary antibodies (anti-rabbit/anti-mouse IgG for the primary antibodies). Bands were visualized using an ECL system (Lightning Blot System, PerkinElmer, Waltham, MA, USA). For data quantification, band intensity of each blot was calculated as ratio relative to β-actin. The intensity ratio of the control group was set as 100 %, and the intensity of other treatment groups was expressed as percentage to the control group.
Data Analysis
Results were expressed as the mean ± SEM. Data were evaluated using one-way analysis of variance (ANOVA) for post hoc comparisons (SPSS 13.0). Data that passed the homogeneity test were analyzed by the one-way ANOVA least significant difference (LSD) test. Data that did not pass the homogeneity test were analyzed by the one-way ANOVA Dunnett’s T3 test. In all cases, p < 0.05 was considered statistically significant.
Results
Modulation of Anxiety and Depressive Behaviors in UCMS Mice
Administration of ZBD-2 (3 mg/kg, p.o., once daily for 2 weeks) increased open-arm entries and time spent in open arms in UCMS mice in the EPM test (Fig. 1a, b). In the OF test, the total distance traveled and the time in the center area were notably decreased in the UCMS mice (Fig. 1c, d). ZBD-2 administration significantly increased the time in the center area and the total distance traveled in the OF test. In the FST and TST tests, ZBD-2 decreased the mean immobility period of the UCMS mice (Fig. 1e, f). These effects were abolished by the TSPO antagonist PK11195 (3 mg/kg).

ZBD-2 reduces anxiety-like and depressive behaviors. a, b ZBD-2 (3 mg/kg) reversed the open-arm entries and the time spent in open arm of UCMS mice in EPM test. c, d ZBD-2 (3 mg/kg) reversed the time in the center area and total distance traveled of UCMS mice in OF test. e, f ZBD-2 (3 mg/kg) decreased the immobility time in UCMS mice in the FST and TST tests. Above effects of ZBD-2 were abolished by PK11195 (3 mg/kg). UCMS means the mice treated with chronic unpredictable mild stress. Con Control, Veh Vehicle. n = 6 in each group. *p < 0.05; **p < 0.01 compared with the control mice, # p < 0.05; ## p < 0.01 compared with the UCMS Veh group
Effects of ZBD-2 on Neurotransmitter Levels
Administration of ZBD-2 (3 mg/kg) significantly reversed the down-regulation of DA (Fig. 2a), 5-HT (Fig. 2b), 5-HIAA (Fig. 2c), DA metabolite 3,4-dihydroxyphenylacetic acid (DOPAC) (Fig. 2d), NE (Fig. 2e), DA metabolite homovanillic acid (HVA) (Fig. 2f), and GABA (Fig. 2g) in the hippocampus of UCMS mice. The effects of ZBD-2 were abolished by the PK11195 (3 mg/kg).

Effects of ZBD-2 on the monoamine levels. ZBD-2 (3 mg/kg) increased the content of DA (a), 5-HT (b), 5-HIAA (c), DOPAC (DA metabolite 3,4-dihydroxyphenylacetic acid) (d), NE (e), HVA (homovanillic acid) (f), and GABA (g) in the hippocampus of UCMS mice. The effects of ZBD-2 were blocked by PK11195 (3 mg/kg). Con Control, Veh Vehicle. n = 6 mice in each group. *p < 0.05; **p < 0.01 compared with the control mice, # p < 0.05; ## p < 0.01 compared with the UCMS Veh group
ZBD-2 Regulates the Levels of TSPO
It has been reported that chronic stress induces a decrease in TSPO density in the brain and peripheral organs in rodents (Weizman and Gavish 1993). At the same time, a decrease in lymphocyte or platelet TSPO levels in patients with post-traumatic stress disorder and panic disorder has been reported (Rocca et al. 1998; Gavish et al. 1999). In the present study, levels of TSPO in the hippocampus were decreased in UCMS mice. Treatment of ZBD-2 (3 mg/kg) for 2 weeks significantly increased the levels of TSPO in the hippocampus (Fig. 3). This effect was blocked by a TSPO antagonist PK11195 (3 mg/kg).

Effects of ZBD-2 on levels of TSPO. a Representative bands of Western blot analysis for TSPO in the hippocampus. b ZBD-2 (3 mg/kg) increased the levels of TSPO in the hippocampus of UCMS mice. This effect was blocked by PK11195 (3 mg/kg). Con Control, Veh Vehicle. n = 6 in each group. **p < 0.01 compared with the control mice, ## p < 0.01 compared with the UCMS Veh group
Effects of ZBD-2 on Serotonin Receptors
Serotonin receptors 5-HT1A and 5-HT2A are all involved in the clinical effects of SSRIs. Activation of 5-HT1A or blockade of 5-HT2A receptors seems to improve the clinical effects of SSRIs (Celada et al. 2004). Levels of 5-HT1A were significantly reduced, whereas levels of 5-HT2A were increased in the hippocampus of UCMS mice (Fig. 4a–c). ZBD-2 treatment restored the levels of 5-HT1A and 5-HT2A to the basal levels in the UCMS mice. PK11195 (3 mg/kg) abolished these effects.

Effects of ZBD-2 on levels of serotonin receptor. a Representative Western blot analysis for 5-HT1A and 5-HT2A in the hippocampus. b ZBD-2 (3 mg/kg) increased the levels of 5-HT1A in UCMS mice. c ZBD-2 (3 mg/kg) decreased the levels of 5-HT2A in UCMS mice. Effects of ZBD-2 were blocked by the PK11195 (3 mg/kg). Con Control, Veh Vehicle. n = 6 in each group. **p < 0.01 compared with the control mice, ## p < 0.01 compared with the UCMS Veh group
Effects of ZBD-2 on the Excitatory Synaptic Proteins
Inotropic glutamate receptors, including NMDA and AMPA receptors, play pivotal roles in emotional disorders (Myers et al. 2011). First, we examined the expression of AMPA and NMDA receptor subunits in hippocampus. Levels of GluN2B-containing NMDA receptors were significantly decreased in UCMS mice, but GluN2A-containing NMDA receptors were not affected (Fig. 5a–c). Furthermore, levels of PSD-95 (Fig. 5d), GluA1 (Fig. 5e), p-GluA1-Ser 845 (Fig. 5f), and p-GluA1-Ser 831 (Fig. 5g) were decreased in the UCMS mice. ZBD-2 (3 mg/kg) reversed these alterations in the hippocampus. Changes in CaMKIIs are related to stress and antidepressant response (Suenaga et al. 2004; Du et al. 2004). UCMS induced an up-regulation of CaMKII in the hippocampus, and this was reversed by ZBD-2 treatment (Fig. 5h). These effects of ZBD-2 on excitatory synaptic proteins were blocked by PK11195 (3 mg/kg).

Effects of ZBD-2 on the levels of excitatory synaptic proteins. a Representative Western blot analysis for excitatory synaptic proteins in the hippocampus. b ZBD-2 (3 mg/kg) did not affect the levels of GluN2A-containing NMDA receptors in the hippocampus of UCMS mice. Levels of GluN2B-containing NMDA receptors (c), PSD-95 (d), GluA1 (e), p-GluA1-Ser 845 (f), and p-GluA1-Ser 831 (g) were decreased in the hippocampus of UCMS mice. ZBD-2 (3 mg/kg) reversed the down-regulation of these proteins in UCMS mice. h ZBD-2 (3 mg/kg) reversed the up-regulation of CaMKII in hippocampus of UCMS mice. Effects of ZBD-2 were abolished by PK11195 (3 mg/kg). Con Control, Veh Vehicle. n = 6 in each group. **p < 0.01 compared with the control mice, ## p < 0.01 compared with the UCMS Veh group
Effects of ZBD-2 on GABAergic Proteins and iNOS in the UCMS Mice
Levels of GABAA-a2 were decreased in the hippocampus of UCMS mice, whereas the levels of GABA synthetase GAD67 were not changed (Fig. 6a, b). UCMS induced the neuroinflammation as shown by the increase in iNOS in the hippocampus (Fig. 6c). ZBD-2 (3 mg/kg) significantly increased the levels of GABAA-a2 and inhibited the iNOS in the hippocampus of UCMS mice. These effects were abolished by the treatment of PK11195 (3 mg/kg).

Effects of ZBD-2 on the levels of GAD67, GABAA-α2, and iNOS. a Representative bands of Western blot analysis for GAD67, GABAA-α2, and iNOS. b ZBD-2 (3 mg/kg) did not affect the levels of GAD67. c ZBD-2 (3 mg/kg) reversed the down-regulation of GABAA-α2 in the hippocampus of UCMS mice. d ZBD-2 (3 mg/kg) reversed the up-regulation of iNOS levels in the hippocampus of UCMS mice. Effects of ZBD-2 were abolished by PK11195 (3 mg/kg). Con Control, Veh Vehicle. n = 6 in each group. **p < 0.01 compared with the control mice, ## p < 0.01 compared with the UCMS Veh group
Discussion
In the present study, we evaluated the pharmacological profile of ZBD-2, a novel TSPO ligand, in UCMS mice. Our results demonstrate that ZBD-2 ameliorates the behavioral disorders associated with UCMS-induced anxiety- and depression-like behaviors. The underlying mechanisms involve the restoration of the monoamine levels, anti-inflammation, and regulation of GluN2B receptors and GABAergic receptors in the hippocampus.
UCMS paradigm recapitulates the role of socioenvironmental stressors precipitating depressive episodes and induces a syndrome reminiscent of depressive symptoms (Zhu et al. 2014). This model is particularly well suited to investigate causal factors involved in vulnerability to stress or in development of distinct alterations. UCMS was shown to elicit anxiety-like behaviors, increased aggressiveness, behavioral despair in the TST (depression), and modification of plasma corticosterone levels. All of these alterations were shown to be reversed by chronic exposure to antidepressant treatments in a time course, which parallels the clinical feature of these drugs (Ibarguen-Vargas et al. 2009). Thus, UCMS is suitable for the evaluation of anxiolytic and antidepressant drugs. In present study, we found that activity of UCMS mice was decreased in the open field test. Hypoactivity is a classic phenotype of depression. Hypoactivity may affect the performance in elevated plus maze test, forced swimming test, and tail suspension test. Therefore, we need to combine multiple behavioral tests to evaluate the anxiety- and depressive-like behaviors. In the present study, 3 mg/kg of ZBD-2 produced significant anxiolytic-like effects in the OF and EPM tests and anti-depressive-like effects in the FST and TST tests in the UCMS mice. Our previous study revealed the effective anxiolytic dose of ZBD-2 ranged from 0.15 and 1.5 mg/kg in chronic pain mice (Wang et al. 2015), which is similar to other TSPO agonists XBD173 (0.1–10 mg/kg in rat) and AC-5216 (the light/dark box test in rat 0.1–3 mg/kg) (Kita et al. 2004). The anxiolytic-like and antidepressant-like effects of ZBD-2 were blocked by PK11195, improving the evidence of targeting on the TSPO.
TSPO has been shown to be involved in stress responses. The expression of TSPO is up-regulated in microglia, astrocytes, and infiltrating macrophages, as well as occasionally in neurons in response to injury and neurodegenerative diseases (Girard et al. 2012). Our previous study revealed that TSPO in the amygdala was up-regulated in the mice suffering the chronic pain, and ZBD-2 treatment reversed this alteration (Wang et al. 2015). However, it has been reported that chronic stress induces a decrease in TSPO density in the brain and peripheral organs in rodents (Weizman and Gavish 1993). In the present UCMS model, TSPO in the hippocampus was down-regulated and restored to the basal level by the ZBD-2 treatment. Thus, the down-regulation of TSPO may be the response to the UCMS and the main cause for the emotional disorders.
Hippocampus is a critical brain region for the emotional disorder. Monoamines, including the 5-HT, DA, and NE, are the crucial neuromodulators in the development of emotional disorders (Elhwuegi 2004). Hippocampus is a critical brain region for the emotional disorder. Monoamines, including the 5-HT, DA, and NE, are the crucial neuromodulators in the development of emotional disorders (Elhwuegi 2004). In the present study, levels of monoamine were significantly decreased in the hippocampus of UCMS mice. This is consistent with the other reports (Liang et al. 2015). Repeated treatment of ZBD-2 restored the levels of 5-HT, NE, DA, and the metabolites in the hippocampus. Interestingly, we also found that levels of GABA were decreased in the UCMS mice, which was reversed by ZBD-2 treatment. Furthermore, ZBD-2 increased the levels of GABAA receptor; however, it did not affect the levels of GABA synthetase GDA67. These findings indicate that ZBD-2 attenuates the anxiety/depression dyad, at least partially, through modulation of monoamine levels in the brain.
Our previous study reported that levels of excitatory glutamate receptors and inhibitory GABA receptors were altered in the mice underlying acute stress (Tian et al. 2013). In the present study, administration of ZBD-2 reversed the down-regulation of excitatory glutamate receptors, including the NR2B-containing NMDARs, GluR1 receptors, PSD-95, and the up-regulation of CaMKII in the hippocampus from UCMS mice. It has been reported that changes in CaMKIIs are related to stress and antidepressant response (Suenaga et al. 2004; Du et al. 2004). Acute and repeated (4 days), but not chronic (14 days) stress significantly increases phospho-CaMKII levels without affecting the levels of total CaMKII in hippocampus (Suenaga et al. 2004). In the present study, we find that total levels of CaMKII are increased in the chronic unpredictable mild stress (UCMS) mice. The phospho-CaMKII levels are changeable due to the duration of stress. Thus, the anxiolytic-like and anti-depressive-like effects of ZBD-2 may be due to the maintenance of the balance between excitatory and inhibitory transmission in the hippocampus. We suppose that the changes of synaptic proteins are directly caused by stress and drug treatment.
Earlier studies have documented alterations in 5-HT receptors in several cortical and forebrain brain regions of subjects with major depression from PET imaging and human postmortem brain (Sullivan et al. 2015; Lothe et al. 2012). 5-HT1A and 5-HT2A play different roles in modulation of serotonin release or cellular function, and activation of 5-HT1A or blockade of 5-HT2A receptors seems to improve the clinical effects of SSRIs (Celada et al. 2004). Treated with ZBD-2 caused down-regulation of 5-HT2A but up-regulation of 5-HT1A in the hippocampus. Subsequent PK11195 treatment reversed the alteration of serotonin receptors, suggesting that the activity of ZBD-2 may indirectly regulate the serotonin synaptic transmission.
Lastly, it is reported that hippocampal-expressed TNFα serves as a therapeutic target for management of major depressive disorder, emphasizing the fundamental role of the inflammation in the development and maintenance of both disorders (Fasick et al. 2015). Treated with ZBD-2 inhibited the levels of iNOS in the hippocampus, suggesting the indirect anti-inflammation activities of ZBD-2.
The direct target of ZBD-2 is TSPO; however, we found that treatment of ZBD-2 could regulate the alteration of biogenic amines, CamKII, and neuroinflammation in the model mice. We could not conclude all these effects subsequent to the regulation of an overarching mitochondrial dysfunction. Further study is needed to specify the direct and indirect activities of ZBD-2 in the emotional disorders.
In summary, present findings provide the evidences that ZBD-2 attenuates the anxiety/depression dyad by regulating multiple molecules involved in the development of dyad. This study is helpful for the clinical treatment of emotional disorders by TSPO ligand.
Abbreviations
- 5-HT:
-
Serotonin
- AMPAR:
-
α-Amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor
- CNS:
-
Central nervous system
- DA:
-
Dopamine
- EPM:
-
Elevated plus maze
- FST:
-
Forced swim test
- GABA:
-
γ-Aminobutyric acid
- NE:
-
Norepinephrine
- NMDA:
-
N-Methyl-d-aspartate
- OF:
-
Open field
- SSRIs:
-
Selective serotonin reuptake inhibitors
- TSPO:
-
Translocator protein (18 kDa)
- TST:
-
Tail suspension test
- UCMS:
-
Chronic unpredictable mild stress
- ZBD-2:
-
N-Benzyl-N-ethyl-2-(7,8-dihydro-7-benzyl-8-oxo-2-phenyl-9H-purin-9-yl) acetamide
References
Albert, P. R., Vahid-Ansari, F., & Luckhart, C. (2014). Serotonin-prefrontal cortical circuitry in anxiety and depression phenotypes: Pivotal role of pre- and post-synaptic 5-HT1A receptor expression. Frontiers in Behavioral Neuroscience, 8, 199. doi:10.3389/fnbeh.2014.00199.
Anderson, I. M., Nutt, D. J., & Deakin, J. F. (2000). Evidence-based guidelines for treating depressive disorders with antidepressants: A revision of the 1993 British Association for Psychopharmacology guidelines. British Association for Psychopharmacology. Journal of Psychopharmacology, 14(1), 3–20.
Anholt, R. R., Pedersen, P. L., De Souza, E. B., & Snyder, S. H. (1986). The peripheral-type benzodiazepine receptor. Localization to the mitochondrial outer membrane. Journal of Biological Chemistry, 261(2), 576–583.
Belelli, D., & Lambert, J. J. (2005). Neurosteroids: Endogenous regulators of the GABA(A) receptor. Nature Reviews Neuroscience, 6(7), 565–575. doi:10.1038/nrn1703.
Bourin, M., Mocaer, E., & Porsolt, R. (2004). Antidepressant-like activity of S 20098 (agomelatine) in the forced swimming test in rodents: Involvement of melatonin and serotonin receptors. Journal of Psychiatry and Neuroscience, 29(2), 126–133.
Cates, L. N., Roberts, A. J., Huitron-Resendiz, S., & Hedlund, P. B. (2013). Effects of lurasidone in behavioral models of depression. Role of the 5-HT(7) receptor subtype. Neuropharmacology, 70, 211–217. doi:10.1016/j.neuropharm.2013.01.023.
Celada, P., Puig, M., Amargos-Bosch, M., Adell, A., & Artigas, F. (2004). The therapeutic role of 5-HT1A and 5-HT2A receptors in depression. Journal of Psychiatry and Neuroscience, 29(4), 252–265.
De Benedetto, G. E., Fico, D., Pennetta, A., Malitesta, C., Nicolardi, G., Lofrumento, D. D., et al. (2014). A rapid and simple method for the determination of 3,4-dihydroxyphenylacetic acid, norepinephrine, dopamine, and serotonin in mouse brain homogenate by HPLC with fluorimetric detection. Journal of Pharmaceutical and Biomedical Analysis, 98, 266–270. doi:10.1016/j.jpba.2014.05.039.
Dranovsky, A., & Hen, R. (2006). Hippocampal neurogenesis: Regulation by stress and antidepressants. Biological Psychiatry, 59(12), 1136–1143. doi:10.1016/j.biopsych.2006.03.082.
Du, J., Szabo, S. T., Gray, N. A., & Manji, H. K. (2004). Focus on CaMKII: A molecular switch in the pathophysiology and treatment of mood and anxiety disorders. International Journal of Neuropsychopharmacology, 7(3), 243–248. doi:10.1017/S1461145704004432.
Elhwuegi, A. S. (2004). Central monoamines and their role in major depression. Progress in Neuropsychopharmacology and Biological Psychiatry, 28(3), 435–451. doi:10.1016/j.pnpbp.2003.11.018.
Fasick, V., Spengler, R. N., Samankan, S., Nader, N. D., & Ignatowski, T. A. (2015). The hippocampus and TNF: Common links between chronic pain and depression. Neuroscience and Biobehavioral Reviews, 53, 139–159. doi:10.1016/j.neubiorev.2015.03.014.
Gavish, M., Bachman, I., Shoukrun, R., Katz, Y., Veenman, L., Weisinger, G., et al. (1999). Enigma of the peripheral benzodiazepine receptor. Pharmacological Reviews, 51(4), 629–650.
Gavish, M., Laor, N., Bidder, M., Fisher, D., Fonia, O., Muller, U., et al. (1996). Altered platelet peripheral-type benzodiazepine receptor in posttraumatic stress disorder. Neuropsychopharmacology, 14(3), 181–186. doi:10.1016/0893-133X(95)00078-R.
Girard, C., Liu, S., Adams, D., Lacroix, C., Sineus, M., Boucher, C., et al. (2012). Axonal regeneration and neuroinflammation: Roles for the translocator protein 18 kDa. Journal of Neuroendocrinology, 24(1), 71–81. doi:10.1111/j.1365-2826.2011.02215.x.
Guo, Y. Y., Liu, S. B., Cui, G. B., Ma, L., Feng, B., Xing, J. H., et al. (2012). Acute stress induces down-regulation of large-conductance Ca2+-activated potassium channels in the lateral amygdala. Journal of Physiology, 590(Pt 4), 875–886. doi:10.1113/jphysiol.2011.223784.
Haase, J., & Brown, E. (2015). Integrating the monoamine, neurotrophin and cytokine hypotheses of depression—A central role for the serotonin transporter? Pharmacology & Therapeutics, 147, 1–11. doi:10.1016/j.pharmthera.2014.10.002.
Ibarguen-Vargas, Y., Surget, A., Vourc’h, P., Leman, S., Andres, C. R., Gardier, A. M., et al. (2009). Deficit in BDNF does not increase vulnerability to stress but dampens antidepressant-like effects in the unpredictable chronic mild stress. Behavioural Brain Research, 202(2), 245–251. doi:10.1016/j.bbr.2009.03.040.
Kallarackal, A. J., Kvarta, M. D., Cammarata, E., Jaberi, L., Cai, X., Bailey, A. M., et al. (2013). Chronic stress induces a selective decrease in AMPA receptor-mediated synaptic excitation at hippocampal temporoammonic-CA1 synapses. Journal of Neuroscience, 33(40), 15669–15674. doi:10.1523/JNEUROSCI.2588-13.2013.
Khoshnoodi, M., Fakhraei, N., & Dehpour, A. R. (2015). Possible involvement of nitric oxide in antidepressant-like effect of silymarin in male mice. Pharmaceutical Biology, 53(5), 739–745. doi:10.3109/13880209.2014.942787.
Kita, A., Kohayakawa, H., Kinoshita, T., Ochi, Y., Nakamichi, K., Kurumiya, S., et al. (2004). Antianxiety and antidepressant-like effects of AC-5216, a novel mitochondrial benzodiazepine receptor ligand. British Journal of Pharmacology, 142(7), 1059–1072. doi:10.1038/sj.bjp.0705681.
Koo, J. W., & Duman, R. S. (2008). IL-1beta is an essential mediator of the antineurogenic and anhedonic effects of stress. Proceedings of the National Academy of Sciences, 105(2), 751–756. doi:10.1073/pnas.0708092105.
Li, X. B., Guo, H. L., Shi, T. Y., Yang, L., Wang, M., Zhang, K., et al. (2015). Neuroprotective effects of a novel translocator protein (18 kDa) ligand, ZBD-2, against focal cerebral ischemia in vivo and NMDA-induced neurotoxicity in vitro. Clinical and Experimental Pharmacology and Physiology. doi:10.1111/1440-1681.12460.
Li, K., Zhou, T., Liao, L., Yang, Z., Wong, C., Henn, F., et al. (2013). betaCaMKII in lateral habenula mediates core symptoms of depression. Science, 341(6149), 1016–1020. doi:10.1126/science.1240729.
Liang, B. F., Huang, F., Wang, H. T., Wang, G. H., Yuan, X., Zhang, M. Z., et al. (2015). Involvement of norepinephrine and serotonin system in antidepressant-like effects of hederagenin in the rat model of unpredictable chronic mild stress-induced depression. Pharmaceutical Biology, 53(3), 368–377. doi:10.3109/13880209.2014.922586.
Liu, B., Xu, C., Wu, X., Liu, F., Du, Y., Sun, J., et al. (2015). Icariin exerts an antidepressant effect in an unpredictable chronic mild stress model of depression in rats and is associated with the regulation of hippocampal neuroinflammation. Neuroscience, 294, 193–205. doi:10.1016/j.neuroscience.2015.02.053.
Lothe, A., Saoud, M., Bouvard, S., Redoute, J., Lerond, J., & Ryvlin, P. (2012). 5-HT(1A) receptor binding changes in patients with major depressive disorder before and after antidepressant treatment: A pilot [(1)(8)F]MPPF positron emission tomography study. Psychiatry Research, 203(1), 103–104. doi:10.1016/j.pscychresns.2011.09.001.
Murata, Y., Yanagihara, Y., Mori, M., Mine, K., & Enjoji, M. (2015). Chronic treatment with tandospirone, a serotonin 1A receptor partial agonist, inhibits psychosocial stress-induced changes in hippocampal neurogenesis and behavior. Journal of Affective Disorders, 180, 1–9. doi:10.1016/j.jad.2015.03.054.
Myers, K. M., Carlezon, W. A, Jr., & Davis, M. (2011). Glutamate receptors in extinction and extinction-based therapies for psychiatric illness. Neuropsychopharmacology, 36(1), 274–293. doi:10.1038/npp.2010.88.
Nelson, E. M., & Philbrick, A. M. (2012). Avoiding serotonin syndrome: The nature of the interaction between tramadol and selective serotonin reuptake inhibitors. Annals of Pharmacotherapy, 46(12), 1712–1716. doi:10.1345/aph.1Q748.
Nothdurfter, C., Baghai, T. C., Schule, C., & Rupprecht, R. (2012). Translocator protein (18 kDa) (TSPO) as a therapeutic target for anxiety and neurologic disorders. European Archives of Psychiatry and Clinical Neuroscience, 262(Suppl 2), S107–S112. doi:10.1007/s00406-012-0352-5.
Papadopoulos, V., Lecanu, L., Brown, R. C., Han, Z., & Yao, Z. X. (2006). Peripheral-type benzodiazepine receptor in neurosteroid biosynthesis, neuropathology and neurological disorders. Neuroscience, 138(3), 749–756. doi:10.1016/j.neuroscience.2005.05.063.
Rocca, P., Beoni, A. M., Eva, C., Ferrero, P., Zanalda, E., & Ravizza, L. (1998). Peripheral benzodiazepine receptor messenger RNA is decreased in lymphocytes of generalized anxiety disorder patients. Biological Psychiatry, 43(10), 767–773.
Rupprecht, R., Rammes, G., Eser, D., Baghai, T. C., Schule, C., Nothdurfter, C., et al. (2009). Translocator protein (18 kD) as target for anxiolytics without benzodiazepine-like side effects. Science, 325(5939), 490–493. doi:10.1126/science.1175055.
Schmidt, M. V., Trumbach, D., Weber, P., Wagner, K., Scharf, S. H., Liebl, C., et al. (2010). Individual stress vulnerability is predicted by short-term memory and AMPA receptor subunit ratio in the hippocampus. Journal of Neuroscience, 30(50), 16949–16958. doi:10.1523/JNEUROSCI.4668-10.2010.
Suenaga, T., Morinobu, S., Kawano, K., Sawada, T., & Yamawaki, S. (2004). Influence of immobilization stress on the levels of CaMKII and phospho-CaMKII in the rat hippocampus. International Journal of Neuropsychopharmacology, 7(3), 299–309. doi:10.1017/S1461145704004304.
Sullivan, G. M., Oquendo, M. A., Milak, M., Miller, J. M., Burke, A., Ogden, R. T., et al. (2015). Positron emission tomography quantification of serotonin(1A) receptor binding in suicide attempters with major depressive disorder. JAMA Psychiatry, 72(2), 169–178. doi:10.1001/jamapsychiatry.2014.2406.
Tian, Z., Wang, Y., Zhang, N., Guo, Y. Y., Feng, B., Liu, S. B., et al. (2013). Estrogen receptor GPR30 exerts anxiolytic effects by maintaining the balance between GABAergic and glutamatergic transmission in the basolateral amygdala of ovariectomized mice after stress. Psychoneuroendocrinology, 38(10), 2218–2233. doi:10.1016/j.psyneuen.2013.04.011.
Tomaz, V. S., Cordeiro, R. C., Costa, A. M., de Lucena, D. F., Nobre Junior, H. V., de Sousa, F. C., et al. (2014). Antidepressant-like effect of nitric oxide synthase inhibitors and sildenafil against lipopolysaccharide-induced depressive-like behavior in mice. Neuroscience, 268, 236–246. doi:10.1016/j.neuroscience.2014.03.025.
Vasquez, C. E., Riener, R., Reynolds, E., & Britton, G. B. (2014). NMDA receptor dysregulation in chronic state: A possible mechanism underlying depression with BDNF downregulation. Neurochemistry International, 79, 88–97. doi:10.1016/j.neuint.2014.09.007.
Veenman, L., & Gavish, M. (2006). The peripheral-type benzodiazepine receptor and the cardiovascular system. Implications for drug development. Pharmacology & Therapeutics, 110(3), 503–524. doi:10.1016/j.pharmthera.2005.09.007.
Wang, D. S., Tian, Z., Guo, Y. Y., Guo, H. L., Kang, W. B., Li, S., et al. (2015). Anxiolytic-like effects of translocator protein (TSPO) ligand ZBD-2 in an animal model of chronic pain. Molecular Pain, 11, 16. doi:10.1186/s12990-015-0013-6.
Weizman, R., & Gavish, M. (1993). Molecular cellular and behavioral aspects of peripheral-type benzodiazepine receptors. Clinical Neuropharmacology, 16(5), 401–417.
Willner, P., Towell, A., Sampson, D., Sophokleous, S., & Muscat, R. (1987). Reduction of sucrose preference by chronic unpredictable mild stress, and its restoration by a tricyclic antidepressant. Psychopharmacology (Berl), 93(3), 358–364.
Zhu, S., Wang, J., Zhang, Y., Li, V., Kong, J., He, J., et al. (2014). Unpredictable chronic mild stress induces anxiety and depression-like behaviors and inactivates AMP-activated protein kinase in mice. Brain Research, 1576, 81–90. doi:10.1016/j.brainres.2014.06.002.
Zorumski, C. F., Paul, S. M., Izumi, Y., Covey, D. F., & Mennerick, S. (2013). Neurosteroids, stress and depression: Potential therapeutic opportunities. Neuroscience and Biobehavioral Reviews, 37(1), 109–122. doi:10.1016/j.neubiorev.2012.10.005.
Acknowledgments
This work was supported by the National Natural Science Foundation of China Nos. 81325022, 31470052, 2011ZXJ09106-01C, and Clinical Science and Technology Project Funding in Jiangsu Province (No. BL2012002).
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Dong-sheng Wang, Jing Han and Shuo Li have contributed equally to this work.
Rights and permissions
About this article

Cite this article
Wang, Ds., Han, J., Li, S. et al. Antidepressant-Like and Anxiolytic-Like Effects of ZBD-2, a Novel Ligand for the Translocator Protein (18 kDa). Neuromol Med 19, 57–68 (2017). https://doi.org/10.1007/s12017-016-8425-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12017-016-8425-7