
Abstract
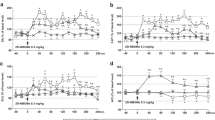
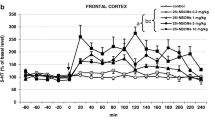
The effects of diazepam quipazine, lysergic acid diethylamide (LSD), and 2,5-dimethoxy-4-methylamphetamine (DOM) were examined on a conditioned suppression paradigm. Food-deprived rats were trained to drink a liquid diet from a tube. Subsequently, intermittent 7-s tones were presented during the daily 10-min sessions, the tube being electrified during the last 5 s of each tone. The subject gradually learned to suppress contact with the tube during the tone periods to a low stable level (punished responding) and consumed stable volumes of the liquid diet during the silent periods (unpunished responding). Treatment with diazepam caused large increases (1,000% of control) in punished responding. The hallucinogens produced only modest increases (200–300%), while quipazine did not significantly increase punished responding. Metergoline pretreatment (0.1–2.0 mg/kg, 180 min) had no effect on punished responding itself, and there was no significant alteration of the diazepam dose-response pattern. The weak increase in punished responding by LSD was antagonized by metergoline, but the interaction between metergoline and DOM was variable and inconsistent. Diazepam, quipazine, LSD, and DOM caused dose-dependent decreases in unpunished responding (fluid intake). Metergoline alone decreased unpunished responding only at 2.0 mg/kg. Metergoline pretreatment (1.0 mg/kg) only slightly antagonized the LSD effect on unpunished behavior, but shifted the dose-response curves of DOM and quipazine for decrease in fluid intake to the right approximately eight fold. On the contrary, the dose-response curve of diazepam to decrease fluid intake was shifted to the left by metergoline pretreatment. These data suggest that altered activity of brain serotonin (5-HT) neurons is not responsible for the dramatic increase in punished responding by diazepam. The hallucinogens, quipazine, and diazepam all produce a decrease in unpunished responding, but they appear to do so by different neuropharmacological mechanisms. In addition, there may be at least slight differences in the mechanism by which LSD produces its effects as compared with that of quipazine and DOM.
Similar content being viewed by others

References
Aghajanian GK, Haigler HJ, Bennett JL (1975) Amine receptors in CNS. III. 5-Hydroxytryptamine in brain. In: Iversen L, Iversen S, Snyder S (eds) Handbook of Psychopharmacology, vol VI. Plenum, New York, pp 63–96
Andén NE, Corrodi H, Fuxe K, Meed JL (1974) Hallucinogenic phenethylamines: Interactions with serotonin turnover and receptors. Eur J Pharmacol 25:176–184
Beer B, Chasin M, Clody DE, Vogel JR, Horovitz ZP (1972) Cyclic adenosine monophosphate phosphodiesterase in brain: Effect on anxiety. Science 176:428–430
Blakely TA, Parker LF (1973) The effects of p-chlorophenylalanine on experimentally induced conflict behavior. Pharmacol Biochem Behav 1:609–613
Commissaris RL, Lyness WH, Rech RH (1981a) The effects of d-lysergic acid diethylamide (LSD), 2,5-dimethoxy-4-methylamphetamine (DOM), pentobarbital and methaqualone on punished behavior in control and 5,7-dihydroxytryptamine-treated rats. Pharmacol Biochem Behav 14:617–623
Commisaris RL, Lyness WH, Rech RH, Moore KE (1981b) Differential antagonism by metergoline of the behavioral effects of indolealkylamine and phenethylamine hallucinogens in the rat. J Pharmacol Exp Ther 219:170–174
Cook L, Sepinwall J (1975) Behavioral analysis of the effects and mechanism of action of benzodiazepines In: Costa E, Greengard P (eds) Mechanism of action of benzodiazepines. Raven Press, New York, pp 1–28
Ford RD, Rech RH, Commissaris RL, Mayer L (1979) Effects of acute and chronic interactions of diazepam and d-amphetamine on punished and unpunished behavior of rats. Psychopharmacology 65:197–204
Geller I, Blum K (1970) The effects of 5-HTP on p-chlorophenylalanine (PCPA) attenuation of “conflict” behavior. Eur J Pharmacol 9:319–324
Geller I, Hartmann RJ, Croy DJ (1974) Attenuation of conflict behavior with cinanserin, a serotonin antagonist: Reversal of the effect with 5-hydroxytryptophan and α-methyltryptamine. Res Commun Chem Pathol Pharmacol 7:165–174
Graeff FG (1974) Tryptamine antagonists and punished behavior., J Pharmacol Exp Ther 189:344–350
Kilts CD, Commissaris RL, Rech RH (1981) Comparison of anti-conflict effects in three experimental animal models of anxiety. Psychopharmacology 74:290–296
Kuhn DM, White FJ, Appel JB (1978) The discriminative stimulus properties of LSD: Mechanisms of action. Neuropharmacology 17:257–263
Samanin R, Bendotti C, Candelaresi G, Garattini S (1978) Specificity of serotonergic involvement in the decrease of food intake induced by quipazine in the rat. Life Sci 21:1259–1266
Schoenfeld RI (1976) Lysergic acid diethylamide-and mescaline-induced attenuation of the effect of punishment in the rat. Science 192:801–803
Sloviter R, Drust E, Dammiano B, Connor J (1980) A common mechanism for lysergic acid, indolealkylamine and phenethylamine hallucinogens: Serotonergic mediation of behavioral effects in rats. J Pharmacol Exp Ther 214:231–238
Stein L, Wise CD, Belluzzi JD (1975) Effect of benzodiazepines on central serotonergic mechanisms. In: Costa E, Greengard P (eds) Mechanisms of action of benzodiazepines. Raven Press New York, pp 29–44
Tye NC, Everitt BJ, Iversen SD (1977) 5-Hydroxytryptamine and punishment. Nature 268:741–743
Winter JC (1979) Quipazine-induced stimulus control in the rat. Psychopharmacology 60:265–269
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Commissaris, R.L., Rech, R.H. Interactions of metergoline with diazepam, quipazine, and hallucinogenic drugs on a conflict behavior in the rat. Psychopharmacology 76, 282–285 (1982). https://doi.org/10.1007/BF00432562
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1007/BF00432562