Complete Chloroplast Genomes of Three Medicinal Alpinia Species: Genome Organization, Comparative Analyses and Phylogenetic Relationships in Family Zingiberaceae


Abstract
:1. Introduction
2. Results and Discussion
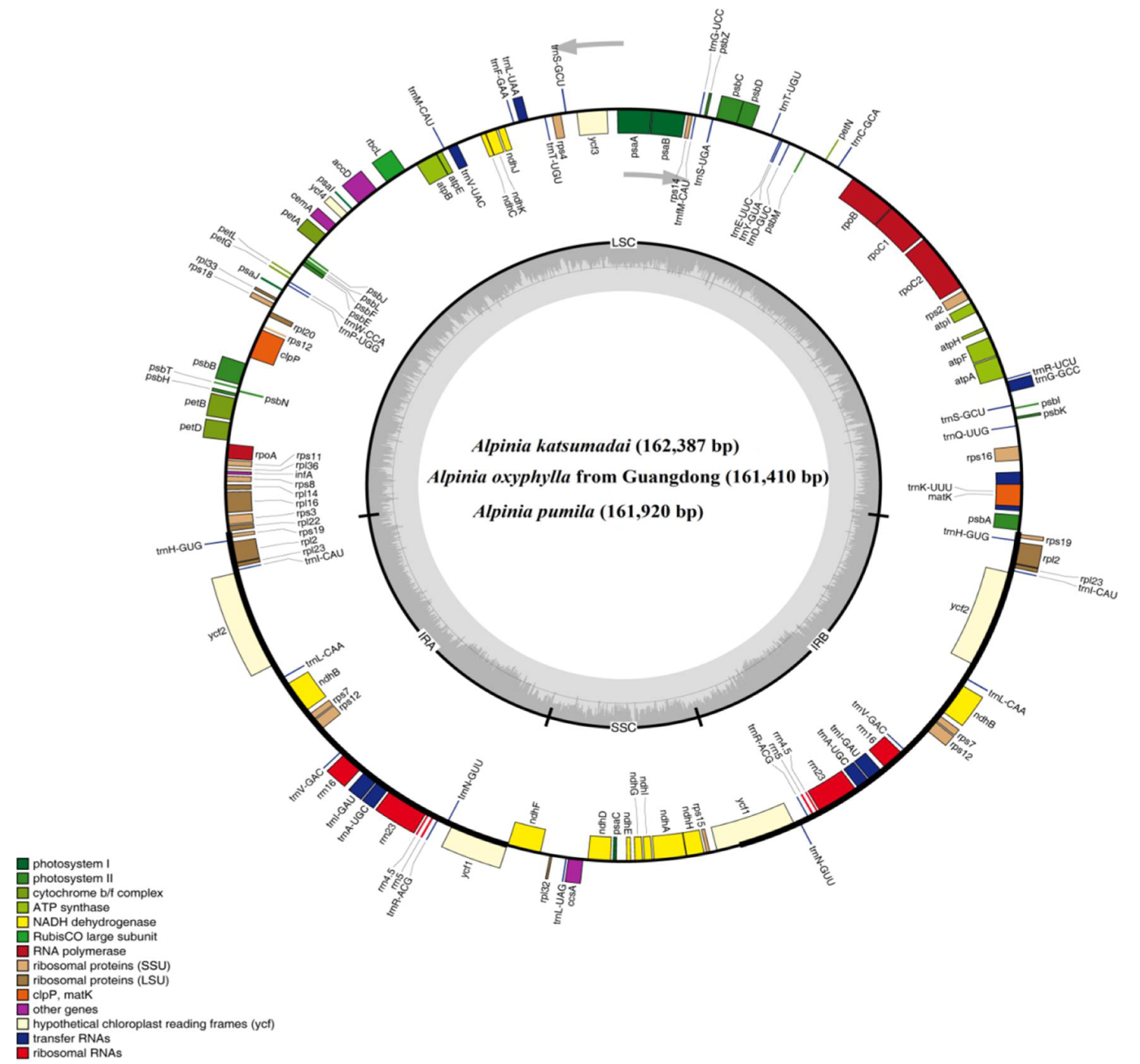
2.1. The Chloroplast Genome Features of Alpinia Species
2.2. Codon Usage and Predicted RNA Editing Sites Analyses
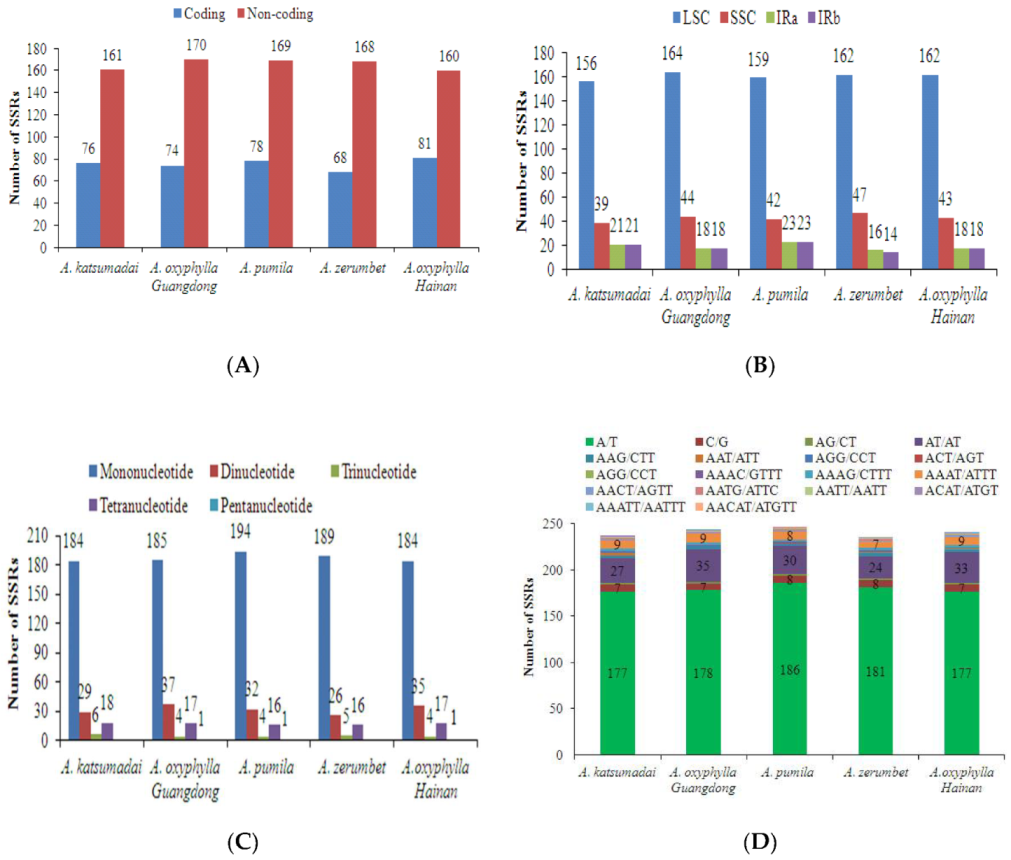
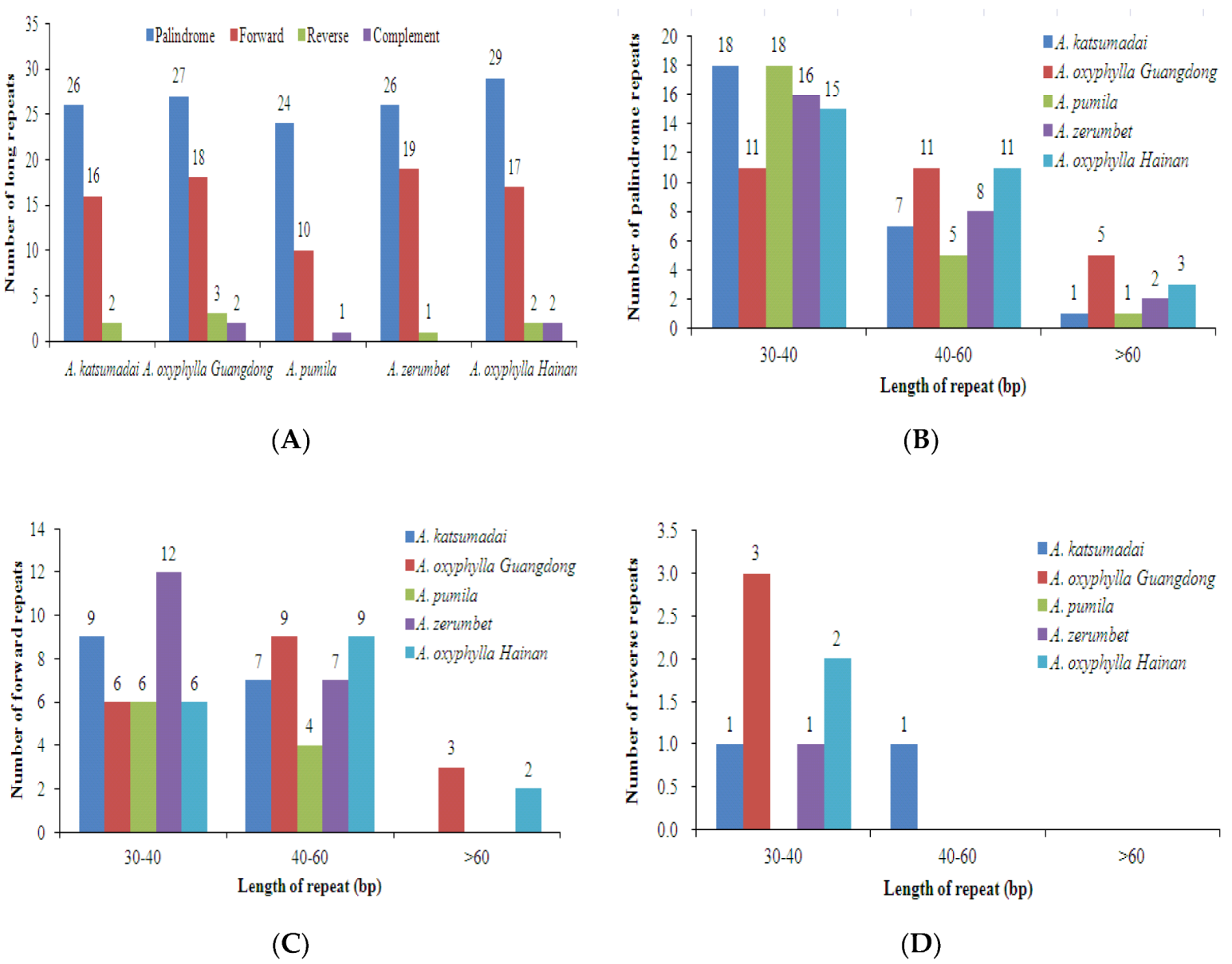
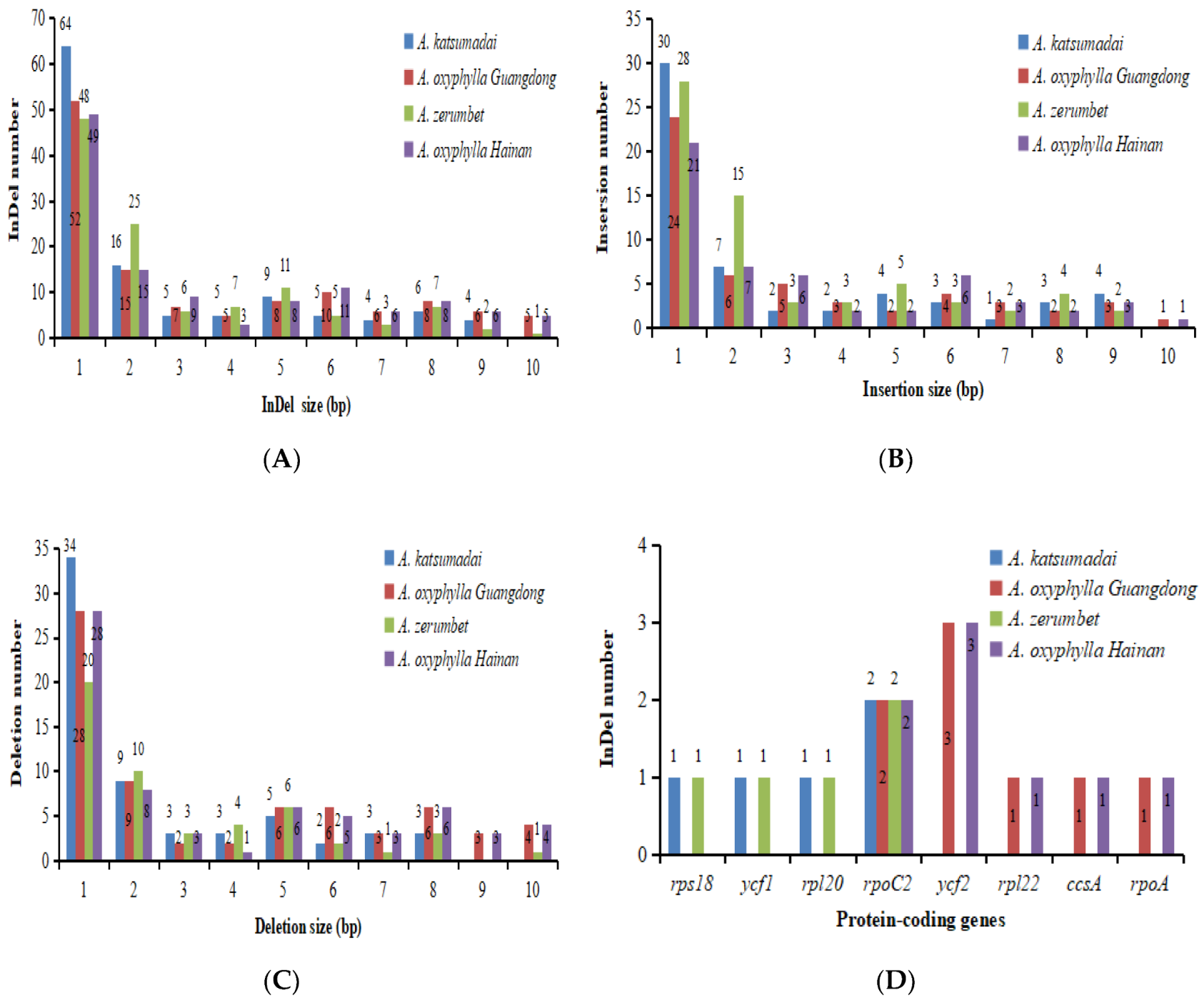
2.3. SSRs and Long Repeats Analyses
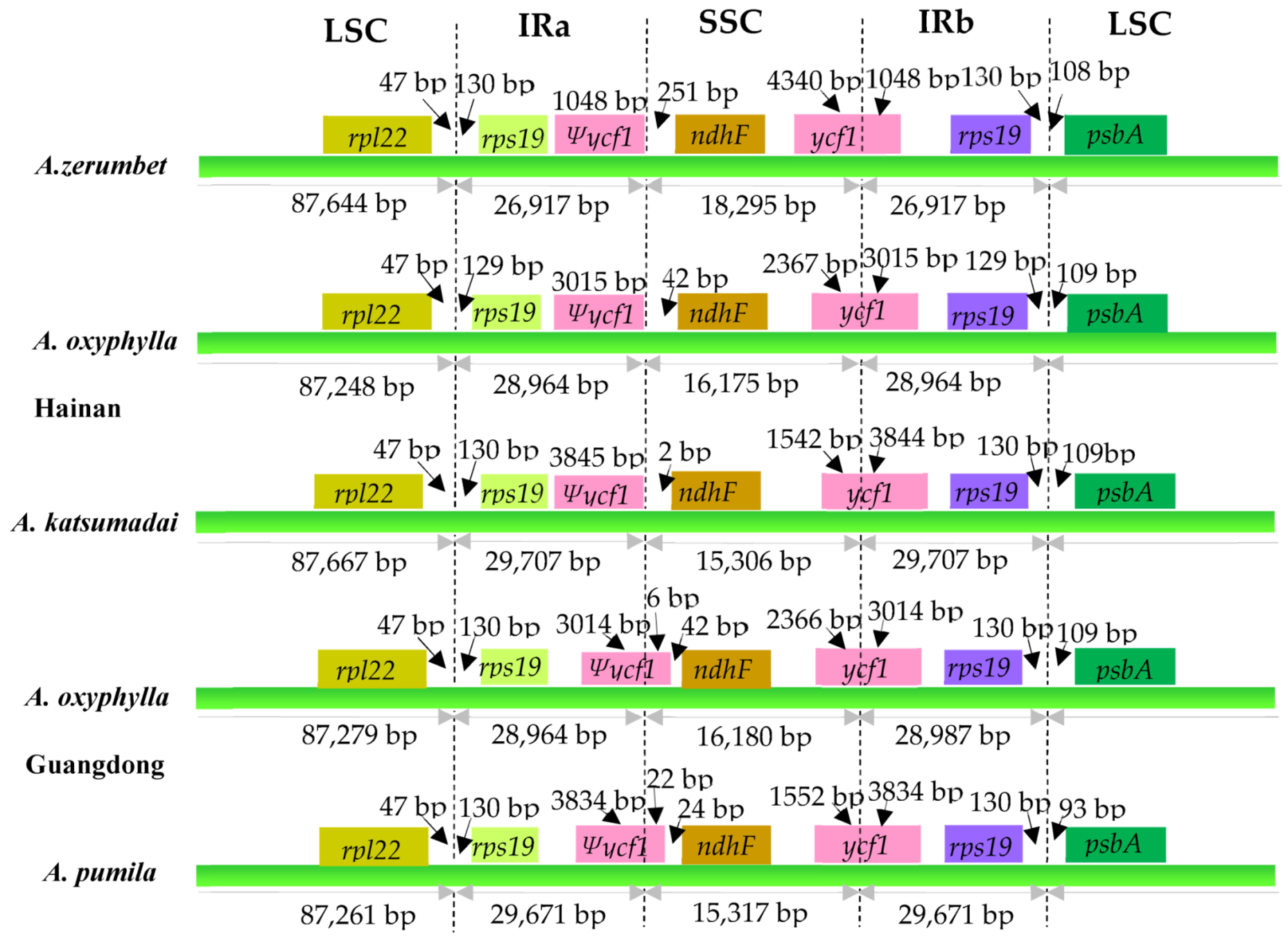
2.4. IR Contraction and Expansion Analyses
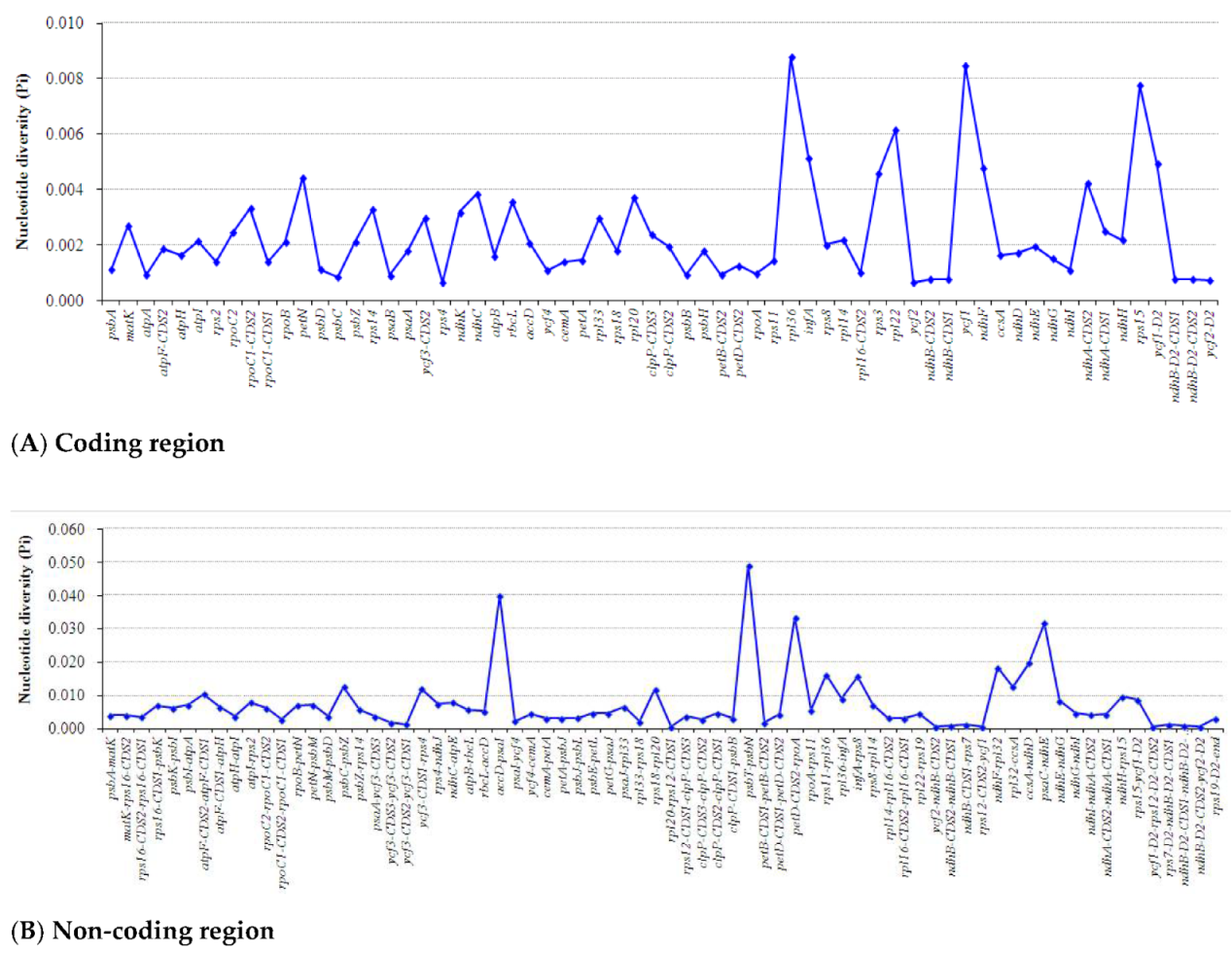
2.5. Divergence Hotspot Regions Analyses
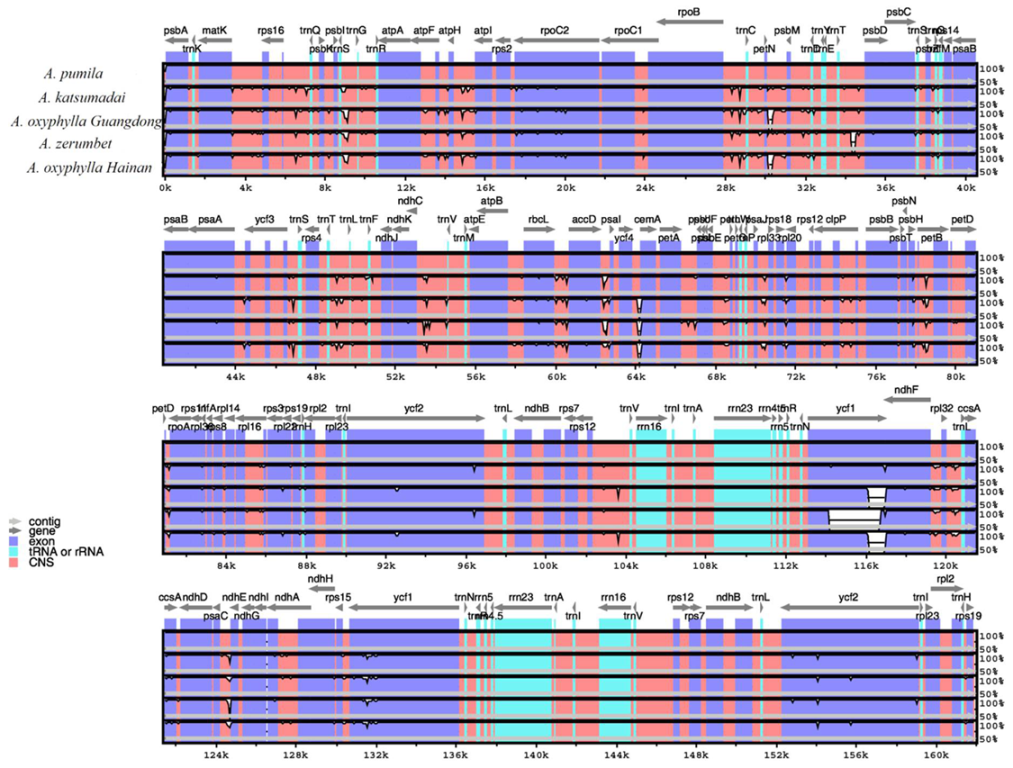
2.6. Interspecific Analyses of Alpinia Chloroplast Genomes
2.7. Intraspecific Analyses of two Chloroplast Genomes of A. oxyphylla
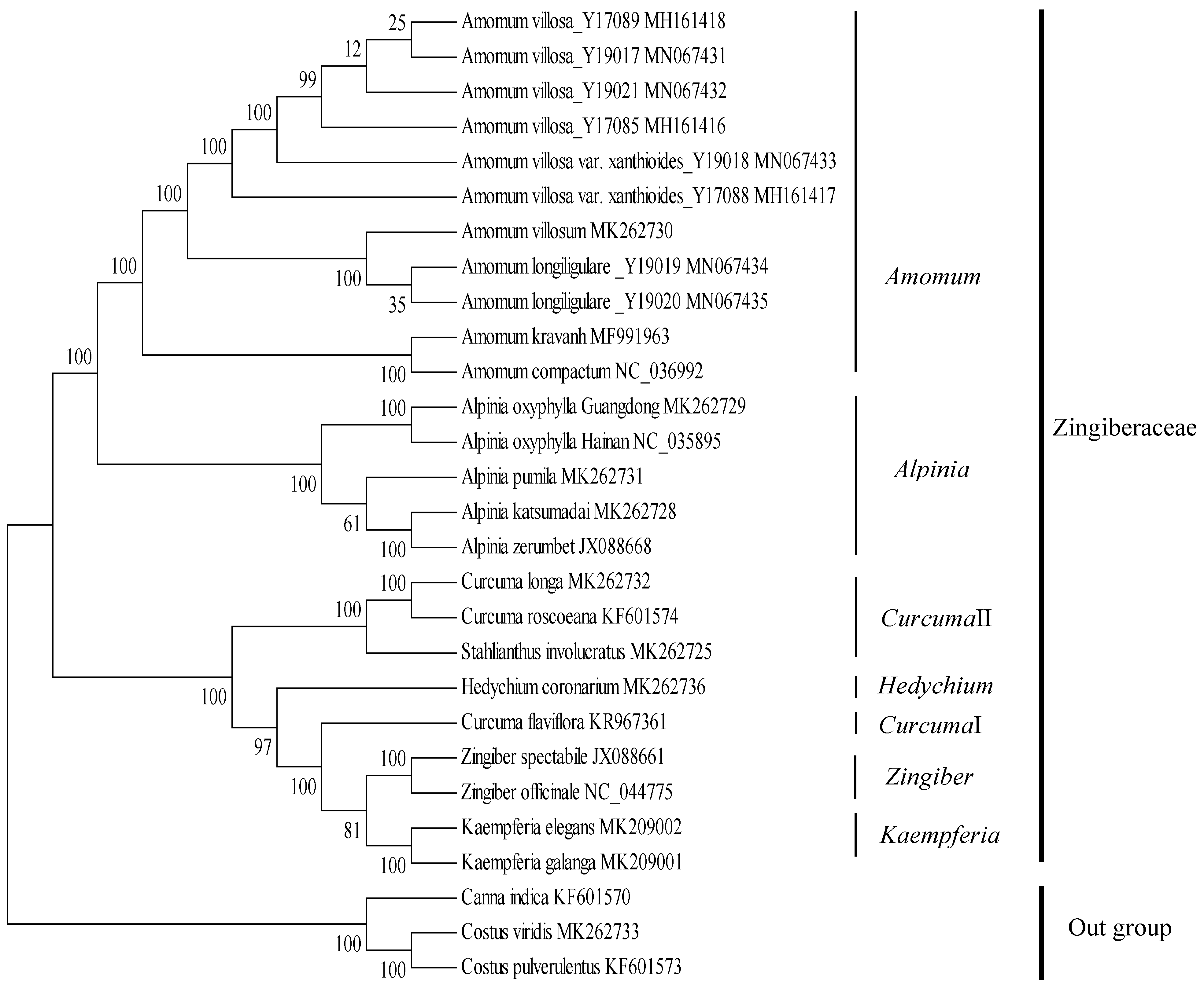
2.8. Phylogenetic Analyses
3. Materials and Methods
3.1. Plant Material and DNA Extraction
3.2. Chloroplast Genome Sequencing and Assembly
3.3. Chloroplast Genome Annotation and Structure Analysis
3.4. Codon Usage and Prediction of RNA Editing Sites
3.5. Genome Comparison and Divergence Analyses
3.6. Phylogenetic Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| bp | base pairs |
| BLAST | Basic Local Alignment Search Tool |
| NCBI | National Center for Biotechnology Information |
| IR | Inverted repeat |
| LSC | Large single copy |
| SSC | Small single copy |
| SSRs | Simple Sequence Repeats |
| A | Adenine |
| T | Thymine |
| G | Guanine |
| C | Cytosine |
| DNA | Deoxyribonucleic Acid |
| RNA | Ribonucleic acid |
| tRNA | Transfer RNA |
| rRNA | Ribosomal RNA |
| SNPs | Single-Nucleotide Polymorphisms |
| Indels | Insertion/deletions |
References
- Wu, D.; Larsen, K. Zingiberaceae. In Flora of China; Science Press: Beijing, China, 2000; Volume 24, pp. 322–377. [Google Scholar]
- Wu, D.; Liu, N.; Ye, Y. The Zingiberaceous Resources in China; Huazhong University of Science and Technology University Press: Wuhan, China, 2016; Volume 1, pp. 12, 20, 30, 33. [Google Scholar]
- Branney, T.M.E. Hardy Gingers: Including Hedychium, Roscoea and Zingiber; Timber Press, Inc.: Porland, OR, USA, 2005; pp. 41–63. [Google Scholar]
- Zhang, J.Q.; Li, Y.B. Zhongguo Yaoyong Jiangke Zhiwu; China Medical Science Press: Beijing, China, 2015; Volume 1, pp. 133–220. [Google Scholar]
- Pogačar, M.Š.; Klančnik, A.; Bucar, F.; Langerholc, T.; Možina, S.S. Alpinia katsumadai extracts inhibit adhesion and invasion of campylobacter jejuni in animal and human foetal small intestine cell lines. Phytother. Res. 2015, 29, 1585–1589. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Y.; Huang, S.S.; Lee, M.M.; Deng, J.S.; Huang, G.J. Anti-inflammatory activities of cardamonin from Alpinia katsumadai through heme oxygenase-1 induction and inhibition of NF-κB and MAPK signaling pathway in the carrageenan-induced pawedema. Int. Immunopharmacol. 2015, 25, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.J.; Lee, S.J.; Lee, S.; Jung, K.; Lee, S.W.; Rho, M.C. Acyclic triterpenoids from Alpinia katsumadai inhibit IL-6-induced STAT3 activation. Molecules 2017, 22, 1611. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.B.; Yang, C.S.; Luo, J.G.; Zhang, C.; Luo, J.; Yang, M.H.; Kong, L.Y. Experimental and theoretical calculation studies on the structure elucidation and absolute configuration of calyxins from Alpinia katsumadai. Fitoterapia 2017, 119, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Li, H.L.; Tan, Y.F.; Guan, W.W.; Zhang, J.Q.; Li, Y.H.; Zhao, Y.S.; Qin, Z.M. Different accumulation profiles of multiple components between pericarp and seed of Alpinia oxyphylla capsular fruit as determined by UFLC-MS/MS. Molecules 2014, 19, 4510–4523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, Y.; Cheng, X.; Jing, H.; Yan, T.; Xiao, F.; Wu, B.; Bi, K.; Jia, Y. Comparative pharmacokinetic study of the components in Alpinia oxyphylla Miq.-Schisandra chinensis (Turcz.) Baill. herb pair and its single herb between normal and Alzheimer’s disease rats by UPLC-MS/MS. J. Pharm. Biomed. Anal. 2020, 177, 112874. [Google Scholar] [CrossRef] [PubMed]
- Delectis Florae Reipublicae Popularis Sinicae Agendae Academiae Sinicae Edita. In Flora Reipublicae Popularis Sinicae; Science press: Beijing, China, 1981.
- Kress, W.J.; Prince, L.M.; Williams, K.J. The phylogeny and a new classification of the gingers (Zingiberaceae) evidence from molecular data. Am. J. Bot. 2002, 89, 1682–1696. [Google Scholar] [CrossRef]
- Gao, B.; Yuan, L.; Tang, T.; Hou, J.; Pan, K.; Wei, N. The complete chloroplast genome sequence of Alpinia oxyphylla Miq. and comparison analysis within the Zingiberaceae family. PLoS ONE 2019, 14, e0218817. [Google Scholar] [CrossRef]
- Wicke, S.; Schneeweiss, G.M.; DePamphilis, C.W.; Muller, K.F.; Quandt, D. The evolution of the plastid chromosome in land plants: Gene content, gene order, gene function. Plant Mol. Biol. 2011, 76, 273–297. [Google Scholar] [CrossRef] [Green Version]
- Daniell, H.; Lin, C.S.; Yu, M.; Chang, W.J. Chloroplast genomes: Diversity, evolution, and applications in genetic engineering. Genome Biol. 2016, 17, 134. [Google Scholar] [CrossRef] [Green Version]
- Eid, J.; Fehr, A.; Gray, J.; Luong, K.; Lyle, J.; Otto, G.; Peluso, P.; Rank, D.; Baybayan, P.; Bettman, B.; et al. Real-time DNA sequencing from single polymerase molecules. Science 2009, 323, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Ferrarini, M.; Moretto, M.; Ward, J.A.; Surbanovski, N.; Stevanovic, V.; Giongo, L.; Viola, R.; Cavalieri, D.; Velasco, R.; Cestaro, A.; et al. An evaluation of the PacBio RS platform for sequencing and de novo assembly of a chloroplast genome. BMC Genom. 2013, 14, 670. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Gui, S.; Guan, Z.; Pan, L.; Wang, S.; Ke, W.; Liang, D.; Ding, Y. A precise chloroplast genome of Nelumbo nucifera (Nelumbonaceae) evaluated with Sanger, Illumina MiSeq, and PacBio RS II sequencing platforms: Insight into the plastid evolution of basal eudicots. BMC Plant Biol. 2014, 14, 289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chomicki, G.; Renner, S.S. Watermelon origin solved with molecular phylogenetics including Linnaen material: Another example of museomics. New Phytol. 2015, 205, 526–532. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Li, Q.; Hu, Z.; Li, X.; Chen, S. The complete Amomum kravanh chloroplast genome sequence and phylogenetic analysis of the commelinids. Molecules 2017, 22, 1875. [Google Scholar] [CrossRef] [Green Version]
- Lin, M.; Qi, X.; Chen, J.; Sun, L.; Zhong, Y.; Fang, J.; Hu, C. The complete chloroplast genome sequence of Actinidia arguta using the PacBio RSII platform. PLoS ONE 2018, 13, e0197393. [Google Scholar]
- Zhou, Y.; Nie, J.; Xiao, L.; Hu, Z.; Wang, B. Comparative chloroplast genome analysis of rhubarb botanical origins and development of specific identification markers. Molecules 2018, 23, 2811. [Google Scholar] [CrossRef] [Green Version]
- Li, D.M.; Zhao, C.Y.; Liu, X.F. Complete chloroplast genome sequences of Kaempferia galanga and Kaempferia elegans: Molecular structures and comparative analysis. Molecules 2019, 24, 474. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.; Chen, X.; Nie, L.; Sun, W.; Hu, H.; Lin, Y.; Li, H.; Zheng, X.; Song, J.; Yao, H. Comparison and phylogenetic analysis of chloroplast genomes of three medicinal and edible Amomum species. Int. J. Mol. Sci. 2019, 20, 4040. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.; Nie, L.; Sun, W.; Xu, Z.; Wang, Y.; Yu, J.; Song, J.; Yao, H. Comparative and phylogenetic analyses of ginger (Zingiber officinale) in the family Zingiberaceae based on the complete chloroplast genome. Plants 2019, 8, 283. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zuo, Y.; Zhu, X.; Liao, S.; Ma, J. Complete chloroplast genomes and comparative analysis of sequences evolution among seven Aristolochia (Aristolochiaceae) medicinal species. Int. J. Mol. Sci. 2019, 20, 1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiapella, J.O.; Barfuss, M.H.J.; Xue, Z.Q.; Greimler, J. The plastid genome of Deschampsia cespitosa (Poaceae). Molecules 2019, 24, 216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Zhang, C.; Guo, X.; Liu, Q.; Wang, K. Complete chloroplast genome of Camellia japonica genome structures, comparative and phylogenetic analysis. PLoS ONE 2019, 14, e0216645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, C.; Deng, Y.; Wang, J. The complete chloroplast genomes of Echinacanthus species (Acanthaceae): Phylogenetic relationships, adaptive evolution, and screening of molecular markers. Front. Plant Sci. 2019, 9, 1989. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Sylvester, S.P.; Li, M.; Zhang, C.; Li, X.; Duan, Y.; Wang, X. The complete plastid genome of Magnolia zenii and genetic comparison to Magnoliaceae species. Molecules 2019, 24, 261. [Google Scholar] [CrossRef] [Green Version]
- Li, D.M.; Xu, Y.C.; Zhu, G.F. Complete Chloroplast genome of the plant Stahlianthus involucratus (Zingiberaceae). Mitochondrial DNA Part B 2019, 4, 2702–2703. [Google Scholar] [CrossRef] [Green Version]
- Li, D.M.; Zhao, C.Y.; Zhu, G.F.; Xu, Y.C. Complete chloroplast genome sequence of Hedychium coronarium. Mitochondrial DNA Part B 2019, 4, 2806–2807. [Google Scholar] [CrossRef] [Green Version]
- Li, D.M.; Zhao, C.Y.; Xu, Y.C. Characterization and phylogenetic analysis of the complete chloroplast genome of Curcuma longa (Zingiberaceae). Mitochondrial DNA Part B 2019, 4, 2974–2975. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Deng, J.; Li, Y.; Gao, G.; Ding, C.; Zhang, L.; Zhou, Y.; Yang, R. The complete chloroplast genome sequence of Curcuma flaviflora (Curcuma). Mitochondrial DNA Part A 2016, 27, 3644–3645. [Google Scholar] [CrossRef]
- Saina, J.K.; Li, Z.Z.; Gichira, A.W.; Liao, Y.Y. The complete chloroplast genome sequence of tree of heaven (Ailanthus altissima (Mill.)) (Sapindales: Simaroubaceae), an important pantropical tree. Int. J. Mol. Sci. 2018, 19, 929. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Huang, Y.; An, W.; Zheng, X.; Huang, S.; Liang, L. Sequencing and structural analysis of the complete chloroplast genome of the medicinal plant Lycium chinense Mill. Plants 2019, 8, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.H.; Yang, J.; Park, J.; Yamada, T.; Maki, M.; Kim, S.C. Comparison of whole plastome sequences between thermogenic skunk cabbage Symplocarpus renifolius and nonthermogenic S. nipponicus (Orontioideae; Araceae) in east Asia. Int. J. Mol. Sci. 2019, 20, 4678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, N.; Han, L.; Chen, C.; Wang, Z. The complete chloroplast genome sequence of Epipremnum aureum and its comparative analysis among eight Araceae species. PLoS ONE 2018, 13, e0192956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Yu, H.; Wang, J.; Lei, W.; Gao, J.; Qiu, X.; Wang, J. The complete chloroplast genome sequences of the medicinal plant Forsythia suspense (Oleaceae). Int. J. Mol. Sci. 2017, 18, 2288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- George, B.; Bhatt, B.S.; Awasthi, M.; George, B.; Singh, A.K. Comparative analysis of microsatellites in chloroplast genomes of lower and higher plants. Curr. Genet. 2015, 61, 665–677. [Google Scholar] [CrossRef] [PubMed]
- Tong, W.; Kim, T.S.; Park, Y.J. Rice chloroplast genome variation architecture and phylogenetic dissection in diverse Oryza species assessed by whole-genome resequencing. Rice 2016, 9, 57. [Google Scholar] [CrossRef] [Green Version]
- Dong, W.L.; Wang, R.N.; Zhang, N.Y.; Fan, W.B.; Fang, M.F.; Li, Z.H. Molecular evolution of chloroplast genomes of orchid species: Insights into phylogenetic relationship and adaptive evolution. Int. J. Mol. Sci. 2018, 19, 716. [Google Scholar] [CrossRef] [Green Version]
- Gu, C.; Ma, L.; Wu, Z.; Chen, K.; Wang, Y. Comparative analyses of chloroplast genomes from 22 Lythraceae species: Inferences for phylogenetic relationships and genome evolution within Myrtales. BMC Plant Biol. 2019, 19, 281. [Google Scholar] [CrossRef]
- Yin, K.; Zhang, Y.; Li, Y.; Du, F.K. Different natural selection pressures on the atpF gene in evergreen sclerophyllous and deciduous oak species: Evidence from comparative analysis of the complete chloroplast genome of Quercus aquifolioides with other oak species. Int. J. Mol. Sci. 2018, 19, 1042. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; Bi, Y.; Yang, F.; Zhang, M.; Chen, X.; Xue, J.; Zhang, X. Complete chloroplast genome sequences of Lilium: Insights into evolutionary dynamics and phylogenetic analyses. Sci. Rep. 2017, 7, 5751. [Google Scholar] [CrossRef] [Green Version]
- Lu, Q.; Ye, W.; Lu, R.; Xu, W.; Qiu, Y. Phylogenomic and comparative analyses of complete plastomes of Croomia and Stemona (Stemonaceae). Int. J. Mol. Sci. 2018, 19, 2383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Chen, S.; Zhang, X. Whole-genome comparison reveals heterogeneous divergence and mutation hotspots in chloroplast genome of Eucommia ulmoides oliver. Int. J. Mol. Sci. 2018, 19, 1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, D.; Zhao, Z.; Zhang, T.; Zhong, W.; Liu, C.; Yuan, Q.; Huang, L. The chloroplast genome sequence of Scutellaria baicalensis provides insight into intraspecific and interspecific chloroplast genome diversity in Scutellaria. Genes 2017, 8, 227. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Hu, Z.; Lin, X.; Li, Q.; Gao, H.; Luo, G.; Chen, S. High-throughput pyrosequencing of the complete chloroplast genome of Magnolia officinalis and its application in species identification. Acta Pharm. Sin. 2012, 47, 124–130. [Google Scholar]
- Luo, R.; Liu, B.; Xie, Y.; Li, Z.; Huang, W.; Yuan, J.; He, G.; Chen, Y.; Pan, Q.; Liu, Y. SOAPdenovo2: An empirically improved memory-efficient short-end de novo assembler. Gigascience 2012, 1, 18. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stoneshavas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Chaisson, M.J.; Tesler, G. Mapping single molecule sequencing reads using basic local alignment with successive refinement (BLASR): Application and theory. BMC Bioinform. 2012, 13, 238. [Google Scholar] [CrossRef] [Green Version]
- Denisov, G.; Walenz, B.; Halpern, A.L.; Miller, J.; Axerlrod, N.; Levy, S.; Sutton, G. Consensus generation and variant detection by celera assembler. Bioinformatics 2008, 24, 1035–1040. [Google Scholar] [CrossRef] [Green Version]
- Wyman, S.K.; Jansen, R.K.; Boore, J.L. Automatic annotation of organellar genomes with DOGMA. Bioinformatics 2004, 20, 3252–3255. [Google Scholar] [CrossRef] [Green Version]
- Lowe, T.M.; Chan, P.P. tRNAscan-SE On-line: Search and Contextual Analysis of Transfer RNA Genes. Nucleic Acids Res. 2016, 44, W54–W57. [Google Scholar] [CrossRef]
- Greiner, S.; Lehwark, P.; Bock, R. OrganellarGenomeDRAW (OGDRAW) version 1.3.1: Expanded toolkit for the graphical visualization of organellar genomes. Nucleic Acids Res. 2019, 47, W59–W64. [Google Scholar] [CrossRef] [Green Version]
- MISA-Microsatellite Identification Tool. Available online: http://pgrc.ipk-gatersleben.de/misa/ (accessed on 20 September 2017).
- Kurtz, S.; Choudhuri, J.V.; Ohlebusch, E.; Schleiermacher, C.; Stoye, J.; Giegerich, R. REPuter: The manifold applications of repeat analysis on a genomic scale. Nucleic Acids Res. 2001, 29, 4633–4642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. Mega7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mower, J.P. The PREP Suite: Predictive RNA editors for plant mitochondrial genes, chloroplast genes and user-defined alignments. Nucleic Acids Res. 2009, 37, W253–W259. [Google Scholar] [CrossRef] [PubMed]
- Frazer, K.A.; Pachter, L.; Poliakov, A.; Rubin, E.M.; Dubchak, I. VISTA: Computational tools for comparative genomics. Nucleic Acids Res. 2004, 32, W273–W279. [Google Scholar] [CrossRef] [PubMed]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcais, G.; Delcher, A.L.; Phillippy, A.M.; Coston, R.; Salzberg, S.L.; Zimin, A. MUMmer4: A fast and versatile genome alignment system. PLoS Comput. Biol. 2018, 14, e1005944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A. Se-Al: Sequence Alignment Editor; Version 2.0. Available online: http://tree.bio.ed.ac.uk/software (accessed on 30 September 2017).










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genome Characteristics | A. katsumadai | A. oxyphylla Guangdong | A. pumila | A. zerumbet | A. oxyphylla Hainan |
|---|---|---|---|---|---|
| GenBank number | MK262728 | MK262729 | MK262731 | JX088668 | NC_035895 |
| Genome size (bp) | 162,387 | 161,410 | 161,920 | 159,773 | 161,351 |
| LSC length (bp) | 87,667 | 87,279 | 87,261 | 87,644 | 87,248 |
| SSC length (bp) | 15,306 | 16,180 | 15,317 | 18,295 | 16,175 |
| IR length (bp) | 29,707 | 28,964/28,987 | 29,671 | 26,917 | 28,964 |
| Total genes | 133 | 133 | 133 | 132 | 132 |
| Protein-coding genes | 87 | 87 | 87 | 86 | 87 |
| tRNA genes | 38 | 38 | 38 | 38 | 37 |
| rRNA genes | 8 | 8 | 8 | 8 | 8 |
| GC content (%) | 36.15 | 36.16 | 36.17 | 36.27 | 36.17 |
| Category | Gene Names | Amount |
|---|---|---|
| Photosystem Ⅰ | psaA, psaB, psaC, psaI, psaJ | 5 |
| Photosystem Ⅱ | psbA, psbB, psbC, psbD, psbE, psbF, psbH, psbI, psbJ, psbK, psbL, psbM, psbN, psbT, psbZ | 15 |
| Cytochrome b/f complex | petA, petB *, petD *, petG, petL, petN | 6 |
| ATP synthase | atpA, atpB, atpE, atpF *, atpH, atpI | 6 |
| NADH dehydrogenase | ndhA *, ndhB(×2) *, ndhC, ndhD, ndhE, ndhF, ndhG, ndhH, ndhI, ndhJ, ndhK | 12 |
| Rubisco | rbcL | 1 |
| RNA polymerase | rpoA, rpoB, rpoC1 *, rpoC2 | 4 |
| Large subunit ribosomal proteins | rpl2(×2) *, rpl14, rpl16 *, rpl20, rpl22, rpl23(×2), rpl32, rpl33, rpl36 | 11 |
| Small subunit ribosomal proteins | rps2, rps3, rps4, rps7(×2), rps8, rps11, rps12(×2) *, rps14, rps15, rps16 *, rps18, rps19(×2) | 15 |
| Other proteins | accD, ccsA, cemA, clpP **, infA, matK | 6 |
| Proteins of unknown function | ycf1(×2), ycf2(×2), ycf3 **, ycf4 | 6 |
| Ribosomal RNAs | rrn4.5(×2), rrn5(×2), rrn16(×2), rrn23(×2) | 8 |
| Transfer RNAs | trnA-UGC(×2) *, trnC-GCA, trnD-GUC, trnE-UUC, trnF-GAA, trnfM-CAU, trnG-GCC *, trnG-UCC, trnH-GUG(×2), trnI-CAU(×2), trnI-GAU(×2) *, trnK-UUU(×2) *, trnL-CAA(×2), trnL-UAA *, trnL-UAG, trnM-CAU, trnN-GUU(×2), trnP-UGG, trnQ-UUG, trnR-ACG(×2), trnR-UCU, trnS-GCU(×2), trnS-UGA, trnT-UGU, trnV-GAC(×2), trnV-UAC *, trnW-CCA, trnY-GUA | 38 |
| Genes | A. katsumadai | A. oxyphylla Guangdong | A. zerumbet | A. oxyphylla Hainan | Location | ||||
|---|---|---|---|---|---|---|---|---|---|
| S | N | S | N | S | N | S | N | ||
| psbA | 3 | 0 | 3 | 0 | 3 | 0 | 3 | 0 | LSC |
| matK | 2 | 1 | 3 | 3 | 2 | 2 | 3 | 3 | LSC |
| atpA | 1 | 1 | 2 | 1 | 1 | 1 | 2 | 1 | LSC |
| atpF | - | - | - | - | 2 | 0 | - | - | LSC |
| atpH | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | LSC |
| atpI | 1 | 1 | - | - | 2 | 1 | - | - | LSC |
| rps2 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | LSC |
| rpoC2 | 5 | 7 | 8 | 8 | 6 | 7 | 8 | 8 | LSC |
| rpoC1 | 0 | 2 | 6 | 3 | 0 | 2 | 6 | 3 | LSC |
| rpoB | 7 | 2 | 7 | 3 | 6 | 3 | 7 | 3 | LSC |
| petN | - | - | - | - | 1 | 0 | - | - | LSC |
| psbD | - | - | - | - | 1 | 2 | - | - | LSC |
| psbC | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | LSC |
| psbZ | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | LSC |
| rps14 | - | - | 0 | 1 | 1 | 0 | 0 | 1 | LSC |
| psaB | 2 | 0 | 2 | 1 | 1 | 0 | 2 | 1 | LSC |
| psaA | 6 | 1 | 4 | 1 | 5 | 1 | 4 | 1 | LSC |
| ycf3 | - | - | 1 | 0 | - | - | 1 | 0 | LSC |
| rps4 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | LSC |
| ndhK | 0 | 2 | 1 | 1 | 1 | 3 | 1 | 1 | LSC |
| ndhC | 1 | 1 | 0 | 2 | 0 | 1 | 0 | 2 | LSC |
| atpB | - | - | 2 | 2 | 1 | 1 | 3 | 2 | LSC |
| rbcL | 3 | 4 | 5 | 1 | 3 | 3 | 5 | 1 | LSC |
| accD | 2 | 1 | 3 | 2 | 2 | 1 | 3 | 2 | LSC |
| ycf4 | - | - | 1 | 0 | - | - | 1 | 0 | LSC |
| cemA | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | LSC |
| petA | 0 | 2 | 1 | 2 | 0 | 2 | 1 | 2 | LSC |
| rpl33 | - | - | 0 | 1 | - | - | 0 | 1 | LSC |
| rps18 | - | - | 1 | 0 | - | - | 1 | 0 | LSC |
| rpl20 | - | - | 2 | 0 | 1 | 0 | 2 | 0 | LSC |
| clpP | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | LSC |
| psbB | 1 | 0 | - | - | 3 | 0 | - | - | LSC |
| psbH | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | LSC |
| petB | 1 | 0 | - | - | 1 | 0 | - | - | LSC |
| petD | - | - | 0 | 1 | - | - | 0 | 1 | LSC |
| rpoA | 1 | 0 | 0 | 1 | - | - | 0 | 1 | LSC |
| rps11 | - | - | 0 | 1 | - | - | 0 | 1 | LSC |
| rpl36 | 1 | 0 | 2 | 0 | 1 | 0 | 2 | 0 | LSC |
| infA | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | LSC |
| rps8 | 2 | 0 | 2 | 0 | 2 | 0 | 2 | 0 | LSC |
| rpl14 | 1 | 0 | 1 | 0 | 1 | 1 | 1 | 0 | LSC |
| rpl16 | - | - | - | - | 0 | 1 | - | - | LSC |
| rps3 | 3 | 2 | 2 | 3 | 2 | 3 | 2 | 3 | LSC |
| rpl22 | 0 | 3 | 0 | 1 | 0 | 3 | 0 | 1 | LSC |
| ndhF | 5 | 7 | 8 | 5 | 6 | 6 | 8 | 5 | SSC |
| ccsA | 0 | 3 | 0 | 2 | 1 | 2 | 0 | 2 | SSC |
| ndhD | - | - | 3 | 0 | - | - | 3 | 0 | SSC |
| ndhE | - | - | 1 | 0 | - | - | 1 | 0 | SSC |
| ndhG | 2 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | SSC |
| ndhI | - | - | 1 | 0 | - | - | 1 | 0 | SSC |
| ndhA | 2 | 2 | 2 | 1 | 2 | 2 | 2 | 1 | SSC |
| ndhH | 2 | 2 | 1 | 0 | 2 | 2 | 1 | 0 | SSC |
| rps15 | 1 | 1 | 0 | 3 | 0 | 1 | 0 | 3 | SSC |
| ycf1 | 2 | 15 | 6 | 19 | 3 | 18 | 6 | 19 | IRa/b |
| Total | 67 | 61 | 90 | 72 | 73 | 69 | 91 | 72 | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, D.-M.; Zhu, G.-F.; Xu, Y.-C.; Ye, Y.-J.; Liu, J.-M. Complete Chloroplast Genomes of Three Medicinal Alpinia Species: Genome Organization, Comparative Analyses and Phylogenetic Relationships in Family Zingiberaceae. Plants 2020, 9, 286. https://doi.org/10.3390/plants9020286
Li D-M, Zhu G-F, Xu Y-C, Ye Y-J, Liu J-M. Complete Chloroplast Genomes of Three Medicinal Alpinia Species: Genome Organization, Comparative Analyses and Phylogenetic Relationships in Family Zingiberaceae. Plants. 2020; 9(2):286. https://doi.org/10.3390/plants9020286
Chicago/Turabian StyleLi, Dong-Mei, Gen-Fa Zhu, Ye-Chun Xu, Yuan-Jun Ye, and Jin-Mei Liu. 2020. "Complete Chloroplast Genomes of Three Medicinal Alpinia Species: Genome Organization, Comparative Analyses and Phylogenetic Relationships in Family Zingiberaceae" Plants 9, no. 2: 286. https://doi.org/10.3390/plants9020286