Interaction between Macrophages and Nanoparticles: In Vitro 3D Cultures for the Realistic Assessment of Inflammatory Activation and Modulation of Innate Memory

 ,
,  , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Synthesis and Characterization of AuNP
2.1.1. AuNP Synthesis and Purification
2.1.2. Nanoparticle Characterization
2.2. LAL Assay
2.3. Human Monocyte Isolation and Culture
2.4. Cell Activation and Induction of Innate Memory
2.5. Cytokine Analysis
2.6. Statistical Analysis
3. Results
3.1. Gold Nanoparticle Characterisation
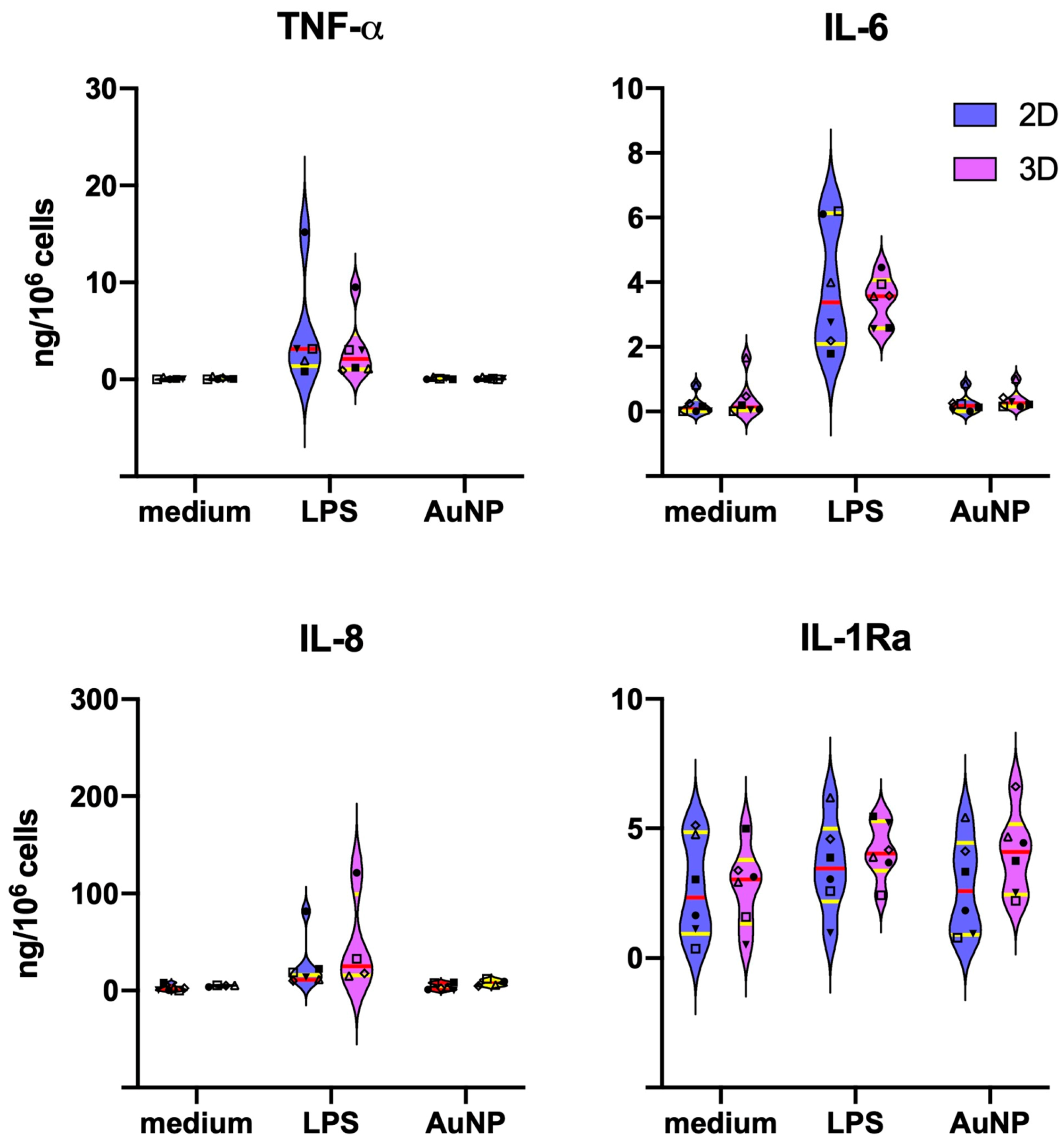
3.2. In Vitro Development of Macrophage Innate Memory in 2D vs. 3D Cultures
3.3. Differences in the Primary Monocyte Reactivity in 2D vs. 3D Cultures
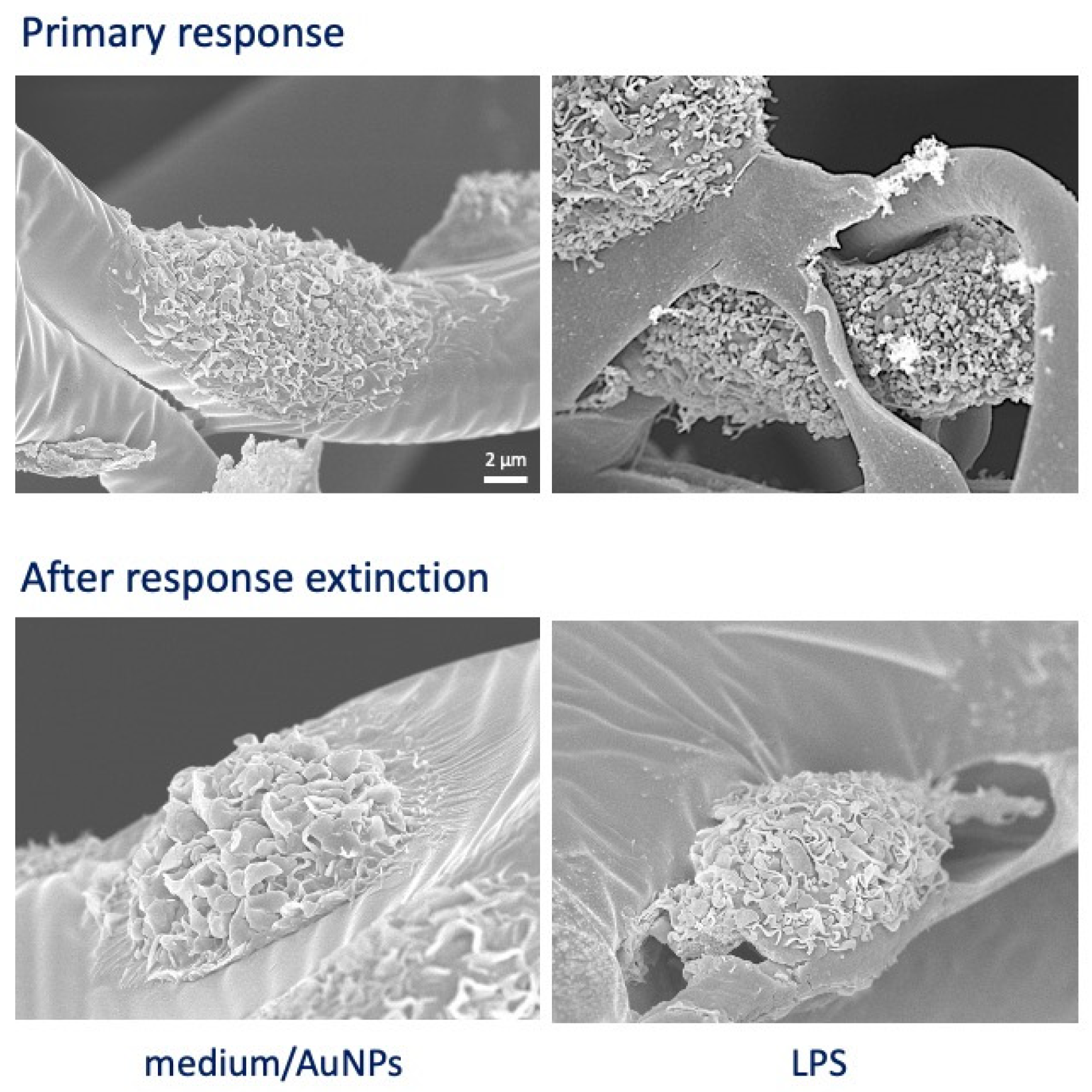
3.4. Extinction of Inflammatory Activation in Macrophages Six Days after Priming
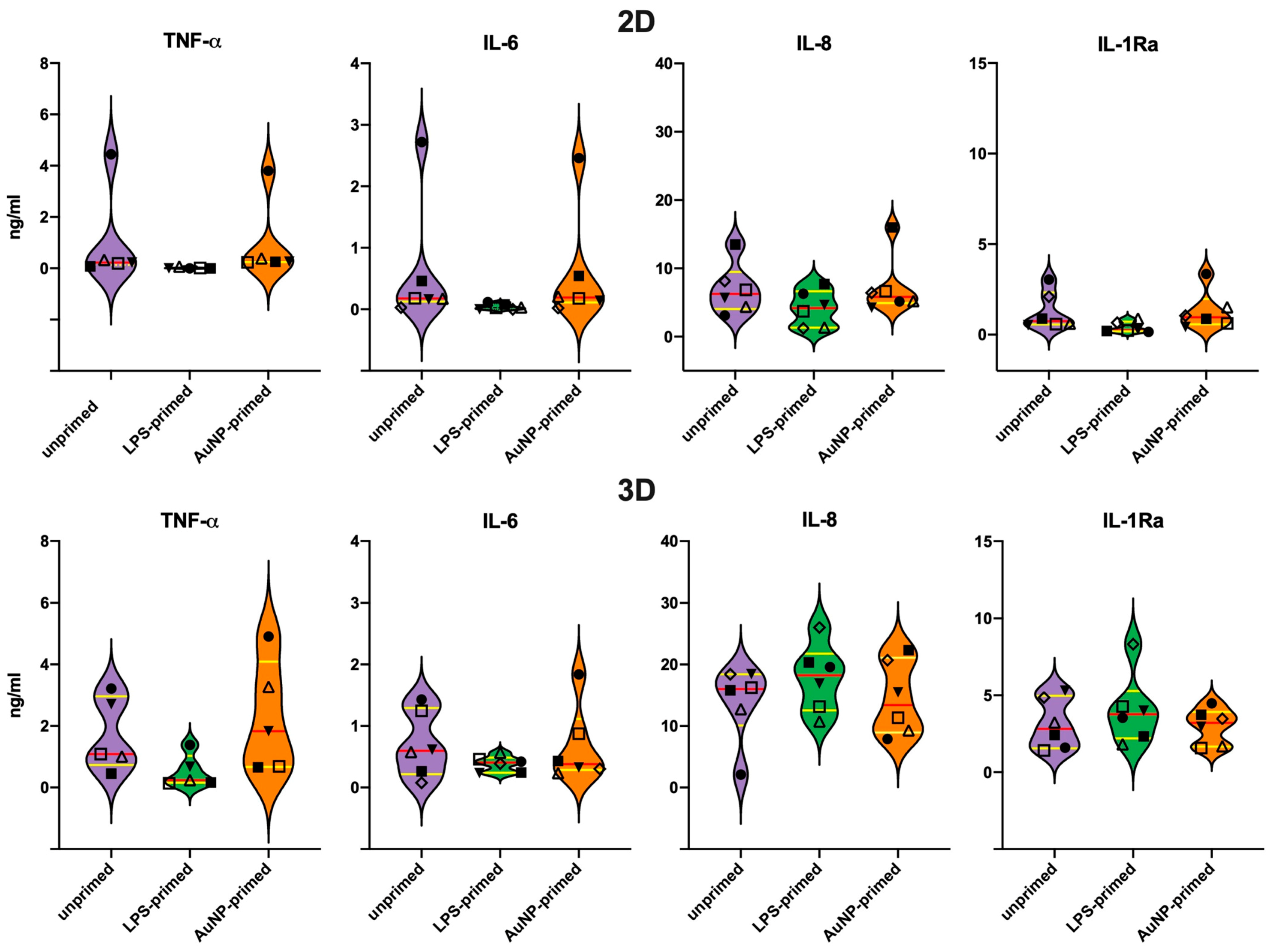
3.5. Secondary Response of Unprimed Macrophages
3.6. Memory Response of LPS-Primed Macrophages
3.7. Memory Response of AuNP-Primed Macrophages
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dobrovolskaia, M.A.; Shurin, M.; Shvedova, A.A. Current understanding of interactions between nanoparticles and the immune system. Toxicol. Appl. Pharmacol. 2016, 299, 78–89. [Google Scholar] [CrossRef] [Green Version]
- Fadeel, B. Hide and seek: Nanomaterial interactions with the immune system. Front. Immunol. 2019, 10, 133. [Google Scholar] [CrossRef]
- Boraschi, D.; Italiani, P.; Palomba, R.; Decuzzi, P.; Duschl, A.; Fadeel, B.; Moghimi, S.M. Nanoparticles and innate immunity: New perspectives on host defence. Sem. Immunol. 2017, 34, 33–51. [Google Scholar] [CrossRef]
- Medzhitov, R.; Janeway, C.A. Innate immunity: The virtues of a non-clonal system of recognition. Cell 1997, 91, 295–298. [Google Scholar] [CrossRef] [Green Version]
- Carta, S.; Yassi, S.; Pettinati, I.; Delfino, L.; Dinarello, C.A.; Rubartelli, A. The rate of IL-1β secretion in different myeloid cells varies with the extent of redox response to TLR triggering. J. Biol. Chem. 2011, 286, 27069–27080. [Google Scholar] [CrossRef] [Green Version]
- Italiani, P.; Boraschi, D. From monocytes to M1/M2 macrophages: Phenotypical vs. functional differentiation. Front. Immunol. 2014, 5, 514. [Google Scholar] [CrossRef] [Green Version]
- Mills, C.D.; Lenz, L.L.; Ley, K. M1/M2 Macrophages: The Arginine Fork in the Road to Health and Disease; Frontiers Media: Lausanne, Switzerland, 2015; p. 280. [Google Scholar] [CrossRef]
- Holst, O.; Ulmer, A.J.; Brade, H.; Flad, H.D.; Rietschel, E.T. 1996. Biochemistry and cell biology of bacterial endotoxins. FEMS Immunol. Med. Microbiol. 1996, 16, 83–104. [Google Scholar] [CrossRef] [PubMed]
- Mestas, J.; Hughues, C.C.W. Of mice and not men: Differences between mouse and human immunology. J. Immunol. 2004, 172, 2731–2738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, M.M. A prescription for human immunology. Immunity 2008, 29, 835–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Italiani, P.; Mazza, E.M.C.; Lucchesi, D.; Cifola, I.; Gemelli, C.; Grande, A.; Battaglia, C.; Bicciato, S.; Boraschi, D. Transcriptomic profiling of the development of the inflammatory response in human monocytes in vitro. PLoS ONE 2014, 9, e87680. [Google Scholar] [CrossRef]
- Li, Y.; Italiani, P.; Casals, E.; Valkenborg, D.; Mertens, I.; Baggerman, G.; Nelissen, I.; Puntes, V.; Boraschi, D. Assessing the immunosafety of engineered nanoparticles with a novel in vitro model based on human primary monocytes. ACS Appl. Mater. Interfaces 2016, 8, 28437–28447. [Google Scholar] [CrossRef] [PubMed]
- Ferretti, A.M.; Usseglio, S.; Mondini, S.; Drago, C.; La Mattina, R.; Chini, B.; Verderio, C.; Leonzino, M.; Cagnoli, C.; Joshi, P.; et al. Towards bio-compatible magnetic nanoparticles: Immune-related effects, in vitro internalization, and in vivo bio-distribution of zwitterionic ferrite nanoparticles with unexpected renal clearance. J. Colloid Interface Sci. 2020, 582, 678–700. [Google Scholar] [CrossRef] [PubMed]
- Beeson, P.B. Development of tolerance to typhoid bacterial pyrogen and its abolition by reticulo-endothelial blockade. Proc. Soc. Exp. Biol. Med. 1946, 61, 248–250. [Google Scholar] [CrossRef] [PubMed]
- Howard, J.G.; Biozzi, G.; Halpern, B.N.; Stiffel, C.; Mouton, D. The effect of Mycobacterium tuberculosis (BCG) infection on the resistance of mice to bacterial endotoxin and Salmonella enteritidis infection. Br. J. Exp. Pathol. 1959, 40, 281–290. [Google Scholar] [PubMed]
- Bistoni, F.; Vecchiarelli, A.; Cenci, E.; Puccetti, P.; Marconi, P.; Cassone, A. Evidence for macrophage-mediated protection against lethal Candida albicans infection. Infect. Immun. 1986, 51, 668–674. [Google Scholar] [CrossRef] [Green Version]
- Netea, M.G.; Quintin, J.; van der Meer, J.W.M. Trained immunity: A memory for innate host defense. Cell Host Microbe 2011, 9, 355–361. [Google Scholar] [CrossRef] [Green Version]
- Milutinovi’c, B.; Kurtz, J. Immune memory in invertebrates. Sem. Immunol. 2016, 28, 328–342. [Google Scholar] [CrossRef]
- Cooper, D.; Eleftherianos, I. Memory and specificity in the insect immune system: Current perspectives and future challenges. Front. Immunol. 2017, 8, 539. [Google Scholar] [CrossRef]
- Gourbal, B.; Pinaud, S.; Beckers, G.J.M.; Van Der Meer, J.W.M.; Conrath, U.; Netea, M.G. Innate immune memory: An evolutionary perspective. Immunol. Rev. 2018, 283, 21–40. [Google Scholar] [CrossRef]
- Arts, R.J.W.; Joosten, L.A.B.; Netea, M.G. The potential role of trained immunity in autoimmune and autoinflammatory disorders. Front. Immunol. 2018, 9, 298. [Google Scholar] [CrossRef]
- Salam, A.P.; Borsini, A.; Zunszain, P.A. Trained innate immunity: A salient factor in the pathogenesis of neuroimmune psychiatric disorders. Mol. Psychiatry 2018, 23, 170–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salani, F.; Sterbini, V.; Sacchinelli, E.; Garramone, M.; Bossu, P. Is innate memory a double-edge sword in Alzheimer’s Disease? A reappraisal of new concepts and old data. Front. Immunol. 2019, 10, 1768. [Google Scholar] [CrossRef] [PubMed]
- Töpfer, E.; Boraschi, D.; Italiani, P. Innate immune memory: The latest frontier of adjuvanticity. J. Immunol. Res. 2015, 2015, 478408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, Z.; Afkhami, S.; Bavananthasivam, J.; Fritz, D.K.; D’Agostino, M.R.; Vaseghi-Shanjani, M.; Yao, Y.; Jeyanathan, M. Innate immune memory of tissue-resident macrophages and trained innate immunity: Re-vamping vaccine concept and strategies. J. Leukocyte Biol. 2020, 108, 825–834. [Google Scholar] [CrossRef]
- Sánchez-Ramón, S.; Conejero, L.; Netea, M.G.; Sancho, D.; Palomares, Ó.; Subiza, J.L. Trained immunity-based vaccines: A new paradigm for the development of broad-spectrum anti-infectious formulations. Front. Immunol. 2018, 9, 2936. [Google Scholar] [CrossRef] [PubMed]
- Mulder, W.J.M.; Ochando, J.; Joosten, L.A.B.; Fayad, Z.A.; Netea, M.G. Therapeutic targeting of trained immunity. Nat. Rev. Drug Discov. 2019, 18, 553–566. [Google Scholar] [CrossRef]
- Italiani, P.; Boraschi, D. Induction of innate immune memory by engineered nanoparticles: A hypothesis that may become true. Front. Immunol. 2017, 8, 734. [Google Scholar] [CrossRef] [Green Version]
- Lebre, F.; Boland, J.B.; Gouveia, P.; Gorman, A.; Lundahl, M.; O’Brien, F.J.; Coleman, J.; Lavelle, E.C. Pristine graphene induces innate immune training. Nanoscale 2020, 12, 11192–11200. [Google Scholar] [CrossRef]
- Swartzwelter, B.J.; Barbero, F.; Verde, A.; Mangini, M.; Pirozzi, M.; De Luca, A.C.; Puntes, V.F.; Leite, L.C.C.; Italiani, P.; Boraschi, D. Gold nanoparticles modulate BCG-induced innate iummune memory in nhuman monocytes by shifting the memory response towards tolerance. Cells 2020, 9, 284. [Google Scholar] [CrossRef] [Green Version]
- Auguste, M.; Balbi, T.; Ciacci, C.; Canonico, B.; Papa, S.; Borello, A.; Vezzulli, L.; Canesi, L. Shift in immune parameters after repeated exposure to nanoplastics in the Marine Bivalve Mytilus. Front. Immunol. 2020, 11, 426. [Google Scholar] [CrossRef]
- Italiani, P.; Della Camera, G.; Boraschi, D. Induction of innate immune memory by engineered nanoparticles in monocytes/macrophages: From hypothesis to reality. Front. Immunol. 2020, 11, 566309. [Google Scholar] [CrossRef]
- Franceschi, C.; Salvioli, S.; Garagnani, P.; de Eguileor, M.; Monti, D.; Capri, M. Immunobiography and the heterogeneity of immune responses in the elderly: A focus on inflammaging and trained immunity. Front. Immunol. 2017, 8, 982. [Google Scholar] [CrossRef]
- Ojea-Jiménez, I.; Bastús, N.G.; Puntes, V. Influence of the sequence of the reagents addition in the citrate-mediated synthesis of gold nanoparticles. J. Phys. Chem. C 2011, 115, 15752–15757. [Google Scholar] [CrossRef]
- Li, Y.; Italiani, P.; Casals, E.; Tran, N.; Puntes, V.F.; Boraschi, D. Optimising the use of commercial LAL assays for the analysis of endotoxin contamination in metal colloids and metal oxide nanoparticles. Nanotoxicology 2015, 9, 462–473. [Google Scholar] [CrossRef] [Green Version]
- Oostingh, G.J.; Casals, E.; Italiani, P.; Colognato, R.; Stritzinger, R.; Ponti, J.; Pfaller, T.; Kohl, Y.; Ooms, D.; Favilli, F.; et al. Problems and challenges in the development and validation of human cell-based assays to determine nanoparticle-induced immunomodulatory effects. Particle Fibre Toxicol. 2011, 8, 8. [Google Scholar] [CrossRef] [Green Version]
- Piella, J.; Bastús, N.G.; Puntes, V. Size-dependent protein-nanoparticle interaction in citrate-stabilized gold nanoparticles: The emergence of the protein corona. Bioconj. Chem. 2017, 28, 88–97. [Google Scholar] [CrossRef]
- Comenge, J.; Sotelo, C.; Romero, F.; Gallego, O.; Barnadas, A.; Garcia-Caballero Parada, T.; Dominguez, F.; Puntes, V.F. Detoxifying antitumoral drugs via nanoconjugation: The case of gold nanoparticles and cisplatin. PLoS ONE 2012, 7, e47562. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Shi, Z.; Radauer-Preiml, I.; Andosch, A.; Casals, E.; Luetz-Meidl, U.; Cobaleda, M.; Lin, Z.; Jaberi-Douraki, M.; Italiani, P.; et al. Bacterial endotoxin (LPS) binds to the surface of gold nanoparticles, interferes with biocorona formation and induces human monocyte inflammatory activation. Nanotoxicology 2017, 11, 1157–1175. [Google Scholar] [CrossRef] [Green Version]
- Montanaro, M.; Meloni, M.; Anemona, L.; Giurato, L.; Scimeca, M.; Izzo, V.; Servadei, F.; Smirnov, A.; Candi, E.; Mauriello, A.; et al. Macrophage activation and M2 polarization in wound bed of diabetic patients treated by dermal/epidermal substitute Nevelia. Int. J. Lower Extr. Wounds 2020. online ahead of print. [Google Scholar] [CrossRef]
- Röszer, T. Understanding the misterious M2 macrophage through activation markers and effector mechanisms. Med. Inflamm. 2015, 2015, 816460. [Google Scholar] [CrossRef] [Green Version]
- Sherwood, J.; Bertrand, J.; Nalesso, G.; Poulet, B.; Pitsillides, A.; Brandolini, L.; Karystinou, A.; De Bari, C.; Luyten, F.P.; Pitzalis, C.; et al. A homeostatic function of CXCR2 signalling in articular cartilage. Ann. Rheum. Dis. 2015, 74, 2207–2215. [Google Scholar] [CrossRef] [Green Version]
- Nicolás-Ávila, J.A.; Adrover, J.M.; Hidalgo, A. Neutrophils in homeostasis, immunity and cancer. Immunity 2017, 46, 15–28. [Google Scholar] [CrossRef] [Green Version]
- Pekalski, M.L.; Rubio Garcia, A.; Ferreira, R.C.; Rainbow, D.B.; Smyth, D.J.; Mashar, M.; Brady, J.; Savinykh, N.; Castro Dopico, X.; Mahmood, S.; et al. Neonatal and adult recent thymic emigrants produce IL-8 and express complement receptors CR1 and CR2. JCI Insight 2017, 2, e93739. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Swartzwelter, B.J.; Verde, A.; Rehak, L.; Madej, M.; Puntes, V.F.; De Luca, A.C.; Boraschi, D.; Italiani, P. Interaction between Macrophages and Nanoparticles: In Vitro 3D Cultures for the Realistic Assessment of Inflammatory Activation and Modulation of Innate Memory. Nanomaterials 2021, 11, 207. https://doi.org/10.3390/nano11010207
Swartzwelter BJ, Verde A, Rehak L, Madej M, Puntes VF, De Luca AC, Boraschi D, Italiani P. Interaction between Macrophages and Nanoparticles: In Vitro 3D Cultures for the Realistic Assessment of Inflammatory Activation and Modulation of Innate Memory. Nanomaterials. 2021; 11(1):207. https://doi.org/10.3390/nano11010207
Chicago/Turabian StyleSwartzwelter, Benjamin J., Alessandro Verde, Laura Rehak, Mariusz Madej, Victor. F. Puntes, Anna Chiara De Luca, Diana Boraschi, and Paola Italiani. 2021. "Interaction between Macrophages and Nanoparticles: In Vitro 3D Cultures for the Realistic Assessment of Inflammatory Activation and Modulation of Innate Memory" Nanomaterials 11, no. 1: 207. https://doi.org/10.3390/nano11010207