
Context-Dependent Roles of RNA Modifications in Stress Responses and Diseases



Department of Medicine, Section of Dermatology, University of Chicago, Chicago, IL 60637, USA
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2021, 22(4), 1949; https://doi.org/10.3390/ijms22041949
Submission received: 31 January 2021
/
Revised: 5 February 2021
/
Accepted: 8 February 2021
/
Published: 16 February 2021
(This article belongs to the Special Issue Cell Death in Biology and Diseases 2.0)
Abstract
:RNA modifications are diverse post-transcriptional modifications that regulate RNA metabolism and gene expression. RNA modifications, and the writers, erasers, and readers that catalyze these modifications, serve as important signaling machineries in cellular stress responses and disease pathogenesis. In response to stress, RNA modifications are mobilized to activate or inhibit the signaling pathways that combat stresses, including oxidative stress, hypoxia, therapeutic stress, metabolic stress, heat shock, DNA damage, and ER stress. The role of RNA modifications in response to these cellular stressors is context- and cell-type-dependent. Due to their pervasive roles in cell biology, RNA modifications have been implicated in the pathogenesis of different diseases, including cancer, neurologic and developmental disorders and diseases, and metabolic diseases. In this review, we aim to summarize the roles of RNA modifications in molecular and cellular stress responses and diseases.
1. Introduction
RNA modifications are covalent chemical modifications of RNA molecules. To date, over 100 chemical modifications of RNA species have been identified [1,2]. The regulation and function of RNA modifications have recently emerged as pivotal mechanisms that regulate a wide range of biological and pathological processes, giving rise to the field known as epitranscriptomics.
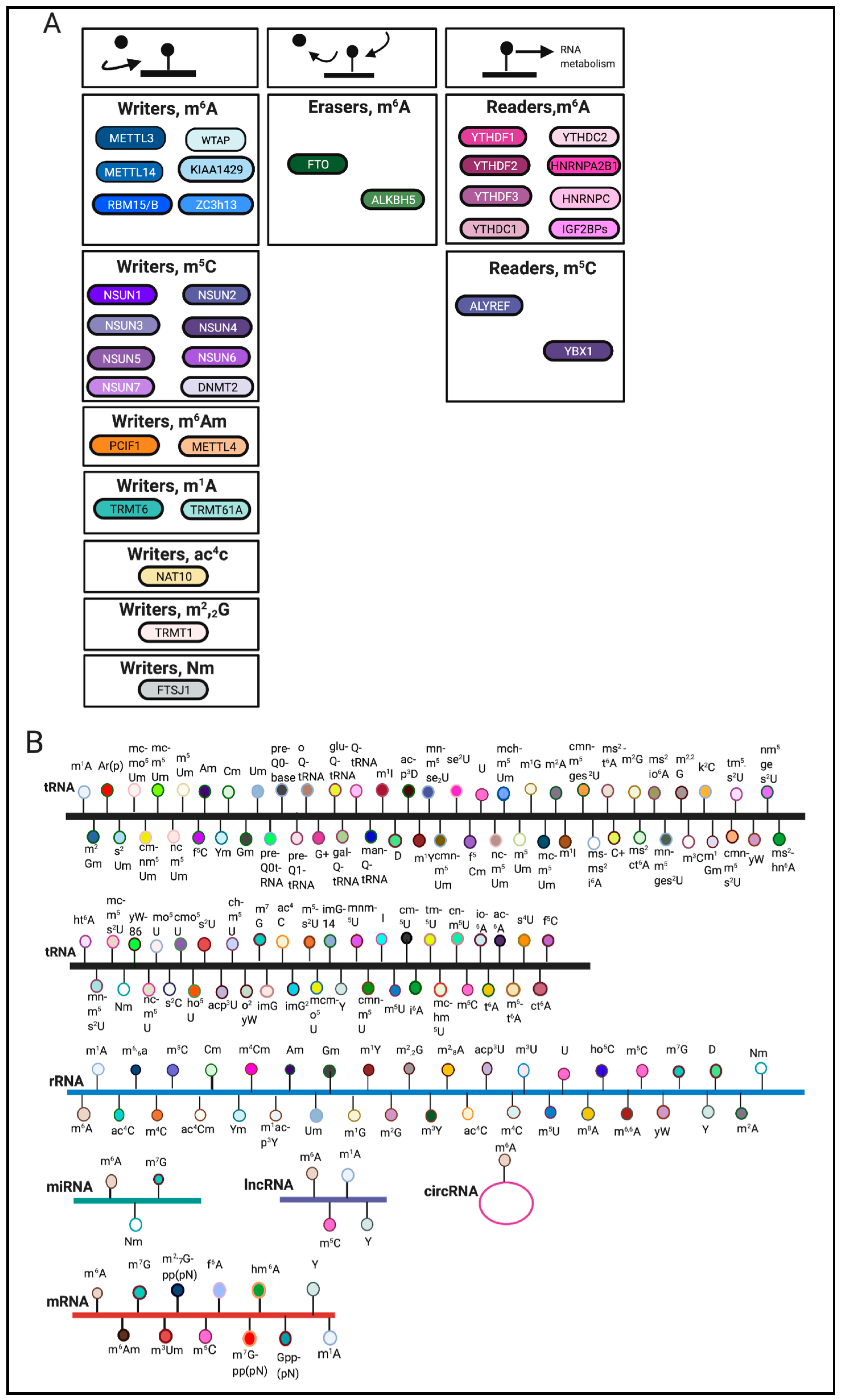
RNA modifications are regulated through the coordination of ‘writers’, ‘erasers’ and ‘readers’, which deposit, remove, and recognize RNA modifications, respectively (Figure 1A). These enzymes represent key elements in patterning the epitranscriptomic landscape.
RNA modifications can occur on various RNA species including mRNA, tRNA, rRNA and other non-coding RNAs [3] (Figure 1B). Among these RNA species, transfer RNAs (tRNAs) contain the most RNA modifications [4]. One of the most abundant modifications on tRNA and rRNA is 5-methylcytosine (m5C) [5]. In comparison, mRNA modifications were more difficult to identify and characterize due to their low-abundance. The advent of sophisticated sequencing technologies and methods has generated a renewed interest in mRNA modifications.
The most abundant internal mRNA modification is N6-methyladenosine (m6A) [2]. Other mRNA modifications are reviewed elsewhere [2,3,12]. m6A writers include methyltransferase-like family members 3,14 (METTL3, METTL14), Wilms tumor 1-associated protein (WTAP), KIAA1429, RBM15/RBM15B, and Zc3h13 [2,13,14]. m6A erasers include fat mass and obesity associated (FTO) and Alkb homologue 5 (ALKBH5) [2]. Furthermore, m6A readers include YTH m6A-binding protein 1,2 and 3 (YTHDF1, YTHDF2, YTHDF3), Heterogeneous nuclear ribonucleoprotein A2/B1 and C (HNRNPA2B1, HNRNPC), YTH domain-containing 1 and 2 (YTHDC1, YTHDC2), and IGF2BPs [2,9,15]. m6A writers, erasers and readers are reviewed elsewhere [2,16]. In this review, we aim to summarize the role of RNA modifications in cellular stress response pathways and within various diseases. m6A remains the best-studied RNA modification and will therefore be the main focus of this review.
2. RNA Modifications in Stress Responses
2.1. Oxidative Stress
Both m6A and m5C pathways play important roles in regulating the cellular response to oxidative stress, a condition caused by disrupted redox homeostasis, including the generation of reactive oxygen species (ROS).
2.1.1. Writers
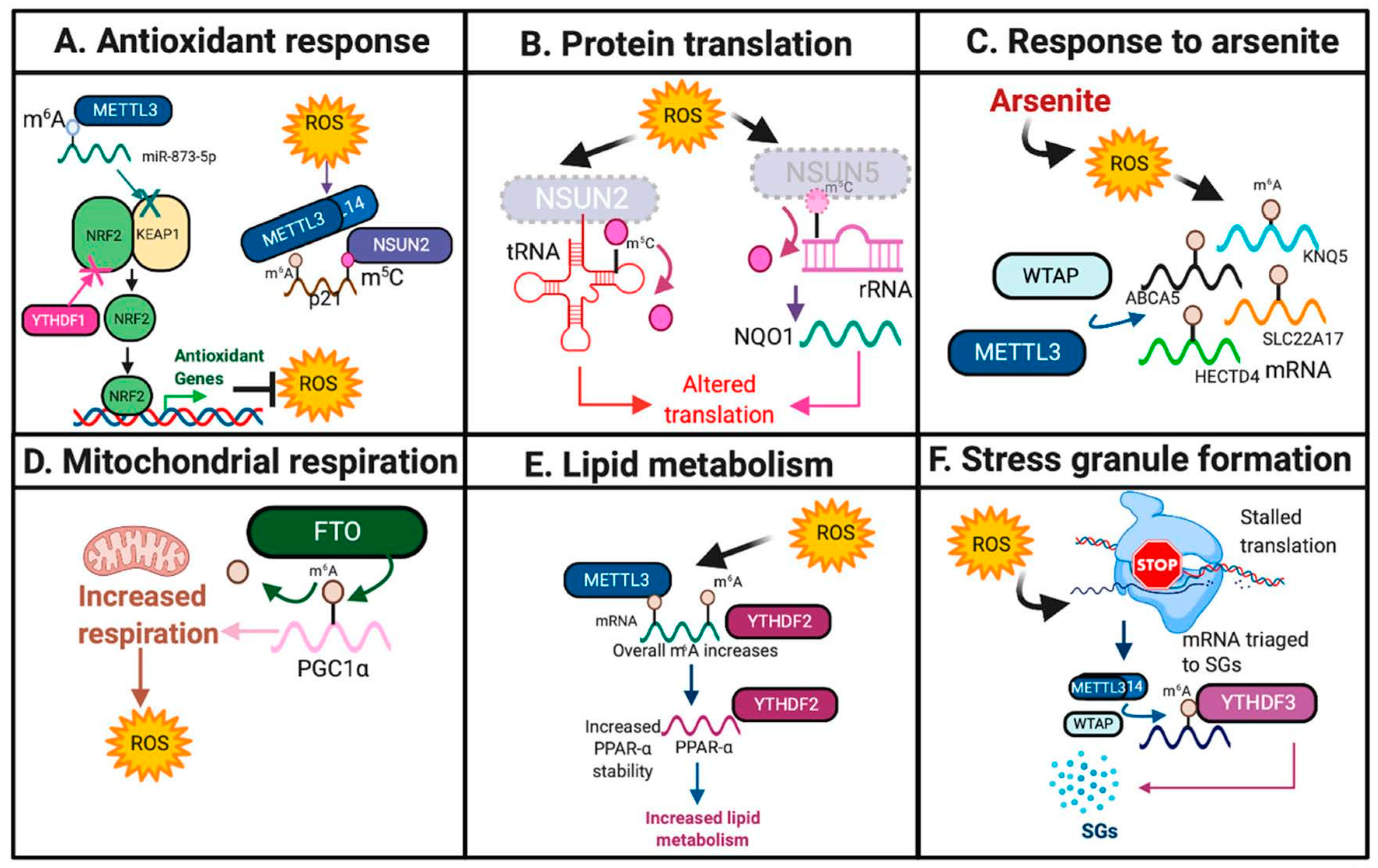
Previous studies have suggested that METTL3 may serve a protective role against oxidative stress. In mouse renal tubular epithelial cells, METTL3-mediated m6A deposition onto miR-873-5P and promoted its recognition and maturation by DGCR8, an miRNA processing complex, leading to inhibition of KEAP1 and activation of the NRF2 antioxidant pathway in response to colistin-induced oxidative stress [17,18] (Figure 2A and Figure 3A, Table 1).
Furthermore, m5C writers have also been implicated in the oxidative stress response. Using colon cancer cell lines and HeLa cells, Li and colleagues identified that NSUN2 catalyzes the deposition of m5C, and METTL3/METTL14 catalyze the deposition of m6A, in the 3′ UTR of p21, which has been previously found to up-regulate NRF2 in response to oxidative stress and induce cellular senescence [20,60]. These results therefore suggest that these methyltransferases synergize to increase p21 expression in response to oxidative stress [20]. Furthermore, nsun2 expression was decreased upon oxidative stress in mouse keratinocytes, leading to site-specific reductions in m5C on tRNAs, stalling protein translation, and forcing the cell to enter into a catabolic state [23]. These results suggest that loss of NSUN2 may be necessary to induce the cellular stress response [23] (Figure 2B and Figure 3A, Table 1).
In human keratinocytes, ROS was induced by arsenite, an environmental carcinogen, resulting in increased expression of WTAP and METTL14 and overall m6A levels [22,61]. Specific m6A increases were also identified on cell surface receptor SLC22A17, potassium channel KCNQ5, ATP binding cassette subfamily A member 5 (ABCA5) and HECT domain E3 Ubiquitin Protein Ligase 4 (HECTD4), leading to decreased mRNA expression of these genes [22] (Figure 2C and Figure 3A, Table 1). Further studies are needed to elucidate the functional significance of these targets in this context.
Additionally, in glioma, epigenetic loss of NSUN5 resulted in hypomethylation of 28S rRNA at position C3782, leading to decreased overall protein translation in response to oxidative stress, but increased translation of proteins that promote survival and adaptation to oxidative stress, including antioxidant NQO1 [24] (Figure 2B and Figure 3A, Table 1).
2.1.2. Erasers
m6A demethylases have also been implicated in the oxidative stress response. Zhuang and colleagues identified that in clear cell renal carcinoma, FTO induces oxidative stress through m6A demethylation at the 3′UTR of PGC1α, a major regulator of mitochondrial metabolism, which resulted in increased PGC1α mRNA stability and translation and increased ROS production [27] (Figure 2D and Figure 3A, Table 1). Furthermore, overexpression of FTO in hepatocytes and myotubes resulted in increased lipogenesis and mitochondrial dysfunction, which, in turn, increased ROS levels and induced oxidative stress [25,26].
2.1.3. Readers
m6A readers serve diverse roles in response to oxidative stress. Loss of clock protein BMAL1 increased ROS production in human HepG2 and mouse Hepa1-6 cells, which resulted in specific METTL3-mediated m6A increases on the nuclear receptor peroxisome proliferator-activator α (PPARα) locus, increased YTHDF2 expression, which mediates PPARα stability, and increased lipid metabolism [21] (Figure 2E and Figure 3A, Table 1). However, YTHDF1 and YTHDF2 may serve context-dependent functions in mediating oxidative stress. YTHDF1 may serve as a negative regulator of the KEAP1-NRF2 antioxidant pathway as YTHDF1 knockdown in human bronchial epithelium cells (BEAS-2B) increased NRF2 expression and antioxidant production [28].
Oxidative stress also induced METTL3/METTL14/WTAP-mediated m6A deposition on 5′UTR of stress granules (SGs), which are assemblies of mRNA that are stalled within translation initiation, and form in response to stress [19,62,63]. YTHDF3 has been shown to mediate the triage of mRNAs into SGs in response to oxidative stress in HEK293 and U2OS osteosarcoma cells [19] (Figure 2F and Figure 3A, Table 1).
2.2. Hypoxia
While the genetic mechanisms that mediate the hypoxia response have been a subject of study for many years, post-transcriptional regulation of the hypoxia response is not as well-elucidated. Emerging evidence suggests that m6A RNA methylation has an active role in cellular response to hypoxia (Figure 3B, Table 1).
2.2.1. Writers
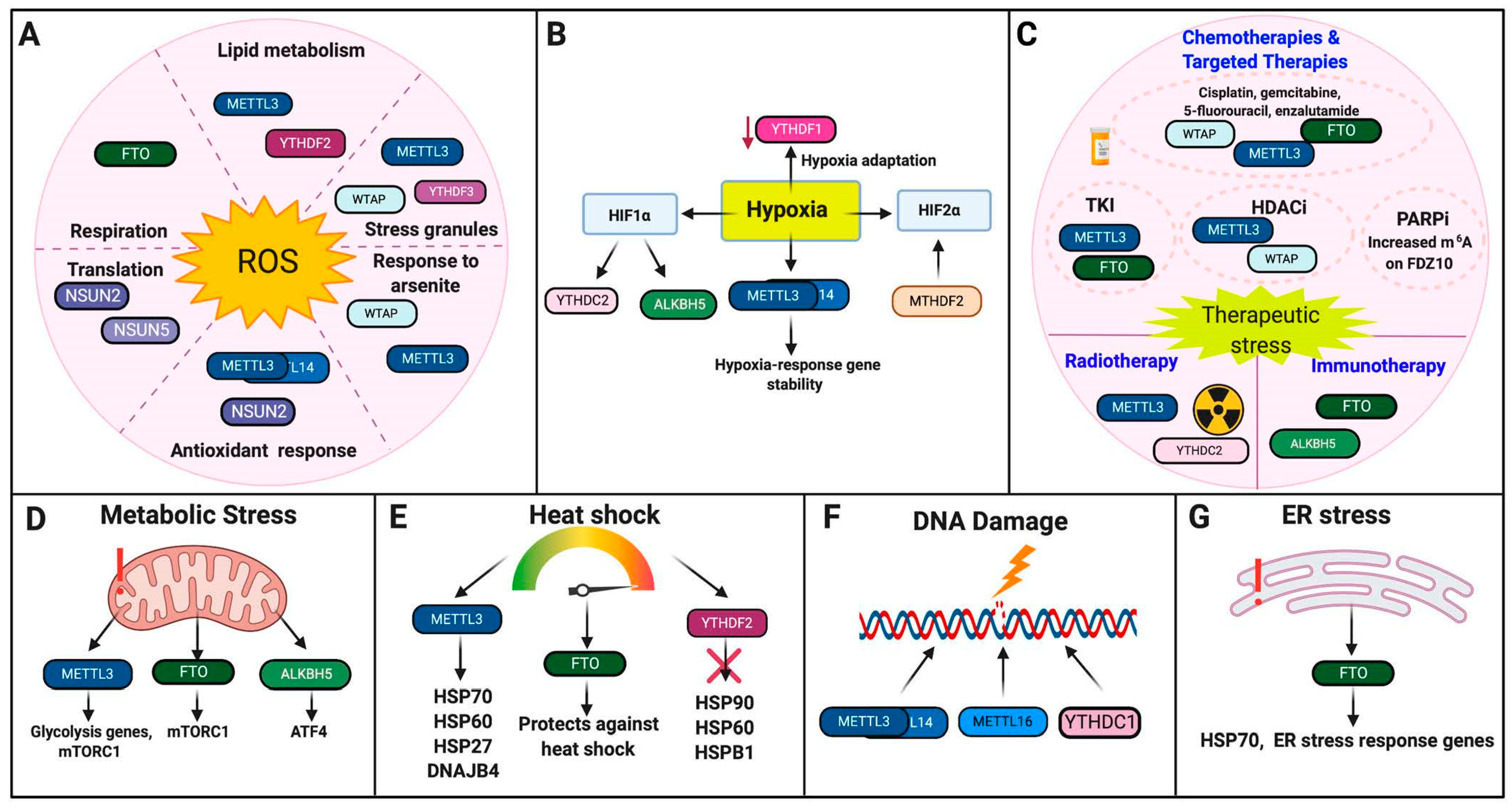
METTL3/METTL14 activity and m6A deposition can be regulated by hypoxia. Overall m6A levels were increased under hypoxia in HEK293T cells, including specific m6A increases on hypoxia-responsive genes GLUT1 and c-MYC [29]. Additionally, METTL3/14 promoted the mRNA stability of hypoxia-response genes, suggesting that m6A is involved in post-transcriptional stabilization of hypoxia-induced mRNAs [29]. Furthermore, METTL3 was identified as a hypoxia-responsive gene in both endothelial cells and adipocyte stem cells upon differentiation into vascular smooth muscle [30,31].
However, other groups have found that m6A levels increased in response to hypoxia irrespective of METTL3/METTL14 or ALKBH5 activity in transformed human mammary epithelial cells, suggesting that there may be other novel hypoxia-induced mechanisms that regulate m6A levels [64]. Correspondingly, methylenetetrahydrofolate dehydrogenase 2 (MTHDF2), an enzyme that is important in one-carbon metabolism, is believed to regulate m6A deposition on HIF-2α in renal cell carcinoma (RCC), which leads to increased HIF-2α translation, metabolic reprogramming, and tumor progression [65,66].
2.2.2. Erasers
Thalhammer and colleagues identified that the ALKBH5 promoter contains two putative binding sites for HIF-1α and was up-regulated in response to hypoxia across several different cancer cell lines, including U2OS, breast cancer cell line MCF7, and neuroblastoma cell line IMR32 [32]. However, the hypoxia-induced changes in m6A demethylation and patterning across these cell lines requires further study. Additionally, ALKBH5 and ZNF217 were found to regulate a breast cancer stem cell (BCSC) phenotype in hypoxia, resulting in decreased m6A methylation of NANOG and KLF4 and increased expression of these pluripotency factors [33,34].
2.2.3. Readers
The role of m6A readers in response to hypoxia requires further study. Developmentally, lower YTHDF1 expression was seen in mammals that live at low oxygen altitudes, suggesting that loss of YTHDF1 may serve as an evolutionary adaptation to hypoxia [28]. Furthermore, YTHDC2 was found to promote HIF-1α mRNA translation in colon cancer [35].
2.3. Therapeutic Stress
Resistance to therapies remains a pressing issue for many types of cancer. Writers, erasers, and readers for m6A RNA methylation are shown to play important roles in response to therapeutic agents, including targeted therapies, immunotherapies, and conventional therapies (Figure 3C, Table 1). Further elucidating the epigenetic mechanisms that mediate chemotherapy resistance remains an active area of research.
Chemotherapies and Targeted Therapies
Cisplatin, gemcitabine, 5-fluorouracil, enzalutamide. METTL3 expression may induce resistance to chemotherapy as METTL3 knockdown increased sensitivity to gemcitabine, 5-fluorouracil (5-FU) and cisplatin in pancreatic cancer, potentially through activation of MAPK signaling [38]. In glioma, METTL3 may also promote temozolomide resistance through m6A-mediated stabilization of SOX2, which mediates glioma stem cell formation, as METTL3 knockdown increased temozolomide sensitivity in this context [39]. Additionally, in AML, WTAP knockdown increased sensitivity to etoposide, suggesting that WTAP may mediate etoposide resistance [40]. Furthermore, FTO increased resistance to cisplatin in cervical squamous cell carcinoma (CSCC) through m6A demethylation of the β-CATENIN transcript, leading to increased β-CATENIN mRNA and protein expression [45]. FTO also contributed to enzalutamide resistance in castration-resistant prostate cancer by mediating alterations within androgen receptor-regulated enhancer RNAs (AR-eRNAs) [44]. Therefore, whether m6A methylation or demethylation mediates chemotherapy resistance is cancer-cell type and context dependent.
Kinase Inhibitors. Yan and colleagues reported increased FTO-mediated m6A demethylation increased resistance to tyrosine kinase inhibitors (TKIs) through enhanced mRNA stability of anti-apoptotic genes and pro-proliferation genes, such as BCL-2, in leukemia [42]. In liver cancer, decreased METTL3 expression increased resistance to sorafenib through decreased mRNA stability of FOXO3, a negative regulator for autophagy, leading to the activation of autophagy-mediated therapeutic resistance [37].
Histone Deacetylase (HDAC) Inhibitors. In non-small cell lung cancer (NSCLC), chidamide, an HDAC inhibitor, inhibited c-MET expression by decreasing the m6A methylation of c-MET, potentially through targeting WTAP and METTL3 [36]. Additionally, chidamide-mediated decreases in m6A and c-MET expression also increased sensitivity to critzotinib [36].
PARP Inhibitors (PARPi). Increased m6A deposition on FZD10 mRNA resulted in increased FZD10 stability, which mediated resistance to PARPi through activation of the WNT/β-CATENIN pathway in BRCA-mutant epithelial ovarian cancer cells [67].
Immunotherapies. In melanoma, FTO promoted the expression of PDCD1, which expresses the PD-1 protein, as well as CXCR4 and SOX10, promoting melanoma tumorigenesis and resistance to immunotherapy [43]. Decreased FTO expression therefore increased response to the PD-1 blockade immunotherapy as well as IFN-induced tumor cell killing [43]. ALKBH5 knockdown also increased PD-1 therapy efficiency in both melanoma and colorectal cancer models by decreasing populations of immunosuppressive Tregs and myeloid-derived suppressor cells (MDSCs) [41]. The role of m6A in mediating the immune response is complex and remains an active area of study.
Ionizing Radiation. METTL3 expression promoted radioresistance in glioma by promoting glioblastoma stem cell maintenance through m6A-mediated stabilization of SOX2, and subsequent METTL3 knockdown resulted in increased radiosensitivity [39]. In addition to promoting cisplatin resistance, FTO also contributed to radiation resistance in CSCC through up-regulation of [45]. YTHDC2 contributed to radioresistance in nasopharyngeal carcinoma (NPC) cells by binding to IGF1R mRNA and promoting the translation of the IGF1R transcript, leading to downstream activation of the IGF1R-AKT/S6 signaling axis [46].
2.4. Metabolic Stress
2.4.1. Writers
METTL3 may serve pivotal roles in glycolysis and metabolism and subsequent loss of METTL3 may therefore promote metabolic stress. In hepatocellular carcinoma (HCC), METTL3 expression positively correlated with glycolysis genes SLC2A1, HK2, PFKM and PKM [47]. Knockdown of METTL3 decreased mTORC1 activity, a major regulator of cellular metabolism, and also sensitized HCC cells to 2-deoxyglucose (2-DG), a glycolysis inhibitor, promoting glycolytic stress and the subsequent induction of cell death [47,68].
2.4.2. Erasers
ATF4 is a master regulator of amino acid metabolism [69]. Upon amino acid starvation and subsequent induction of nutrient stress in mouse embryonic fibroblasts (MEFs), alkbh5 was recruited to atf4 mRNA and promoted atf4 translation, suggesting that m6A regulation at the atf4 transcript contributes to the cellular stress response in response to nutrient stress [48].
Additionally, FTO was found to activate the mTORC1 pathway in MEFs as fto knockdown decreased mTORC1 activation and increased autophagy [49]. Furthermore, fto was down-regulated upon amino acid starvation, potentially as a means to regulate mTORC1 activation upon amino acid stress [49]. However, in melanoma, FTO was up-regulated in response to metabolic stress and promoted adaptation to metabolic stress, therefore reflecting the context-dependent roles of m6A in response to metabolic stress [43].
2.5. Heat Shock
The best-studied heat shock response involves the translation and regulation of heat shock proteins (HSPs), which function to mediate adaptation to heat stress.
2.5.1. Writers
METTL3 was decreased in HepG2 cells upon heat shock [52]. METTL3 may also target HSP70, HSP60, and HSP27 downstream, as METTL3 knockdown decreased the mRNA expression of these genes [52]. m6A-mediated regulation of HSPs is further corroborated by the identification of m6A sites on HSPs [50]. Other studies have also identified increased METTL3-mediated m6A deposition on the 5′UTR of HSP70 post-heat shock, which is hypothesized to regulate cap-independent translation in response to heat stress [53]. Similarly, m6A increases were also identified in the 5′UTR of DNAJB4 (HSP40 Homolog) upon heat shock, but it is unclear whether METTL3 alone mediates this m6A increase [51]. These studies therefore highlight the context-dependent regulation of HSPs mediated by m6A.
2.5.2. Erasers
The role of m6A erasers in response to heat shock is not well-established. An overall decrease of m6A and increase in fto was noted in response to a mild heat challenge in the hypothalamus of male Cobb chicks, suggesting that FTO serves a protective role against heat stress [54]. However, during a harsher heat challenge, m6A levels increased, suggesting that m6A regulation may be temperature-dependent [54].
2.5.3. Readers
YTHDF2 mRNA was increased upon heat shock, but also may serve as a negative regulator of HSP90, HSP60, and HSPB1 as YTHDF2 knockdown resulted in increased mRNA expression of these HSPs in HepG2 cells [52]. Zhou and colleagues also discovered that m6A is preferentially deposited on the 5′UTR of newly transcribed HSP70 mRNA and is read by YTHDF2 upon heat shock, which may be functionally significant as previous reports have found that m6A deposition at the 5′UTR mediates cap-independent translation [55,70].
2.6. DNA Damage
Writers
In response to UVC/UVA radiation, m6A and DNA Pol were rapidly recruited (2–4 min post-UV) to sites of DNA damage by METTL3/METTL14 [56]. METTL16, a U6 snRNA methyltransferase, was also recruited to DNA damage sites at a later time point (20–30 min post-UV) and was found to methylate small nuclear and nucleolar RNAs that were also recruited to DNA damage sites upon UV radiation [58]. m6A was only recruited to DNA damage lesions only in the presence of cyclobutane pyrimidine dimers (CPDs), which form in response to UV exposure [58]. Furthermore, in response to UV, m6A RNA modifications may utilize the nucleotide excision repair (NER) pathway rather than non-homologous end-joining (NHEJ), as knockout of two NHEJ-specific enzymes, SUV391H/H2, had no effect on m6A recruitment [58]. In response to double-stranded breaks (DSBs), METTL3 localized to DNA damage sites, depositing m6A on DNA-damage associated RNA, increasing accumulation of DNA-RNA hybrids, and recruiting YTHDC1 and DNA-damage associated proteins RAD51 and BRCA to initiate homologous recombination (HR) [57].
2.7. ER Stress
The role of RNA modifications in ER stress remains understudied in mammalian systems. Work in non-mammalian systems suggests that ER stress may influence or induce m6A deposition in response to viral infection and innate immune signaling [71]. The limited work in mammalian systems suggests that FTO stabilizes HSP70 and other ER stress-associated genes in response to genotoxic stress in osteoblasts [59] (Figure 3G, Table 1).
3. RNA Modifications in Diseases
3.1. Cancer
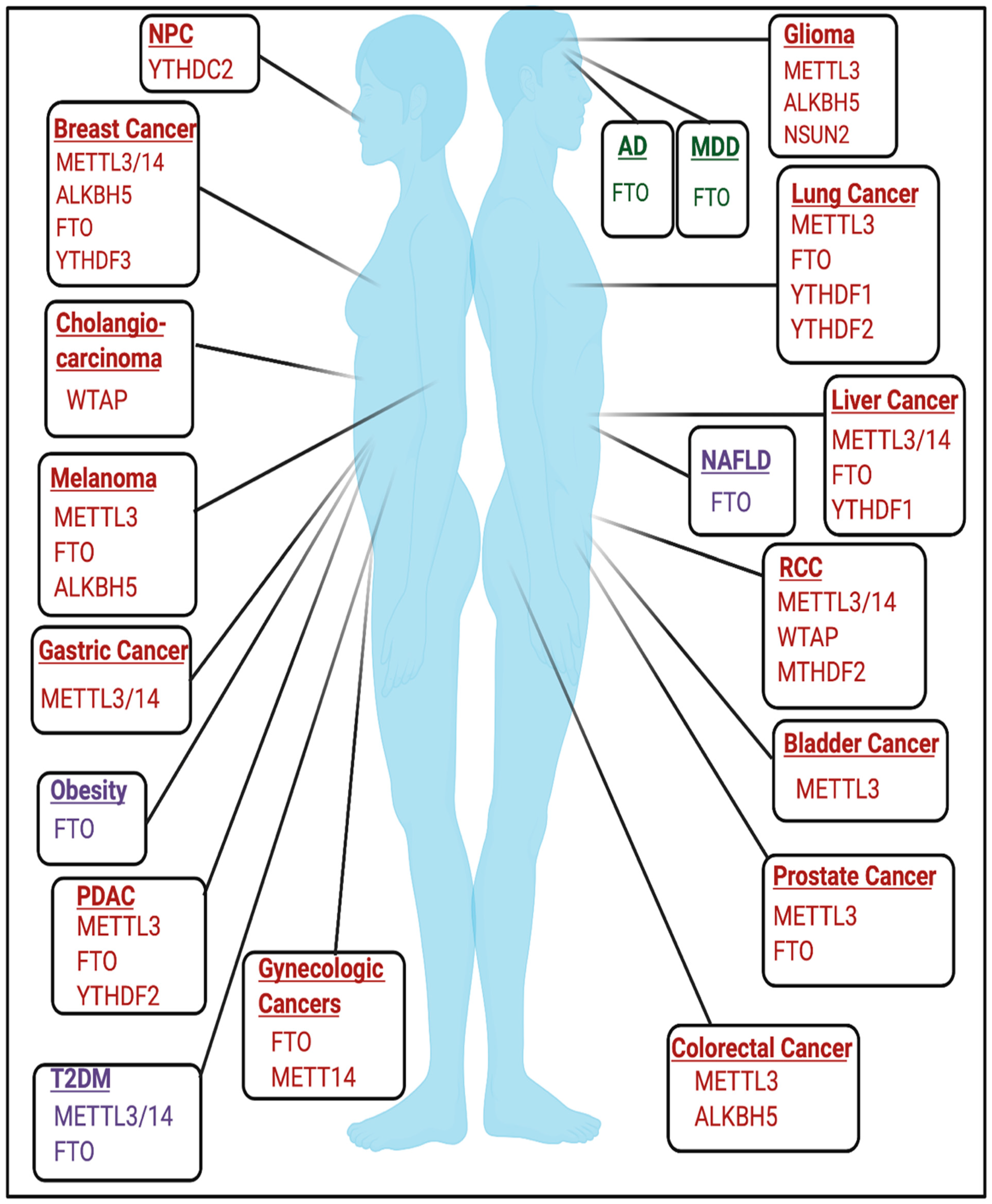
The role of RNA modifications in cancer is cell-type and context-dependent and has been reviewed extensively in various types of cancer [16,72,73,74,75,76,77,78,79,80,81] and are highlighted in Figure 4. Rare RNA modifications have been described in cancer type-specific contexts, such as glioblastoma, and are detailed in [82]. An active area of research seeks to elucidate the role of RNA modifications in non-coding RNAs and other RNA species in the context of cancer, which are reviewed elsewhere [83].
3.2. Developmental and Neurologic Disorders
The role of RNA modifications in the context of developmental and neurological disorders remains an active area of study. m6A has been previously found to play important roles in embryonic development and neurobiological functions [84,85]. The roles of m6A and other RNA modifications in mediating neurologic function are further discussed elsewhere [84,86,87,88,89].
The necessity of m6A in development is emphasized by early embryonic lethality in mettl3 KO mice [90]. Conditional mettl3 knockout in murine brains also resulted in severe developmental defects within the cerebrum and cortex and induced apoptosis in cerebella granule cells (CGCs) [91]. FTO may also be important in mediating development as expression of catalytically inactive mutant FTO(R316Q) resulted in severe growth defects [92]. Mutations in tRNA methyltransferases have been implicated in developmental disorders and are detailed in [93]. NSUN2 mutations have been linked to microcephaly, intellectual disability, and Dubowitz Syndrome, which is characterized by growth and mental retardation [94,95,96]. Additionally, homozygous frameshift mutations in TRMT1, a writer for m2,2G, have been linked to intellectual disability [97]. Mutations and polymorphisms in FTSJ1, a writer for 2′O-methylribose, have also been linked to X-linked mental retardation [98,99,100,101,102]. Furthermore, targets of fragile X mental retardation protein (FMRP), a protein that is commonly mutated in Fragile X Syndrome, were enriched for m6A, and FMRP targets were targeted for degradation by YTHDF2 [103].
3.2.1. Alzheimer’s Disease
m6A increases and distinct m6A patterning were found in the cortex and hippocampus of APP/PS1 transgenic mice, which are used to model Alzheimer’s Disease (AD) [104]. Additionally, AD-associated SNPs that decreased FTO expression were identified in Caucasian and Caribbean Hispanic populations [105]. AD patients also showed changes in small RNA modifications, which are detailed in [89].
3.2.2. Major Depressive Disorder
m6A and m6Am may be also implicated in major depressive disorder (MDD) as m6A and m6Am patterning were dysregulated in patients with MDD [106]. Conversely, the FTO variant rs9939609 was associated with a lower risk of developing MDD [107,108]. FTO may be also be involved in the development of anxiety, as fto−/− mice show increased anxiety-like behavior and hyperactivation of the hypothalamic-pituitary-adrenal (HPA) axis [109].
3.3. Metabolic Disorders and Diseases
FTO is a major driver in contributing to the pathogenesis of several metabolic diseases and therefore serves as a therapeutic target in this context.
3.3.1. Obesity
One of the strongest predictors of obesity is believed to be SNP rs9939609 in FTO [110,111,112,113,114,115]. FTO SNPs rs17817449 and rs3751812 also increased obesity risk in north Indian and Pakistani populations, respectively [116,117]. fto overexpression in mice also resulted in a dose-dependent increase in body mass and increased food intake [118,119]. However, these studies are m6A-independent. Conversely, previous studies have identified that FTO-mediated m6A demethylation regulates mRNA splicing in adipocytes and genes involved in sterol metabolism, which therefore may provide a mechanism by which FTO promotes obesity at the molecular level [120]. The role of FTO in metabolism is further reviewed elsewhere [121]. Identifying the m6A-dependent and m6A-independent functions of FTO in mediating obesity remains an active area of research.
3.3.2. Diabetes
In addition to obesity, m6A sequencing of type-two diabetes mellitus (T2DM) patients revealed overall changes in m6A patterning and hypomethylation of mRNA transcripts involved in insulin biogenesis, secretion, and pancreatic -cell biology [122,123]. FTO mRNA expression was also higher in some T2DM patients [124]. Additionally, METTL3/METTL14 expression was decreased in -cells of patients with T2DM, and METTL14 specifically may be essential for insulin secretion and -cell survival [122,123,125]. However, the role of m6A in mediating T2DM may be tissue and context-dependent as METTL3 and m6A levels were increased in liver tissue from T2DM patients [126].
3.3.3. Non-Alcoholic Fatty Liver Disease
Increased expression of fto was induced by a high-fat diet, resulting in increased lipogenesis and induction of non-alcoholic fatty liver disease (NAFLD), a disease that is commonly associated with obesity [26,127,128]. Identifying the m6A-dependent and m6A-independent functions of FTO in this context requires future study.
4. Conclusions and Perspectives
Emerging evidence demonstrates the crucial role of RNA modifications in stress responses and diseases. These findings not only shine new lights on molecular and cellular responses to a wide range of stress conditions, but also may provide new opportunities in targeting the RNA modification pathways to modulate stress responses and thus may prevent and treat diseases. The field of epitranscriptomics will benefit from the following advances in the near future. First, elucidating the context and cell-type-specific role of RNA modifications will allow us to fully understand the precise and diverse roles and functions of these modifications in biology and diseases. Throughout this review, we have highlighted the role of RNA modifications in promoting or resisting certain cellular processes that are cell-type and context-dependent. For example, what mediates the changes in RNA modifications and their role on gene transcription as a cell transitions from homeostasis to stress, or within oncogenic transformation? Second, while many sequencing technologies have been developed to detect RNA modifications, there is a need to detect these modifications directly at the single base resolution across all regions within the gene body. Last, there is also a need to further characterize the specificity with which RNA modifications are deposited. Accordingly, identifying the molecular mechanisms that determine which RNAs, RNA species, RNA secondary and tertiary structures, and adenosines, or other nucleosides, are targeted for modification will be critical to determining the sequence specificity of these modifications. These important advances will further facilitate the understanding of the precise role for RNA modifications in physiology and pathology, and may reveal new opportunities for disease diagnosis, prognosis, prevention, or treatment.
Author Contributions
Conceptualization, E.W. and Y.-Y.H.; writing—original draft preparation, E.W., Y.-H.C. and Y.-Y.H.; writing—review and editing, E.W., Y.-H.C. and Y.-Y.H.; visualization, E.W., Y.-H.C., Y.-Y.H.; funding acquisition, Y.-Y.H. All authors have read and agreed to the published version of the manuscript.
Funding
Work in the authors’ laboratory was supported in part by NIH grants 5T32CA009594-32 (E.W.), ES031534 (Y.-Y.H), ES024373 (Y.-Y.H.), ES030576 (Y.-Y.H.), the CACHET (NIH ES027792), the University of Chicago Comprehensive Cancer Center (NIH CA014599), the CTSA (NIH UL1 TR000430), and the University of Chicago Friends of Dermatology Endowment Fund.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We apologize to those investigators whose work could not be directly referenced owing to space limitations. We thank Ann Motten for her critical reading of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Boccaletto, P.; Machnicka, M.A.; Purta, E.; Piatkowski, P.; Baginski, B.; Wirecki, T.K.; de Crécy-Lagard, V.; Ross, R.; Limbach, P.A.; Kotter, A.; et al. MODOMICS: A database of RNA modification pathways. 2017 update. Nucleic Acids Res. 2018, 46, D303–D307. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.S.; Roundtree, I.A.; He, C. Post-transcriptional gene regulation by mRNA modifications. Nat. Rev. Mol. Cell Biol. 2017, 18, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Motorin, Y.; Helm, M. RNA nucleotide methylation. Wiley Interdiscip. Rev. RNA 2011, 2, 611–631. [Google Scholar] [CrossRef]
- Helm, M.; Motorin, Y. Detecting RNA modifications in the epitranscriptome: Predict and validate. Nat. Rev. Genet. 2017, 18, 275–291. [Google Scholar] [CrossRef] [PubMed]
- Bohnsack, K.E.; Höbartner, C.; Bohnsack, M.T. Eukaryotic 5-methylcytosine (m⁵C) RNA Methyltransferases: Mechanisms, Cellular Functions, and Links to Disease. Genes (Basel) 2019, 10, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alarcón, C.R.; Lee, H.; Goodarzi, H.; Halberg, N.; Tavazoie, S.F. N6-methyladenosine marks primary microRNAs for processing. Nature 2015, 519, 482–485. [Google Scholar] [CrossRef]
- Dai, Q.; Moshitch-Moshkovitz, S.; Han, D.; Kol, N.; Amariglio, N.; Rechavi, G.; Dominissini, D.; He, C. Nm-seq maps 2′-O-methylation sites in human mRNA with base precision. Nat. Methods 2017, 14, 695–698. [Google Scholar] [CrossRef] [PubMed]
- Dinescu, S.; Ignat, S.; Lazar, A.D.; Constantin, C.; Neagu, M.; Costache, M. Epitranscriptomic Signatures in lncRNAs and Their Possible Roles in Cancer. Genes (Basel) 2019, 10, 52. [Google Scholar] [CrossRef] [Green Version]
- Esteve-Puig, R.; Bueno-Costa, A.; Esteller, M. Writers, readers and erasers of RNA modifications in cancer. Cancer Lett. 2020, 474, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Pandolfini, L.; Barbieri, I.; Bannister, A.J.; Hendrick, A.; Andrews, B.; Webster, N.; Murat, P.; Mach, P.; Brandi, R.; Robson, S.C.; et al. METTL1 Promotes let-7 MicroRNA Processing via m7G Methylation. Mol. Cell 2019, 74, 1278–1290.e1279. [Google Scholar] [CrossRef] [Green Version]
- Romano, G.; Veneziano, D.; Nigita, G.; Nana-Sinkam, S.P. RNA Methylation in ncRNA: Classes, Detection, and Molecular Associations. Front. Genet. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Shi, H.; Chai, P.; Jia, R.; Fan, X. Novel insight into the regulatory roles of diverse RNA modifications: Re-defining the bridge between transcription and translation. Mol. Cancer 2020, 19, 78. [Google Scholar] [CrossRef]
- Patil, D.P.; Chen, C.K.; Pickering, B.F.; Chow, A.; Jackson, C.; Guttman, M.; Jaffrey, S.R. m(6)A RNA methylation promotes XIST-mediated transcriptional repression. Nature 2016, 537, 369–373. [Google Scholar] [CrossRef]
- Wen, J.; Lv, R.; Ma, H.; Shen, H.; He, C.; Wang, J.; Jiao, F.; Liu, H.; Yang, P.; Tan, L.; et al. Zc3h13 Regulates Nuclear RNA m(6)A Methylation and Mouse Embryonic Stem Cell Self-Renewal. Mol. Cell 2018, 69, 1028–1038.e1026. [Google Scholar] [CrossRef] [Green Version]
- Liao, S.; Sun, H.; Xu, C. YTH Domain: A Family of N6-methyladenosine (m6A) Readers. Genom. Proteom. Bioinform. 2018, 16, 99–107. [Google Scholar] [CrossRef]
- Uddin, M.B.; Wang, Z.; Yang, C. Dysregulations of Functional RNA Modifications in Cancer, Cancer Stemness and Cancer Therapeutics. Theranostics 2020, 10, 3164–3189. [Google Scholar] [CrossRef]
- Han, J.; Lee, Y.; Yeom, K.H.; Kim, Y.K.; Jin, H.; Kim, V.N. The Drosha-DGCR8 complex in primary microRNA processing. Genes Dev. 2004, 18, 3016–3027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Ishfaq, M.; Xu, L.; Xia, C.; Chen, C.; Li, J. METTL3/m(6)A/miRNA-873-5p Attenuated Oxidative Stress and Apoptosis in Colistin-Induced Kidney Injury by Modulating Keap1/Nrf2 Pathway. Front. Pharmacol. 2019, 10, 517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anders, M.; Chelysheva, I.; Goebel, I.; Trenkner, T.; Zhou, J.; Mao, Y.; Verzini, S.; Qian, S.-B.; Ignatova, Z. Dynamic m6A methylation facilitates mRNA triaging to stress granules. Life Sci. Alliance 2018, 1, e201800113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Li, X.; Tang, H.; Jiang, B.; Dou, Y.; Gorospe, M.; Wang, W. NSUN2-Mediated m5C Methylation and METTL3/METTL14-Mediated m6A Methylation Cooperatively Enhance p21 Translation. J. Cell. Biochem. 2017, 118, 2587–2598. [Google Scholar] [CrossRef]
- Zhong, X.; Yu, J.; Frazier, K.; Weng, X.; Li, Y.; Cham, C.M.; Dolan, K.; Zhu, X.; Hubert, N.; Tao, Y.; et al. Circadian Clock Regulation of Hepatic Lipid Metabolism by Modulation of m(6)A mRNA Methylation. Cell Rep. 2018, 25, 1816–1828.e1814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, T.; Li, X.; Sun, D.; Zhang, Z. Oxidative stress: One potential factor for arsenite-induced increase of N(6)-methyladenosine in human keratinocytes. Environ. Toxicol. Pharmacol. 2019, 69, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Gkatza, N.A.; Castro, C.; Harvey, R.F.; Heiß, M.; Popis, M.C.; Blanco, S.; Bornelöv, S.; Sajini, A.A.; Gleeson, J.G.; Griffin, J.L.; et al. Cytosine-5 RNA methylation links protein synthesis to cell metabolism. PLoS Biol. 2019, 17, e3000297. [Google Scholar] [CrossRef] [PubMed]
- Janin, M.; Ortiz-Barahona, V.; de Moura, M.C.; Martínez-Cardús, A.; Llinàs-Arias, P.; Soler, M.; Nachmani, D.; Pelletier, J.; Schumann, U.; Calleja-Cervantes, M.E.; et al. Epigenetic loss of RNA-methyltransferase NSUN5 in glioma targets ribosomes to drive a stress adaptive translational program. Acta Neuropathol. 2019, 138, 1053–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bravard, A.; Lefai, E.; Meugnier, E.; Pesenti, S.; Disse, E.; Vouillarmet, J.; Peretti, N.; Rabasa-Lhoret, R.; Laville, M.; Vidal, H.; et al. FTO is increased in muscle during type 2 diabetes, and its overexpression in myotubes alters insulin signaling, enhances lipogenesis and ROS production, and induces mitochondrial dysfunction. Diabetes 2011, 60, 258–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Ren, W.; Li, A.; Ding, Y.; Guo, W.; Su, D.; Hu, C.; Xu, K.; Chen, H.; Xu, X.; et al. Fat Mass and Obesity-Associated Gene Enhances Oxidative Stress and Lipogenesis in Nonalcoholic Fatty Liver Disease. Dig. Dis. Sci. 2013, 58, 1004–1009. [Google Scholar] [CrossRef]
- Zhuang, C.; Zhuang, C.; Luo, X.; Huang, X.; Yao, L.; Li, J.; Li, Y.; Xiong, T.; Ye, J.; Zhang, F.; et al. N6-methyladenosine demethylase FTO suppresses clear cell renal cell carcinoma through a novel FTO-PGC-1α signalling axis. J. Cell Mol. Med. 2019, 23, 2163–2173. [Google Scholar] [CrossRef]
- Shi, Y.; Fan, S.; Wu, M.; Zuo, Z.; Li, X.; Jiang, L.; Shen, Q.; Xu, P.; Zeng, L.; Zhou, Y.; et al. YTHDF1 links hypoxia adaptation and non-small cell lung cancer progression. Nat. Commun. 2019, 10, 12801–12806. [Google Scholar] [CrossRef] [Green Version]
- Fry, N.J.; Law, B.A.; Ilkayeva, O.R.; Holley, C.L.; Mansfield, K.D. N6-methyladenosine is required for the hypoxic stabilization of specific mRNAs. RNA 2017, 23, 1444–1455. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Zhu, Q.; Huang, J.; Cai, R.; Kuang, Y. Hypoxia Promotes Vascular Smooth Muscle Cell (VSMC) Differentiation of Adipose-Derived Stem Cell (ADSC) by Regulating Mettl3 and Paracrine Factors. Stem Cells Int. 2020, 2020, 1–11. [Google Scholar] [CrossRef]
- Yao, M.D.; Jiang, Q.; Ma, Y.; Liu, C.; Zhu, C.Y.; Sun, Y.N.; Shan, K.; Ge, H.M.; Zhang, Q.Y.; Zhang, H.Y.; et al. Role of METTL3-Dependent N(6)-Methyladenosine mRNA Modification in the Promotion of Angiogenesis. Mol. Ther. 2020, 28, 2191–2202. [Google Scholar] [CrossRef]
- Thalhammer, A.; Bencokova, Z.; Poole, R.; Loenarz, C.; Adam, J.; O’Flaherty, L.; Schödel, J.; Mole, D.; Giaslakiotis, K.; Schofield, C.J.; et al. Human AlkB homologue 5 is a nuclear 2-oxoglutarate dependent oxygenase and a direct target of hypoxia-inducible factor 1α (HIF-1α). PLoS ONE 2011, 6, e16210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Samanta, D.; Lu, H.; Bullen, J.W.; Zhang, H.; Chen, I.; He, X.; Semenza, G.L. Hypoxia induces the breast cancer stem cell phenotype by HIF-dependent and ALKBH5-mediated m⁶A-demethylation of NANOG mRNA. Proc. Natl. Acad. Sci. USA 2016, 113, E2047–E2056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Zhi, W.I.; Lu, H.; Samanta, D.; Chen, I.; Gabrielson, E.; Semenza, G.L. Hypoxia-inducible factors regulate pluripotency factor expression by ZNF217- and ALKBH5-mediated modulation of RNA methylation in breast cancer cells. Oncotarget 2016, 7, 64527–64542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanabe, A.; Tanikawa, K.; Tsunetomi, M.; Takai, K.; Ikeda, H.; Konno, J.; Torigoe, T.; Maeda, H.; Kutomi, G.; Okita, K.; et al. RNA helicase YTHDC2 promotes cancer metastasis via the enhancement of the efficiency by which HIF-1α mRNA is translated. Cancer Lett. 2016, 376, 34–42. [Google Scholar] [CrossRef]
- Ding, N.; You, A.; Tian, W.; Gu, L.; Deng, D. Chidamide increases the sensitivity of Non-small Cell Lung Cancer to Crizotinib by decreasing c-MET mRNA methylation. Int. J. Biol. Sci. 2020, 16, 2595–2611. [Google Scholar] [CrossRef]
- Lin, Z.; Niu, Y.; Wan, A.; Chen, D.; Liang, H.; Chen, X.; Sun, L.; Zhan, S.; Chen, L.; Cheng, C.; et al. RNA m6A methylation regulates sorafenib resistance in liver cancer through FOXO3-mediated autophagy. EMBO J. 2020, 39, e103181. [Google Scholar] [CrossRef] [PubMed]
- Taketo, K.; Konno, M.; Asai, A.; Koseki, J.; Toratani, M.; Satoh, T.; Doki, Y.; Mori, M.; Ishii, H.; Ogawa, K. The epitranscriptome m6A writer METTL3 promotes chemo- and radioresistance in pancreatic cancer cells. Int. J. Oncol. 2018, 52, 621–629. [Google Scholar] [CrossRef] [Green Version]
- Visvanathan, A.; Patil, V.; Arora, A.; Hegde, A.S.; Arivazhagan, A.; Santosh, V.; Somasundaram, K. Essential role of METTL3-mediated m6A modification in glioma stem-like cells maintenance and radioresistance. Oncogene 2018, 37, 522–533. [Google Scholar] [CrossRef]
- Bansal, H.; Yihua, Q.; Iyer, S.P.; Ganapathy, S.; Proia, D.; Penalva, L.O.; Uren, P.J.; Suresh, U.; Carew, J.S.; Karnad, A.B.; et al. WTAP is a novel oncogenic protein in acute myeloid leukemia. Leukemia 2014, 28, 1171–1174. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Kang, Y.; Wang, L.; Huff, S.; Tang, R.; Hui, H.; Agrawal, K.; Gonzalez, G.M.; Wang, Y.; Patel, S.P.; et al. ALKBH5 regulates anti–PD-1 therapy response by modulating lactate and suppressive immune cell accumulation in tumor microenvironment. Proc. Natl. Acad. Sci. USA 2020, 117, 20159–20170. [Google Scholar] [CrossRef]
- Yan, F.; Al-Kali, A.; Zhang, Z.; Liu, J.; Pang, J.; Zhao, N.; He, C.; Litzow, M.R.; Liu, S. A dynamic N6-methyladenosine methylome regulates intrinsic and acquired resistance to tyrosine kinase inhibitors. Cell Res. 2018, 28, 1062–1076. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Wei, J.; Cui, Y.-H.; Park, G.; Shah, P.; Deng, Y.; Aplin, A.E.; Lu, Z.; Hwang, S.; He, C.; et al. m6A mRNA demethylase FTO regulates melanoma tumorigenicity and response to anti-PD-1 blockade. Nat. Commun. 2019, 10, 2782. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhao, Y.; Wang, L.; Zhang, J.; Karnes, R.J.; Kohli, M.; Wang, G.; Huang, H. Alterations of androgen receptor-regulated enhancer RNAs (eRNAs) contribute to enzalutamide resistance in castration-resistant prostate cancer. Oncotarget 2016, 7, 38551–38565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, S.; Bai, Z.L.; Xia, D.; Zhao, Z.J.; Zhao, R.; Wang, Y.Y.; Zhe, H. FTO regulates the chemo-radiotherapy resistance of cervical squamous cell carcinoma (CSCC) by targeting β-catenin through mRNA demethylation. Mol. Carcinog. 2018, 57, 590–597. [Google Scholar] [CrossRef]
- He, J.-J.; Li, Z.; Rong, Z.-X.; Gao, J.; Mu, Y.; Guan, Y.-D.; Ren, X.-X.; Zi, Y.-Y.; Liu, L.-Y.; Fan, Q.; et al. m6A Reader YTHDC2 Promotes Radiotherapy Resistance of Nasopharyngeal Carcinoma via Activating IGF1R/AKT/S6 Signaling Axis. Front. Oncol. 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Wei, X.; Jian, Z.; Zhang, X. METTL3 expression is associated with glycolysis metabolism and sensitivity to glycolytic stress in hepatocellular carcinoma. Cancer Med. 2020, 9, 2859–2867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Wan, J.; Shu, X.E.; Mao, Y.; Liu, X.M.; Yuan, X.; Zhang, X.; Hess, M.E.; Brüning, J.C.; Qian, S.B. N(6)-Methyladenosine Guides mRNA Alternative Translation during Integrated Stress Response. Mol. Cell 2018, 69, 636–647.e637. [Google Scholar] [CrossRef] [Green Version]
- Gulati, P.; Cheung, M.K.; Antrobus, R.; Church, C.D.; Harding, H.P.; Tung, Y.-C.L.; Rimmington, D.; Ma, M.; Ron, D.; Lehner, P.J.; et al. Role for the obesity-related FTO gene in the cellular sensing of amino acids. Proc. Natl. Acad. Sci. USA 2013, 110, 2557–2562. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Hu, Y.; Hou, Z.; Sun, Q.; Jia, Y.; Zhao, R. Chronic corticosterone exposure induces liver inflammation and fibrosis in association with m(6)A-linked post-transcriptional suppression of heat shock proteins in chicken. Cell Stress Chaperones 2020, 25, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Miao, W.; Li, L.; Zhao, Y.; Dai, X.; Chen, X.; Wang, Y. HSP90 inhibitors stimulate DNAJB4 protein expression through a mechanism involving N6-methyladenosine. Nat. Commun. 2019, 10, 3613. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Li, Y.; Wang, T.; Zhong, X. Modification of N6-methyladenosine RNA methylation on heat shock protein expression. PLoS ONE 2018, 13, e0198604. [Google Scholar] [CrossRef] [PubMed]
- Meyer, K.D.; Patil, D.P.; Zhou, J.; Zinoviev, A.; Skabkin, M.A.; Elemento, O.; Pestova, T.V.; Qian, S.B.; Jaffrey, S.R. 5′ UTR m(6)A Promotes Cap-Independent Translation. Cell 2015, 163, 999–1010. [Google Scholar] [CrossRef] [Green Version]
- Kisliouk, T.; Rosenberg, T.; Ben-Nun, O.; Ruzal, M.; Meiri, N. Early-Life m6A RNA Demethylation by Fat Mass and Obesity-Associated Protein (FTO) Influences Resilience or Vulnerability to Heat Stress Later in Life. Eneuro 2020, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Wan, J.; Gao, X.; Zhang, X.; Jaffrey, S.R.; Qian, S.-B. Dynamic m6A mRNA methylation directs translational control of heat shock response. Nature 2015, 526, 591–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, Y.; Laurent, B.; Hsu, C.-H.; Nachtergaele, S.; Lu, Z.; Sheng, W.; Xu, C.; Chen, H.; Ouyang, J.; Wang, S.; et al. RNA m6A methylation regulates the ultraviolet-induced DNA damage response. Nature 2017, 543, 573–576. [Google Scholar] [CrossRef]
- Zhang, C.; Chen, L.; Peng, D.; Jiang, A.; He, Y.; Zeng, Y.; Xie, C.; Zhou, H.; Luo, X.; Liu, H.; et al. METTL3 and N6-Methyladenosine Promote Homologous Recombination-Mediated Repair of DSBs by Modulating DNA-RNA Hybrid Accumulation. Mol. Cell 2020, 79, 425–442.e427. [Google Scholar] [CrossRef]
- Svobodová Kovaříková, A.; Stixová, L.; Kovařík, A.; Komůrková, D.; Legartová, S.; Fagherazzi, P.; Bártová, E. N(6)-Adenosine Methylation in RNA and a Reduced m(3)G/TMG Level in Non-Coding RNAs Appear at Microirradiation-Induced DNA Lesions. Cells 2020, 9, 360. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Riddle, R.C.; Yang, Q.; Rosen, C.R.; Guttridge, D.C.; Dirckx, N.; Faugere, M.-C.; Farber, C.R.; Clemens, T.L. The RNA demethylase FTO is required for maintenance of bone mass and functions to protect osteoblasts from genotoxic damage. Proc. Natl. Acad. Sci. USA 2019, 116, 17980–17989. [Google Scholar] [CrossRef] [Green Version]
- Villeneuve, N.F.; Sun, Z.; Chen, W.; Zhang, D.D. Nrf2 and p21 regulate the fine balance between life and death by controlling ROS levels. Cell Cycle 2009, 8, 3255–3256. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Li, J.; Lou, B.; Wu, R.; Wang, G.; Lu, C.; Wang, H.; Pi, J.; Xu, Y. The Role of Reactive Oxygen Species in Arsenic Toxicity. Biomolecules 2020, 10, 240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, S.; Wheeler, J.R.; Walters, R.W.; Agrawal, A.; Barsic, A.; Parker, R. ATPase-Modulated Stress Granules Contain a Diverse Proteome and Substructure. Cell 2016, 164, 487–498. [Google Scholar] [CrossRef] [Green Version]
- Protter, D.S.W.; Parker, R. Principles and Properties of Stress Granules. Trends Cell Biol. 2016, 26, 668–679. [Google Scholar] [CrossRef] [Green Version]
- Fry, N.J.; Law, B.A.; Ilkayeva, O.R.; Carraway, K.R.; Holley, C.L.; Mansfield, K.D. N(6)-methyladenosine contributes to cellular phenotype in a genetically-defined model of breast cancer progression. Oncotarget 2018, 9, 31231–31243. [Google Scholar] [CrossRef] [Green Version]
- Green, N.H.; Galvan, D.L.; Badal, S.S.; Chang, B.H.; LeBleu, V.S.; Long, J.; Jonasch, E.; Danesh, F.R. MTHFD2 links RNA methylation to metabolic reprogramming in renal cell carcinoma. Oncogene 2019, 38, 6211–6225. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Leung, G.K.K. More Than a Metabolic Enzyme: MTHFD2 as a Novel Target for Anticancer Therapy? Front. Oncol. 2020, 10, 658. [Google Scholar] [CrossRef] [PubMed]
- Fukumoto, T.; Zhu, H.; Nacarelli, T.; Karakashev, S.; Fatkhutdinov, N.; Wu, S.; Liu, P.; Kossenkov, A.V.; Showe, L.C.; Jean, S.; et al. N6-Methylation of Adenosine of FZD10 mRNA Contributes to PARP Inhibitor Resistance. Cancer Res. 2019, 79, 2812–2820. [Google Scholar] [CrossRef] [Green Version]
- Pajak, B.; Siwiak, E.; Sołtyka, M.; Priebe, A.; Zieliński, R.; Fokt, I.; Ziemniak, M.; Jaśkiewicz, A.; Borowski, R.; Domoradzki, T.; et al. 2-Deoxy-d-Glucose and Its Analogs: From Diagnostic to Therapeutic Agents. Int. J. Mol. Sci. 2019, 21, 234. [Google Scholar] [CrossRef] [Green Version]
- Pakos-Zebrucka, K.; Koryga, I.; Mnich, K.; Ljujic, M.; Samali, A.; Gorman, A.M. The integrated stress response. EMBO Rep. 2016, 17, 1374–1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shatsky, I.N.; Terenin, I.M.; Smirnova, V.V.; Andreev, D.E. Cap-Independent Translation: What’s in a Name? Trends Biochem. Sci. 2018, 43, 882–895. [Google Scholar] [CrossRef]
- Gokhale, N.S.; McIntyre, A.B.R.; Mattocks, M.D.; Holley, C.L.; Lazear, H.M.; Mason, C.E.; Horner, S.M. Altered m(6)A Modification of Specific Cellular Transcripts Affects Flaviviridae Infection. Mol. Cell 2020, 77, 542–555.e548. [Google Scholar] [CrossRef]
- Barbieri, I.; Kouzarides, T. Role of RNA modifications in cancer. Nat. Rev. Cancer 2020, 20, 303–322. [Google Scholar] [CrossRef]
- Delaunay, S.; Frye, M. RNA modifications regulating cell fate in cancer. Nat. Cell Biol. 2019, 21, 552–559. [Google Scholar] [CrossRef]
- Huang, H.; Weng, H.; Deng, X.; Chen, J. RNA Modifications in Cancer: Functions, Mechanisms, and Therapeutic Implications. Annu. Rev. Cancer Biol. 2020, 4, 221–240. [Google Scholar] [CrossRef] [Green Version]
- Huo, F.-C.; Zhu, Z.-M.; Pei, D.-S. N6-methyladenosine (m6A) RNA modification in human cancer. Cell Prolif. 2020, 53, e12921. [Google Scholar] [CrossRef] [PubMed]
- Ianniello, Z.; Paiardini, A.; Fatica, A. N6-Methyladenosine (m6A): A Promising New Molecular Target in Acute Myeloid Leukemia. Front. Oncol. 2019, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Wang, Y.; Wu, J.; Liu, J.; Qin, Z.; Fan, H. N(6)-Methyladenosine: A Potential Breakthrough for Human Cancer. Mol. Ther. Nucleic Acids 2020, 19, 804–813. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Wan, A.; Lin, Z.; Lu, X.; Wan, G. N6-Methyladenosine modification: A novel pharmacological target for anti-cancer drug development. Acta Pharm. Sin. B 2018, 8, 833–843. [Google Scholar] [CrossRef]
- Song, H.; Liu, D.; Dong, S.; Zeng, L.; Wu, Z.; Zhao, P.; Zhang, L.; Chen, Z.-S.; Zou, C. Epitranscriptomics and epiproteomics in cancer drug resistance: Therapeutic implications. Signal Transduct. Target. Ther. 2020, 5, 193. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Chai, P.; Jia, R.; Jia, R. Novel insights on m6A RNA methylation in tumorigenesis: A double-edged sword. Mol. Cancer 2018, 17, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, G.; Sun, Z.; Zhang, N. Reshaping the role of m6A modification in cancer transcriptome: A review. Cancer Cell Int. 2020, 20, 353. [Google Scholar] [CrossRef]
- Dong, Z.; Cui, H. The Emerging Roles of RNA Modifications in Glioblastoma. Cancers (Basel) 2020, 12, 736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fazi, F.; Fatica, A. Interplay Between N6-Methyladenosine (m6A) and Non-coding RNAs in Cell Development and Cancer. Front. Cell Dev. Biol. 2019, 7. [Google Scholar] [CrossRef]
- Widagdo, J.; Anggono, V. The m6A-epitranscriptomic signature in neurobiology: From neurodevelopment to brain plasticity. J. Neurochem. 2018, 147, 137–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Zhai, Y.; Zhang, S.; Dai, X.; Li, Z. Roles of N6-Methyladenosine (m(6)A) in Stem Cell Fate Decisions and Early Embryonic Development in Mammals. Front. Cell Dev. Biol. 2020, 8, 782. [Google Scholar] [CrossRef]
- Chokkalla, A.K.; Mehta, S.L.; Vemuganti, R. Epitranscriptomic regulation by m6A RNA methylation in brain development and diseases. J. Cereb. Blood Flow Metab. 2020, 40, 2331–2349. [Google Scholar] [CrossRef] [PubMed]
- Engel, M.; Chen, A. The emerging role of mRNA methylation in normal and pathological behavior. Genes Brain Behav. 2018, 17, e12428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, C.; Chang, M.; Lv, H.; Zhang, Z.-W.; Zhang, W.; He, X.; Wu, G.; Zhao, S.; Zhang, Y.; Wang, D.; et al. RNA m6A methylation participates in regulation of postnatal development of the mouse cerebellum. Genome Biol. 2018, 19, 68. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Trebak, F.; Souza, L.A.C.; Shi, J.; Zhou, T.; Kehoe, P.G.; Chen, Q.; Feng Earley, Y. Small RNA modifications in Alzheimer’s disease. Neurobiol. Dis. 2020, 145, 105058. [Google Scholar] [CrossRef] [PubMed]
- Geula, S.; Moshitch-Moshkovitz, S.; Dominissini, D.; Mansour, A.A.; Kol, N.; Salmon-Divon, M.; Hershkovitz, V.; Peer, E.; Mor, N.; Manor, Y.S.; et al. m6A mRNA methylation facilitates resolution of naïve pluripotency toward differentiation. Science 2015, 347, 1002–1006. [Google Scholar] [CrossRef]
- Wang, C.X.; Cui, G.S.; Liu, X.; Xu, K.; Wang, M.; Zhang, X.X.; Jiang, L.Y.; Li, A.; Yang, Y.; Lai, W.Y.; et al. METTL3-mediated m6A modification is required for cerebellar development. PLoS Biol. 2018, 16, e2004880. [Google Scholar] [CrossRef] [PubMed]
- Boissel, S.; Reish, O.; Proulx, K.; Kawagoe-Takaki, H.; Sedgwick, B.; Yeo, G.S.H.; Meyre, D.; Golzio, C.; Molinari, F.; Kadhom, N.; et al. Loss-of-function mutation in the dioxygenase-encoding FTO gene causes severe growth retardation and multiple malformations. Am. J. Hum. Genet. 2009, 85, 106–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, A.G.; Batlle, E.; Ribas de Pouplana, L. Role of tRNA modifications in human diseases. Trends Mol. Med. 2014, 20, 306–314. [Google Scholar] [CrossRef]
- Abbasi-Moheb, L.; Mertel, S.; Gonsior, M.; Nouri-Vahid, L.; Kahrizi, K.; Cirak, S.; Wieczorek, D.; Motazacker, M.M.; Esmaeeli-Nieh, S.; Cremer, K.; et al. Mutations in NSUN2 cause autosomal-recessive intellectual disability. Am. J. Hum. Genet. 2012, 90, 847–855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanco, S.; Dietmann, S.; Flores, J.V.; Hussain, S.; Kutter, C.; Humphreys, P.; Lukk, M.; Lombard, P.; Treps, L.; Popis, M.; et al. Aberrant methylation of tRNAs links cellular stress to neuro-developmental disorders. EMBO J. 2014, 33, 2020–2039. [Google Scholar] [CrossRef]
- Martinez, F.J.; Lee, J.H.; Lee, J.E.; Blanco, S.; Nickerson, E.; Gabriel, S.; Frye, M.; Al-Gazali, L.; Gleeson, J.G. Whole exome sequencing identifies a splicing mutation in NSUN2 as a cause of a Dubowitz-like syndrome. J. Med. Genet. 2012, 49, 380–385. [Google Scholar] [CrossRef] [Green Version]
- Najmabadi, H.; Hu, H.; Garshasbi, M.; Zemojtel, T.; Abedini, S.S.; Chen, W.; Hosseini, M.; Behjati, F.; Haas, S.; Jamali, P.; et al. Deep sequencing reveals 50 novel genes for recessive cognitive disorders. Nature 2011, 478, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, C.; Grégoire, M.J.; Brochet, K.; Raffo, E.; Leheup, B.; Jonveaux, P. Pure de-novo 5 Mb duplication at Xp11.22–p11.23 in a male: Phenotypic and molecular characterization. J. Hum. Genet. 2006, 51, 815. [Google Scholar] [CrossRef]
- Dai, L.; Xing, L.; Gong, P.; Zhang, K.; Gao, X.; Zheng, Z.; Zhou, J.; Guo, Y.; Guo, S.; Zhang, F. Positive association of the FTSJ1 gene polymorphisms with nonsyndromic X-linked mental retardation in young Chinese male subjects. J. Hum. Genet. 2008, 53, 592–597. [Google Scholar] [CrossRef]
- Freude, K.; Hoffmann, K.; Jensen, L.R.; Delatycki, M.B.; des Portes, V.; Moser, B.; Hamel, B.; van Bokhoven, H.; Moraine, C.; Fryns, J.P.; et al. Mutations in the FTSJ1 gene coding for a novel S-adenosylmethionine-binding protein cause nonsyndromic X-linked mental retardation. Am. J. Hum. Genet. 2004, 75, 305–309. [Google Scholar] [CrossRef]
- Ramser, J.; Winnepenninckx, B.; Lenski, C.; Errijgers, V.; Platzer, M.; Schwartz, C.E.; Meindl, A.; Kooy, R.F. A splice site mutation in the methyltransferase gene FTSJ1 in Xp11.23 is associated with non-syndromic mental retardation in a large Belgian family (MRX9). J. Med. Genet. 2004, 41, 679–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takano, K.; Nakagawa, E.; Inoue, K.; Kamada, F.; Kure, S.; Goto, Y. A loss-of-function mutation in the FTSJ1 gene causes nonsyndromic X-linked mental retardation in a Japanese family. Am. J. Med. Genet. B Neuropsychiatry Genet. 2008, 147b, 479–484. [Google Scholar] [CrossRef]
- Zhang, F.; Kang, Y.; Wang, M.; Li, Y.; Xu, T.; Yang, W.; Song, H.; Wu, H.; Shu, Q.; Jin, P. Fragile X mental retardation protein modulates the stability of its m6A-marked messenger RNA targets. Hum. Mol. Genet. 2018, 27, 3936–3950. [Google Scholar] [CrossRef]
- Han, M.; Liu, Z.; Xu, Y.; Liu, X.; Wang, D.; Li, F.; Wang, Y.; Bi, J. Abnormality of m6A mRNA Methylation Is Involved in Alzheimer’s Disease. Front. Neurosci. 2020, 14. [Google Scholar] [CrossRef] [PubMed]
- Reitz, C.; Tosto, G.; Mayeux, R.; Luchsinger, J.A. Genetic variants in the Fat and Obesity Associated (FTO) gene and risk of Alzheimer’s disease. PLoS ONE 2012, 7, e50354. [Google Scholar] [CrossRef] [PubMed]
- Engel, M.; Eggert, C.; Kaplick, P.M.; Eder, M.; Röh, S.; Tietze, L.; Namendorf, C.; Arloth, J.; Weber, P.; Rex-Haffner, M.; et al. The Role of m(6)A/m-RNA Methylation in Stress Response Regulation. Neuron 2018, 99, 389–403.e389. [Google Scholar] [CrossRef] [PubMed]
- Milaneschi, Y.; Lamers, F.; Mbarek, H.; Hottenga, J.J.; Boomsma, D.I.; Penninx, B.W.J.H. The effect of FTO rs9939609 on major depression differs across MDD subtypes. Mol. Psychiatry 2014, 19, 960–962. [Google Scholar] [CrossRef] [Green Version]
- Samaan, Z.; Anand, S.; Zhang, X.; Desai, D.; Rivera, M.; Pare, G.; Thabane, L.; Xie, C.; Gerstein, H.; Engert, J.C.; et al. The protective effect of the obesity-associated rs9939609 A variant in fat mass- and obesity-associated gene on depression. Mol. Psychiatry 2013, 18, 1281–1286. [Google Scholar] [CrossRef]
- Spychala, A.; Rüther, U. FTO affects hippocampal function by regulation of BDNF processing. PLoS ONE 2019, 14, e0211937. [Google Scholar] [CrossRef]
- Frayling, T.M.; Timpson, N.J.; Weedon, M.N.; Zeggini, E.; Freathy, R.M.; Lindgren, C.M.; Perry, J.R.B.; Elliott, K.S.; Lango, H.; Rayner, N.W.; et al. A common variant in the FTO gene is associated with body mass index and predisposes to childhood and adult obesity. Science (N. Y.) 2007, 316, 889–894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreasen, C.H.; Stender-Petersen, K.L.; Mogensen, M.S.; Torekov, S.S.; Wegner, L.; Andersen, G.; Nielsen, A.L.; Albrechtsen, A.; Borch-Johnsen, K.; Rasmussen, S.S.; et al. Low physical activity accentuates the effect of the FTO rs9939609 polymorphism on body fat accumulation. Diabetes 2008, 57, 95–101. [Google Scholar] [CrossRef] [Green Version]
- Scuteri, A.; Sanna, S.; Chen, W.M.; Uda, M.; Albai, G.; Strait, J.; Najjar, S.; Nagaraja, R.; Orrú, M.; Usala, G.; et al. Genome-wide association scan shows genetic variants in the FTO gene are associated with obesity-related traits. PLoS Genet. 2007, 3, e115. [Google Scholar] [CrossRef]
- Cecil, J.E.; Tavendale, R.; Watt, P.; Hetherington, M.M.; Palmer, C.N. An obesity-associated FTO gene variant and increased energy intake in children. N. Engl. J. Med. 2008, 359, 2558–2566. [Google Scholar] [CrossRef]
- Grant, S.F.; Li, M.; Bradfield, J.P.; Kim, C.E.; Annaiah, K.; Santa, E.; Glessner, J.T.; Casalunovo, T.; Frackelton, E.C.; Otieno, F.G.; et al. Association analysis of the FTO gene with obesity in children of Caucasian and African ancestry reveals a common tagging SNP. PLoS ONE 2008, 3, e1746. [Google Scholar] [CrossRef]
- Hotta, K.; Nakata, Y.; Matsuo, T.; Kamohara, S.; Kotani, K.; Komatsu, R.; Itoh, N.; Mineo, I.; Wada, J.; Masuzaki, H.; et al. Variations in the FTO gene are associated with severe obesity in the Japanese. J. Hum. Genet. 2008, 53, 546–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prakash, J.; Srivastava, N.; Awasthi, S.; Agarwal, C.G.; Natu, S.M.; Rajpal, N.; Mittal, B. Association of FTO rs17817449 SNP with obesity and associated physiological parameters in a north Indian population. Ann. Hum. Biol. 2011, 38, 760–763. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, S.A.; Mumtaz, A.; Shahid, S.U.; Shabana, N.A. rs3751812, a common variant in fat mass and obesity-associated (FTO) gene, is associated with serum high- and low-density lipoprotein cholesterol in Pakistani individuals. Nutrition 2017, 39–40, 92–95. [Google Scholar] [CrossRef] [PubMed]
- Fischer, J.; Koch, L.; Emmerling, C.; Vierkotten, J.; Peters, T.; Brüning, J.C.; Rüther, U. Inactivation of the Fto gene protects from obesity. Nature 2009, 458, 894–898. [Google Scholar] [CrossRef]
- Church, C.; Moir, L.; McMurray, F.; Girard, C.; Banks, G.T.; Teboul, L.; Wells, S.; Brüning, J.C.; Nolan, P.M.; Ashcroft, F.M.; et al. Overexpression of Fto leads to increased food intake and results in obesity. Nat. Genet. 2010, 42, 1086–1092. [Google Scholar] [CrossRef]
- Zhao, X.; Yang, Y.; Sun, B.-F.; Shi, Y.; Yang, X.; Xiao, W.; Hao, Y.-J.; Ping, X.-L.; Chen, Y.-S.; Wang, W.-J.; et al. FTO-dependent demethylation of N6-methyladenosine regulates mRNA splicing and is required for adipogenesis. Cell Res. 2014, 24, 1403–1419. [Google Scholar] [CrossRef]
- Wu, J.; Frazier, K.; Zhang, J.; Gan, Z.; Wang, T.; Zhong, X. Emerging role of m6A RNA methylation in nutritional physiology and metabolism. Obes. Rev. 2020, 21, e12942. [Google Scholar] [CrossRef]
- De Jesus, D.F.; Zhang, Z.; Kahraman, S.; Brown, N.K.; Chen, M.; Hu, J.; Gupta, M.K.; He, C.; Kulkarni, R.N. m6A mRNA methylation regulates human β-cell biology in physiological states and in type 2 diabetes. Nat. Metab. 2019, 1, 765–774. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, J.; Lin, Z.; Zhang, W.; Wang, S.; Wang, W.; Wang, Q.; Ning, G. m6A mRNA Methylation Controls Functional Maturation in Neonatal Murine β-Cells. Diabetes 2020, 69, 1708–1722. [Google Scholar] [CrossRef]
- Shen, F.; Huang, W.; Huang, J.-T.; Xiong, J.; Yang, Y.; Wu, K.; Jia, G.-F.; Chen, J.; Feng, Y.-Q.; Yuan, B.-F.; et al. Decreased N6-Methyladenosine in Peripheral Blood RNA From Diabetic Patients Is Associated With FTO Expression Rather Than ALKBH5. J. Clin. Endocrinol. Metab. 2015, 100, E148–E154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Luo, G.; Sun, J.; Men, L.; Ye, H.; He, C.; Ren, D. METTL14 is essential for β-cell survival and insulin secretion. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 2138–2148. [Google Scholar] [CrossRef]
- Xie, W.; Ma, L.L.; Xu, Y.Q.; Wang, B.H.; Li, S.M. METTL3 inhibits hepatic insulin sensitivity via N6-methyladenosine modification of Fasn mRNA and promoting fatty acid metabolism. Biochem. Biophys. Res. Commun. 2019, 518, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, S.; Li, J.; Han, C.; Wang, Z.; Li, C.; Wang, X.; Liu, Z.; Wen, J.; Zheng, L. Expression and significance of fat mass and obesity associated gene and forkhead transcription factor O1 in non-alcoholic fatty liver disease. Chin. Med. J. (Engl.) 2014, 127, 3771–3776. [Google Scholar] [PubMed]
- Benedict, M.; Zhang, X. Non-alcoholic fatty liver disease: An expanded review. World J. Hepatol. 2017, 9, 715–732. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Overview of RNA modifications. (A). Writers, erasers and readers involved in catalyzing various RNA modifications. (B). Noted are RNA modifications that have been identified on tRNA, rRNA, mRNA, miRNA, lncRNA, circRNA [1,2,6,7,8,9,10,11]. The schematic was created using BioRender.

Figure 2.
RNA modifications in response to oxidative stress. Highlighted are the diverse pathways and mechanisms by which RNA modifications respond to oxidative stress. Featured pathways include: (A). Antioxidant response. (B). Protein translation. (C). Response to arsenite. (D). Mitochondrial respiration. (E). Lipid metabolism. (F). Stress granule formation. The schematic was created using BioRender.
Figure 2.
RNA modifications in response to oxidative stress. Highlighted are the diverse pathways and mechanisms by which RNA modifications respond to oxidative stress. Featured pathways include: (A). Antioxidant response. (B). Protein translation. (C). Response to arsenite. (D). Mitochondrial respiration. (E). Lipid metabolism. (F). Stress granule formation. The schematic was created using BioRender.

Figure 3.
RNA Modifications in cellular stress responses. Highlighted are the roles of RNA modifications in response to cellular stresses. (A). Oxidative Stress. (B). Hypoxia. (C). Therapeutic Stress. (D). Metabolic Stress. (E). Heat Shock. (F). DNA damage. (G). ER Stress. The schematic was made using BioRender.
Figure 3.
RNA Modifications in cellular stress responses. Highlighted are the roles of RNA modifications in response to cellular stresses. (A). Oxidative Stress. (B). Hypoxia. (C). Therapeutic Stress. (D). Metabolic Stress. (E). Heat Shock. (F). DNA damage. (G). ER Stress. The schematic was made using BioRender.

Figure 4.
RNA Modifications in diseases. Highlighted are the regulators of RNA modifications that have established roles in regulating disease pathogenesis across genders as well as sex-specific diseases such as breast cancer, gynecologic cancers, and prostate cancer. Windows in red are modifiers implicated in cancer. Windows in purple are metabolic diseases. Windows in green are neurologic diseases. Not pictured are developmental disorders. The schematic was created using BioRender.
Figure 4.
RNA Modifications in diseases. Highlighted are the regulators of RNA modifications that have established roles in regulating disease pathogenesis across genders as well as sex-specific diseases such as breast cancer, gynecologic cancers, and prostate cancer. Windows in red are modifiers implicated in cancer. Windows in purple are metabolic diseases. Windows in green are neurologic diseases. Not pictured are developmental disorders. The schematic was created using BioRender.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary of the role of RNA modifications in stress responses.
| Stressor | Regulators | Response to Stresses | Cells or Organisms | References |
|---|---|---|---|---|
| Oxidative Stress | METTL3/14 | Increases in response to ROS, activates p21/NRF2, deposits m6A onto SGs | mRTECs, HeLa, colon cancer cell lines, HepG2, Hepa1-6, HEK293, U2OS | [18,19,20,21] |
| METTL14 | Increased upon arsenite-induced ROS | Keratinocytes | [22] | |
| WTAP | Increased upon arsenite-induced ROS | Keratinocytes | [22] | |
| NSUN2 | Alters protein synthesis, enhances p21 translation | HeLa, colon cancer cell lines, keratinocytes, | [20,23] | |
| NSUN5 | Alters protein synthesis | Glioma cell lines | [24] | |
| FTO | Promotes mitochondrial and lipogenesis-induced ROS | HEK293T/kidney cancer cell lines, myotubes, L02 cells | [25,26,27] | |
| YTHDF1 | Negative regulator of NRF2 | Tibetan mammals | [28] | |
| YTHDF2 | Increases in response to ROS | HepG2, Hepa1-6 | [21] | |
| YTHDF3 | Selective SG-reader in response to oxidative stress | HEK293T, U2OS | [19] | |
| Hypoxia | METTL3 | Promotes stability of hypoxia-response genes | HEK293T | [29] |
| METTL3/14 | Hypoxia-response gene | ADSCs, HUVECs | [30,31] | |
| ALKBH5 | Induced by HIF1, promotes BCSC phenotype | U20S, MCF7 and other breast cancer cell lines, IMR32, HeLa | [32,33,34] | |
| YTHDF1 | Promotes hypoxia adaptation | Tibetan mammals | [28] | |
| YTHDC2 | Promotes HIF1 translation | HT29, HCT116, COS | [35] | |
| Therapeutic Stress | METTL3 | Sensitive to sorafenib but increased resistance to gemcitabine, 5-FU, cisplatin, temozolomide, and radiotherapy; targeted by chidamide | HEK293T, HCC, NSCLC, PDAC and AML cell lines, GBM tissues and GSCs | [36,37,38,39] |
| WTAP | Promotes etoposide resistance and is targeted by chidamide | NSCLC and AML cell lines | [36,40] | |
| ALKBH5 | Promotes resistance to anti-PD-1 therapy | Melanoma cell lines | [41] | |
| FTO | Promotes resistance to TKIs, cisplatin, enzalutamide, and anti-PD-1 therapies | Leukemia, melanoma, CSCC, protstate cancer, and ovarian cancer cell lines | [42,43,44,45] | |
| YTHDC2 | Promotes radioresistance | NPC cell lines | [46] | |
| Metabolic Stress | METTL3 | Combats glycolytic stress | HCC cell lines | [47] |
| ALKBH5 | Promotes ATF4 translation | MEF | [48] | |
| FTO | Promotes adaptation to metabolic stress and regulates mTORC1 | Melanoma cell lines, MEF | [43,49] | |
| Heat Shock | METTL3 | Regulates m6A deposition onto HSP70, HSP60, HSP27, and DNAJB4 | HepG2, M14, HeLa, HEK293T, male chickens, | [50,51,52,53] |
| FTO | Serves a protective role | Male Cobb chicks | [54] | |
| YTHDF2 | Negative regulator of HSP90, HSP60 and HSPB1; HSP70 reader | HepG2, HeLa, MEF | [52,55] | |
| DNA Damage | METTL3 | Recruited to DNA damage sites post-UV | HEK293T, U2OS, HeLa, A375, MEF, CAL-27 | [56,57] |
| METTL16 | Recruited to DNA damage sites post-UV | MEF, HaCaT, U2OS, HeLa | [58] | |
| YTHDC1 | Recruited to DNA-RNA hybrids, recruit HR proteins | HEK293T, U2OS, CAL-27 | [57] | |
| ER Stress | FTO | Induces ER stress pathways post-genotoxic damage | Osteoblasts | [59] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wilkinson, E.; Cui, Y.-H.; He, Y.-Y. Context-Dependent Roles of RNA Modifications in Stress Responses and Diseases. Int. J. Mol. Sci. 2021, 22, 1949. https://doi.org/10.3390/ijms22041949
AMA Style
Wilkinson E, Cui Y-H, He Y-Y. Context-Dependent Roles of RNA Modifications in Stress Responses and Diseases. International Journal of Molecular Sciences. 2021; 22(4):1949. https://doi.org/10.3390/ijms22041949
Chicago/Turabian StyleWilkinson, Emma, Yan-Hong Cui, and Yu-Ying He. 2021. "Context-Dependent Roles of RNA Modifications in Stress Responses and Diseases" International Journal of Molecular Sciences 22, no. 4: 1949. https://doi.org/10.3390/ijms22041949
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.