Potential Mechanisms of AtNPR1 Mediated Resistance against Huanglongbing (HLB) in Citrus

 ,
, 
Abstract
:1. Introduction
2. Results
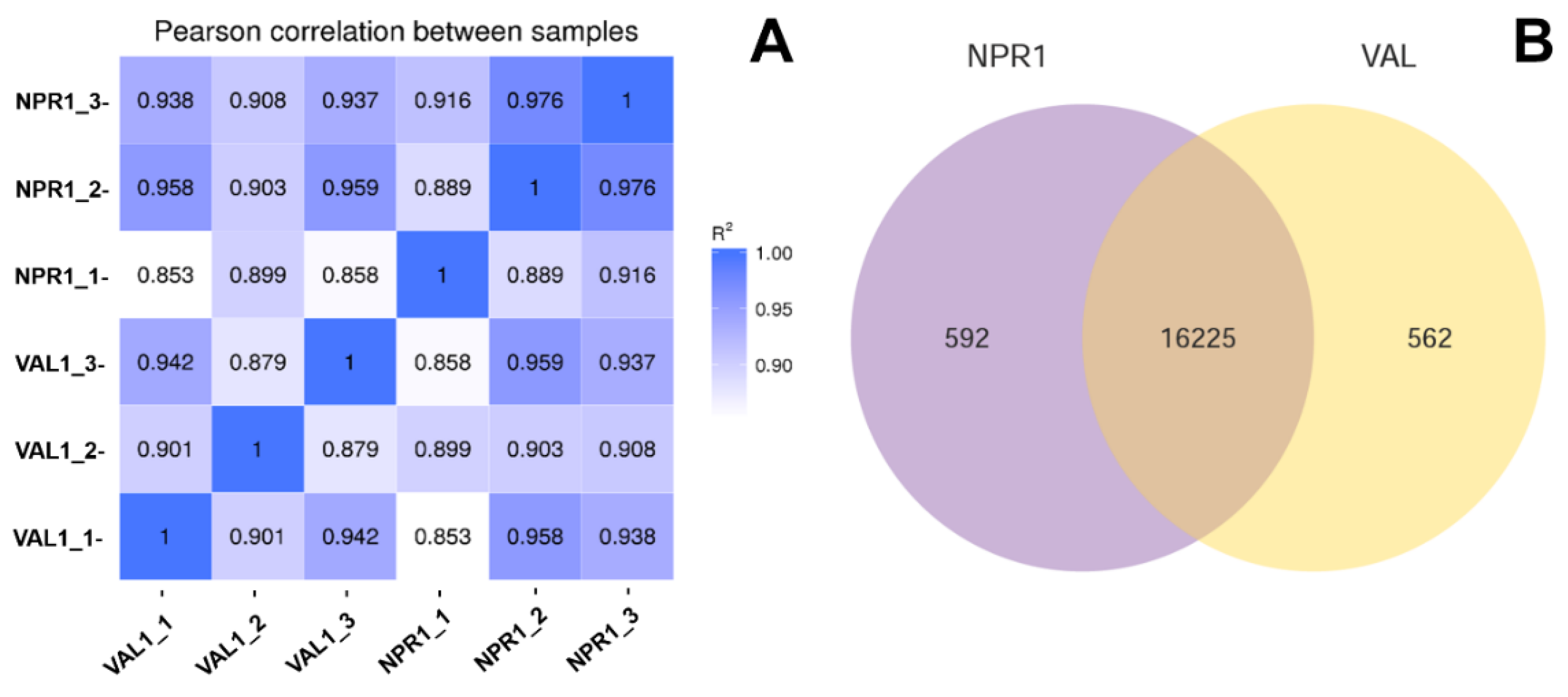
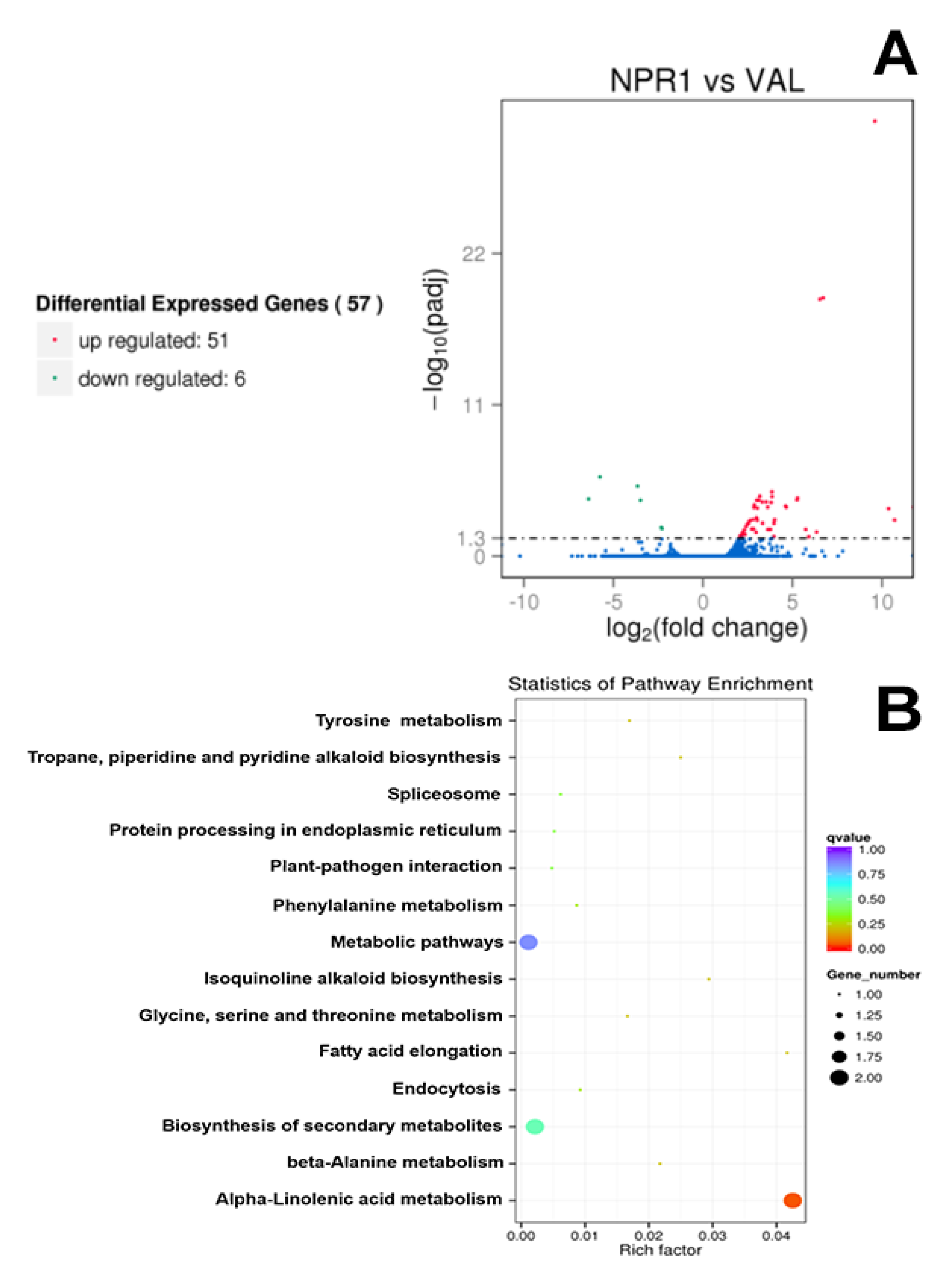
2.1. Transcriptome Sequencing and Differentially Expressed Genes (DEGs) Identification
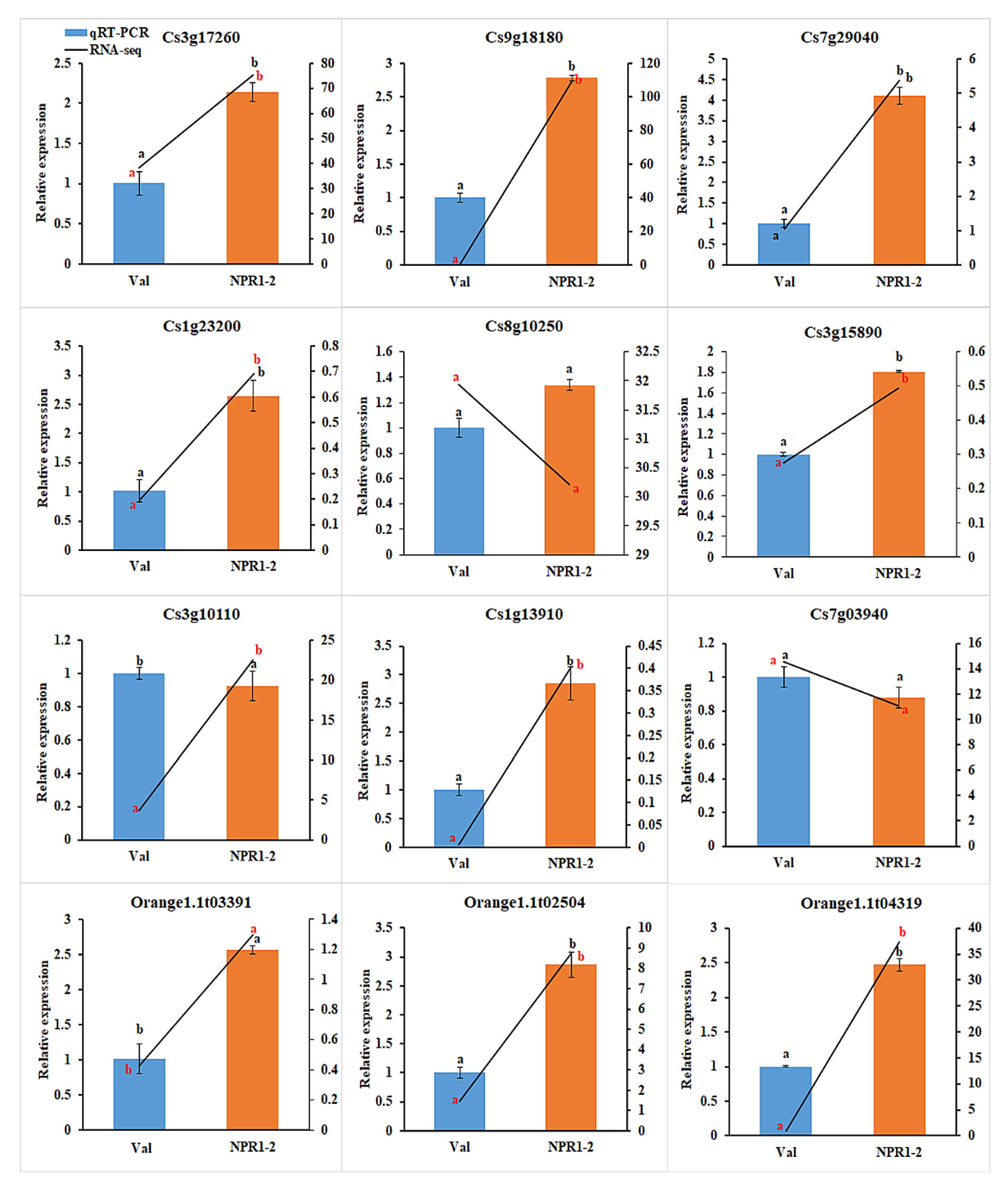
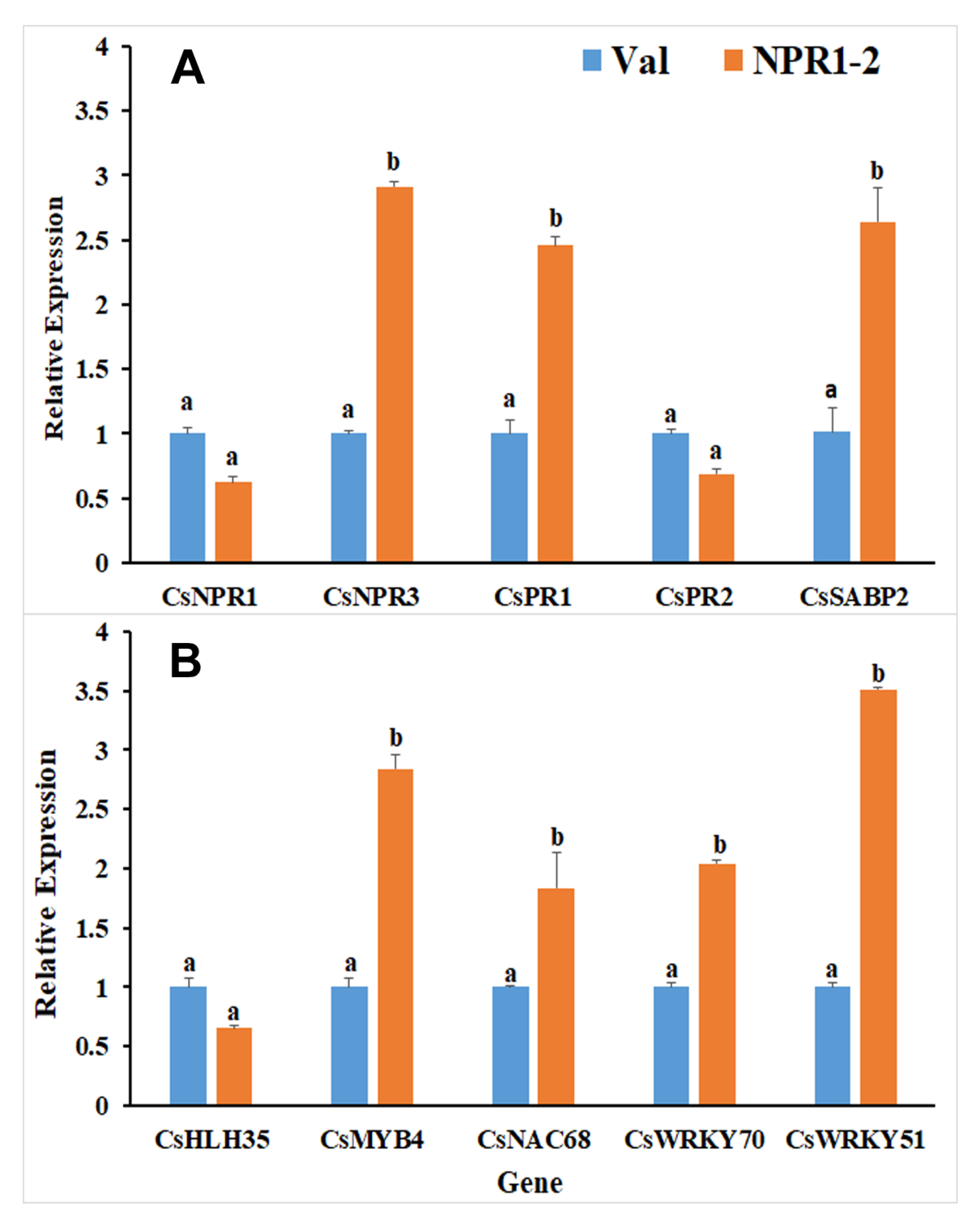
2.2. Expression of DEGs Candidate in RNAseq Correlates with qRT-PCR
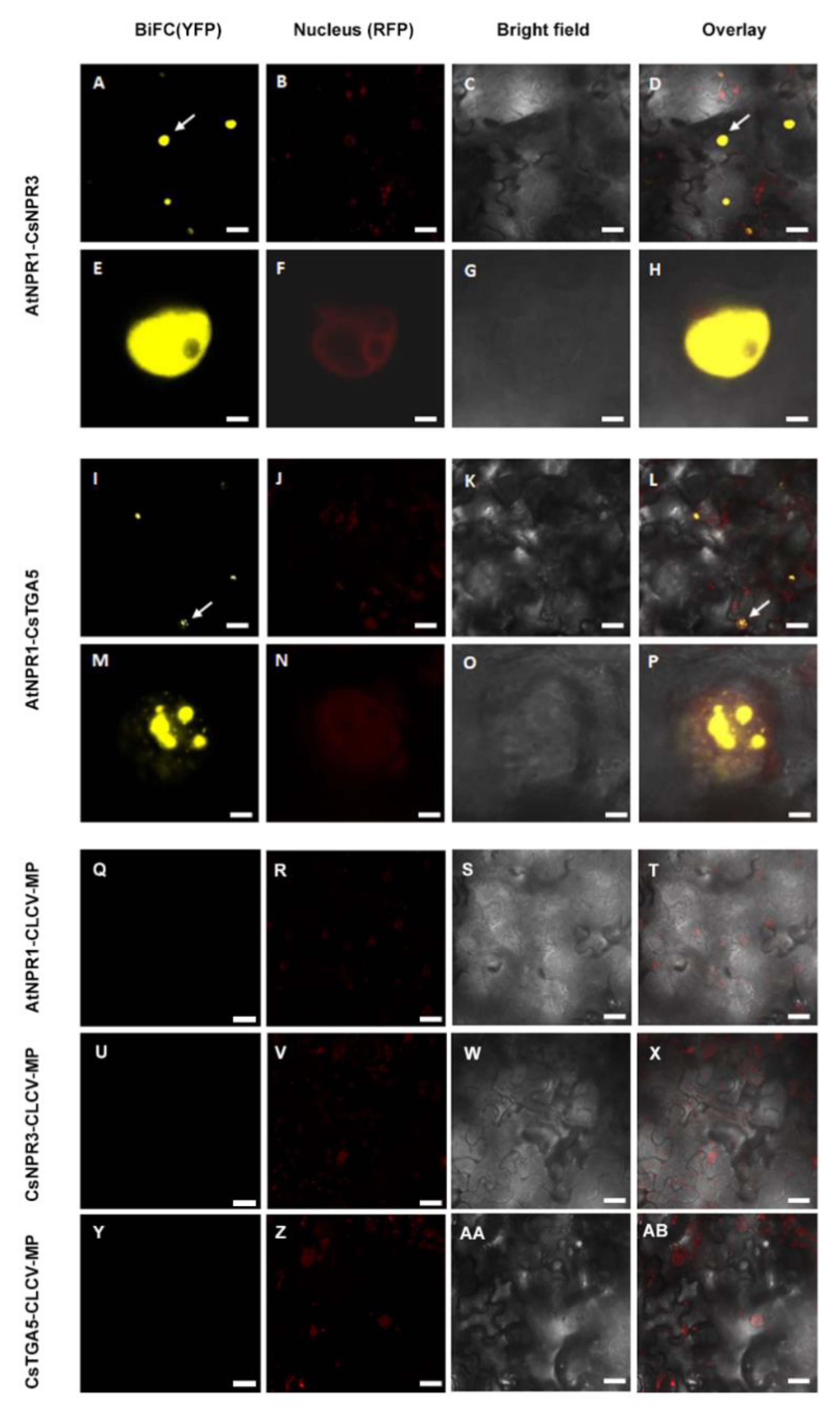
2.3. AtNPR1 Interacts with Citrus Homolog Proteins
3. Discussion
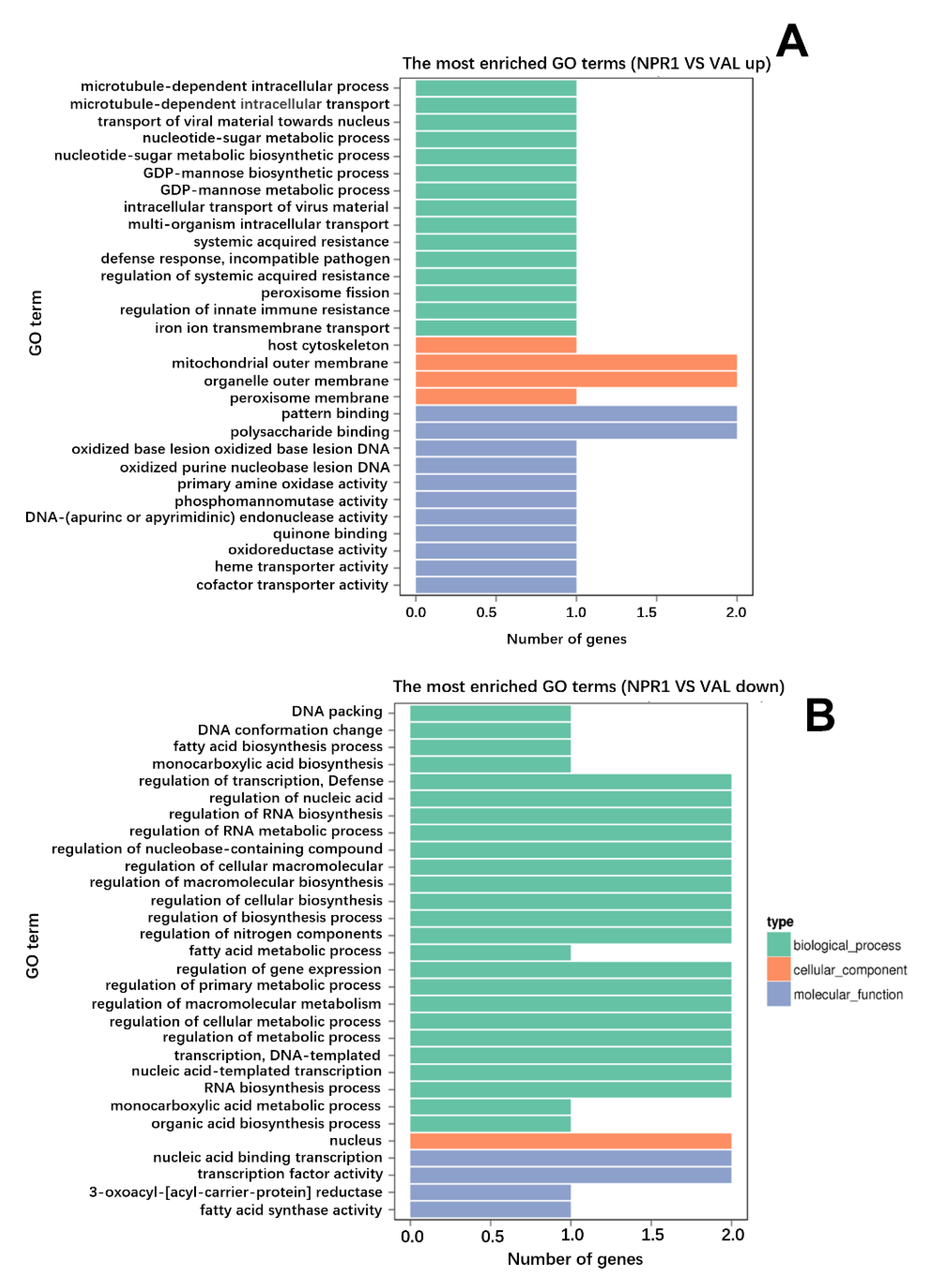
3.1. The Transcriptional Change in NPR1-2 Transgenic Line
3.2. AtNPR1 Interacted with NPR Orthologs Proteins
4. Materials and Methods
4.1. Plant Materials
4.2. RNA Extraction and cDNA Synthesis
4.3. RNAseq Libraries Construction and Sequencing
4.4. Data Mapping and Differentially Expressed Genes Identification
4.5. GO and KEGG Pathway Analysis
4.6. Quantitative Real-Time RT-PCR (qRT-PCR) Analysis
4.7. Vector Construction Using Gateway Technology
4.8. Bimolecular Fluorescence Complementation Assays
4.9. Yeast Two-Hybrid Assays
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Duan, Y.; Zhou, L.; Hall, D.G.; Li, W.; Doddapaneni, H.; Lin, H.; Liu, L.; Vahling, C.M.; Gabriel, D.W.; Williams, K.P.; et al. Complete genome sequence of citrus Huanglongbing bacterium, ‘Candidatus Liberibacter Asiaticus’ obtained through Metagenomics. Mol. Plant Microbe Interact. 2009, 22, 1011–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Q.; Pitino, M.; Zhang, S.; Krystel, J.; Cano, L.M.; Shatters, R.G.; Hall, D.G.; Stover, E. Temporal and spatial detection of Candidatus Liberibacter asiaticus putative effector transcripts during interaction with Huanglongbing-susceptible, −tolerant, and -resistant citrus hosts. BMC Plant Biol. 2019, 19, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qureshi, J.A.; Kostyk, B.C.; Stansly, P.A. Insecticidal suppression of Asian citrus Psyllid Diaphorina citri (Hemiptera: Liviidae) vector of Huanglongbing pathogens. PLoS ONE 2014, 9, e112331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N. The citrus Huanglongbing crisis and potential solutions. Mol. Plant 2019, 12, 607–609. [Google Scholar] [CrossRef]
- Zhang, M.; Guo, Y.; Powell, C.A.; Doud, M.S.; Yang, C.; Duan, Y. Effective antibiotics against ‘Candidatus Liberibacter asiaticus’ in HLB-affected citrus plants identified via the graft-based evaluation. PLoS ONE 2014, 9, e111032. [Google Scholar] [CrossRef] [Green Version]
- Young, M.; Ozcan, A.; Myers, M.E.; Johnson, E.G.; Graham, J.H.; Santra, S. Multimodal generally recognized as safe ZnO/nanocopper composite: A novel antimicrobial material for the management of citrus phytopathogens. J. Agric. Food Chem. 2017, 66, 6604–6608. [Google Scholar] [CrossRef]
- Hoffman, M.T.; Doud, M.S.; Williams, L.; Zhang, M.-Q.; Ding, F.; Stover, E.; Hall, D.; Zhang, S.; Jones, L.; Gooch, M. Heat treatment eliminates ‘Candidatus Liberibacter asiaticus’ from infected citrus trees under controlled conditions. Phytopathology 2013, 103, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Jiang, J.; Wang, N. Control of citrus Huanglongbing via trunk injection of plant defense activators and antibiotics. Phytopathology 2017, 108, 186–195. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Trivedi, P.; Wang, N. Field evaluation of plant defense inducers for the control of citrus huanglongbing. Phytopathology 2015, 106, 37–46. [Google Scholar] [CrossRef]
- Stansly, P.A.; Arevalo, H.A.; Qureshi, J.A.; Jones, M.M.; Hendricks, K.; Roberts, P.D.; Roka, F.M. Vector control and foliar nutrition to maintain economic sustainability of bearing citrus in Florida groves affected by huanglongbing. Pest Manag. Sci. 2014, 70, 415–426. [Google Scholar] [CrossRef]
- Dong, X. NPR1, all things considered. Curr. Opin. Plant Biol. 2004, 7, 547–552. [Google Scholar] [CrossRef] [PubMed]
- Pieterse, C.M.J.; Van Loon, L.C. NPR1: The spider in the web of induced resistance signaling pathways. Curr. Opin. Plant Biol. 2004, 7, 456–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Köhler, C.; Merkle, T.; Roby, D.; Neuhaus, G. Developmentally regulated expression of a cyclic nucleotide-gated ion channel from Arabidopsis indicates its involvement in programmed cell death. Planta 2001, 213, 327–332. [Google Scholar] [CrossRef]
- Cao, H.; Bowling, S.A.; Gordon, A.S.; Dong, X. Characterization of an arabidopsis mutant that is nonresponsive to inducers of systemic acquired resistance. Plant Cell 1994, 6, 1583–1592. [Google Scholar] [CrossRef] [PubMed]
- Delaney, T.; Friedrich, L.; Ryals, J. Arabidopsis signal transduction mutant defective in chemically and biologically induced disease resistance. Proc. Natl. Acad. Sci. USA 1995, 92, 6602–6606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mou, Z.; Fan, W.; Dong, X. Inducers of plant systemic acquired resistance regulate NPR1 function through redox changes. Cell 2003, 113, 935–944. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Tessaro, M.J.; Lassner, M.; Li, X. Knockout analysis of Arabidopsis transcription factors TGA2, TGA5, and TGA6 reveals their redundant and essential roles in systemic acquired resistance. Plant Cell 2003, 15, 2647–2653. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Sun, T.; Ao, K.; Peng, Y.; Zhang, Y.; Li, X.; Zhang, Y. Opposite roles of salicylic acid receptors NPR1 and NPR3/NPR4 in transcriptional regulation of plant immunity. Cell 2018, 173, 1454–1467.e15. [Google Scholar] [CrossRef]
- Zhang, Y.; Cheng, Y.T.; Qu, N.; Zhao, Q.; Bi, D.; Li, X. Negative regulation of defense responses in Arabidopsis by two NPR1 paralogs. Plant J. 2006, 48, 647–656. [Google Scholar] [CrossRef]
- Fitzgerald, H.A.; Chern, M.S.; Navarre, R.; Ronald, P.C. Overexpression of (At)NPR1 in rice leads to a BTH- and environment-induced lesion-mimic/cell death phenotype. Mol. Plant Microbe Interact. 2004, 17, 140–151. [Google Scholar] [CrossRef] [Green Version]
- Molla, K.A.; Karmakar, S.; Chanda, P.K.; Sarkar, S.N.; Datta, S.K.; Datta, K. Tissue-specific expression of Arabidopsis NPR1 gene in rice for sheath blight resistance without compromising phenotypic cost. Plant Sci. 2016, 250, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Makandar, R.; Essig, J.S.; Schapaugh, M.A.; Trick, H.N.; Shah, J. Genetically engineered resistance to Fusarium head blight in wheat by expression of Arabidopsis NPR1. Mol. Plant Microbe Interact. 2006, 19, 123–129. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Gao, R.; Cui, R.; Lü, B.; Li, X.; Zhao, Y.; You, Z.; Tian, S.; Dong, H. Tobacco TTG2 suppresses resistance to pathogens by sequestering NPR1 from the nucleus. J. Cell Sci. 2012, 125, 4913–4922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Gao, H.; Li, R.; Han, D.; Wang, L.; Wu, J.; Xu, P.; Zhang, S. GmBTB/POZ, a novel BTB/POZ domain-containing nuclear protein, positively regulates the response of soybean to Phytophthora sojae infection. Mol. Plant Pathol. 2019, 20, 78–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.; Zhang, L.; Li, L.; Chen, M.; Shi, P.; Shen, Q.; Yan, T.; Hao, X.; Fu, X.; Ma, Y.; et al. Interaction of AaTGA6 with salicylic acid signaling modulates Artemisinin biosynthesis in Artemisia Annua. J. Exp. Bot. 2019, 70, 3969–3979. [Google Scholar] [CrossRef] [Green Version]
- Babourina, O.; Rengel, Z.; Jayakannan, M.; Bose, J.; Shabala, S.; Poschenrieder, C.; Massart, A. The NPR1-dependent salicylic acid signalling pathway is pivotal for enhanced salt and oxidative stress tolerance in Arabidopsis. J. Exp. Bot. 2015, 66, 1865–1875. [Google Scholar] [CrossRef] [Green Version]
- La, V.H.; Lee, B.R.; Islam, M.T.; Park, S.-H.; Jung, H.I.; Bae, D.W.; Kim, T.H. Characterization of salicylic acid-mediated modulation of the drought stress responses: Reactive oxygen species, proline, and redox state in Brassica napus. Environ. Exp. Bot. 2019, 157, 1–10. [Google Scholar] [CrossRef]
- Olate, E.; Jiménez-Gómez, J.M.; Holuigue, L.; Salinas, J. NPR1 mediates a novel regulatory pathway in cold acclimation by interacting with HSFA1 factors. Nat. Plants 2018, 4, 811–823. [Google Scholar] [CrossRef]
- Silva, K.J.P.; Mahna, N.; Mou, Z.; Folta, K.M. NPR1 as a transgenic crop protection strategy in horticultural species. Hortic. Res. 2018, 5, 15. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Francis, M.I.; Dawson, W.O.; Graham, J.H.; Orbović, V.; Triplett, E.W.; Mou, Z. Over-expression of the Arabidopsis NPR1 gene in citrus increases resistance to citrus canker. Eur. J. Plant Pathol. 2010, 128, 91–100. [Google Scholar] [CrossRef]
- Dutt, M.; Barthe, G.; Irey, M.; Grosser, J. Transgenic citrus expressing an Arabidopsis NPR1 gene exhibit enhanced resistance against Huanglongbing (HLB; Citrus Greening). PLoS ONE 2015, 10, e0137134. [Google Scholar] [CrossRef] [PubMed]
- Pajerowska-Mukhtar, K.M.; Emerine, D.K.; Mukhtar, M.S. Tell me more: Roles of NPRs in plant immunity. Trends Plant Sci. 2013, 18, 402–411. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.L.; Zhao, Y. Transcriptome comparison and gene coexpression network analysis provide a systems view of citrus response to ‘Candidatus Liberibacter asiaticus’ infection. BMC Genom. 2013, 14, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, Y.; Cheng, C.Z.; Jiang, N.H.; Jiang, B.; Zhang, Y.Y.; Wu, B.; Hu, M.L.; Zeng, J.W.; Yan, H.X.; Yi, G.J.; et al. Comparative Transcriptome and iTRAQ proteome analyses of citrus root responses to Candidatus Liberibacter Asiaticus infection. PLoS ONE 2015, 10, e0126973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rawat, N.; Kiran, S.P.; Du, D.; Gmitter, F.G.; Deng, Z. Comprehensive meta-analysis, co-expression, and miRNA nested network analysis identifies gene candidates in citrus against Huanglongbing disease. BMC Plant Biol. 2015, 15, 184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nwugo, C.C.; Doud, M.S.; Duan, Y.-P.; Lin, H. Proteomics analysis reveals novel host molecular mechanisms associated with thermotherapy of ‘Ca. Liberibacter asiaticus’-infected citrus plants. BMC Plant Biol. 2016, 16, 253. [Google Scholar] [CrossRef] [Green Version]
- Killiny, N.; Jones, S.E.; Nehela, Y.; Hijaz, F.; Dutt, M.; Gmitter, F.G.; Grosser, J.W. All roads lead to Rome: Towards understanding different avenues of tolerance to huanglongbing in citrus cultivars. Plant Physiol. Biochem. 2018, 129, 1–10. [Google Scholar] [CrossRef]
- Yao, L.; Yu, Q.; Huang, M.; Hung, W.; Grosser, J.; Chen, S.; Wang, Y.; Gmitter, F.G. Proteomic and metabolomic analyses provide insight into the off-flavour of fruits from citrus trees infected with ‘Candidatus Liberibacter asiaticus’. Hortic. Res. 2019, 6, 31. [Google Scholar] [CrossRef]
- Wang, Y.; Zhou, L.; Yu, X.; Stover, E.; Luo, F.; Duan, Y. Transcriptome profiling of Huanglongbing (HLB) tolerant and susceptible citrus plants reveals the role of basal resistance in HLB tolerance. Front. Plant Sci. 2016, 7, 933. [Google Scholar] [CrossRef] [Green Version]
- Yan, C.; Fan, M.; Yang, M.; Zhao, J.; Zhang, W.; Su, Y.; Xiao, L.; Deng, H.; Xie, D. Injury activates Ca2+/Calmodulin-dependent phosphorylation of JAV1-JAZ8-WRKY51 complex for jasmonate biosynthesis. Mol. Cell 2018, 70, 136–149. [Google Scholar] [CrossRef] [Green Version]
- Ma, W.; Ali, R.; Berkowitz, G.A. Characterization of plant phenotypes associated with loss-of-function of AtCNGC1, a plant cyclic nucleotide gated cation channel. Plant Physiol. Biochem. 2006, 44, 494–505. [Google Scholar] [CrossRef]
- Tian, W.; Hou, C.; Ren, Z.; Wang, C.; Zhao, F.; Dahlbeck, D.; Hu, S.; Zhang, L.; Niu, Q.; Li, L.; et al. A calmodulin-gated calcium channel links pathogen patterns to plant immunity. Nature 2019, 572, 131–135. [Google Scholar] [CrossRef]
- Yoshioka, K.; Moeder, W.; Kang, H.G.; Kachroo, P.; Masmoudi, K.; Berkowitz, G.; Klessig, D.F. The chimeric Arabidopsis CYCLIC NUCLEOTIDE-GATED ION CHANNEL11/12 activates multiple pathogen resistance responses. Plant Cell 2006, 18, 747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urquhart, W.; Gunawardena, A.H.; Moeder, W.; Ali, R.; Berkowitz, G.A.; Yoshioka, K. The chimeric cyclic nucleotide-gated ion channel ATCNGC11/12 constitutively induces programmed cell death in a Ca2+ dependent manner. Plant Mol. Biol. 2007, 65, 747–761. [Google Scholar] [CrossRef]
- Romeis, T. Protein kinases in the plant defence response. Curr. Opin. Plant Biol. 2001, 4, 407–414. [Google Scholar] [CrossRef]
- Hurni, S.; Scheuermann, D.; Krattinger, S.G.; Kessel, B.; Wicker, T.; Herren, G.; Fitze, M.N.; Breen, J.; Presterl, T.; Ouzunova, M.; et al. The maize disease resistance gene Htn1 against northern corn leaf blight encodes a wall-associated receptor-like kinase. Proc. Natl. Acad. Sci. USA 2015, 112, 8780–8785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.L.; Yu, Q.Y.; Tang, L.L.; Ji, W.; Bai, X.; Cai, H.; Liu, X.F.; Ding, X.D.; Zhu, Y.M. GsSRK, a G-type lectin S-receptor-like serine/threonine protein kinase, is a positive regulator of plant tolerance to salt stress. J. Plant Physiol. 2013, 170, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Voorrips, R.E.; van’t Westende, W.; van Kaauwen, M.; Visser, R.G.F.; Vosman, B. Aphid resistance in Capsicum maps to a locus containing LRR-RLK gene analogues. Theor. Appl. Genet. 2019. [Google Scholar] [CrossRef] [Green Version]
- Pandey, S.P.; Somssich, I.E. The role of WRKY transcription factors in plant immunity. Plant Physiol. 2009, 150, 1648–1655. [Google Scholar] [CrossRef] [Green Version]
- Gao, Q.-M.; Venugopal, S.; Navarre, D.; Kachroo, A. Low oleic acid-derived repression of Jasmonic acid-inducible defense responses requires the WRKY50 and WRKY51 proteins. Plant Physiol. 2011, 155, 464–476. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Brader, G.; Kariola, T.; Tapio Palva, E. WRKY70 modulates the selection of signaling pathways in plant defense. Plant J. 2006, 46, 477–491. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.Q.; Yan, S.; Saleh, A.; Wang, W.; Ruble, J.; Oka, N.; Mohan, R.; Spoel, S.H.; Tada, Y.; Zheng, N. NPR3 and NPR4 are receptors for the immune signal salicylic acid in plants. Nature 2012, 486, 228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Zhang, D.; Chu, J.Y.; Boyle, P.; Wang, Y.; Brindle, I.D.; De Luca, V.; Després, C. The Arabidopsis NPR1 protein is a receptor for the plant defense hormone salicylic acid. Cell Rep. 2012, 1, 639–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Liu, Q.; Tao, N.; Deng, X. Efficient isolation of RNA from fruit peel and pulp of ripening navel orange (Citrus sinensis Osbeck). J. Huazhong Agric. Univ. 2006, 25, 300–304. [Google Scholar]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [Green Version]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mafra, V.; Kubo, K.S.; Alves-Ferreira, M.; Ribeiro-Alves, M.; Stuart, R.M.; Boava, L.P.; Rodrigues, C.M.; Machado, M.A. Reference genes for accurate transcript normalization in citrus genotypes under different experimental conditions. PLoS ONE 2012, 7, e31263. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Burow, M.D.; Chlan, C.A.; Sen, P.; Lisca, A.; Murai, N. High-frequency generation of transgenic tobacco plants after modified leaf disk cocultivation with Agrobacterium tumefaciens. Plant Mol. Biol. Report. 1990, 8, 124–139. [Google Scholar] [CrossRef]
- Bracha-Drori, K.; Shichrur, K.; Katz, A.; Oliva, M.; Angelovici, R.; Yalovsky, S.; Ohad, N. Detection of protein–protein interactions in plants using bimolecular fluorescence complementation. Plant J. 2004, 40, 419–427. [Google Scholar] [CrossRef]
- Martin, K.; Kopperud, K.; Chakrabarty, R.; Banerjee, R.; Brooks, R.; Goodin, M.M. Transient expression in Nicotiana benthamiana fluorescent marker lines provides enhanced definition of protein localization, movement and interactions in planta. Plant J. 2009, 59, 150–162. [Google Scholar] [CrossRef] [PubMed]
- Sparkes, I.A.; Runions, J.; Kearns, A.; Hawes, C. Rapid, transient expression of fluorescent fusion proteins in tobacco plants and generation of stably transformed plants. Nat. Protoc. 2006, 1, 2019–2025. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample * | Clean Reads | Clean Bases | ≥Q30 (%) | GC Content (%) | Total Mapped (%) | Uniquely Mapped (%) |
|---|---|---|---|---|---|---|
| NPR1_1 | 62,607,264 | 7.91G | 92.89 | 44.84 | 47,656,347 (76.12%) | 46,101,606 (73.64%) |
| NPR1_2 | 59,175,150 | 7.40G | 92.79 | 45.23 | 45,972,884 (77.69%) | 44,572,333 (75.32%) |
| NPR1_3 | 59,225,298 | 7.41G | 92.58 | 44.92 | 47,206,105 (79.71%) | 45,624,108 (77.03%) |
| Val_1 | 65,551,728 | 8.20G | 92.44 | 44.4 | 54,260,656 (82.78%) | 52,711,952 (80.41%) |
| Val_2 | 65,372,880 | 8.18G | 92.84 | 44.19 | 54,287,742 (83.04%) | 52,522,887 (80.34%) |
| Val_3 | 61,717,406 | 7.72G | 92.7 | 44.59 | 51,647,046 (83.68%) | 50,005,354 (81.02%) |
| Total/Average | 373,649,726 | 47.00G | 92.71 | 44.69 | 50,171,797 (80.50%) | 48,589,707 (77.96%) |
| Gene ID | log2 (Fold Change) | padj | Functional Description | E-Value |
|---|---|---|---|---|
| Cs9g18210 | Inf | 2.76E-04 | Cyclic nucleotide-gated ion channel 1 | 1.26E-100 |
| orange1.1t01447 | 10.70 | 2.35E-03 | Transactivator/viroplasmin protein | 2.95E-06 |
| orange1.1t01448 | 10.36 | 3.52E-04 | Putative Polyprotein CP | 2.51E-07 |
| Cs9g18180 | 9.60 | 2.48E-32 | Cyclic nucleotide-gated ion channel 1 | 2.41E-98 |
| orange1.1t04318 | 6.71 | 1.70E-19 | Ankyrin repeat-containing protein | 1.39E-15 |
| orange1.1t04316 | 6.52 | 2.20E-19 | Ankyrin repeat-containing protein | 5.38E-21 |
| Cs1g13910 | 6.34 | 1.76E-02 | Wall-associated receptor kinase-like 10 | 3.06E-167 |
| Cs9g18240 | 5.91 | 3.92E-02 | Cyclic nucleotide-gated ion channel 1 | 1.81E-129 |
| Cs1g03870 | 5.27 | 5.92E-05 | Probable WRKY transcription factor 51 | 8.66E-38 |
| orange1.1t04319 | 5.26 | 8.15E-05 | Ankyrin repeat-containing protein | 5.68E-07 |
| Cs2g27430 | 4.00 | 2.22E-03 | MYB-related protein Myb4 | 1.25E-50 |
| orange1.1t04443 | 3.97 | 3.98E-03 | TMV resistance protein N | 1.09E-39 |
| orange1.1t03802 | 3.96 | 3.80E-02 | Protein Enhanced disease susceptibility 1 | 3.56E-135 |
| Cs2g27410 | 3.86 | 4.63E-05 | MYB-related protein Myb4 | 8.93E-51 |
| orange1.1t04313 | 3.85 | 2.11E-05 | Huamn Ankyrin-2 | 1.79E-15 |
| Cs2g01090 | 3.81 | 1.23E-04 | UPF0481 protein | 8.09E-36 |
| Cs7g11940 | 3.77 | 1.13E-02 | Cytosolic sulfotransferase 12 | 4.84E-116 |
| Cs1g24440 | 3.62 | 2.76E-04 | Salicylate carboxymethyl transferase | 6.62E-158 |
| orange1.1t03083 | 3.33 | 3.98E-03 | Putative ribonuclease H protein | 3.42E-20 |
| Cs1g11960 | 3.31 | 1.15E-04 | G-type lectin S-receptor-like serine/threonine-protein kinase | 6.12-131 |
| Cs5g16310 | 3.17 | 4.40E-05 | Flavanone 3-dioxygenase | 2.30E-69 |
| orange1.1t02071 | 3.13 | 8.55E-05 | UPF0481 protein | 2.10E-28 |
| Cs2g21000 | 3.04 | 3.17E-04 | Linolenate hydroperoxide lyase, chloroplastic | 0 |
| Cs2g13280 | 3.03 | 8.73E-05 | Probable receptor-like protein kinase | 5.18E-98 |
| orange1.1t04913 | 3.02 | 2.44E-03 | TMV resistance protein N | 1.45E-54 |
| Cs7g29570 | 3.00 | 8.73E-05 | Probable WRKY transcription factor 70 | 1.61E-41 |
| Cs4g04210 | 2.84 | 2.92E-04 | Transcription factor bHLH35 | 4.47E-67 |
| orange1.1t01840 | 2.83 | 1.05E-02 | TMV resistance protein N | 8.85E-172 |
| Cs4g05900 | 2.72 | 1.06E-02 | Ankyrin repeat-containing protein | 3.72E-27 |
| Cs3g10110 | 2.62 | 2.54E-03 | LRR receptor-like serine/threonine-protein kinase | 9.01E-17 |
| orange1.1t02504 | 2.55 | 3.90E-03 | Probable disease resistance protein | 3.26E-175 |
| Cs4g18320 | 2.49 | 4.46E-03 | Glutaredoxin-C6 | 2.03E-57 |
| Cs9g12160 | 2.40 | 6.88E-03 | LRR receptor-like serine/threonine-protein kinase GSO1 | 3.67E-14 |
| Cs7g29040 | 2.34 | 2.61E-02 | Heat shock cognate 70 kDa protein | 0 |
| Cs7g15430 | 2.31 | 1.25E-02 | Cytochrome P450 89A2 | 3.06E-171 |
| Cs5g04250 | 2.25 | 2.55E-02 | Mitochondrial outer membrane protein porin of 36 kDa | 3.27E-51 |
| Cs6g01870 | 2.18 | 2.61E-02 | Probable LRR receptor-like serine/threonine-protein kinase | 6.63E-13 |
| orange1.1t00589 | 2.14 | 3.59E-02 | NAC domain-containing protein 68 | 1.50E-20 |
| Cs5g06000 | 2.13 | 3.10E-02 | Copper methylamine oxidase | 3.84E-167 |
| Cs1g03700 | 2.09 | 3.15E-02 | Mechanosensitive ion channel protein 10 | 1.20E-13 |
| Cs5g12280 | −6.39 | 6.99E-05 | Agamous-like MADS-box protein AGL8 | 3.38E-33 |
| Cs5g12290 | −5.76 | 1.70E-06 | Agamous-like MADS-box protein AGL8 | 9.58E-34 |
| Cs7g11800 | −3.65 | 8.18E-06 | Developmental protein SEPALLATA 2 | 8.38E-33 |
| Cs9g07970 | −2.33 | 8.25E-03 | 3-ketoacyl-CoA synthase 11 | 0 |
| orange1.1t04139 | −2.29 | 1.01E-02 | Geraniol 8-hydroxylase | 0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiu, W.; Soares, J.; Pang, Z.; Huang, Y.; Sun, Z.; Wang, N.; Grosser, J.; Dutt, M. Potential Mechanisms of AtNPR1 Mediated Resistance against Huanglongbing (HLB) in Citrus. Int. J. Mol. Sci. 2020, 21, 2009. https://doi.org/10.3390/ijms21062009
Qiu W, Soares J, Pang Z, Huang Y, Sun Z, Wang N, Grosser J, Dutt M. Potential Mechanisms of AtNPR1 Mediated Resistance against Huanglongbing (HLB) in Citrus. International Journal of Molecular Sciences. 2020; 21(6):2009. https://doi.org/10.3390/ijms21062009
Chicago/Turabian StyleQiu, Wenming, Juliana Soares, Zhiqian Pang, Yixiao Huang, Zhonghai Sun, Nian Wang, Jude Grosser, and Manjul Dutt. 2020. "Potential Mechanisms of AtNPR1 Mediated Resistance against Huanglongbing (HLB) in Citrus" International Journal of Molecular Sciences 21, no. 6: 2009. https://doi.org/10.3390/ijms21062009