Mutation of ACX1, a Jasmonic Acid Biosynthetic Enzyme, Leads to Petal Degeneration in Chinese Cabbage (Brassica campestris ssp. pekinensis)

Abstract
:1. Introduction
2. Results
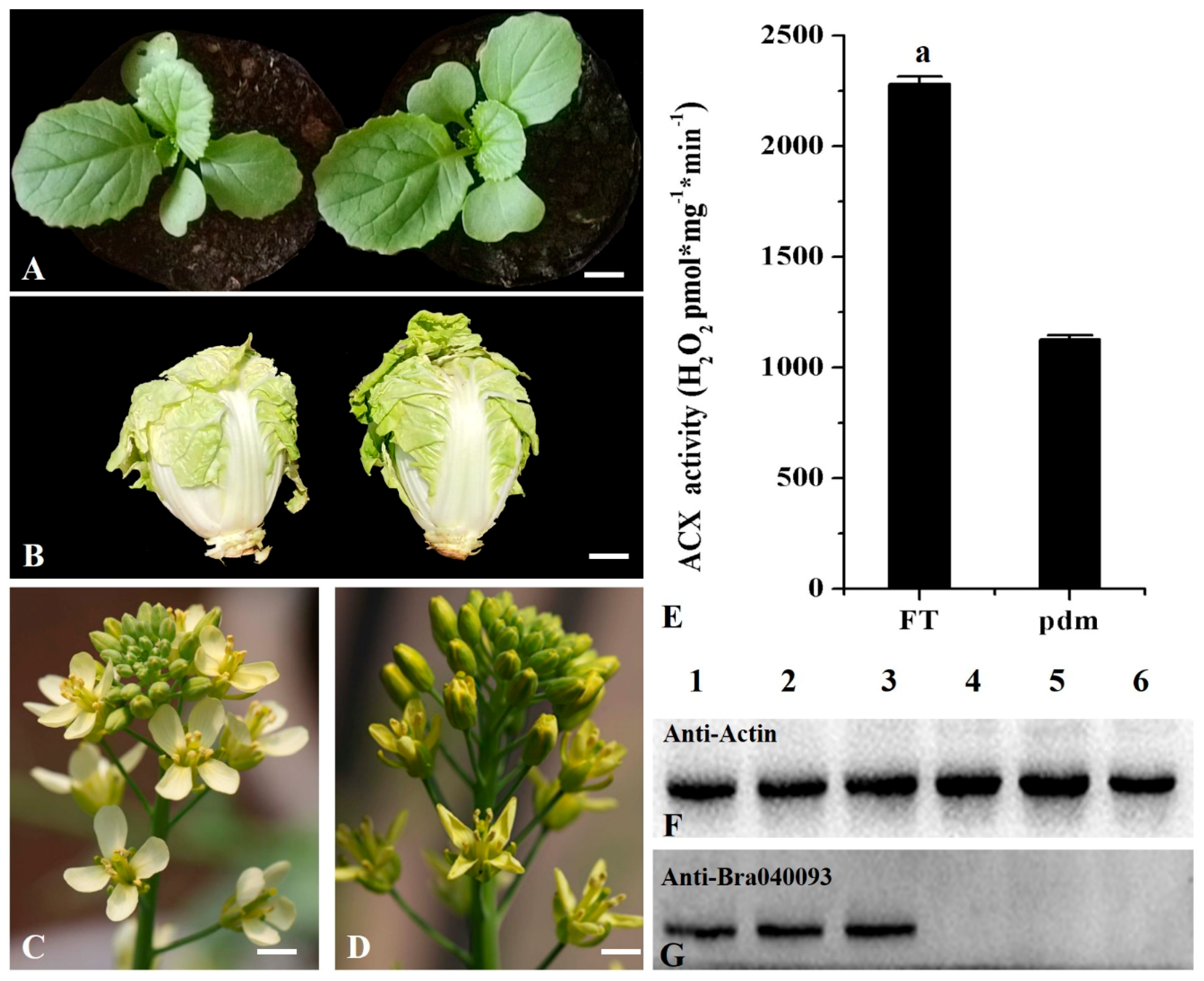
2.1. ACX Activity is Decreased in Pdm Plants
2.2. Analysis of Protein Expression Patterns
2.3. Analysis of JA and MeJ Contents
2.4. Effects of Exogenous Application of JA and Meja on Petal Phenotype
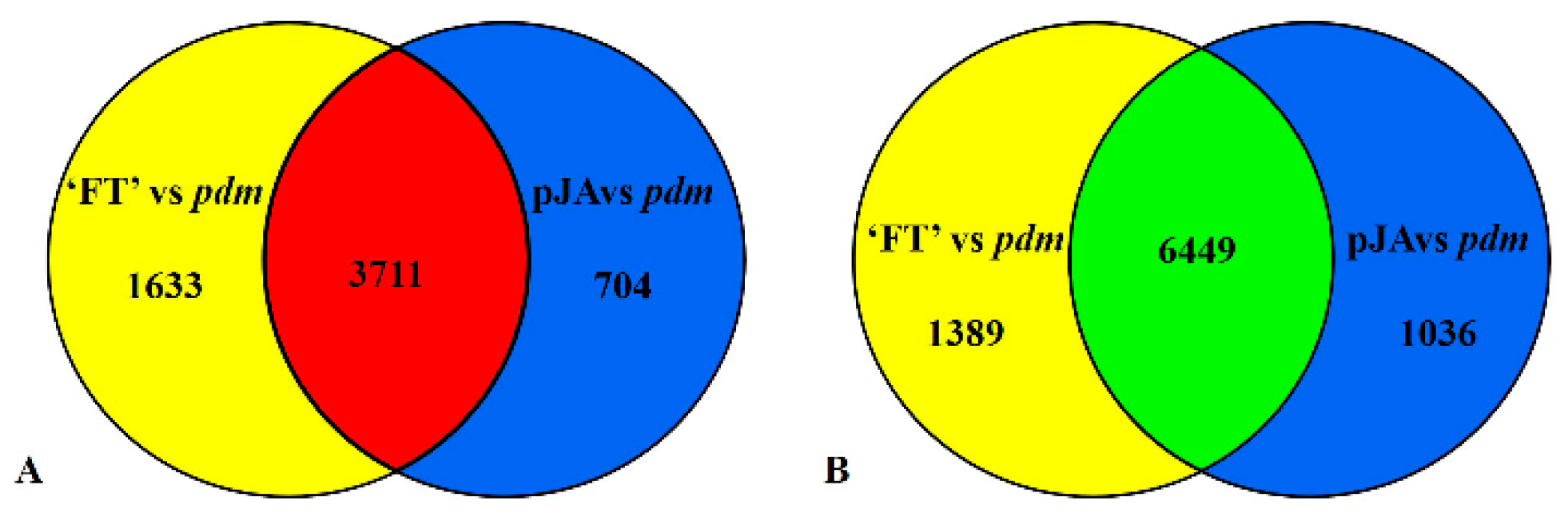
2.5. RNA-Seq Analysis
2.6. DEGs Related to Petal Development
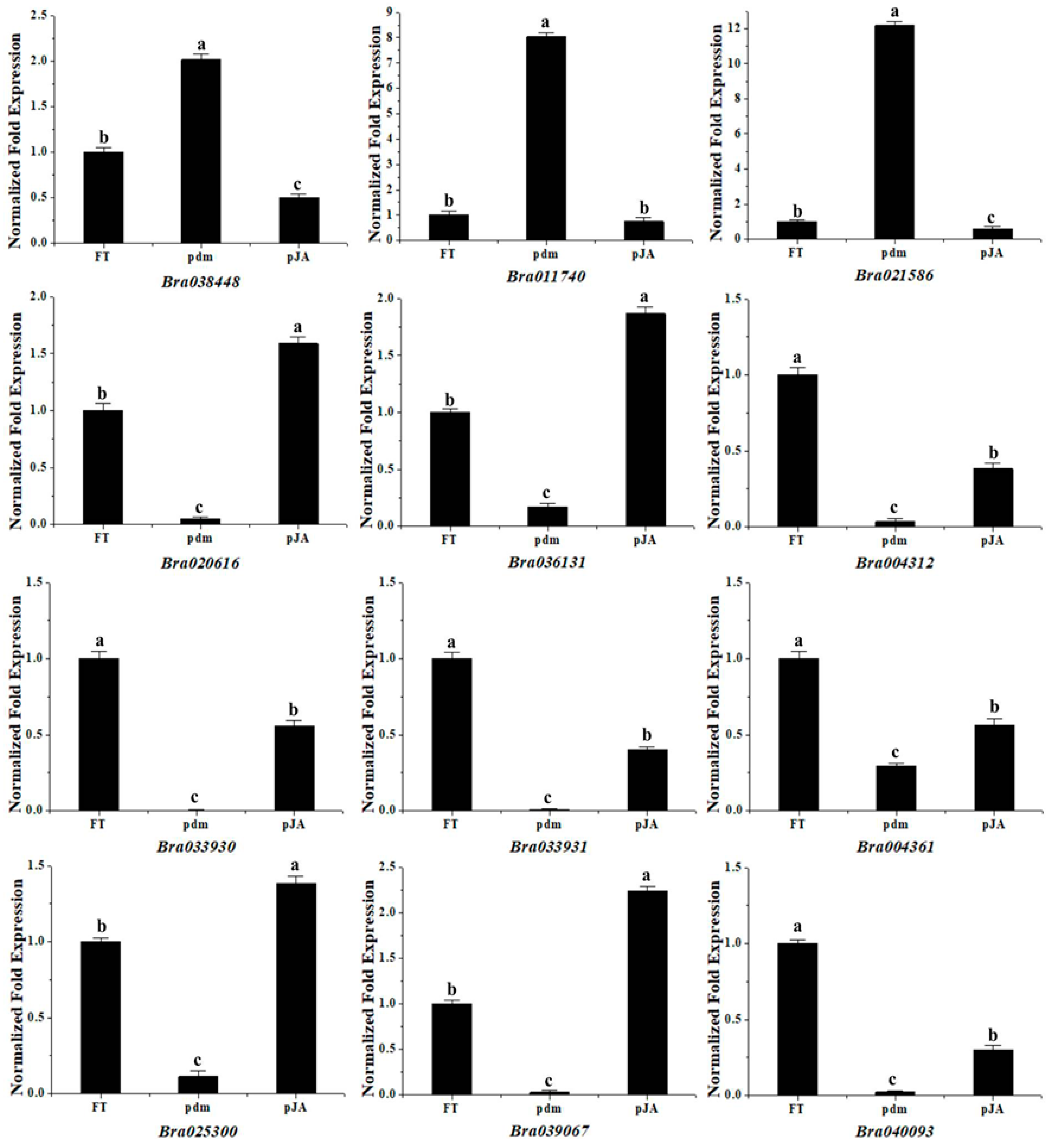
2.7. qRT-PCR Analysis of Gene Expression Patterns
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Measurement of ACX Activity
4.3. Western Blotting
4.4. Measurement of JA and MeJA Contents
4.5. Exogenous Application of JA and MeJA
4.6. Scanning Electron Microscopy (SEM)
4.7. RNA-Seq, Expression Annotation, GO, and KEGG Pathway Enrichment Analyses
4.8. RNA Isolation, cDNA Library Construction, and Illumina Sequencing for Transcriptome Analysis
4.9. Quantitative Real-Time PCR (qRT-PCR) Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Database
References
- Crepet, W.L. Progress in understanding angiosperm history, success, and relationships: Darwin’s abominably “perplexing phenomenon”. Proc. Natl. Acad. Sci. USA 2000, 97, 12939–12941. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhou, Y.; Ding, L.; Wu, Z.; Liu, R.; Meyerowitz, E.M. Transcription repressor HANABA TARANU controls flower development by integrating the actions of multiple hormones, floral organ specification genes, and GATA3 family genes in Arabidopsis. Plant Cell 2013, 25, 83–101. [Google Scholar] [CrossRef] [PubMed]
- Coen, E.S.; Meyerowitz, E.M. The war of the whorls: Genetic interactions controlling flower development. Nature 1991, 353, 31–37. [Google Scholar] [CrossRef]
- Colombo, L.; Franken, J.; Koetje, E.; van Went, J.; Dons, H.J.; Angenent, G.C.; van Tunen, A.J. The petunia MADS box gene FBP11 determines ovule identity. Plant Cell 1995, 7, 1859–1868. [Google Scholar] [CrossRef] [PubMed]
- Theissen, G. Development of floral organ identity: Stories from the MADS house. Curr. Opin. Plant Biol. 2001, 4, 75–85. [Google Scholar] [CrossRef]
- Theissen, G.; Saedler, H. Plant biology. Floral quartets. Nature 2011, 409, 469–471. [Google Scholar] [CrossRef]
- Galliot, C.; Stuurman, J.; Kuhlemeier, C. The genetic dissection of floral pollination syndromes. Curr. Opin. Plant Biol. 2006, 9, 78–82. [Google Scholar] [CrossRef]
- Mizukami, Y.; Fischer, R.L. Plant organ size control: AINTEGUMENTA regulates growth and cell numbers during organogenesis. Proc. Natl. Acad. Sci. USA 2000, 97, 942–947. [Google Scholar] [CrossRef]
- Anastasiou, E.; Kenz, S.; Gerstung, M.; MacLean, D.; Timmer, J.; Fleck, C.; Lenhard, M. Control of plant organ size by KLUH/CYP78A5-Dependent intercellular signaling. Dev. Cell 2007, 13, 843–856. [Google Scholar] [CrossRef] [PubMed]
- Stransfeld, L.; Eriksson, S.; Adamski, N.M.; Breuninger, H.; Lenhard, M. KLUH/CYP78A5 promotes organ growth without affecting the size of the early primordium. Plant Signal Behav. 2010, 5, 982–984. [Google Scholar]
- Kawade, K. Proliferative control of leaf cells through inter-cell-layer AN3 signaling. Plant Morphol. 2015, 26, 59–63. [Google Scholar] [CrossRef]
- Adamski, N.M.; Anastasiou, E.; Eriksson, S.; O’Neill, C.M.; Lenhard, M. Local maternal control of seed size by KLUH/CYP78A5-dependent growth signaling. Proc. Natl. Acad. Sci. USA 2009, 106, 20115–20120. [Google Scholar] [CrossRef]
- Brioudes, F.; Joly, C.; Szécsi, J.; Varaud, E.; Leroux, J.; Bellvert, F.; Bertrand, C.; Bendahmane, M. Jasmonate controls late development stages of petal growth in Arabidopsis thaliana. Plant J. 2009, 60, 1070–1080. [Google Scholar] [CrossRef]
- Szécsi, J.; Joly, C.; Bordji, K.; Varaud, E.; Cock, J.M.; Dumas, C.; Bendahmane, M. BIGPETALp, a bHLH transcription factor is involved in the control of Arabidopsis petal size. EMBO J. 2006, 25, 3912–3920. [Google Scholar] [CrossRef]
- Varaud, E.; Brioudes, F.; Szécsi, J.; Leroux, J.; Brown, S.; Perrot-Rechenmann, C.; Bendahmane, M. AUXIN RESPONSE FACTOR8 regulates Arabidopsis petal growth by interacting with the bHLH transcription factor BIGPETALp. Plant Cell 2011, 23, 973–983. [Google Scholar] [CrossRef]
- Szécsi, J.; Wippermann, B.; Bendahmane, M. Genetic and phenotypic analyses of petal development in Arabidopsis. Methods Mol. Biol. 2014, 1110, 191–202. [Google Scholar]
- Song, S.; Qi, T.; Wasternack, C.; Xie, D. Jasmonate signaling and crosstalk with gibberellin and ethylene. Curr. Opin. Plant Biol. 2014, 21, 112–119. [Google Scholar] [CrossRef]
- Yuan, Z.; Zhang, D. Roles of jasmonate signalling in plant inflorescence and flower development. Curr. Opin. Plant Biol. 2015, 27, 44–51. [Google Scholar] [CrossRef]
- Yoon, J.Y.; Hamayun, M.; Lee, S.K.; Lee, I.J. Methyl jasmonate alleviated salinity stress in soybean. J. Crop Sci. Biotechnol. 2009, 12, 63–68. [Google Scholar] [CrossRef]
- Wasternack, C.; Stenzel, I.; Hause, B.; Hause, G.; Kutter, C.; Maucher, H.; Neumerkel, J.; Feussner, I.; Miersch, O. The wound response in tomato-role of jasmonic acid. J. Plant Physiol. 2006, 163, 297–306. [Google Scholar] [CrossRef]
- Fugate, K.K.; Oliveira, L.S.D.; Ferrareze, J.P.; Bolton, M.D.; Deckard, E.L.; Finger, F.L. Jasmonic acid causes short and long term alterations to the transcriptome and the expression of defense genes in sugarbeet roots. Plant Gene 2017, 9, 50–63. [Google Scholar] [CrossRef]
- He, Y.; Zhang, H.; Sun, Z.; Li, J.; Hong, G.; Zhu, Q.; Zhou, X.; MacFarlane, S.; Yan, F.; Chen, J. Jasmonic acid-mediated defense suppresses brassinosteroid-mediated susceptibility to Rice black streaked dwarf virus infection in rice. New Phytol. 2017, 214, 388–399. [Google Scholar] [CrossRef]
- Wasternack, C.; Hause, B. Jasmonates: Biosynthesis, perception, signal transduction and action in plant stress response, growth and development. an update to the 2007 review in Annals of Botany. Ann. Bot. 2013, 111, 1021–1058. [Google Scholar] [CrossRef]
- Monzón, G.C.; Pinedo, M.; Lamattina, L.; Canal, L.D.L. Sunflower root growth regulation: The role of jasmonic acid and its relation with auxins. Plant Growth Regul. 2012, 66, 129–136. [Google Scholar] [CrossRef]
- Stintzi, A.; Browse, J. The Arabidopsis male-sterile mutant, opr3, lacks the 12-oxophytodienoic acid reductase required for jasmonate synthesis. Proc. Natl. Acad. Sci. USA 2000, 97, 10625–10630. [Google Scholar] [CrossRef]
- Li, L.; Zhao, Y.; McCaig, B.C.; Wingerd, B.A.; Wang, J.; Whalon, M.E.; Pichersky, E.; Howe, G.A. The tomato homolog of coronatine-insensitive1 is required for the maternal control of seed maturation, jasmonate-signaled defense responses, and glandular trichome development. Plant Cell 2004, 16, 126–143. [Google Scholar] [CrossRef]
- Ishiguro, S.; Kawai-Oda, A.; Ueda, J.; Nishida, I.; Okada, K. The DEFECTIVE IN ANTHER DEHISCEBCE1 gene encodes a novel phospholipase A1 catalyzing the initial step of jasmonic acid biosynthesis, which synchronizes pollen maturation, anther dehiscence, and flower opening in Arabidopsis. Plant Cell 2001, 13, 2191–2209. [Google Scholar] [CrossRef]
- Reeves, P.H.; Ellis, C.M.; Ploense, S.E.; Wu, M.F.; Yadav, V.; Tholl, D.; Chételat, A.; Haupt, I.; Kennerley, B.J.; Hodgens, C.; et al. A regulatory network for coordinated flower maturation. PLoS Genet. 2012, 8, e1002506. [Google Scholar] [CrossRef]
- Ueda, J.; Kato, J. Isolation and identification of a senescence-promoting substance from wormwood (Artemisia absinthium L.). Plant Physiol. 1980, 66, 246–249. [Google Scholar] [CrossRef]
- Liu, L.; Li, H.X.; Zeng, H.L.; Cai, Q.H.; Zhou, X.; Yin, C.X. Exogenous jasmonic acid and cytokinin antagonistically regulate rice flag leaf senescence by mediating chlorophyll degradation, membrane deterioration, and senescence-associated genes expression. Plant Growth Regul. 2016, 35, 366–376. [Google Scholar] [CrossRef]
- Li, H.; Xue, D.; Gao, Z.; Yan, M.; Xu, W.; Xing, Z.; Huang, D.; Qian, Q.; Xue, Y. A putative lipase gene EXTRA GLUME1 regulates both empty-glume fate and spikelet development in rice. Plant J. 2009, 57, 593–605. [Google Scholar] [CrossRef]
- Cai, Q.; Yuan, Z.; Chen, M.; Yin, C.; Luo, Z.; Zhao, X.; Liang, W.; Hu, J.; Zhang, D. Jasmonic acid regulates spikelet development in rice. Nat. Commun. 2014, 5, 3476. [Google Scholar] [CrossRef] [Green Version]
- Pak, H.; Guo, Y.; Chen, M.; Chen, K.; Li, Y.; Hua, S.; Shamsi, I.; Meng, H.; Shi, C.; Jiang, L. The effect of exogenous methyl jasmonate on the flowering time, floral organ morphology, and transcript levels of a group of genes implicated in the development of oilseed rape flowers (Brassica napus L.). Planta 2009, 231, 79–91. [Google Scholar] [CrossRef]
- Mandaokar, A.; Thines, B.; Shin, B.; Lange, B.M.; Choi, G.; Koo, Y.J.; Yoo, Y.J.; Choi, Y.D.; Choi, G.; Browse, J. Transcriptional regulators of stamen development in Arabidopsis identified by transcriptional profiling. Plant J. 2006, 46, 984–1008. [Google Scholar] [CrossRef] [Green Version]
- Cheng, H.; Song, S.; Xiao, L.; Soo, H.M.; Cheng, Z.; Xie, D.; Peng, J. Gibberellin acts through jasmonate to control the expression of MYB21, MYB24, and MYB57 to promote stamen filament growth in Arabidopsis. PLoS Genet. 2009, 5, e1000440. [Google Scholar] [CrossRef]
- Song, S.; Qi, T.; Huang, H.; Ren, Q.; Wu, D.; Chang, C.; Peng, W.; Liu, Y.; Peng, J.; Xie, D. The Jasmonate-ZIM Domain Proteins Interact with the R2R3-MYB Transcription Factors MYB21 and MYB24 to Affect Jasmonate-Regulated Stamen Development in Arabidopsis. Plant Cell 2011, 23, 1000–1013. [Google Scholar] [CrossRef]
- León, J. Role of plant peroxisomes in the production of jasmonic acid-based signals. Subcell. Biochem. 2013, 69, 299–313. [Google Scholar]
- Li, C.; Schilmiller, A.L.; Liu, G.; Lee, G.I.; Howe, G.A. Role of -oxidation in jasmonate biosynthesis and systemic wound signaling in tomato. Plant Cell 2005, 17, 971–986. [Google Scholar] [CrossRef]
- Schilmiller, A.L.; Howe, K.G.A. Functional diversification of acyl-coenzyme a oxidases in jasmonic acid biosynthesis and action. Plant Physiol. 2007, 143, 812–824. [Google Scholar] [CrossRef]
- Agarwal, A.K.; Qi, Y.; Bhat, D.G.; Woerner, B.M.; Brown, S.M. Gene isolation and characterization of two acyl coa oxidases from soybean with broad substrate specificities and enhanced expression in the growing seedling axis. Plant Mol. Biol. 2001, 47, 519–531. [Google Scholar] [CrossRef]
- Xin, Z.J.; Chen, S.L.; Ge, L.G.; Li, X.W.; Sun, X.L. The involvement of a herbivore-induced acyl-CoA oxidase gene, CsACX1, in the synthesis of jasmonic acid and its expression in flower opening in tea plant (Camellia sinensis). Plant Physiol. Biochem. 2019, 135, 132–140. [Google Scholar] [CrossRef]
- Huang, S.N.; Liu, Z.Y.; Li, D.Y.; Yao, R.P.; Meng, Q.; Feng, H. Screening of Chinese cabbage mutants produced by 60Coγ-ray mutagenesis of isolated microspore cultures. Plant Breed. 2014, 133, 480–488. [Google Scholar] [CrossRef]
- Huang, S.N.; Liu, Z.Y.; Yao, R.P.; Li, D.Y.; Zhang, T.; Li, X.; Hou, L.; Wang, Y.H.; Tang, X.Y.; Feng, H. Candidate gene prediction for a petal degeneration mutant, pdm, of the Chinese cabbage (Brassica campestris, ssp. pekinensis) by using fine mapping and transcriptome analysis. Mol. Breed. 2016, 36, 26. [Google Scholar] [CrossRef]
- Huang, S.N.; Liu, Z.Y.; Yao, R.P.; Li, D.Y.; Feng, H. Comparative transcriptome analysis of the petal degeneration mutant pdm, in Chinese cabbage (Brassica campestris, ssp. pekinensis) using RNA-seq. Mol. Genet. Genom. 2015, 290, 1833–1847. [Google Scholar] [CrossRef]
- Horiguchi, G.; Ferjani, A.; Fujikura, U.; Tsukaya, H. Coordination of cell proliferation and cell expansion in the control of leaf size in Arabidopsis thaliana. J. Plant Res. 2006, 119, 37–42. [Google Scholar] [CrossRef]
- Lazarow, P.B.; De Duve, C. A fatty acyl-CoA oxidizing system in rat liver peroxisomes; enhancement by clofibrate, a hypolipidemic drug. Proc. Natl. Acad. Sci. USA 1976, 73, 2043–2046. [Google Scholar] [CrossRef]
- Kondo, S.; Setha, S.; Rudell, D.R.; Buchanan, D.A.; Mattheis, J.P. Aroma volatile biosynthesis in apples affected by 1-MCP and methyl jasmonate. Postharvest Biol. Technol. 2005, 36, 61–68. [Google Scholar] [CrossRef]
- Powers, R.A.; Rife, C.L.; Schilmiller, A.L.; Howe, G.A.; Garavito, R.M. Structure determination and analysis of acyl-coa oxidase (ACX1) from tomato. Acta Crystallogr. 2006, 62, 683–686. [Google Scholar] [CrossRef]
- Wasternack, C. Jasmonates: An update on biosynthesis, signal transduction and action in plant stress response, growth and development. Ann. Bot. 2007, 100, 681–697. [Google Scholar] [CrossRef]
- Koo, A.J.; Howe, G.A. Catabolism and deactivation of the lipid-derived hormone jasmonoyl-isoleucine. Front. Plant Sci. 2012, 3, 19. [Google Scholar] [CrossRef]
- Wiszniewski, A.A.; Bussell, J.D.; Long, R.L.; Smith, S.M. Knockout of the two evolutionarily conserved peroxisomal 3-ketoa cyl-CoA thiolases in Arabidopsis recapitulates the abnormal inflorescence meristem 1 phenotype. J. Exp. Bot. 2014, 65, 6723–6733. [Google Scholar] [CrossRef]
- Hooks, M.A.; Kellas, F.; Graham, I.A. Long-chain acyl-CoA oxidases of Arabidopsis. Plant J. 1999, 20, 1–13. [Google Scholar] [CrossRef]
- Gustafson-Brown, C.; Savidge, B.; Yanofsky, M.F. Regulation of the Arabidopsis floral homeotic gene APETALA1. Cell 1994, 76, 131–143. [Google Scholar] [CrossRef]
- Basu, D.; Le, J.; Zakharova, T.; Mallery, E.L.; Szymanski, D.B. A SPIKE1 signaling complex controls actin-dependent cell morphogenesis through the heteromeric WAVE and APR2/3 complexes. Proc. Natl. Acad. Sci. USA 2008, 105, 4044–4049. [Google Scholar] [CrossRef]
- Qiu, J.L.; Jilk, R.; Marks, M.D.; Szymanski, D.B. The Arabidopsis SPIKE1 gene is required for normal cell shape control and tissue development. Plant Cell 2002, 14, 101–118. [Google Scholar] [CrossRef]
- Ren, H.; Dang, X.; Yang, Y.; Huang, D.; Liu, M.; Gao, X.; Lin, D. SPIKE1 Activates ROP GTPase to Modulate Petal Growth and Shape. Plant Physiol. 2016, 172, 358–371. [Google Scholar] [CrossRef]
- Groszmann, M.; Bylstra, Y.; Lampugnani, E.R.; Smyth, D.R. Regulation of tissue-specific expression of SPATULA, a bHLH gene involved in carpel development, seedling germination, and lateral organ growth in Arabidopsis. J. Exp. Bot. 2010, 61, 1495–1508. [Google Scholar] [CrossRef]
- Heisler, M.G.; Atkinson, A.; Bylstra, Y.H.; Walsh, R.; Smyth, D.R. SPATULA, a gene that controls development of carpel margin tissues in Arabidopsis, encodes a bHLH protein. Development 2001, 128, 1089–1098. [Google Scholar]
- Penfield, S.; Josse, E.M.; Kannangara, R.; Gilday, A.D.; Halliday, K.J.; Graham, I.A. Cold and Light Control Seed Germination through the bHLH Transcription Factor SPATULA. Curr. Biol. 2005, 15, 1998–2006. [Google Scholar] [CrossRef] [Green Version]
- Nag, A.; King, S.; Jack, T. miR319a targeting of TCP4 is critical for petal growth and development in Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 22534–22539. [Google Scholar] [CrossRef]
- Lee, B.H.; Ko, J.H.; Lee, S.; Lee, Y.; Pak, J.H.; Kim, J.H. The Arabidopsis GRF-INTERACTING FACTOR gene family performs an overlapping function in determining organ size as well as multiple developmental properties. Plant Physiol. 2009, 151, 655–668. [Google Scholar] [CrossRef]
- Kim, J.H.; Kende, H. A transcriptional coactivator, AtGIF1, is involved in regulating leaf growth and morphology in Arabidopsis. Proc. Natl. Acad. Sci. USA 2004, 101, 13374–13379. [Google Scholar] [CrossRef]
- Dinneny, J.R.; Yadegari, R.; Fischer, R.L.; Yanofsky, M.F.; Weigel, D. The role of JAGGED in shaping lateral organs. Development 2004, 131, 1101–1110. [Google Scholar] [CrossRef]
- Schiessl, K.; Muiño, J.M.; Sablowshi, R. Arabidopsis JAGGED links floral organ patterning to tissue growth by repressing Kip-related cell cycle. Proc. Natl. Acad. Sci. USA 2014, 111, 2830–2835. [Google Scholar] [CrossRef]
- Adham, A.R.; Zolman, B.K.; Millius, A.; Bartel, B. Mutations in Arabidopsis acyl-CoA oxidase genes reveal distinct and overlapping roles in beta-oxidation. Plant J. 2005, 41, 859–874. [Google Scholar] [CrossRef]
- Liu, W.L.; Jia, H.J.; Zhang, X. Determination of acyl Coenzyme a Oxidase activity in Peach Fruit. Zhejiang Agric. Sci. 2010, 1, 1072–1075. [Google Scholar]
- You, C.C.; Zhu, H.L.; Xu, B.B.; Huang, W.X.; Wang, S.H.; Ding, Y.F.; Liu, Z.H.; Li, G.H.; Lin, C.; Ding, C.Q.; et al. Effect of removing superior spikelets on grain filling of inferior spikelets in rice. Front. Plant Sci. 2016, 7, 1161. [Google Scholar] [CrossRef]
- Liu, T.; Xu, J.; Li, J.; Hu, X. No is involved in JA- and H2O2-mediated ala-induced oxidative stress tolerance at low temperatures in tomato. Environ. Exp. Bot. 2018. [Google Scholar] [CrossRef]
- Krizek, B.A.; Anderson, J.T. Control of flower size. J. Exp. Bot. 2013, 64, 1427–1437. [Google Scholar] [CrossRef] [Green Version]
- Pelaz, S.; Ditta, G.S.; Baumann, E.; Wisman, E.; Yanofsky, M.F. B and C floral organ identity functions require sepallata MADS-box genes. Nature 2000, 405, 200–203. [Google Scholar] [CrossRef]
- Hu, Y.; Poh, H.M.; Chua, N.H. The Arabidopsis ARGOS-LIKE gene regulates cell expansion during organ growth. Plant J. 2006, 47, 1–9. [Google Scholar] [CrossRef]
- Fujikura, U.; Horiguchi, G.; Ponce, M.R.; Micol, J.L.; Tsukaya, H. Coordination of cell proliferation and cell expansion mediated by ribosome-related processes in the leaves of Arabidopsis thaliana. Plant J. 2009, 59, 499–508. [Google Scholar] [CrossRef]
- Feng, G.; Qin, Z.; Yan, J.; Zhang, X.; Hu, Y. Arabidopsis ORGAN SIZE RELATED1 regulates organ growth and final organ size in orchestration with ARGOS and ARL. New Phytol. 2011, 191, 635–646. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Materials | JA (ng/g) | MeJA (ng/g) |
|---|---|---|
| ‘FT’ | 43.07 ± 1.92 * | 1.4 ± 0.06 * |
| pdm | 1.76 ± 0.06 | 0.28 ± 0.02 |
| Gene ID | log2 Fold Change | Up/Down Regulation | Gene Annotation (BLASTX to Arabidopsis thaliana) |
|---|---|---|---|
| Bra038448 | 1.63 | Up | Guanine nucleotide exchange factor SPIKE 1 (SPK1) |
| Bra011740 | 3.93 | Up | Transcription factor SPATULA (SPT) |
| Bra021586 | 4.47 | Up | Transcription factor TCP4 (TCP4) |
| Bra020616 | −2.43 | Down | GRF1-interacting factor 1 (GIF1) |
| Bra036131 | −3.20 | Down | GRF1-interacting factor 1 (GIF1) |
| Bra004312 | −3.01 | Down | Zinc finger protein JAGGED (JAG) |
| Bra033931 | −4.65 | Down | Zinc finger protein JAGGED (JAG) |
| Bra033930 | −4.39 | Down | Zinc finger protein JAGGED (JAG) |
| Bra004361 | −1.10 | Down | Floral homeotic protein APETALA 1 (AP1) |
| Bra025300 | −2.38 | Down | Transcription factor MYB21 (MYB21) |
| Bra039067 | −4.21 | Down | Transcription factor MYB21 (MYB21) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, S.; Huang, S.; Liu, Z.; Feng, H. Mutation of ACX1, a Jasmonic Acid Biosynthetic Enzyme, Leads to Petal Degeneration in Chinese Cabbage (Brassica campestris ssp. pekinensis). Int. J. Mol. Sci. 2019, 20, 2310. https://doi.org/10.3390/ijms20092310
Peng S, Huang S, Liu Z, Feng H. Mutation of ACX1, a Jasmonic Acid Biosynthetic Enzyme, Leads to Petal Degeneration in Chinese Cabbage (Brassica campestris ssp. pekinensis). International Journal of Molecular Sciences. 2019; 20(9):2310. https://doi.org/10.3390/ijms20092310
Chicago/Turabian StylePeng, Shenling, Shengnan Huang, Zhiyong Liu, and Hui Feng. 2019. "Mutation of ACX1, a Jasmonic Acid Biosynthetic Enzyme, Leads to Petal Degeneration in Chinese Cabbage (Brassica campestris ssp. pekinensis)" International Journal of Molecular Sciences 20, no. 9: 2310. https://doi.org/10.3390/ijms20092310