
Structural Characterization and Evolutionary Relationship of High-Molecular-Weight Glutenin Subunit Genes in Roegneria nakaii and Roegneria alashanica


Abstract
:1. Introduction
2. Results
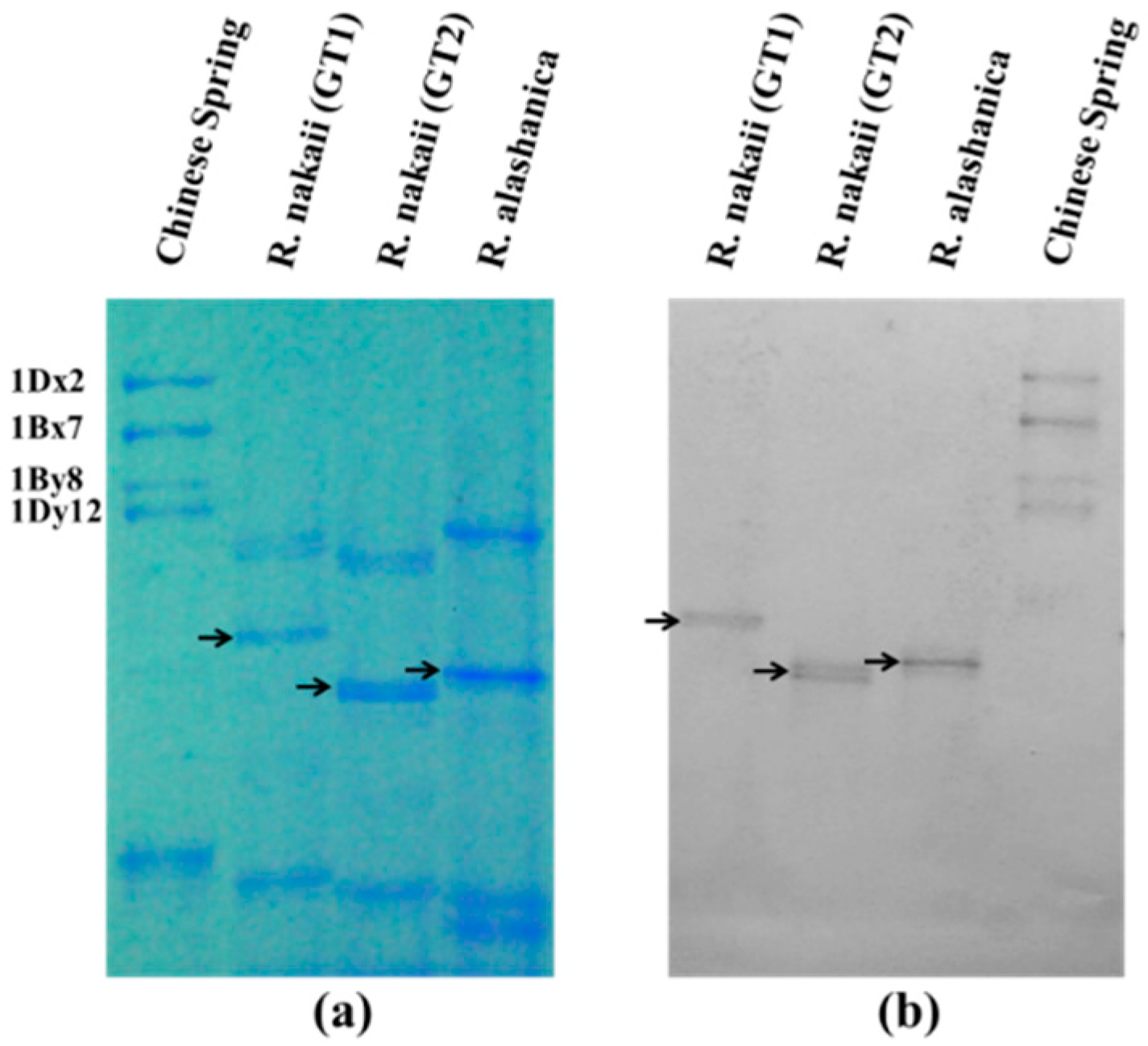
2.1. The Composition of HMW-GSs in R. nakaii and R. alashanica
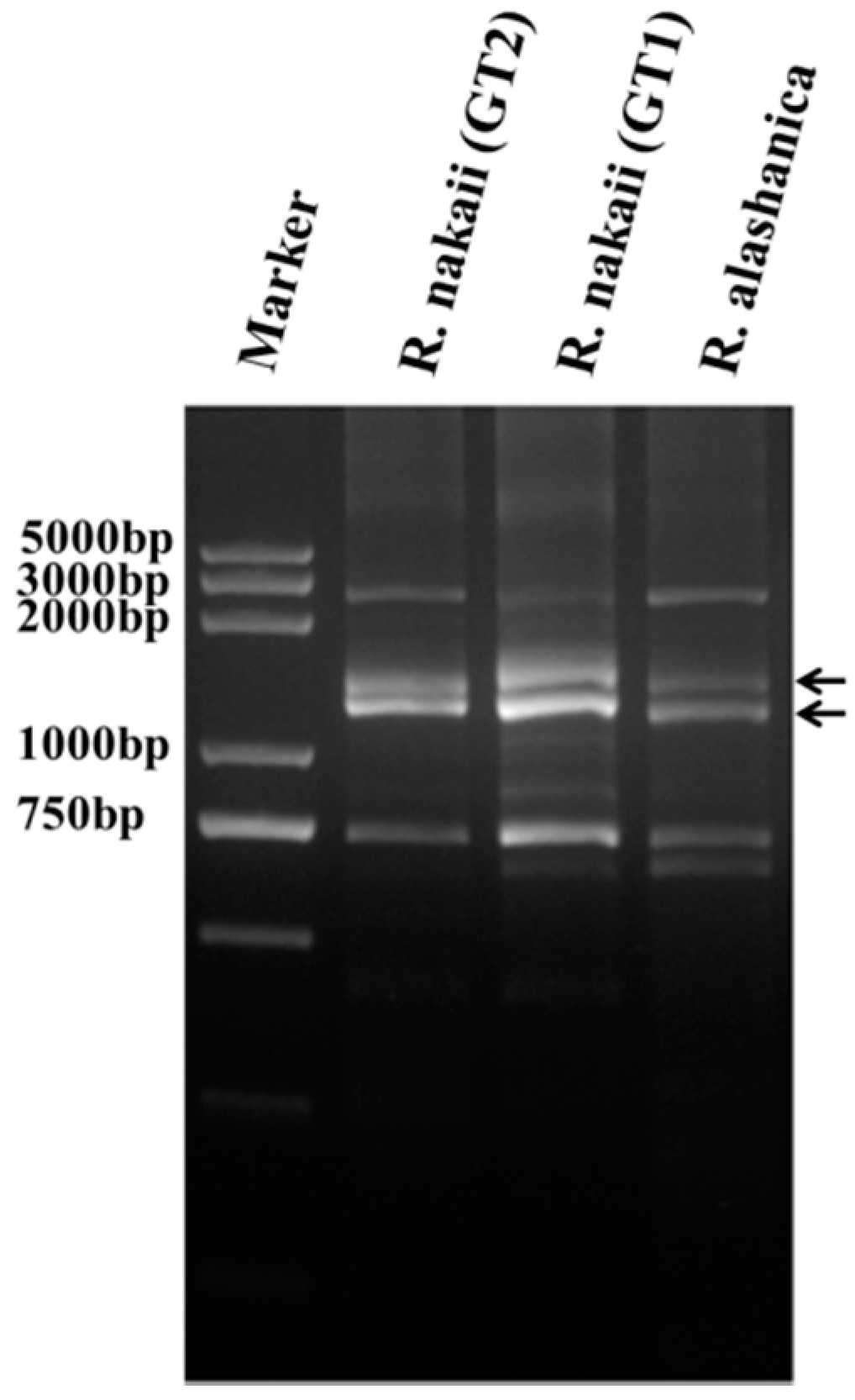
2.2. Cloning and Sequence Analyzing of HMW-GS Genes from R. nakaii and R. alashanica
2.3. Characterization and Comparative Analysis of the Roegneria HMW-GSs
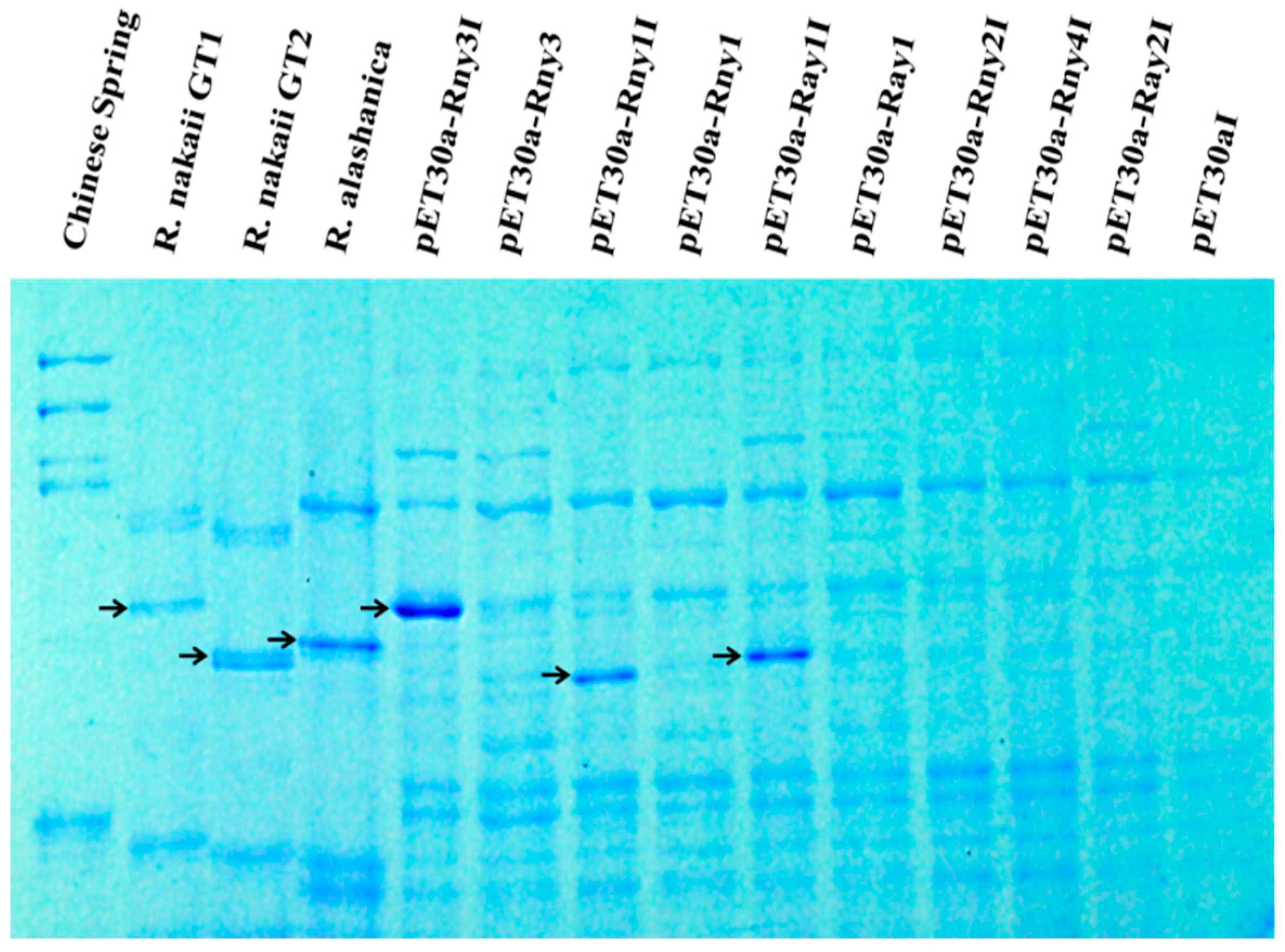
2.4. Heterologous Expression of the Roegneria HMW-GS Genes in Escherichia coli
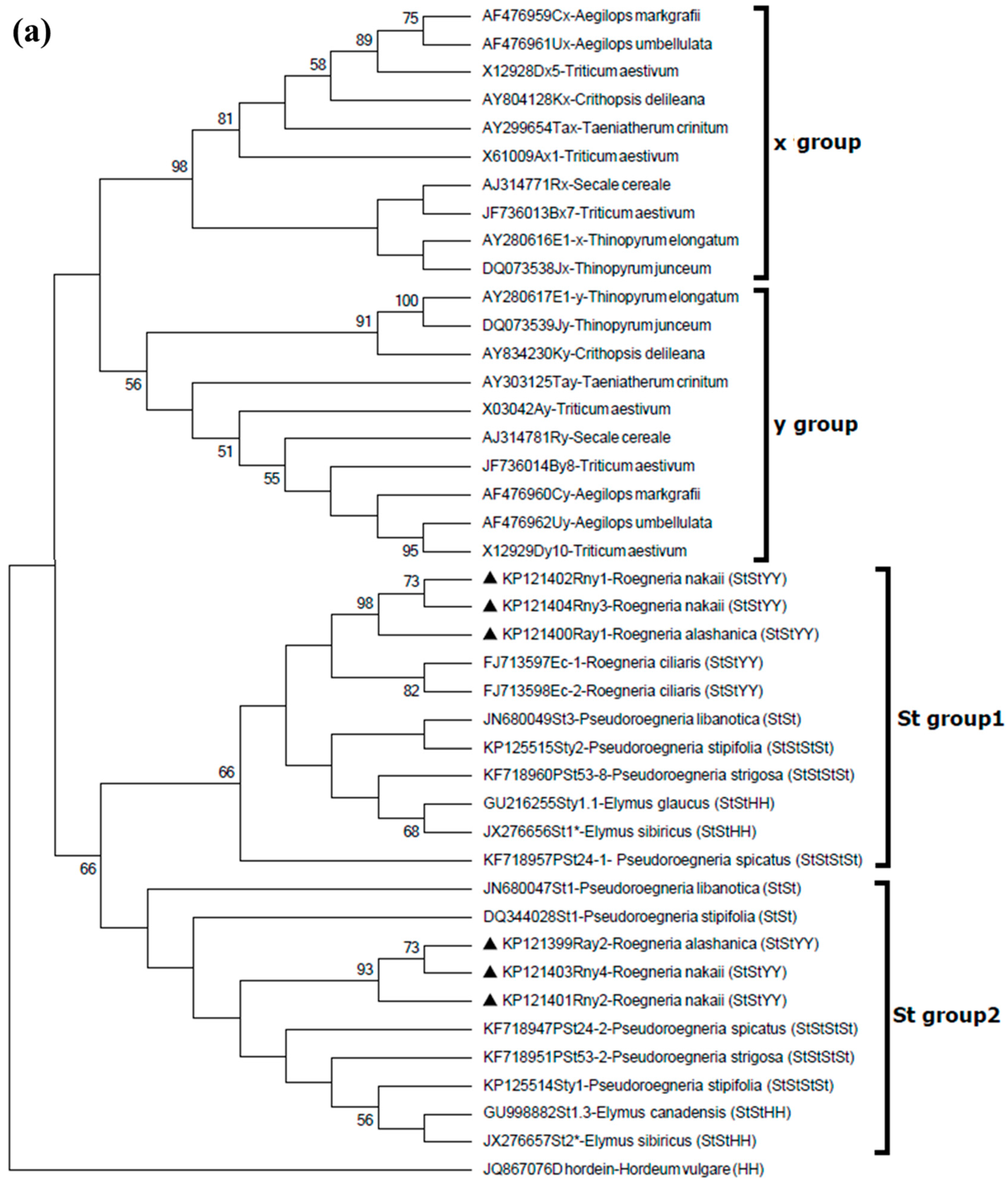
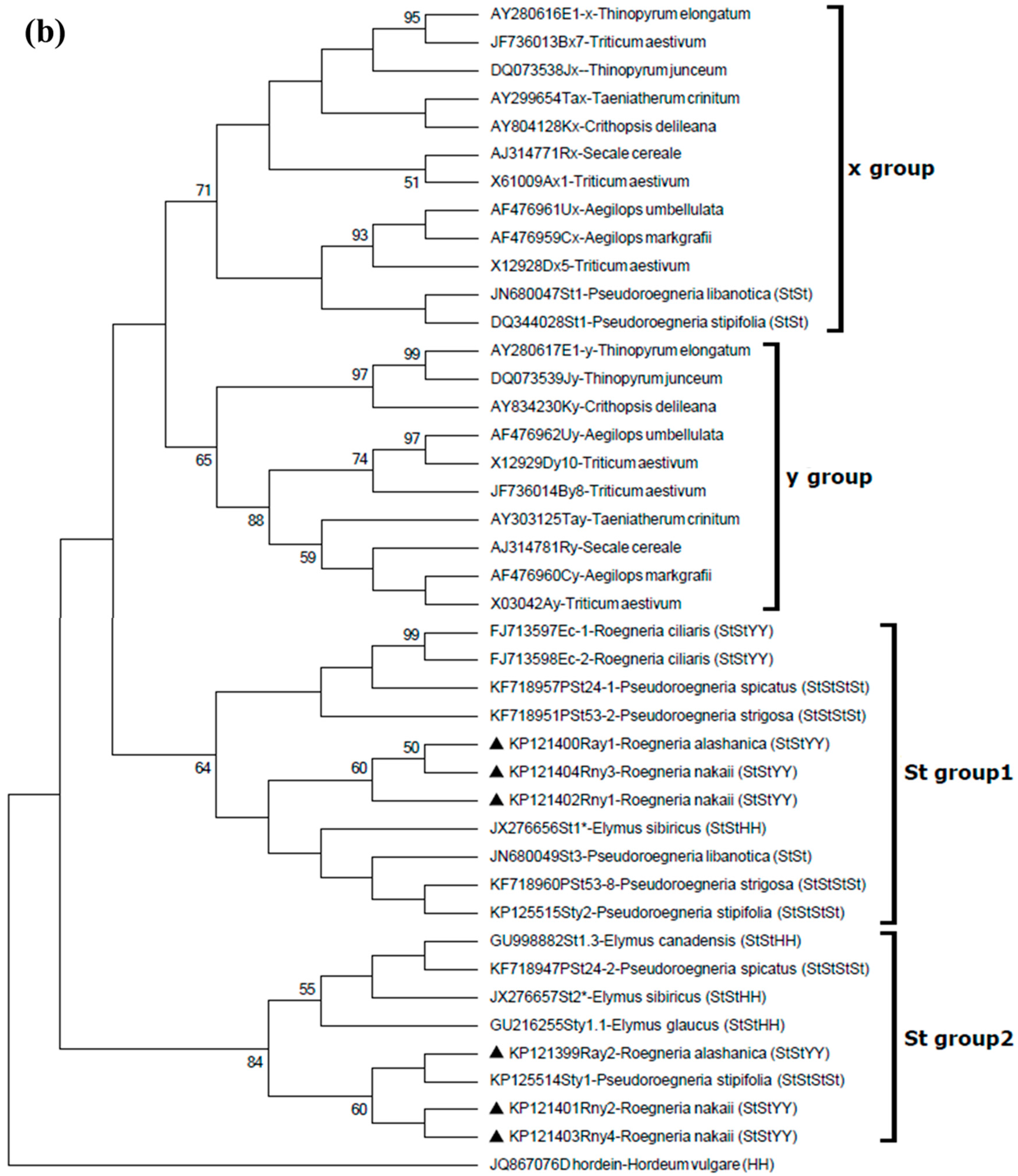
2.5. Phylogenetic Relationships of the New Roegneria HMW-GSs with Orthologous Subunits from Other Species
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. SDS-PAGE
4.3. Western Blotting
4.4. Cloning and Sequencing of the Roegneria HMW-GS Genes
4.5. Heterologous Expression of Cloned HMW-GS Genes in E. coli
4.6. Sequence Comparison and Phylogenetic Analyses of Cloned HMW-GS Genes
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Payne, P.I.; Seekings, J.A.; Worland, A.J.; Jarvis, M.G.; Holt, L.M. Allelic variation of glutenin subunits and gliadins and its effect on breadmaking quality in wheat: Analysis of F5 progeny from Chinese Spring × Chinese Spring (Hope 1A). J. Cereal Sci. 1987, 6, 103–118. [Google Scholar] [CrossRef]
- Shewry, P.R.; Halford, N.G. Cereal seed storage proteins structures properties and role in grain utilization. J. Exp. Bot. 2002, 53, 947–958. [Google Scholar] [CrossRef] [PubMed]
- Payne, P.I.; Holt, L.M.; Worland, A.J.; Law, C.N. Structural and genetical studies on the high-molecular-weight subunits of wheat glutenin: Part 3. Telocentric mapping of the subunit genes on the long arms of the homoeologous group 1 chromosomes. Theor. Appl. Genet. 1982, 63, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Payne, P.I. Genetics of wheat storage proteins and the effect of allelic variation on bread-making quality. Annu. Rev. Plant Physiol. 1987, 38, 141–153. [Google Scholar] [CrossRef]
- Payne, P.I.; Corfield, K.G.; Holt, L.M.; Blackman, J.A. Correlations between the inheritance of certain high-molecular weight subunits of glutenin and bread-making quality in progenies of six crosses of bread wheat. J. Sci. Food Agric. 1981, 32, 51–60. [Google Scholar] [CrossRef]
- Shewry, P.R.; Lafiandra, D.; Tamas, L.; Bekes, F. Genetic manipulation of gluten structure and function. In Gliadin and Glutenin: The Unique Balance of Wheat Quality; Wrigley, C., Bekes, F., Eds.; AACC International, Inc.: St. Paul, MN, USA, 2006; pp. 363–385. [Google Scholar]
- Halford, N.G.; Forde, J.; Anderson, O.D.; Greene, F.C.; Shewry, P.R. The nucleotide and deduced amino acid sequences of an HMW glutenin subunit gene from chromosome 1B of bread wheat (Triticum aestivum L.) and comparison with those of genes from chromosomes 1A and 1D. Theor. Appl. Genet. 1987, 75, 117–126. [Google Scholar] [CrossRef]
- Anderson, O.D.; Greene, F.C.; Yip, R.E.; Halford, N.G.; Shewry, P.R.; Malpica-Romero, J.M. Nucleotide sequences of the two high weight glutenin genes from the D-genome of a hexaploid bread wheat, Triticum aestivum L. cv Cheyenne. Nucleic Acids Res. 1989, 17, 461–462. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wan, Y.; Liu, Z.; Liu, K.; Liu, X.; Li, B.; Li, Z.; Zhang, X.; Dong, Y.; Wang, D. Molecular characterization of HMW glutenin subunit allele 1Bx14: Further insights into the evolution of Glu-B1-1 in wheat and related species. Theor. Appl. Genet. 2004, 109, 1093–1104. [Google Scholar] [CrossRef] [PubMed]
- Pang, B.S.; Zhang, X.Y. Isolation and molecular characterization of high molecular weight glutenin subunit genes 1Bx13 and 1By16 from hexaploid wheat. J. Integr. Plant Biol. 2008, 50, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Shao, H.; Liu, T.; Ran, C.F.; Li, L.Q.; Yu, J.; Gao, X.; Li, X.J. Isolation and molecular characterization of two novel HMW-GS genes from Chinese wheat (Triticum aestivum L.) landrace Banjiemang. Genes Genom. 2015, 37, 45–53. [Google Scholar] [CrossRef]
- Bustos, A.D.; Jouve, N. Characterisation and analysis of new HMW-glutenin alleles encoded by the Glu-R1 locus of Secale cereale. Theor. Appl. Genet. 2003, 107, 74–83. [Google Scholar] [PubMed]
- Guo, Z.F.; Yan, Z.H.; Wang, J.R.; Wei, Y.M.; Zheng, Y.L. Characterization of HMW prolamines and their coding sequences from Crithopsis delileana. Hereditas 2005, 142, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.X.; Zhang, X.Q.; Zhang, H.G.; Cao, S.H.; Wang, D.W.; Hao, S.T.; Li, L.H.; Li, H.J.; Wang, X.P. Isolation and characterization of a novel variant of HMW glutenin subunit gene from the St genome of Pseudoroegneria stipifolia. J. Cereal Sci. 2008, 47, 429–437. [Google Scholar] [CrossRef]
- Liu, S.W.; Gao, X.; Xia, G.M. Characterizing HMW-GS alleles of decaploid Agropyron elongatum in relation to evolution and wheat breeding. Theor. Appl. Genet. 2008, 116, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.T.; Wei, Y.M.; Lu, Z.X.; Pu, Z.E.; Lan, X.J.; Zheng, Y.L. Structural variation and evolutionary relationship of novel HMW glutenin subunits from Elymus glaucus. Hereditas 2010, 147, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.W.; Zhao, F.; Gao, X.; Chen, F.G.; Xia, G.M. A novel high molecular weight glutenin subunit from Australopyrum retrofractum. Amino Acids 2010, 39, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Dai, S.; Pu, Z.; Liu, D.; Wei, Y.; Zheng, Y.; Hu, X.; Yan, Z. Characterization of novel HMW-GS in two diploid species of Eremopyrum. Gene 2013, 519, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Li, Z.; Gong, C.; Xu, H.; Yang, R.; Hao, S.; Wang, X.; Wang, D.; Zhang, X. Identification and characterization of high-molecular-weight glutenin subunits from Agropyron intermedium. PLoS ONE 2014, 9, e87477. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Liang, Y.; Qin, L.; Sun, L.; Xia, G.; Liu, S. Characterization of high molecular weight glutenin subunit genes from the Ns genome of Psathyrostachys juncea. Dev. Genes Evol. 2014, 224, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Pu, Z.; Dai, S.; Pu, X.; Liu, D.; Wu, B.; Lan, X.; Wei, Y.; Zheng, Y.; Yan, Z. Characterization of y-type high-molecular-weight glutenins in tetraploid species of Leymus. Dev. Genes Evol. 2014, 224, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.B.; Han, H.N.; Liang, Y.; Sun, L.; Xia, G.M.; Liu, S.W. Isolation and characterization of novel Glu-St1 alleles from Pseudoroegneria spicata and Pd. strigosa. Genetica 2014, 142, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Wang, S.; Zhen, S.; Shen, X.; Prodanovic, S.; Yan, Y. Molecular characterization and phylogenetic analysis of unusual x-type HMW glutenin subunits from 1SL genome of Aegilops longissima. Genetika 2015, 47, 185–203. [Google Scholar] [CrossRef]
- Shewry, P.R.; Halford, N.G.; Tatham, A.S.; Popineau, Y.; Lafiandra, D.; Belton, P.S. The high molecular weight subunits of wheat glutenin and their role in determining wheat processing properties. Adv. Food Nutr. Res. 2003, 45, 219–302. [Google Scholar] [PubMed]
- Baum, B.R.; Yen, C.; Yang, J.L. Roegneria: Its generic limits and justification for its recognition. Can. J. Bot. 1991, 69, 282–294. [Google Scholar] [CrossRef]
- Yang, J.L.; Baum, B.R.; Yen, C. A revision of the genus Roegneria C. Koch (Triticeae: Poaceae). J. Sichuan Agric. Univ. 2008, 26, 311–381. [Google Scholar]
- Dong, Y.S.; Zhou, R.H.; Xu, S.J.; Li, L.H.; Cauderon, Y.; Wang, R.R.-C. Desirable characteristics in perennial Triticeae collected in China for wheat improvement. Hereditas 1992, 116, 175–178. [Google Scholar] [CrossRef]
- Wan, Y.F.; Yen, C.; Yang, J.L.; Liu, F.Q. Evaluation of Roegneria for resistance to head scab caused by Fusarium graminearum Schwabe. Genet. Resour. Crop Evol. 1987, 44, 211–215. [Google Scholar]
- Wang, X.E.; Chen, P.D.; Liu, D.J.; Zhang, P.; Zhou, B.; Friebe, B.; Gill, B.S. Molecular cytogenetic characterization of Roegneria ciliaris chromosome additions in common wheat. Theor. Appl. Genet. 2001, 102, 651–657. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information (NCBI). GenBank. Available online: https://www.ncbi.nlm.nih.gov/genbank/.
- Shewry, P.R.; Napier, J.A.; Tatham, A.S. Seed storage proteins: Structures and biosynthesis. Plant Cell 1995, 7, 945–956. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.T.; Zhu, X.L.; Tan, Y.; Liu, S.W. Isolation and characterization of Glu-1 genes from the St genome of Pseudoroegneria libanotica. Gene 2012, 499, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Li, S.F.; Zhou, H.B.; Li, L.H.; Wang, X.P.; Zhang, X.Q. Isolation and characterization of high-molecular-weight glutenin subunit genes in Agropyron cristatum. Acta Agric. Sin. 2007, 33, 63–69. [Google Scholar]
- Zhang, L.; Chen, Q.; Su, M.; Yan, B.; Zhang, X.; Jiao, Z. High-molecular-weight glutenin subunit-deficient mutants induced by ion beam and the effects of Glu-1 loci deletion on wheat quality properties. J. Sci. Food Agric. 2016, 96, 1289–1296. [Google Scholar] [CrossRef] [PubMed]
- Denery-Papini, S.; Popineau, Y.; Quillien, L.; van Regenmortel, M.H.V. Specificity of antisera raised against synthetic peptide fragments of high Mr glutenin subunits. J. Cereal Sci. 1996, 23, 133–144. [Google Scholar] [CrossRef]
- Webb, D.M.; Knapp, S.J. DNA extraction from a previously recalcitrant plant genus. Plant Mol. Biol. Rep. 1990, 8, 180–185. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | GenBank Accession Number | ORF Size | Signal Peptide | N-terminal | Repetitive Domain | C-terminal | Total | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Size | Size | Cys | Size | Cys | Size | Cys | Size | Cys | |||
| Rny1 | KP121402 | 1371 bp | 18 | 102 | 5 | 293 | 0 | 42 | 1 | 455 | 6 |
| Rny2 | KP121401 | 1253 bp | 21 | 105 | 5 | 250 | 0 | 41 | 1 | 415 | 6 |
| Rny3 | KP121404 | 1452 bp | 18 | 102 | 5 | 320 | 0 | 42 | 1 | 482 | 6 |
| Rny4 | KP121403 | 1280 bp | 21 | 105 | 5 | 259 | 0 | 41 | 1 | 424 | 6 |
| Ray1 | KP121400 | 1389 bp | 18 | 102 | 5 | 299 | 0 | 42 | 1 | 461 | 6 |
| Ray2 | KP121399 | 1271 bp | 21 | 105 | 5 | 238 | 0 | 41 | 1 | 403 | 6 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Li, Z.; Fan, R.; Wei, B.; Zhang, X. Structural Characterization and Evolutionary Relationship of High-Molecular-Weight Glutenin Subunit Genes in Roegneria nakaii and Roegneria alashanica. Int. J. Mol. Sci. 2016, 17, 1115. https://doi.org/10.3390/ijms17071115
Zhang L, Li Z, Fan R, Wei B, Zhang X. Structural Characterization and Evolutionary Relationship of High-Molecular-Weight Glutenin Subunit Genes in Roegneria nakaii and Roegneria alashanica. International Journal of Molecular Sciences. 2016; 17(7):1115. https://doi.org/10.3390/ijms17071115
Chicago/Turabian StyleZhang, Lujun, Zhixin Li, Renchun Fan, Bo Wei, and Xiangqi Zhang. 2016. "Structural Characterization and Evolutionary Relationship of High-Molecular-Weight Glutenin Subunit Genes in Roegneria nakaii and Roegneria alashanica" International Journal of Molecular Sciences 17, no. 7: 1115. https://doi.org/10.3390/ijms17071115