
Quantitative Expression Analysis of APP Pathway and Tau Phosphorylation-Related Genes in the ICV STZ-Induced Non-Human Primate Model of Sporadic Alzheimer’s Disease

 and
and
Abstract
:1. Introduction
2. Results
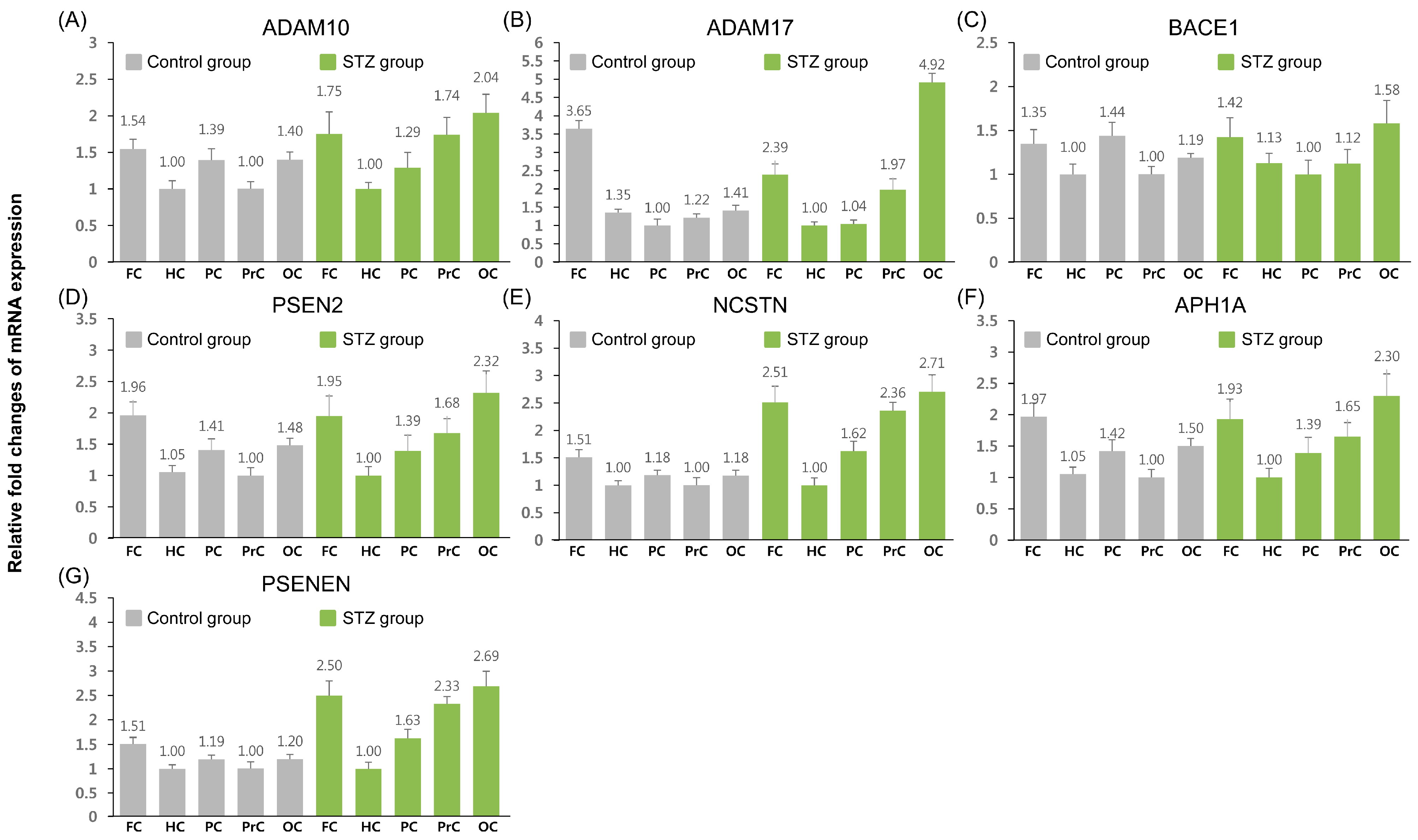
2.1. Relative Expression Analysis of Amyloid Precursor Protein (APP) Pathway-Related Genes


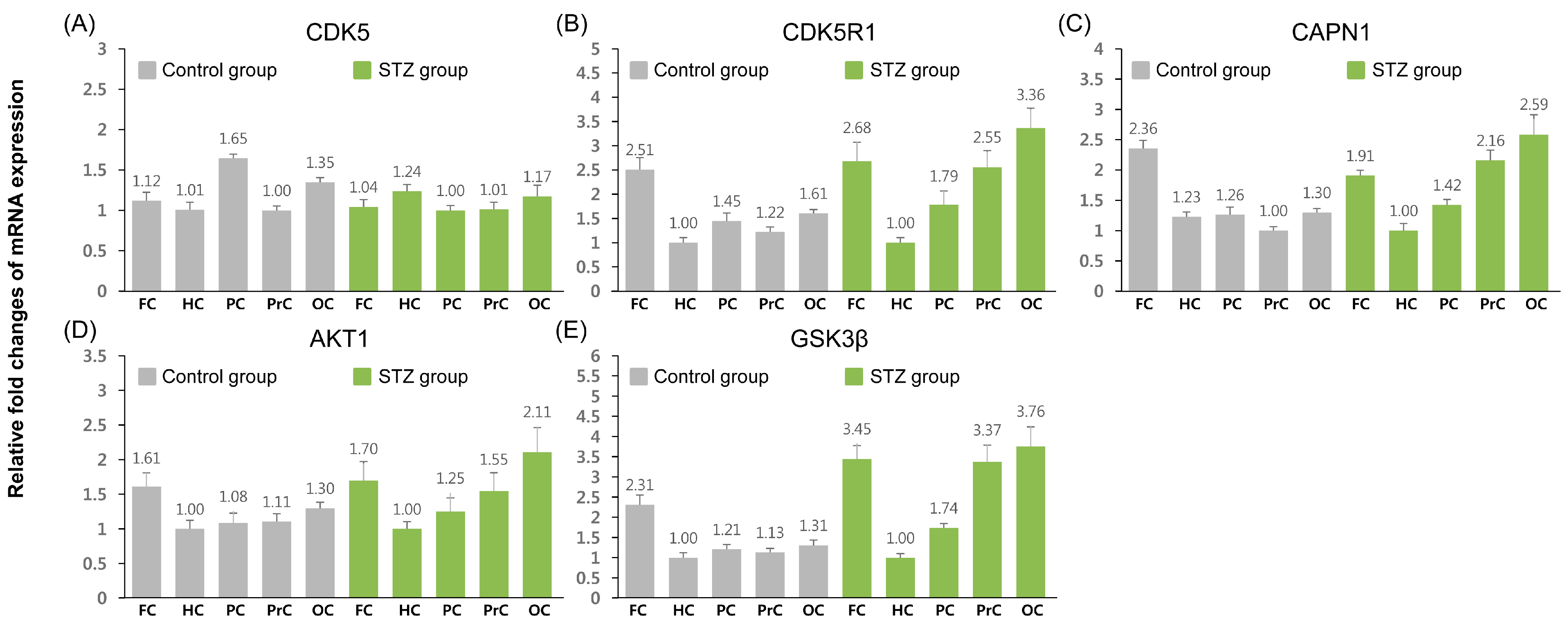
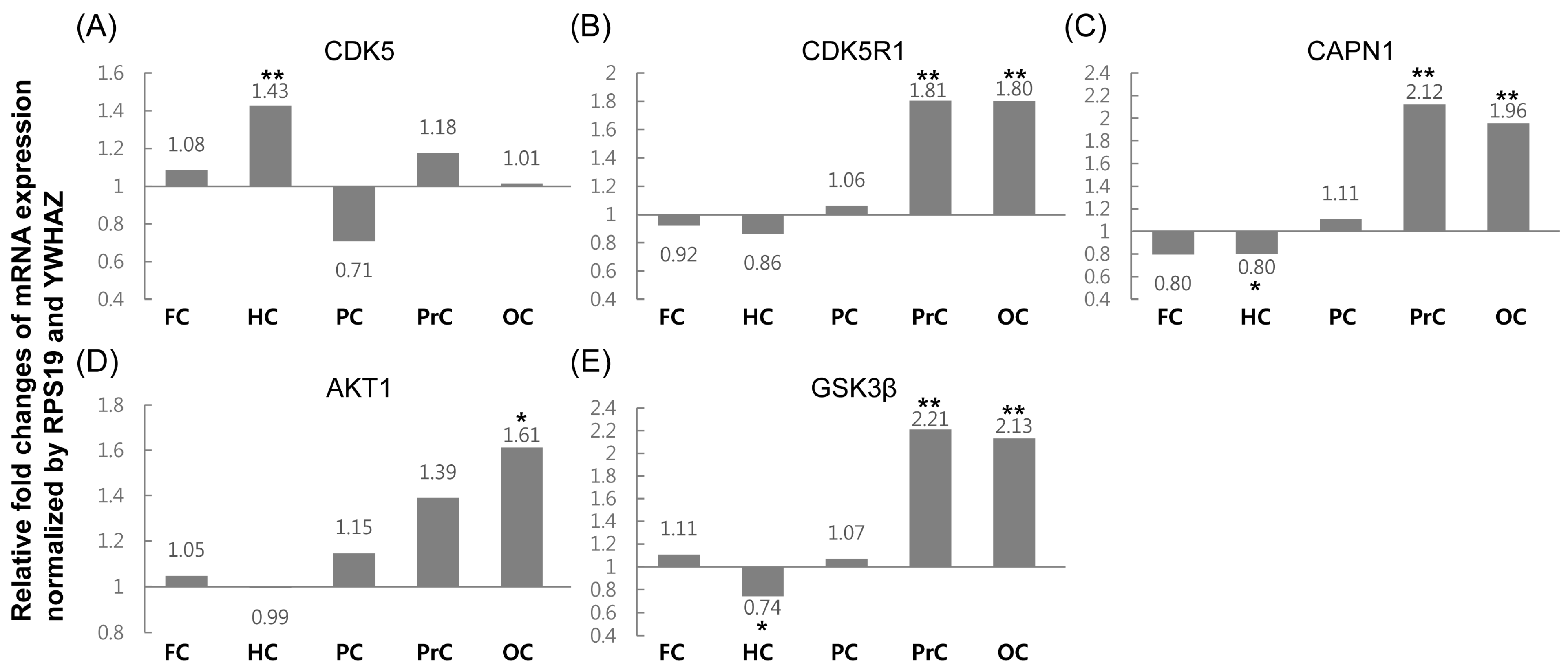
2.2. Relative Expression Analysis of Tau Phosphorylation-Related Genes


3. Discussion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Frontal Cortex | Hippocampus | Posterior Cingulate | Precuneus | Occipital Cortex |
|---|---|---|---|---|---|
| APP | 1.14 | 0.94 | ↑ 1.23 * | ↑ 2.17 * | ↑ 1.44 ** |
| ADAM10 | 1.04 | 0.91 | 0.85 | ↑ 1.58 * | ↑ 1.33 * |
| ADAM17 | ↓ 0.82 ** | 0.54 | 1.18 | ↑ 2.12 * | ↑ 2.07 ** |
| BACE1 | 1.02 | 1.09 | 0.67 | 1.09 | ↑ 1.29 * |
| PSEN2 | 1.00 | 0.96 | 0.99 | ↑ 1.69 * | ↑ 1.57 ** |
| NCSTN | ↑ 1.42 * | 0.85 | 1.17 | ↑ 2.01 * | ↑ 1.96 ** |
| APH1A | 1.00 | 0.96 | 0.99 | ↑ 1.68 ** | ↑ 1.56 ** |
| PSENEN | ↑ 1.43 * | 0.87 | 1.18 | ↑ 2.01 * | ↑ 1.94 ** |
| TAU | 1.14 | 1.10 | 0.96 | 1.18 | 1.21 |
| CDK5 | 1.08 | ↑ 1.43 ** | 0.71 | 1.18 | 1.01 |
| CDK5R1 | 0.92 | 0.86 | 1.06 | ↑ 1.81 ** | ↑ 1.80 ** |
| CAPN1 | 0.80 | 0.80 * | 1.11 | ↑ 2.12 ** | ↑ 1.96 ** |
| AKT1 | 1.05 | 0.99 | 1.15 | 1.39 | ↑ 1.61 ** |
| GSK3β | 1.11 | ↓ 0.74 * | 1.07 | ↑ 2.21 ** | ↑ 2.13 ** |
4. Experimental Section
4.1. Experimental Animals and Sampling
4.2. Ethics Statement
4.3. Total RNA Isolation and cDNA Preparation
4.4. Primer Design and Standard Curve Analysis
| Gene Symbol | Gene Name | Primer * Forward (F)/Reverse (R) 5'–3' | Exon(s) | Amplicon Size (bp) | PCR Efficiency (%) | R2 | NTC ** (Cq) |
|---|---|---|---|---|---|---|---|
| ADAM10 | ADAM metallopeptidase domain 10 | F: TGCAAACTGAAACCTGGGAA | 11 | 121 | 92 | 0.98922 | 31.3 |
| ADAM17 | ADAM metallopeptidase domain 17 | F: CATGAAT/GGCAAATGTGAGAAAC | 15/16 | 168 | 81 | 0.99394 | 33.8 |
| BACE1 | Beta-site APP-cleaving enzyme 1 | F: CGGGTGGAGATCAATGGACA | 5 | 194 | 82 | 0.99029 | N.d. |
| PSEN2 | Presenilin 2 (Alzheimer disease 4) | F: CCGCTGCTACAAG/TTCATCC | 5/6 | 143 | 93 | 0.99045 | 31.9 |
| NCSTN | Nicastrin | F: CTGTGTTCGCCTGCTCAAC | 2 | 123 | 92 | 0.99172 | 34.1 |
| APH1A | Anterior pharynx defective 1 homolog A (C. elegans) | F: ACCTACTGACATCGGGACTG | 5 | 140 | 96 | 0.99049 | 32.7 |
| PSENEN | Presenilin enhancer 2 homolog (C. elegans) | F: ACCTGTGCCGGAAGTACTAC | 2 | 113 | 98 | 0.99198 | 36.9 |
| DK5 | Cyclin-dependent kinase 5 | F: CAGTGGCCCTCTATGACCAA | 10 | 76 | 86 | 0.99153 | N.d. |
| DK5R1 | Cyclin-dependent kinase 5, regulatory subunit 1 (p35) | F: GCTGCCTTGGAAGAGAATCG | 2 | 94 | 91 | 0.99565 | N.d. |
| CAPN1 | Calpain 1, (mu/I) large subunit | F: ATGACCAGATCCAGGCCAAT | 13 | 122 | 93 | 0.99090 | N.d. |
| AKT1 | V-akt murine thymoma viral oncogene homolog 1 | F: CCACGCTACTTCCTCCTCAA | 3 | 159 | 92 | 0.99906 | 35.62 |
| GSK3β | Glycogen synthase kinase 3 beta | F: TCGCCATCAAGAAAGTATTGCA | 2 | 94 | 86 | 0.99641 | N.d. |
| ACTB | Beta-actin | F: ACAGAGCCTCGCCTTTGC | 1 | 160 | 92 | 0.99094 | 32 |
| GAPDH | Glyceraldehyde-3-phospate dehydrogenase | F: ACAACAGCCTCAAGATCGTCAG | 6 | 112 | 90 | 0.99273 | 34.18 |
| RPS19 | Ribosomal protein S19 | F: AGCTTGCTCCCTACGATGAG | 3 | 174 | 93 | 0.99581 | 36.04 |
| YWHAZ | Tyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation protein, zeta polypeptide | F: AGCAGATGGCTCGAGAATACA | 2 | 185 | 97 | 0.99120 | 38.44 |
4.5. RT-qPCR Amplification
4.6. Statistical Analysis
4.7. Minimum Information for Publication of Quantitative Real-Time PCR Experiments (MIQE) Guidelines
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Querfurth, H.W.; LaFerla, F.M. Alzheimer’s disease. N. Engl. J. Med. 2010, 362, 329–344. [Google Scholar] [CrossRef] [PubMed]
- Grunblatt, E.; Hoyer, S.; Riederer, P. Gene expression profile in streptozotocin rat model for sporadic Alzheimer’s disease. J. Neural Transm. 2004, 111, 367–386. [Google Scholar] [CrossRef] [PubMed]
- Hoyer, S. Glucose metabolism and insulin receptor signal transduction in Alzheimer disease. Eur. J. Pharmacol. 2004, 490, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Blennow, K.; de Leon, M.J.; Zetterberg, H. Alzheimer’s disease. Lancet 2006, 368, 387–403. [Google Scholar] [CrossRef] [PubMed]
- Rivera, E.J.; Goldin, A.; Fulmer, N.; Tavares, R.; Wands, J.R.; de la Monte, S.M. Insulin and insulin-like growth factor expression and function deteriorate with progression of Alzheimer’s disease: Link to brain reductions in acetylcholine. J. Alzheimer’s Dis. 2005, 8, 247–268. [Google Scholar]
- Lester-Coll, N.; Rivera, E.J.; Soscia, S.J.; Doiron, K.; Wands, J.R.; de la Monte, S.M. Intracerebral streptozotocin model of type 3 diabetes: Relevance to sporadic Alzheimer’s disease. J. Alzheimer’s Dis. 2006, 9, 13–33. [Google Scholar]
- Grunblatt, E.; Salkovic-Petrisic, M.; Osmanovic, J.; Riederer, P.; Hoyer, S. Brain insulin system dysfunction in streptozotocin intracerebroventricularly treated rats generates hyperphosphorylated tau protein. J. Neurochem. 2007, 101, 757–770. [Google Scholar] [CrossRef]
- Mistur, R.; Mosconi, L.; Santi, S.D.; Guzman, M.; Li, Y.; Tsui, W.; de Leon, M.J. Current challenges for the early detection of Alzheimer’s disease: Brain imaging and CSF studies. J. Clin. Neurol. 2009, 5, 153–166. [Google Scholar] [CrossRef] [PubMed]
- Guglielmotto, M.; Aragno, M.; Tamagno, E.; Vercellinatto, I.; Visentin, S.; Medana, C.; Catalano, M.G.; Smith, M.A.; Perry, G.; Danni, O.; et al. Ages/rage complex upregulates BACE1 via NF-κB pathway activation. Neurobiol. Aging 2012, 33, 196.e113–196.e127. [Google Scholar]
- Shonesy, B.C.; Thiruchelvam, K.; Parameshwaran, K.; Rahman, E.A.; Karuppagounder, S.S.; Huggins, K.W.; Pinkert, C.A.; Amin, R.; Dhanasekaran, M.; Suppiramaniam, V. Central insulin resistance and synaptic dysfunction in intracerebroventricular-streptozotocin injected rodents. Neurobiol. Aging 2012, 33, 430.e5–430.e18. [Google Scholar] [PubMed]
- Sarasa, M.; Pesini, P. Natural non-trasgenic animal models for research in Alzheimer’s disease. Curr. Alzheimer Res. 2009, 6, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.H.; Lee, S.R.; Lee, S.T.; Lee, K.M.; Oh, J.H.; Jang, D.P.; Chang, K.T.; Cho, Z.H. Spatial distribution of glucose hypometabolism induced by intracerebroventricular streptozotocin in monkeys. J. Alzheimer’s Dis. 2011, 25, 517–523. [Google Scholar]
- Kimura, N.; Tanemura, K.; Nakamura, S.; Takashima, A.; Ono, F.; Sakakibara, I.; Ishii, Y.; Kyuwa, S.; Yoshikawa, Y. Age-related changes of Alzheimer’s disease-associated proteins in cynomolgus monkey brains. Biochem. Biophys. Res. Commun. 2003, 310, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Oikawa, N.; Kimura, N.; Yanagisawa, K. Alzheimer-type tau pathology in advanced aged nonhuman primate brains harboring substantial amyloid deposition. Brain Res. 2010, 1315, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; de Preter, K.; Pattyn, F.; Poppe, B.; van Roy, N.; de Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.J.; Kim, Y.H.; Huh, J.W.; Lee, S.R.; Kim, S.H.; Kim, S.U.; Kim, J.S.; Jeong, K.J.; Kim, K.M.; Kim, H.S.; et al. Selection of new appropriate reference genes for RT-qPCR analysis via transcriptome sequencing of cynomolgus monkeys (Macaca fascicularis). PLoS One 2013, 8, e60758. [Google Scholar] [CrossRef]
- Park, S.J.; Huh, J.W.; Kim, Y.H.; Lee, S.R.; Kim, S.H.; Kim, S.U.; Kim, H.S.; Kim, M.K.; Chang, K.T. Selection of internal reference genes for normalization of quantitative reverse transcription polymerase chain reaction (qRT-PCR) analysis in the canine brain and other organs. Mol. Biotechnol. 2013, 54, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Devi, L.; Alldred, M.J.; Ginsberg, S.D.; Ohno, M. Mechanisms underlying insulin deficiency-induced acceleration of β-amyloidosis in a mouse model of Alzheimer’s disease. PLoS One 2012, 7, e32792. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Kim, Y.H.; Lee, Y.; Kim, K.M.; Kim, H.S.; Lee, S.R.; Kim, S.U.; Kim, S.H.; Kim, J.S.; Jeong, K.J.; et al. Selection of appropriate reference genes for RT-qPCR analysis in a streptozotocin-induced Alzheimer’s disease model of cynomolgus monkeys (Macaca fascicularis). PLoS One 2013, 8, e56034. [Google Scholar] [CrossRef]
- Priller, C.; Bauer, T.; Mitteregger, G.; Krebs, B.; Kretzschmar, H.A.; Herms, J. Synapse formation and function is modulated by the amyloid precursor protein. J. Neurosci. 2006, 26, 7212–7221. [Google Scholar] [CrossRef] [PubMed]
- Clarke, J.; Thornell, A.; Corbett, D.; Soininen, H.; Hiltunen, M.; Jolkkonen, J. Over-expression of APP provides neuroprotection in the absence of functional benefit following middle cerebral artery occlusion in rats. Eur. J. Neurosci. 2007, 26, 1845–1852. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Eckman, C.; Younkin, S.; Hsiao, K.K.; Iadecola, C. Increased susceptibility to ischemic brain damage in transgenic mice overexpressing the amyloid precursor protein. J. Neurosci. 1997, 17, 7655–7661. [Google Scholar] [PubMed]
- Matsuyama, S.; Teraoka, R.; Mori, H.; Tomiyama, T. Inverse correlation between amyloid precursor protein and synaptic plasticity in transgenic mice. Neuroreport 2007, 18, 1083–1087. [Google Scholar] [CrossRef] [PubMed]
- Patrick, G.N.; Zukerberg, L.; Nikolic, M.; de la Monte, S.; Dikkes, P.; Tsai, L.H. Conversion of p35 to p25 deregulates CDK5 activity and promotes neurodegeneration. Nature 1999, 402, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S.; Kwon, Y.T.; Li, M.; Peng, J.; Friedlander, R.M.; Tsai, L.H. Neurotoxicity induces cleavage of p35 to p25 by calpain. Nature 2000, 405, 360–364. [Google Scholar] [CrossRef] [PubMed]
- Lebel, M.; Patenaude, C.; Allyson, J.; Massicotte, G.; Cyr, M. Dopamine D1 receptor activation induces tau phosphorylation via CDK5 and GSK3 signaling pathways. Neuropharmacology 2009, 57, 392–402. [Google Scholar] [CrossRef] [PubMed]
- Jope, R.S.; Johnson, G.V. The glamour and gloom of glycogen synthase kinase-3. Trends Biochem. Sci. 2004, 29, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.; Latypova, X.; Wilson, C.M.; Magnaudeix, A.; Perrin, M.L.; Yardin, C.; Terro, F. Tau protein kinases: Involvement in Alzheimer’s disease. Ageing Res. Rev. 2013, 12, 289–309. [Google Scholar] [CrossRef] [PubMed]
- Salkovic-Petrisic, M.; Knezovic, A.; Hoyer, S.; Riederer, P. What have we learned from the streptozotocin-induced animal model of sporadic Alzheimer’s disease, about the therapeutic strategies in Alzheimer’s research. J. Neural Transm. 2013, 120, 233–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Tian, Z.; Liang, Z.; Sun, S.; Dai, C.L.; Lee, M.H.; LaFerla, F.M.; Grundke-Iqbal, I.; Iqbal, K.; Liu, F.; et al. Brain gene expression of a sporadic (icv-STZ mouse) and a familial mouse model (3xTg-AD mouse) of Alzheimer’s disease. PLoS One 2012, 7, e51432. [Google Scholar] [CrossRef]
- Rozen, S.; Skaletsky, H. Primer3 on the www for general users and for biologist programmers. Methods Mol. Biol. 2000, 132, 365–386. [Google Scholar] [PubMed]
- Huh, J.W.; Kim, Y.H.; Park, S.J.; Kim, D.S.; Lee, S.R.; Kim, K.M.; Jeong, K.J.; Kim, J.S.; Song, B.S.; Sim, B.W.; et al. Large-scale transcriptome sequencing and gene analyses in the crab-eating macaque (Macaca fascicularis) for biomedical research. BMC Genomics 2012, 13, 163. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef] [PubMed]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, S.-J.; Kim, Y.-H.; Nam, G.-H.; Choe, S.-H.; Lee, S.-R.; Kim, S.-U.; Kim, J.-S.; Sim, B.-W.; Song, B.-S.; Jeong, K.-J.; et al. Quantitative Expression Analysis of APP Pathway and Tau Phosphorylation-Related Genes in the ICV STZ-Induced Non-Human Primate Model of Sporadic Alzheimer’s Disease. Int. J. Mol. Sci. 2015, 16, 2386-2402. https://doi.org/10.3390/ijms16022386
Park S-J, Kim Y-H, Nam G-H, Choe S-H, Lee S-R, Kim S-U, Kim J-S, Sim B-W, Song B-S, Jeong K-J, et al. Quantitative Expression Analysis of APP Pathway and Tau Phosphorylation-Related Genes in the ICV STZ-Induced Non-Human Primate Model of Sporadic Alzheimer’s Disease. International Journal of Molecular Sciences. 2015; 16(2):2386-2402. https://doi.org/10.3390/ijms16022386
Chicago/Turabian StylePark, Sang-Je, Young-Hyun Kim, Gyu-Hwi Nam, Se-Hee Choe, Sang-Rae Lee, Sun-Uk Kim, Ji-Su Kim, Bo-Woong Sim, Bong-Seok Song, Kang-Jin Jeong, and et al. 2015. "Quantitative Expression Analysis of APP Pathway and Tau Phosphorylation-Related Genes in the ICV STZ-Induced Non-Human Primate Model of Sporadic Alzheimer’s Disease" International Journal of Molecular Sciences 16, no. 2: 2386-2402. https://doi.org/10.3390/ijms16022386