Characterization of Bacterial Communities and Their Antibiotic Resistance Profiles in Wastewaters Obtained from Pharmaceutical Facilities in Lagos and Ogun States, Nigeria

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites and Sampling
2.2. Bacterial Isolation and Counts
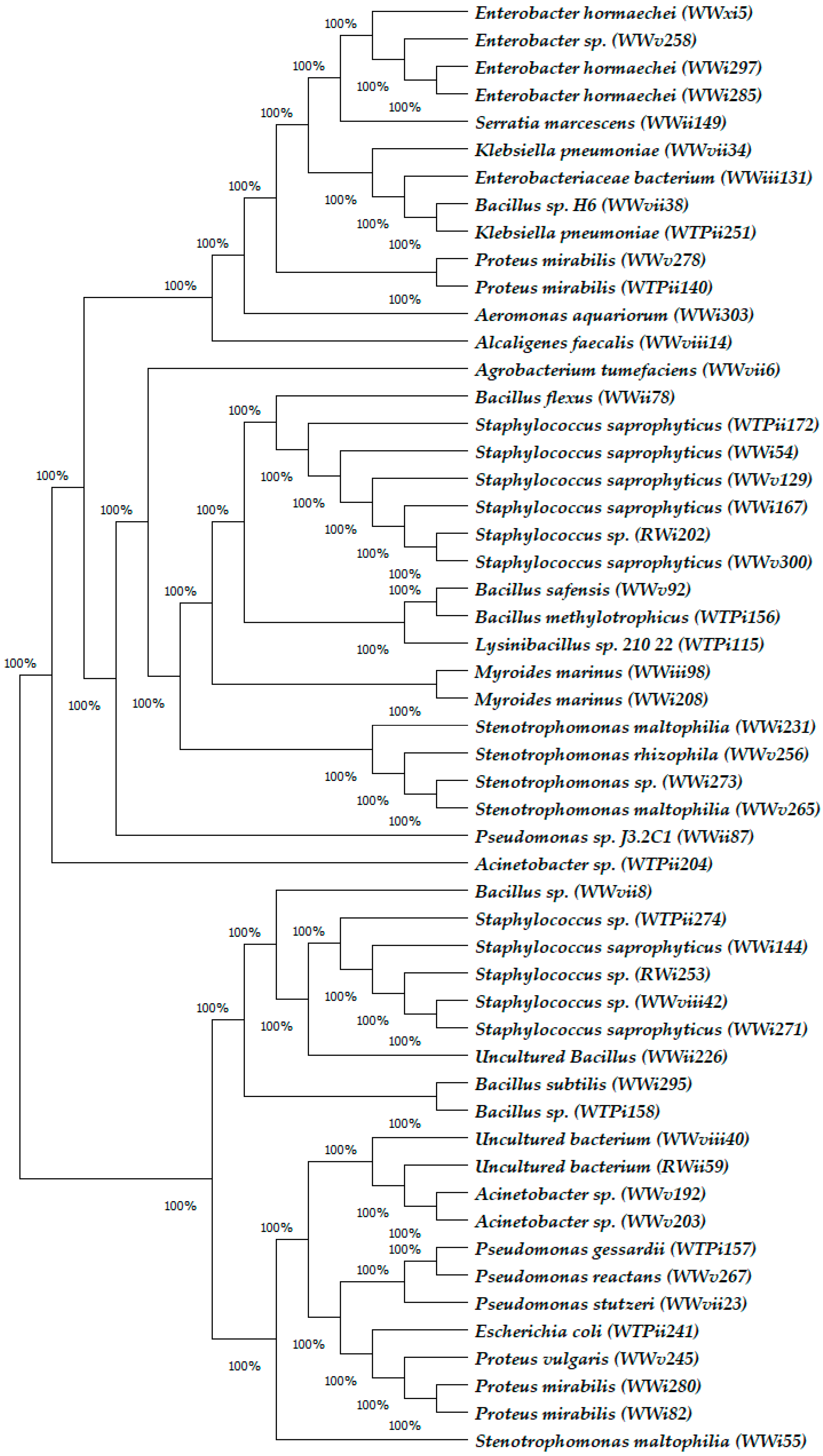
2.3. Bacterial Identification
2.4. Antibiotic Susceptibility Testing
2.5. PCR Detection of Sulfonamide Resistance Genes and Class I Integrons
2.6. Statistical Analysis
2.7. Nucleotide Sequence Accession Numbers
3. Results
3.1. Total Bacterial Counts
3.2. Composition of Bacterial Isolates
3.3. Antibiotics Resistance Prevalence and MDR
3.4. Sulfonamide Resistance Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Berendonk, T.U.; Manaia, C.M.; Merlin, C.; Fatta-Kassinos, D.; Cytryn, E.; Walsh, F.; Burgmann, H.; Sorum, H.; Norstrom, M.; Pons, M.N.; et al. Tackling antibiotic resistance: The environmental framework. Nat. Rev. Microbiol. 2015, 13, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.R.; Stedtfeld, R.D.; Guo, X.; Hashsham, S.A. Antimicrobial resistance in the environment. Water Environ. Res. 2016, 88, 1951–1967. [Google Scholar] [CrossRef] [PubMed]
- Lien, L.T.; Lan, P.T.; Chuc, N.T.; Hoa, N.Q.; Nhung, P.H.; Thoa, N.T.; Tamhankar, A.J.; Lundborg, C.S. Antibiotic Resistance and Antibiotic Resistance Genes in Escherichia coli Isolates from Hospital Wastewater in Vietnam. Int. J. Environ. Res. Public Health 2017, 14, 699. [Google Scholar] [CrossRef] [PubMed]
- Goossens, H.; Ferech, M.; Stichele, R.V.; Elseviers, M. Outpatient antibiotic use in Europe and association with resistance: A cross-national database study. Lancet 2005, 365, 579–587. [Google Scholar] [CrossRef]
- World Health Organization. Antimicrobial Resistance: Global Report on Surveillance 2014; WHO: Geneva, Swizerland, 2015. [Google Scholar]
- Bollenbach, T. Antimicrobial interactions: Mechanisms and implications for drug discovery and resistance evolution. Curr. Opin. Microbiol. 2015, 27, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Levy, S.B.; Marshall, B. Antibacterial resistance worldwide: Causes, challenges and responses. Nat. Med. 2004, 10, S122–S129. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Yu, T.; Zhang, Y.; Yang, M.; Li, Z.; Liu, M.; Qi, R. Antibiotic Resistance Characteristics of Environmental Bacteria from an Oxytetracycline Production Wastewater Treatment Plant and the Receiving River. Appl. Environ. Microbiol. 2010, 76, 3444–3451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, K.D.; Kulis, J.; Thomson, B.; Chapman, T.H.; Mawhinney, D.B. Occurrence of antibiotics in hospital, residential, and dairy effluent, municipal wastewater, and the Rio Grande in New Mexico. Sci. Total Environ. 2006, 366, 772–783. [Google Scholar] [CrossRef] [PubMed]
- Kümmerer, K. Antibiotics in the aquatic environment—A review—Part II. Chemosphere 2009, 75, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Karkman, A.; Do, T.T.; Walsh, F.; Virta, M.P.J. Antibiotic-resistance genes in waste water-review. Trends Microbiol. 2017, 26, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Yang, M.; Hu, J.; Zhang, Y.; Chang, H.; Jin, F. Determination of penicillin G and its degradation products in a penicillin production wastewater treatment plant and the receiving river. Water Res. 2008, 42, 307–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hultman, J.; Tamminen, M.; Parnanen, K.; Cairns, J.; Karkman, A.; Virta, M. Host range of antibiotic resistance genes in wastewater treatment plant influent and effluent. FEMS Microbiol. Ecol. 2018, 94, fiy038. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, J.; Leonard, M.; Greening, G.E.; Lewis, G.D. Influence of wastewater treatment process and the size on human virus profiles in wastewater. Water Res. 2011, 45, 6267–6276. [Google Scholar] [CrossRef] [PubMed]
- Leddy, M.B.; Hasan, N.; Subramanian, P.; Heberling, C.; Cotruvo, J.; Colwell, R.R. Characterization of Microbial Signatures From Advanced Treated Wastewater Biofilms. J. Am. Water Works Assoc. 2017, 109, E503–E512. [Google Scholar] [CrossRef]
- Aw, T.G.; Howe, A.; Rose, J.B. Metagenomic Approaches for Direct and Cell Culture Evaluation of the Virological Quality of Wastewater. J. Virol. Methods 2014, 210, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Iwane, T.; Urase, T.; Yamamoto, K. Possible impact of treated wastewater discharge on incidence of antibiotic resistant bacteria in river water. Water Sci. Technol. 2001, 43, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Yang, M.; Hu, J.; Ren, L.; Zhang, Y.; Li, K. Determination and fate of oxytetracycline and related compounds in oxytetracycline production wastewater and the receiving river. Environ. Toxicol. Chem. 2008, 27, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Szczepanowski, R.; Linke, B.; Krahn, I.; Gartemann, K.; Gutzkow, T.; Eichler, W.; Puhler, A.; Schluter, A. Detection of 140 clinically relevant antibiotic-resistance genes in the plasmid metagenome of wastewater treatment plant bacteria showing reduced susceptibility to selected antibiotics. Microbiology 2009, 155, 2306–2319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le, T.M.; Baker, S.; Le, T.P.; Cao, T.T.; Tran, T.T.; Nguyen, V.M.; Campbell, J.I.; Lam, M.Y.; Nguyen, T.H.; Nguyen, W.; et al. High prevalence of plasmid-mediated quinolone resistance determinants in commensal members of the Enterobacteriaceae in Ho Chi Minh City, Vietnam. J. Med. Microbiol. 2009, 58, 1585–1592. [Google Scholar] [PubMed] [Green Version]
- Quach-Cu, J.; Herrera-Lynch, B.; Marciniak, C.; Adams, S.; Simmerman, A.; Reinke, R.A. The Effect of Primary, Secondary, and Tertiary Wastewater Treatment Processes on Antibiotic Resistance Gene (ARG) Concentrations in Solid and Dissolved Wastewater Fractions. Water 2018, 10, 37. [Google Scholar] [CrossRef]
- D’Costa, V.M.; McGram, K.M.; Hughes, D.W.; Wright, G.D. Sampling the antibiotic resistome. Science 2006, 311, 374–377. [Google Scholar] [CrossRef] [PubMed]
- Dantas, G.; Sommer, M.O.A.; Oluwasegun, R.D.; Church, G.M. Bacteria subsisting on antibiotics. Science 2008, 320, 100–103. [Google Scholar] [CrossRef] [PubMed]
- Luby, E.; Ibekwe, A.M.; Zilles, J.; Pruden, A. Molecular Methods for Assessment of Antibiotic Resistance in Agricultural Ecosystems: Prospects and Challenges. J. Environ. Qual. 2016, 45, 441–453. [Google Scholar] [CrossRef] [PubMed]
- Ibekwe, A.M.; Gonzalez-Rubio, A.; Suarez, D.L. Impact of treated wastewater for irrigation on soil microbial communities. Sci. Total Environ. 2018, 622–623, 1603–1610. [Google Scholar] [CrossRef] [PubMed]
- Akinbowale, O.L.; Peng, H.; Barton, M.D. Class 1 integron mediates antibiotic resistance in Aeromonas spp. from rainbow trout farms in Australia. Int. J. Antimicrob. Agents 2007, 29, S113. [Google Scholar] [CrossRef]
- Agersø, Y.; Petersen, A. The tetracycline resistance determinant, Tet 39 and the Sulfonamide resistance gene sulII are common among resistant Acinetobacter spp. isolated from integrated fish farms in Thailand. J. Antimicrob. Chemother. 2007, 59, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Biyela, P.T. Convergent acquisition of antibiotic resistance determinants amongst the Enterobacteriaceae sp. isolates of the Mhlathuze River, KwaZulu-Natal (RSA). Water SA 2005, 31, 257–260. [Google Scholar] [CrossRef]
- Hu, J.Y.; Shi, J.C.; Chang, H.; Li, D.; Yang, M.; Kamagata, Y.C. Phenotyping and genotyping of antibiotic-resistant Escherichia coli isolated from a natural river basin. Environ. Sci. Technol. 2008, 42, 3415–3420. [Google Scholar] [CrossRef] [PubMed]
- Mohapatra, H.; Mohapatra, S.S.; Mantri, C.K.; Colwell, R.R.; Singh, D.V. Vibrio cholerae non-O1, non-O139 strains isolated before 1992 from Varanasi, India are multiple drug resistant, contain intSXT, dfr18 and aadA5 genes. Environ. Microbiol. 2008, 10, 866–873. [Google Scholar] [CrossRef] [PubMed]
- Tennstedt, T.; Szczepanowski, R.; Braun, S.; Pühler, A.; Schlüter, A. Occurrence of integron-associated resistance gene cassettes located on antibiotic resistance plasmids isolated from a wastewater treatment plant. FEMS Microbiol. Ecol. 2003, 45, 239–252. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, S.; Chakraborty, R. Incidence of class 1 integrons in multiple antibiotic-resistant Gram-negative copiotrophic bacteria from the River Torsa in India. Res. Microbiol. 2006, 157, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Taviani, E.; Ceccarelli, D.; Lazaro, N.; Bani, S.; Cappuccinelli, P.; Colwell, R.R.; Colombo, M.M. Environmental Vibrio spp., isolated in Mozambique, contain a polymorphic group of integrative conjugative elements and class 1 integrons. FEMS Microbiol. Ecol. 2008, 64, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Trobos, M.; Christensen, H.; Sunde, M.; Nordentoft, S.; Agersø, Y.; Simonsen, G.S.; Hammerum, A.M.; Olsen, J.E. Characterization of Sulfonamide resistant Escherichia coli using comparison of sul2 gene sequences and multilocus sequence typing. Microbiology 2009, 155, 831–836. [Google Scholar] [CrossRef] [PubMed]
- Skold, O. Sulfonamide resistance: Mechanisms and trends. Drug Resist. Updates 2000, 3, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Gundogdu, A.; Long, Y.B.; Vollmerhausen, T.L.; Katouli, M. Antimicrobial resistance and distribution of sul genes and integron-associated intI genes among uropathogenic Escherichia coli in Queensland, Australia. J. Med. Microbiol. 2011, 60, 1633–1642. [Google Scholar] [CrossRef] [PubMed]
- Toleman, M.A.; Bennett, P.M.; Bennett, D.M.; Jones, R.N.; Walsh, T.R. Global emergence of trimethoprim/sulfamethoxazole resistance in Stenotrophomonas maltophilia mediated by acquisition of sul genes. Emerg. Infect. Dis. 2007, 13, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Chukwura, E.I.; Okpokwasili, G.C. Impact of Brewery Wastewater on Recipient Aquatic Environments. In Proceedings of the an International Conference Organized by Foundation for African Development through International Biotechnology (FADIB), Enugu, Nigeria, 9–13 February 1997. [Google Scholar]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley: Chichester, UK, 1991; pp. 115–175. [Google Scholar]
- Altschul, F.S.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.R.; Chai, B.; Farris, R.J.; Wang, Q.; Kulam-Syed-Mohideen, A.S.; McGarrell, D.M.; Bandela, A. The ribosomal database project (RDP-II): Introducing my RDP space and quality controlled public data. Nucleic Acids Res. 2007, 35, D169–D172. [Google Scholar] [CrossRef] [PubMed]
- Bauer, A.W.; Kirby, W.M.; Sherris, J.C.; Turck, M. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing; 21st Informational Supplement. M100–S21, Replaces M100-S20 and M100-S20-U; CLSI: Wayne, PA, USA, 2013. [Google Scholar]
- Goldstein, C.; Lee, M.D.; Sanchez, S.; Hudson, C.; Phillips, B.; Register, B.; Grady, M.; Liebert, C.; Summers, A.O.; White, D.G. Incidence of class 1 and 2 integrases in clinical and commensal bacteria from livestock, companion animals, and exotics. Antimicrob. Agents Chemother. 2001, 45, 723–726. [Google Scholar] [CrossRef] [PubMed]
- Peirano, G.; Agersø, Y.; Aarestrup, F.M.; Falavina dos Reis, F.M.; Rodirgues, D.P. Occurrence of integrons and antimicrobial resistance genes among Salmonella enterica from Brazil. J. Antimicrob. Chemother. 2006, 58, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Guardabassi, L.; Petersen, A.; Olsen, J.E.; Dalsgaard, A. Antibiotic resistance in Acinetobacter spp. isolated from sewers receiving waste effluent from a hospital and a pharmaceutical plant. Appl. Environ. Microbiol. 1998, 64, 3499–3502. [Google Scholar] [PubMed]
- Li, D.; Yang, M.; Hu, J.; Zhang, J.; Liu, R.; Gu, X.; Zhang, Y.; Wang, Z. Antibiotic resistance profile in environmental bacteria isolated from penicillin production wastewater treatment plant and the receiving river. Environ. Microbiol. 2009, 11, 1506–1517. [Google Scholar] [CrossRef] [PubMed]
- Davies, J.; Davies, D. Origins and evolution of antibiotic resistance. Microbiol. Mol. Biol. Rev. 2010, 74, 417–433. [Google Scholar] [CrossRef] [PubMed]
- McArthur, J.V.; Tuckfield, R.C. Spatial patterns in antibiotic resistance among stream bacteria: Effects of industrial pollution. Appl. Environ. Microbiol. 2000, 66, 3722–3726. [Google Scholar] [CrossRef] [PubMed]
- Graham, D.W.; Olivares-Rieumont, S.; Knapp, C.W.; Lima, L.; Werner, D.; Bowen, E. Antibiotic Resistance Gene Abundances Associated with Waste Discharges to the Almendares River near Havana, Cuba. Environ. Sci. Technol. 2011, 45, 418–424. [Google Scholar] [CrossRef] [PubMed]
- Manaia, C.M.; Vaz-Moreira, I.; Nunes, O.C. Antibiotic Resistance in Waste Water and Surface Water and Human Health Implications. In Emerging Organic Contaminants and Human Health, Handbook of Environmental Chemistry; Barcelo, D., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; Volume 20, pp. 173–212. [Google Scholar]
- Packer, J.L.; Werner, J.J.; Latch, D.E.; McNeill, K.; Arnold, W.A. Photochemical fate of pharmaceuticals in the environment: Naproxen, diclofenac, clofibric acid, and ibuprofen. Aquat. Sci. Res. Across Bound. 2003, 65, 342–351. [Google Scholar] [CrossRef]
- Jones, O.A.; Voulvoulis, N.; Lester, J.N. Aquatic environmental assessment of the top 25 English prescription pharmaceuticals. Water Res. 2002, 36, 5013–5022. [Google Scholar] [CrossRef]
- Castensson, S. Pharmaceutical Waste. In Pharmaceuticals in the Environment: Sources, Fate, Effects and Risks, 3rd ed.; Kümmerer, K., Ed.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 489–499. [Google Scholar]
- Antunes, P.; Machado, J.; Peixe, L. Dissemination of a new gene cluster comprising sul3 (tnp-sul3-tnp) linked to class 1 integrons with an unusual 3′CS region (qacH) among Salmonella isolates. Int. J. Antimicrob. Agents 2007, 29, S112–S113. [Google Scholar] [CrossRef]
- Gillings, M.R.; Gaze, W.H.; Pruden, A.; Smalla, K.; Tiedje, J.M.; Zhu, Y. Using the class 1 integron-integrase gene as a proxy for anthropogenic pollution. ISME J. 2015, 9, 1269–1279. [Google Scholar] [CrossRef] [PubMed]
- Mazel, D. Integrons: Agents of bacterial evolution. Nat. Rev. Microbiol. 2006, 4, 608–620. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Target Gene | Sequence (5′-3′) | Amplicon Size (bp) | Annealing Temp. (°C) | Reference |
|---|---|---|---|---|
| 27 F | AGAGTTTGATCCTGGCTCAG | 1503 | 55 | [39] |
| 1492 R | TACGGYTACCTTGTTACGACTT | |||
| SulI, F | ATCGCAATAGTTGGCGAAGT | 798 | 55 | [44] |
| SulI, R | GCAAGGCGGAAACCCGCGCC | |||
| SulII, F | GCGCTCAAGGCAGATGGCATT | 284 | 70 | [44] |
| SulII, R | GCGTTTGATACCGGCACCCGT | |||
| intI1, F | CCTCCCGCACGATGATC | 280 | 55 | [45] |
| intI1, R | TCCACGCATCGTCAGGC | |||
| intI2, F | TTATTGCTGGGATTAGGC | 233 | 50 | [45] |
| intI2, R | ACGGCTACCCTCTGTTATC |
| Genus or Species | No of Isolates from | Total Number of Isolates | ||
|---|---|---|---|---|
| IWW | WTP | RW | ||
| Acinetobacter sp. | 25 | 1 | 5 | 31 |
| Aeromonas aquariorum | 3 | - | - | 3 |
| Agrobacterium tumefaciens | 3 | 2 | - | 5 |
| Alcaligenes faecalis | 1 | - | - | 1 |
| Bacillus flexus | 5 | 2 | 1 | 8 |
| Bacillus methylotrophicus | 3 | 1 | 1 | 5 |
| Bacillus safensis | 5 | - | 1 | 6 |
| Bacillus subtilis | 5 | 2 | 1 | 8 |
| Bacillus sp. | 15 | 1 | 2 | 18 |
| Enterobacter hormaechei | 6 | 1 | 1 | 8 |
| Enterobacter sp. | 20 | 1 | 3 | 24 |
| Enterobacteriaceae bacterium | 1 | - | 2 | 3 |
| Escherichia coli | 2 | 1 | - | 3 |
| Klebsiella pneumoniae | 14 | 2 | 2 | 18 |
| Lysinibacillus sp. 210_22 | 2 | 1 | - | 3 |
| Myroides marinus | 6 | 1 | 2 | 9 |
| Proteus mirabilis | 24 | 5 | 6 | 35 |
| Proteus vulgaris | 5 | 2 | 1 | 8 |
| Pseudomonas gessardii | 1 | 1 | - | 2 |
| Pseudomonas reactans | 2 | - | - | 2 |
| Pseudomonas sp. | 6 | 3 | 4 | 13 |
| Pseudomonas stutzeri | 1 | - | - | 1 |
| Serratia marcescens | 3 | 1 | - | 4 |
| Staphylococcus saprophyticus | 5 | 2 | 2 | 9 |
| Staphylococcus sp. | 6 | 1 | 2 | 9 |
| Stenotrophomonas maltophilia | 4 | 1 | 1 | 6 |
| Stenotrophomonas rhizophila | 2 | - | - | 2 |
| Stenotrophomonas sp. | 2 | - | - | 2 |
| Uncultured bacterium | 6 | 1 | 1 | 8 |
| TOTAL | 183 | 33 | 38 | 254 |
| Class of Antibiotics | Group | IWW | WTP | RW | Total |
|---|---|---|---|---|---|
| Penicillin/Clavulanic acid | Augmentin | 178 (97.3) | 30 (90.9) | 35 (92.1) | 243 (95.7) |
| Quinolones | Ofloxacin | 47 (25.7) | 2 (6.1) | 5 (13.2) | 54 (21.3) |
| Nalidixic Acid | 82 (44.8) | 5 (15.2) | 4 (10.5) | 91 (35.8) | |
| Aminoglycosides | Gentamicin | 46 (25.1) | 11 (33.3) | 2 (5.3) | 59 (23.2) |
| Nitrofurans | Nitrofurantoin | 89 (48.6) | 15 (45.5) | 21 (55.3) | 125 (49.2) |
| Sulfonamides | Cotrimoxazole | 158 (86.3) | 14 (42.4) | 16 (42.1) | 188 (74.0) |
| Penicillins | Amoxycillin | 180 (98.3) | 23 (69.7) | 31 (81.6) | 234 (92.1) |
| Tetracyclines | Tetracycline | 127 (69.4) | 15 (45.5) | 5 (13.2) | 147 (57.9) |
| MDR | 3 classes and above | 171 (93.4) | 22 (66.7) | 25 (65.8) | 218 (85.8) |
| NMDR | Less than 3 classes | 12 (6.6) | 11 (33.3) | 13 (34.2) | 36 (14.2) |
| Total Isolates | - | 183 | 33 | 38 | 254 |
| Activity against the Isolates | ||||
|---|---|---|---|---|
| Antibiotics | Resistance Prevalence (%) | MIC (mg L−1) | ||
| Range | 50% | 90% | ||
| Ampicillin (AMP) | 100.0 | 64 to ≥1024 | 1024 | 1024 |
| Amoxicillin (AMO) | 100.0 | 1 to ≥1024 | 1024 | 1024 |
| Streptomycin Sulfate (STR) | 92.3 | 1 to ≥1024 | 128 | 512 |
| Trimethoprim (TRI) | 98.0 | 8 to ≥1024 | 1024 | 1024 |
| Chloramphenicol (CHL) | 100.0 | 256 to ≥1024 | 1024 | 1024 |
| Sulfonamide (SUL) | 100.0 | 512 to ≥1024 | 1024 | 1024 |
| Tetracycline (TET) | 90.6 | 2 to ≥512 | 128 | 256 |
| Oxytetracycline (OXY) | 90.6 | 1 to ≥1024 | 256 | 512 |
| Nalidixic Acid (NAL) | 73.6 | 1 to ≥1024 | 512 | 1024 |
| Erythromycin (ERY) | 92.5 | 4 to ≥1024 | 128 | 512 |
| Spiramycin (SPIRA) | 90.6 | 2 to ≥1024 | 512 | 1024 |
| Kanamycin (KAN) | 54.7 | 1 to ≥1024 | 128 | 1024 |
| Genes Class | Resistance Genes | Bacterial Isolates (%) |
|---|---|---|
| Sulfonamide genes | sul1 | 31.7 |
| sul2 | 21.7 | |
| Mobile genetic elements | Intl1 | 43.3 |
| Intl2 | 0 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Obayiuwana, A.; Ogunjobi, A.; Yang, M.; Ibekwe, M. Characterization of Bacterial Communities and Their Antibiotic Resistance Profiles in Wastewaters Obtained from Pharmaceutical Facilities in Lagos and Ogun States, Nigeria. Int. J. Environ. Res. Public Health 2018, 15, 1365. https://doi.org/10.3390/ijerph15071365
Obayiuwana A, Ogunjobi A, Yang M, Ibekwe M. Characterization of Bacterial Communities and Their Antibiotic Resistance Profiles in Wastewaters Obtained from Pharmaceutical Facilities in Lagos and Ogun States, Nigeria. International Journal of Environmental Research and Public Health. 2018; 15(7):1365. https://doi.org/10.3390/ijerph15071365
Chicago/Turabian StyleObayiuwana, Amarachukwu, Adeniyi Ogunjobi, Min Yang, and Mark Ibekwe. 2018. "Characterization of Bacterial Communities and Their Antibiotic Resistance Profiles in Wastewaters Obtained from Pharmaceutical Facilities in Lagos and Ogun States, Nigeria" International Journal of Environmental Research and Public Health 15, no. 7: 1365. https://doi.org/10.3390/ijerph15071365