Association of ATG4B and Phosphorylated ATG4B Proteins with Tumorigenesis and Prognosis in Oral Squamous Cell Carcinoma

 ,
,
Abstract
:1. Introduction
2. Experimental Procedure
2.1. Tissue Specimens and Tissue Microarray (TMA) Construction
2.2. Immunohistochemistry (IHC)
2.3. Immunohistochemistry Analysis and Score
2.4. Cell Culture and Transient Transfection
2.5. Analysis of Cell Viability, Migration, and Invasion
2.6. Statistical Analysis
3. Results
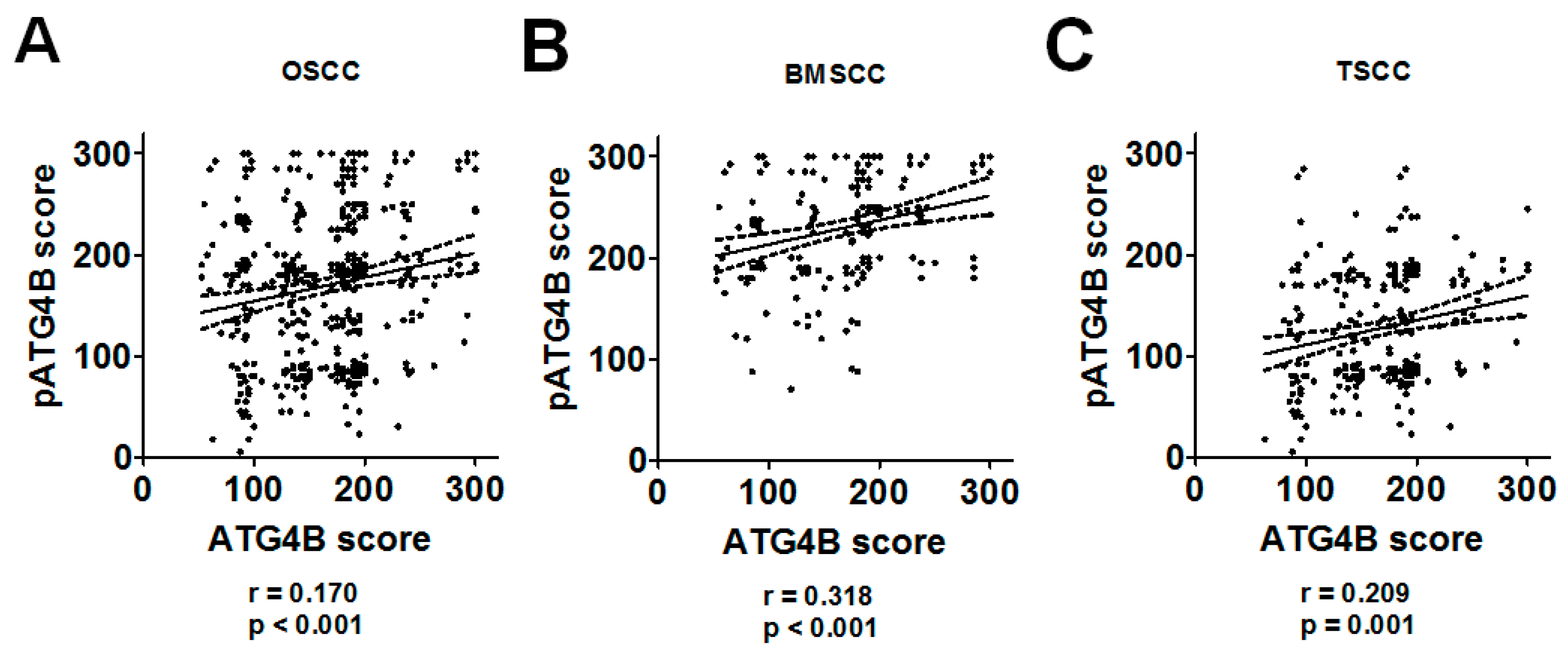
3.1. Association of the ATG4B and Phospho-Ser383/392-ATG4B Protein Levels with Tumorigenesis and Clinicopathological Outcomes
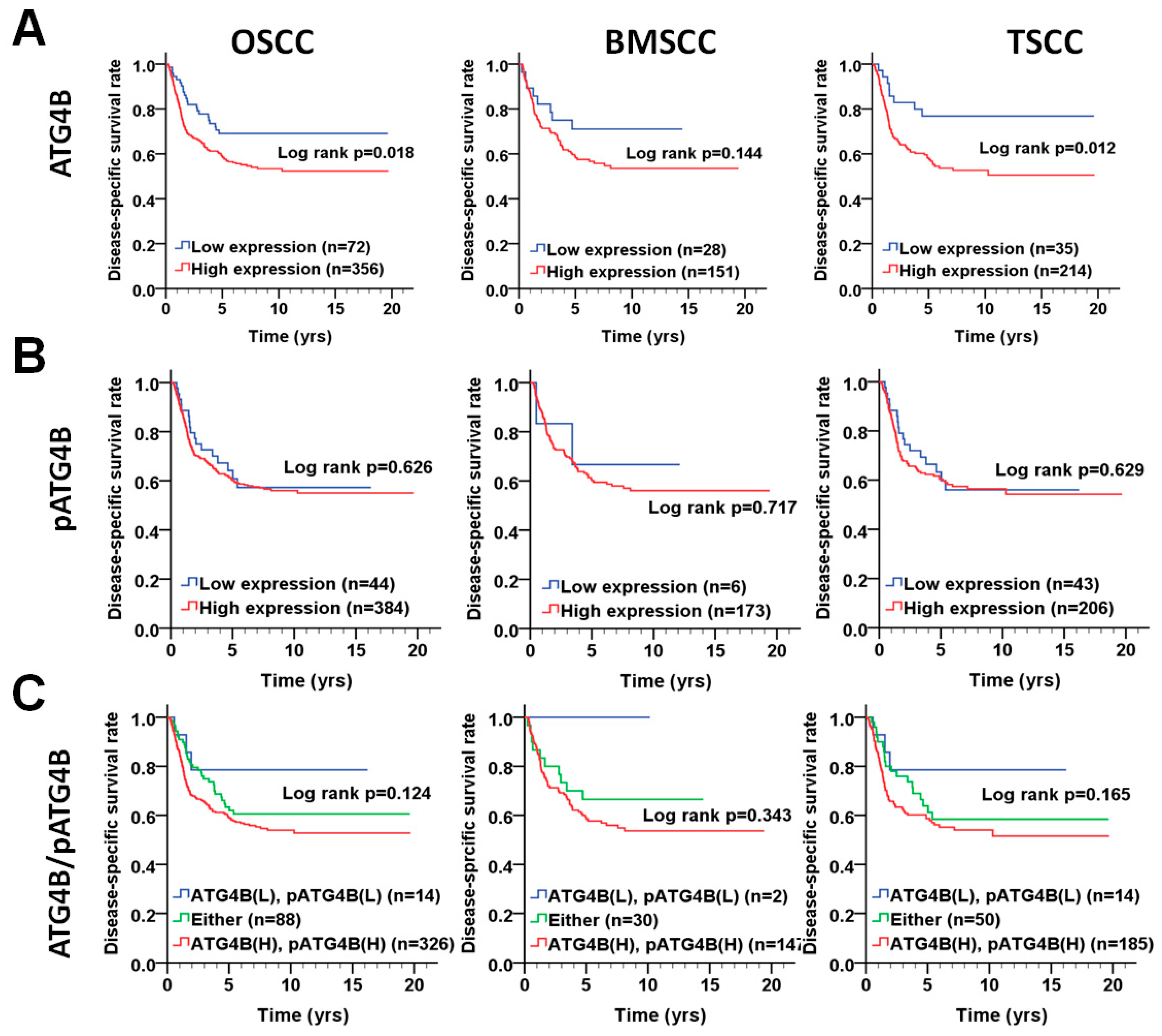
3.2. Protein Levels of ATG4B and Phospho-Ser383/392-ATG4B and DSS of OSCC Patients
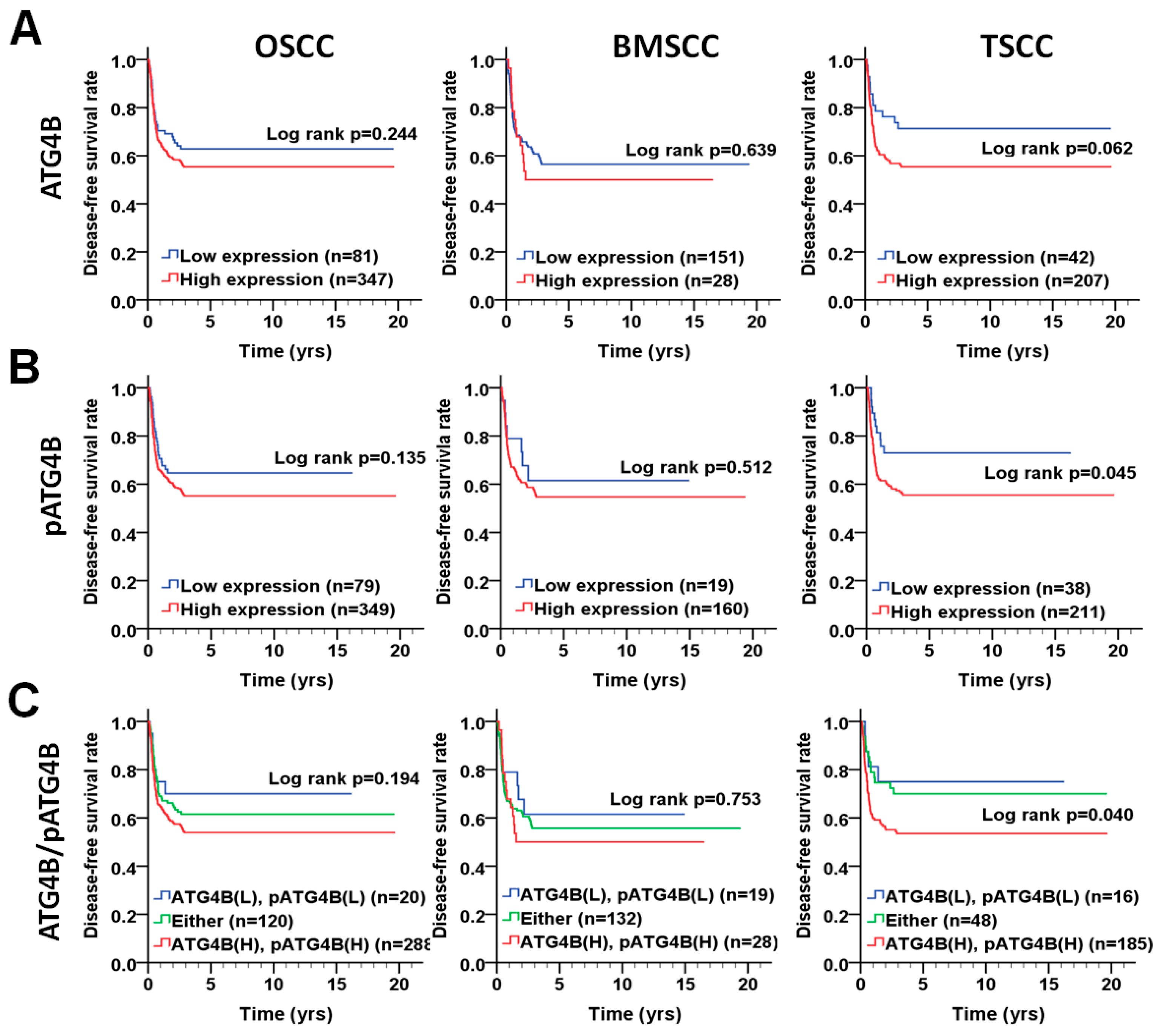
3.3. Association of the ATG4B and Phospho-Ser383/392-ATG4B Protein Levels with Disease-Free Survival in OSCC Patients
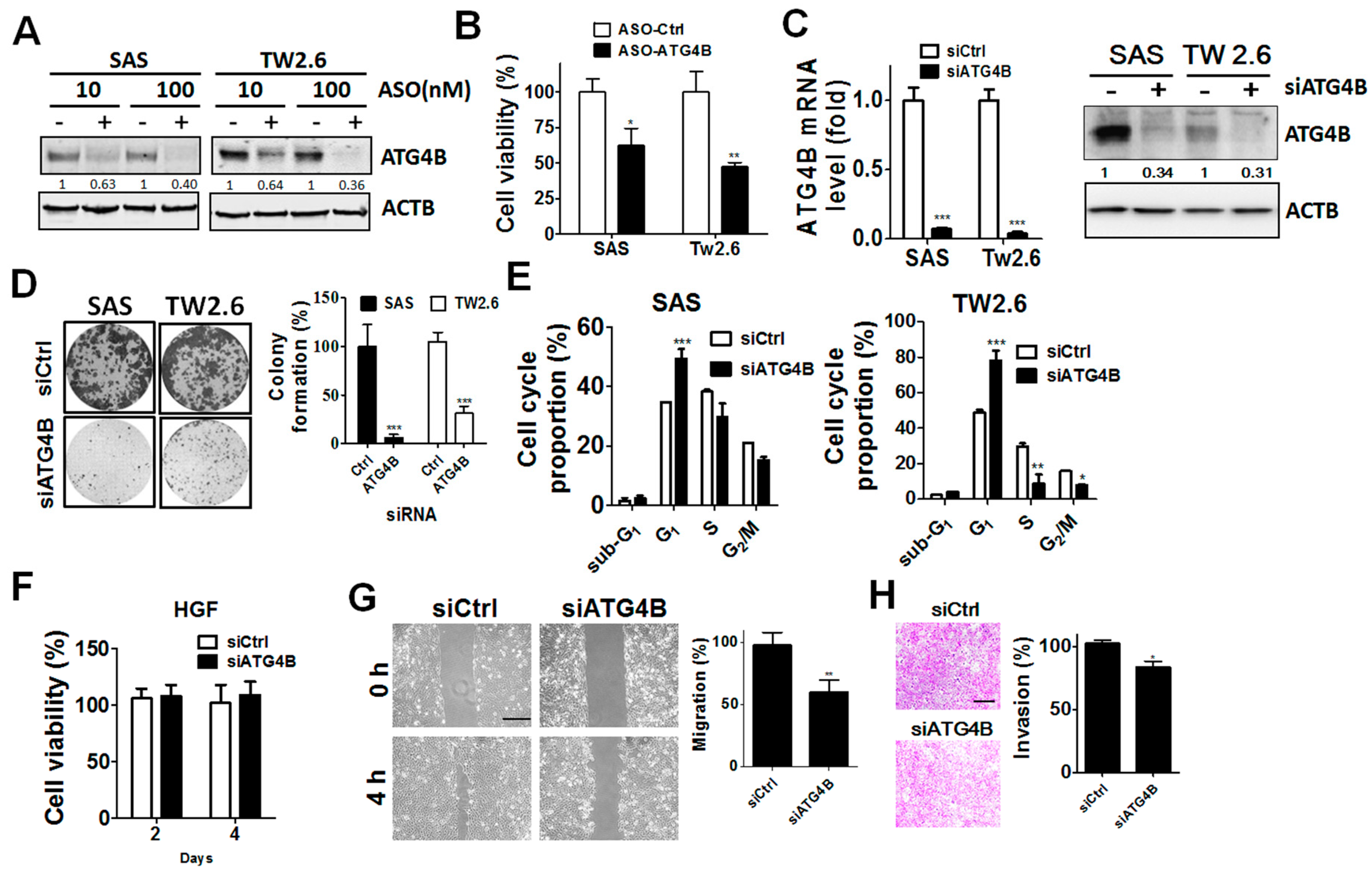
3.4. Function of ATG4B in Cell Proliferation and Invasion of OSCC Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | pATG4B | No. (%) | CHR (95% CI) | p Value * | AHR (95% CI) | p Value † |
|---|---|---|---|---|---|---|
| Sex | ||||||
| Female | Low | 5 (14.7) | 1.00 | 1.00 | ||
| High | 29 (85.3) | 1.43 (0.18–11.30) | 0.735 | 1.09 (0.13–8.98) | 0.939 a | |
| Male | Low | 39 (9.9) | 1.00 | 1.00 | ||
| High | 355 (90.1) | 1.11 (0.66–1.85) | 0.699 | 1.09 (0.65–1.83) | 0.745 a | |
| Age, yrs | ||||||
| ≤50 | Low | 22 (10.9) | 1.00 | 1.00 | ||
| High | 180 (89.1) | 0.94 (0.49–1.81) | 0.845 | 0.90 (0.47–1.74) | 0.751 a | |
| >50 | Low | 22 (9.7) | 1.00 | 1.00 | ||
| High | 204 (90.3) | 1.38 (0.64–2.98) | 0.417 | 1.27 (0.59–2.76) | 0.544 a | |
| Cell differentiation | ||||||
| Well | Low | 5 (6.6) | 1.00 | 1.00 | ||
| High | 71 (93.4) | 1.16 (0.15–8.74) | 0.887 | 1.88 (0.24–14.68) | 0.548 b | |
| Moderate, poor | Low | 39 (11.1) | 1.00 | 1.00 | ||
| High | 313 (88.9) | 1.21 (0.72–2.03) | 0.466 | 1.05 (0.63–1.77) | 0.846 b | |
| AJCC pathological stage | ||||||
| I, II | Low | 31 (11.1) | 1.00 | 1.00 | ||
| High | 248 (88.9) | 1.33 (0.61–2.89) | 0.471 | 1.48 (0.68–3.23) | 0.321 c | |
| III, IV | Low | 13 (8.7) | 1.00 | 1.00 | ||
| High | 136 (91.3) | 0.88 (0.46–1.70) | 0.705 | 0.83 (0.43–1.60) | 0.580 c | |
| T classification | ||||||
| T1, T2 | Low | 35 (10.5) | 1.00 | 1.00 | ||
| High | 297 (89.5) | 1.18 (0.64–2.20) | 0.600 | 1.11 (0.60–2.08) | 0.737 d | |
| T3, T4 | Low | 9 (9.4) | 1.00 | 1.00 | ||
| High | 87 (90.6) | 1.03 (0.44–2.40) | 0.944 | 0.80 (0.33–1.90) | 0.609 d | |
| N classification | ||||||
| N0 | Low | 38 (11.5) | 1.00 | 1.00 | ||
| High | 293 (88.5) | 1.04 (0.57–1.90) | 0.897 | 1.17 (0.64–2.13) | 0.615 e | |
| N1, N2 | Low | 6 (6.2) | 1.00 | 1.00 | ||
| High | 91 (93.8) | 0.83 (0.33–2.06) | 0.685 | 0.70 (0.27–1.80) | 0.460 e | |
| Postoperative RT | ||||||
| No | Low | 30 (9.6) | 1.00 | 1.00 | ||
| High | 281 (90.4) | 1.20 (0.61–2.37) | 0.603 | 1.07 (0.54–2.13) | 0.846 a | |
| Yes | Low | 14 (12.0) | 1.00 | 1.00 | ||
| High | 103 (88.0) | 1.23 (0.59–2.57) | 0.584 | 1.19 (0.57–2.49) | 0.651 a |
| Variable | ATG4B | No. (%) | CHR (95% CI) | p Value * | AHR (95% CI) | p Value † |
|---|---|---|---|---|---|---|
| Sex | ||||||
| Female | Low | 7 (20.6) | 1.00 | 1.00 | ||
| High | 27 (79.4) | 31.66 (0.05–20701.19) | 0.296 | 188230.91 (0.00–1.249E+286) | 0.971 a | |
| Male | Low | 74 (18.8) | 1.00 | 1.00 | ||
| High | 320 (81.2) | 1.15 (0.78–1.71) | 0.484 | 1.13 (0.76–1.67) | 0.555 a | |
| Age, yrs | ||||||
| ≤50 | Low | 33 (16.3) | 1.00 | 1.00 | ||
| High | 169 (83.7) | 1.13 (0.63–2.05) | 0.588 | 0.94 (0.51–1.73) | 0.838 a | |
| >50 | Low | 48 (21.2) | 1.00 | 1.00 | ||
| High | 178 (78.8) | 1.39 (0.82–2.35) | 0.221 | 1.57 (0.92–2.66) | 0.096 a | |
| Cell differentiation | ||||||
| Well | Low | 19 (25.0) | 1.00 | 1.00 | ||
| High | 57 (75.0) | 0.93 (0.34–2.59) | 0.893 | 0.94 (0.34–2.60) | 0.898 b | |
| Moderate, poor | Low | 62 (17.6) | 1.00 | 1.00 | ||
| High | 290 (82.4) | 1.26 (0.82–1.93) | 0.290 | 1.27 (0.83–1.94) | 0.282 b | |
| AJCC pathological stage | ||||||
| I, II | Low | 53 (19.0) | 1.00 | 1.00 | ||
| High | 226 (81.0) | 1.37 (0.82–2.30) | 0.235 | 1.32 (0.79–2.22) | 0.289 c | |
| III, IV | Low | 28 (18.8) | 1.00 | 1.00 | ||
| High | 121 (81.2) | 1.12 (0.61–2.05) | 0.712 | 1.07 (0.58–1.96) | 0.836 c | |
| T classification | ||||||
| T1, T2 | Low | 60 (18.1) | 1.00 | 1.00 | ||
| High | 272 (81.9) | 1.25 (0.80–1.98) | 0.330 | 1.16 (0.74–1.84) | 0.518 d | |
| T3, T4 | Low | 21 (21.9) | 1.00 | 1.00 | ||
| High | 75 (78.1) | 1.29 (0.59–2.81) | 0.519 | 1.13 (0.51–2.50) | 0.755 d | |
| N classification | ||||||
| N0 | Low | 68 (20.5) | 1.00 | 1.00 | ||
| High | 263 (79.5) | 1.33 (0.84–2.11) | 0.226 | 1.30 (0.82–2.06) | 0.266 | |
| N1, N2 | Low | 13 (13.4) | 1.00 | 1.00 | ||
| High | 84 (86.6) | 0.88 (0.42–1.88) | 0.748 | 0.78 (0.37–1.67) | 0.524 e | |
| Postoperative RT | ||||||
| No | Low | 55 (17.7) | 1.00 | 1.00 | ||
| High | 256 (82.3) | 1.43 (0.87–2.36) | 0.159 | 1.44 (0.88–2.38) | 0.150 a | |
| Yes | Low | 26 (22.2) | 1.00 | 1.00 | ||
| High | 91 (77.8) | 1.02 (0.53–1.96) | 0.947 | 1.06 (0.55–2.07) | 0.857 a |
References
- Rivera, C.; Oliveira, A.K.; Costa, R.A.P.; De Rossi, T.; Paes Leme, A.F. Prognostic biomarkers in oral squamous cell carcinoma: A systematic review. Oral Oncol. 2017, 72, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Kumar, G.K.; Abidullah, M.; Elbadawi, L.; Dakhil, S.; Mawardi, H. Epidemiological profile and clinical characteristics of oral potentially malignant disorders and oral squamous cell carcinoma: A pilot study in Bidar and Gulbarga Districts, Karnataka, India. J. Oral Maxillofac. Pathol. 2019, 23, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Adhauliya, N.; Kalappanavar, A.N.; Ali, I.M.; Annigeri, R.G. Autophagy: A boon or bane in oral cancer. Oral Oncol. 2016, 61, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Chiang, C.J.; Lo, W.C.; Yang, Y.W.; You, S.L.; Chen, C.J.; Lai, M.S. Incidence and survival of adult cancer patients in Taiwan, 2002–2012. J. Formos. Med. Assoc. 2016, 115, 1076–1088. [Google Scholar] [CrossRef] [PubMed]
- Dikic, I.; Elazar, Z. Mechanism and medical implications of mammalian autophagy. Nat. Rev. Mol. Cell Biol. 2018, 19, 349–364. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.F.; Chang, H.W.; Cheng, J.S.; Lee, H.P.; Yen, C.Y.; Tsai, W.L.; Cheng, J.T.; Li, Y.J.; Huang, W.C.; Lee, C.H.; et al. Map1lc3b and Sqstm1 Modulated Autophagy for Tumorigenesis and Prognosis in Certain Subsites of Oral Squamous Cell Carcinoma. J. Clin. Med. 2018, 7, 478. [Google Scholar] [CrossRef]
- Ojha, R.; Bhattacharyya, S.; Singh, S.K. Autophagy in Cancer Stem Cells: A Potential Link Between Chemoresistance, Recurrence, and Metastasis. Biores. Open Access 2015, 4, 97–108. [Google Scholar] [CrossRef]
- Zhang, D.; Zhao, Q.; Sun, H.; Yin, L.; Wu, J.; Xu, J.; He, T.; Yang, C.; Liang, C. Defective autophagy leads to the suppression of stem-like features of CD271+ osteosarcoma cells. J. Biomed. Sci. 2016, 23, e82. [Google Scholar] [CrossRef]
- Fu, Y.; Hong, L.; Xu, J.; Zhong, G.; Gu, Q.; Gu, Q.; Guan, Y.; Zheng, X.; Dai, Q.; Luo, X.; et al. Discovery of a small molecule targeting autophagy via ATG4B inhibition and cell death of colorectal cancer cells in vitro and in vivo. Autophagy 2019, 15, 295–311. [Google Scholar] [CrossRef]
- Shu, C.W.; Drag, M.; Bekes, M.; Zhai, D.; Salvesen, G.S.; Reed, J.C. Synthetic substrates for measuring activity of autophagy proteases: autophagins (Atg4). Autophagy 2010, 6, 936–947. [Google Scholar] [CrossRef]
- Liu, P.F.; Leung, C.M.; Chang, Y.H.; Cheng, J.S.; Chen, J.J.; Weng, C.J.; Tsai, K.W.; Hsu, C.J.; Liu, Y.C.; Hsu, P.C.; et al. ATG4B promotes colorectal cancer growth independent of autophagic flux. Autophagy 2014, 10, 1454–1465. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.F.; Hsu, C.J.; Tsai, W.L.; Cheng, J.S.; Chen, J.J.; Huang, I.F.; Tseng, H.H.; Chang, H.W.; Shu, C.W. Ablation of ATG4B Suppressed Autophagy and Activated AMPK for Cell Cycle Arrest in Cancer Cells. Cell. Physiol. Biochem. 2017, 44, 728–740. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.F.; Tsai, K.L.; Hsu, C.J.; Tsai, W.L.; Cheng, J.S.; Chang, H.W.; Shiau, C.W.; Goan, Y.G.; Tseng, H.H.; Wu, C.H.; et al. Drug Repurposing Screening Identifies Tioconazole as an ATG4 Inhibitor that Suppresses Autophagy and Sensitizes Cancer Cells to Chemotherapy. Theranostics 2018, 8, 830–845. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.W.; Liu, P.F.; Tsai, W.L.; Hu, W.H.; Hu, Y.C.; Yang, H.C.; Lin, W.Y.; Weng, J.R.; Shu, C.W. Xanthium strumarium Fruit Extract Inhibits ATG4B and Diminishes the Proliferation and Metastatic Characteristics of Colorectal Cancer Cells. Toxins 2019, 11, 313. [Google Scholar] [CrossRef] [PubMed]
- Rothe, K.; Lin, H.; Lin, K.B.; Leung, A.; Wang, H.M.; Malekesmaeili, M.; Brinkman, R.R.; Forrest, D.L.; Gorski, S.M.; Jiang, X. The core autophagy protein ATG4B is a potential biomarker and therapeutic target in CML stem/progenitor cells. Blood 2014, 123, 3622–3634. [Google Scholar] [CrossRef] [PubMed]
- Bortnik, S.; Choutka, C.; Horlings, H.M.; Leung, S.; Baker, J.H.; Lebovitz, C.; Dragowska, W.H.; Go, N.E.; Bally, M.B.; Minchinton, A.I.; et al. Identification of breast cancer cell subtypes sensitive to ATG4B inhibition. Oncotarget 2016, 7, 66970–66988. [Google Scholar] [CrossRef] [PubMed]
- Toshima, T.; Shirabe, K.; Matsumoto, Y.; Yoshiya, S.; Ikegami, T.; Yoshizumi, T.; Soejima, Y.; Ikeda, T.; Maehara, Y. Autophagy enhances hepatocellular carcinoma progression by activation of mitochondrial beta-oxidation. J. Gastroenterol. 2014, 49, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Wilkie-Grantham, R.P.; Yanagi, T.; Shu, C.W.; Matsuzawa, S.; Reed, J.C. ATG4B (Autophagin-1) phosphorylation modulates autophagy. J. Biol. Chem. 2015, 290, 26549–26561. [Google Scholar] [CrossRef]
- Fu, T.Y.; Wu, C.N.; Sie, H.C.; Cheng, J.T.; Lin, Y.S.; Liou, H.H.; Tseng, Y.K.; Shu, C.W.; Tsai, K.W.; Yen, L.M.; et al. Subsite-specific association of DEAD box RNA helicase DDX60 with the development and prognosis of oral squamous cell carcinoma. Oncotarget 2016, 7, 85097–85108. [Google Scholar] [CrossRef]
- Amin, M.B.; Greene, F.L.; Edge, S.B.; Compton, C.C.; Gershenwald, J.E.; Brookland, R.K.; Meyer, L.; Gress, D.M.; Byrd, D.R.; Winchester, D.P. The Eighth Edition AJCC Cancer Staging Manual: Continuing to build a bridge from a population-based to a more “personalized” approach to cancer staging. CA Cancer J. Clin. 2017, 67, 93–99. [Google Scholar] [CrossRef]
- Goan, Y.G.; Liu, P.F.; Chang, H.W.; Chen, H.C.; Chen, W.C.; Kuo, S.M.; Lee, C.H.; Shu, C.W. Kinome-Wide Screening with Small Interfering RNA Identified Polo-like Kinase 1 as a Key Regulator of Proliferation in Oral Cancer Cells. Cancers 2019, 11, 1117. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.S.; Tsai, W.L.; Liu, P.F.; Goan, Y.G.; Lin, C.W.; Tseng, H.H.; Lee, C.H.; Shu, C.W. The MAP3K7-mTOR Axis Promotes the Proliferation and Malignancy of Hepatocellular Carcinoma Cells. Front. Oncol. 2019, 9, e474. [Google Scholar] [CrossRef] [PubMed]
- Pengo, N.; Agrotis, A.; Prak, K.; Jones, J.; Ketteler, R. A reversible phospho-switch mediated by ULK1 regulates the activity of autophagy protease ATG4B. Nat. Commun. 2017, 8, e294. [Google Scholar] [CrossRef]
- Sheu, S.J.; Chen, J.L.; Bee, Y.S.; Lin, S.H.; Shu, C.W. ERBB2-modulated ATG4B and autophagic cell death in human ARPE19 during oxidative stress. PLoS ONE 2019, 14, e0213932. [Google Scholar] [CrossRef] [PubMed]
- Ni, Z.; He, J.; Wu, Y.; Hu, C.; Dai, X.; Yan, X.; Li, B.; Li, X.; Xiong, H.; Li, Y.; et al. AKT-mediated phosphorylation of ATG4B impairs mitochondrial activity and enhances the Warburg effect in hepatocellular carcinoma cells. Autophagy 2018, 14, 685–701. [Google Scholar] [CrossRef]
- Huang, T.; Kim, C.K.; Alvarez, A.A.; Pangeni, R.P.; Wan, X.; Song, X.; Shi, T.; Yang, Y.; Sastry, N.; Horbinski, C.M.; et al. MST4 Phosphorylation of ATG4B Regulates Autophagic Activity, Tumorigenicity, and Radioresistance in Glioblastoma. Cancer Cell 2017, 32, 840–855. [Google Scholar] [CrossRef]
- Fu, Y.; Huang, Z.; Hong, L.; Lu, J.H.; Feng, D.; Yin, X.M.; Li, M. Targeting ATG4 in Cancer Therapy. Cancers 2019, 11, 649. [Google Scholar] [CrossRef]
- Qiao, S.; Dennis, M.; Song, X.; Vadysirisack, D.D.; Salunke, D.; Nash, Z.; Yang, Z.; Liesa, M.; Yoshioka, J.; Matsuzawa, S.; et al. A REDD1/TXNIP pro-oxidant complex regulates ATG4B activity to control stress-induced autophagy and sustain exercise capacity. Nat. Commun. 2015, 6, e7014. [Google Scholar] [CrossRef]
- Chen, H.T.; Liu, H.; Mao, M.J.; Tan, Y.; Mo, X.Q.; Meng, X.J.; Cao, M.T.; Zhong, C.Y.; Liu, Y.; Shan, H.; et al. Crosstalk between autophagy and epithelial-mesenchymal transition and its application in cancer therapy. Mol. Cancer 2019, 18, e101. [Google Scholar] [CrossRef] [Green Version]





| Variables | No. | Tumor-Adjacent Normal | Tumor | Z | p-Value* | ||
|---|---|---|---|---|---|---|---|
| Mean ± SD | Median | Mean ± SD | Median | ||||
| OSCC | |||||||
| ATG4B | 328 | 145.15 ± 70.90 | 160.00 | 165.75 ± 54.67 | 177.50 | 4.578 | <0.001 |
| pATG4B | 328 | 111.66 ± 84.17 | 85.00 | 168.65 ± 74.32 | 176.25 | 11.302 | <0.001 |
| BMSCC | |||||||
| ATG4B | 127 | 146.81 ± 73.35 | 160.00 | 166.02 ± 62.44 | 180.00 | 2.665 | 0.008 |
| pATG4B | 127 | 175.87 ± 73.63 | 180.00 | 230.85 ± 53.91 | 237.50 | 6.114 | <0.001 |
| TSCC | |||||||
| ATG4B | 201 | 144.10 ± 69.48 | 160.00 | 165.58 ± 49.29 | 175.00 | 3.781 | <0.001 |
| pATG4B | 201 | 71.09 ± 62.42 | 70.00 | 129.35 ± 56.48 | 122.50 | 9.291 | <0.001 |
| Variable | % | OSCC (n = 428) | % | BMSCC (n = 179) | % | TSCC (n = 249) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean ± SD | Median | p Value | Mean ± SD | Median | p Value | Mean ± SD | Median | p Value | ||||
| Sex | ||||||||||||
| Female | 7.9 | 170.76 ± 49.89 | 190.00 | 0.642 * | 2.2 | 179.38 ± 32.04 | 192.50 | 0.662 * | 12.0 | 169.61 ± 52.10 | 190.00 | 0.744 * |
| Male | 92.1 | 166.27 ± 54.25 | 180.00 | 97.8 | 165.99 ± 60.84 | 180.00 | 88.0 | 166.50 ± 48.49 | 175.00 | |||
| Age, y | ||||||||||||
| ≤40 | 15.4 | 165.61 ± 51.38 | 175.00 | 0.166 † | 13.4 | 183.44 ± 57.10 | 180.00 | 0.105 † | 16.9 | 155.42 ± 45.43 | 152.50 | 0.212 † |
| 41–50 | 31.8 | 165.19 ± 56.47 | 170.00 | 30.7 | 158.09 ± 64.01 | 150.00 | 32.5 | 170.01 ± 50.57 | 180.00 | |||
| 51–60 | 29.0 | 160.64 ± 52.92 | 180.00 | 30.2 | 156.76 ± 56.35 | 180.00 | 28.1 | 163.63 ± 50.33 | 177.50 | |||
| >60 | 23.8 | 176.50 ± 52.48 | 185.00 | 25.7 | 178.32 ± 59.83 | 185.00 | 22.5 | 175.00 ± 46.08 | 185.00 | |||
| Subsites | ||||||||||||
| Buccal | 41.8 | 166.28 ± 60.33 | 180.00 | 0.911 * | 100.0 | 166.28 ± 60.33 | 180.00 | |||||
| Tongue | 58.2 | 166.88 ± 48.84 | 180.00 | 100.0 | 166.88 ± 48.84 | 180.00 | ||||||
| Cell differentiation | ||||||||||||
| Well | 17.8 | 164.97 ± 61.05 | 177.50 | 0.558 † | 27.4 | 165.71 ± 63.12 | 180.00 | 0.988 † | 10.8 | 163.61 ± 58.24 | 175.00 | 0.287 † |
| Moderate | 76.2 | 166.15 ± 51.71 | 180.00 | 67.6 | 166.69 ± 60.39 | 180.00 | 82.3 | 165.82 ± 45.98 | 180.00 | |||
| Poor | 6.1 | 177.56 ± 59.18 | 185.00 | 5.0 | 163.89 ± 48.67 | 185.00 | 6.8 | 184.80 ± 64.24 | 195.00 | |||
| AJCC pathological stage | ||||||||||||
| I | 31.8 | 165.66 ± 52.14 | 180.00 | 0.803 † | 35.2 | 168.06 ± 54.54 | 180.00 | 0.806 † | 29.3 | 163.60 ± 50.26 | 180.00 | 0.567 † |
| II | 33.4 | 164.73 ± 52.08 | 175.00 | 26.3 | 165.64 ± 64.73 | 180.00 | 38.6 | 164.29 ± 45.00 | 170.00 | |||
| III | 15.2 | 172.51 ± 60.25 | 180.00 | 10.6 | 176.32 ± 67.70 | 180.00 | 18.5 | 170.94 ± 57.63 | 180.00 | |||
| IV | 19.6 | 166.88 ± 55.11 | 180.00 | 27.9 | 160.85 ± 61.40 | 177.50 | 13.7 | 175.74 ± 43.67 | 190.00 | |||
| T classification | ||||||||||||
| T1 | 33.9 | 164.98 ± 51.80 | 180.00 | 0.389 † | 38.0 | 165.22 ± 54.59 | 177.50 | 0.368 † | 30.9 | 164.77 ± 49.58 | 180.00 | 0.518 † |
| T2 | 43.7 | 171.19 ± 55.78 | 180.00 | 38.0 | 175.00 ± 66.21 | 185.00 | 47.8 | 169.02 ± 49.00 | 175.00 | |||
| T3 | 11.2 | 157.85 ± 46.59 | 175.00 | 6.7 | 154.79 ± 44.19 | 175.00 | 14.5 | 158.87 ± 47.93 | 167.50 | |||
| T4 | 11.2 | 162.60 ± 58.99 | 182.50 | 17.3 | 153.95 ± 63.65 | 170.00 | 6.8 | 178.38 ± 47.10 | 190.00 | |||
| N classification | ||||||||||||
| N0 | 77.3 | 163.40 ± 52.99 a | 175.00 | 0.022 † | 74.9 | 163.34 ± 60.07 | 177.50 | 0.321 † | 79.1 | 163.44 ± 47.74 | 175.00 | 0.049 † |
| N1 | 10.7 | 186.45 ± 60.89 a | 185.00 | 12.3 | 184.32 ± 66.05 | 185.00 | 9.6 | 188.40 ± 57.12 | 183.75 | |||
| N2 | 11.9 | 169.71 ± 49.80 | 180.00 | 12.8 | 166.20 ± 55.66 | 180.00 | 11.2 | 172.59 ± 45.26 | 187.50 | |||
| Variable | % | OSCC (n = 428) | % | BMSCC (n = 179) | % | TSCC (n = 249) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean ± SD | Median | p Value | Mean ± SD | Median | p Value | Mean ± SD | Median | p Value | ||||
| Sex | ||||||||||||
| Female | 7.9 | 135.83 ± 57.99 | 127.50 | 0.006 * | 2.2 | 227.50 ± 46.99 | 222.50 | 0.950 * | 12.0 | 123.61 ± 57.99 | 105.00 | 0.706 * |
| Male | 92.1 | 172.83 ± 74.95 | 180.00 | 97.8 | 229.21 ± 54.19 | 235.00 | 88.0 | 127.78 ± 56.49 | 125.00 | |||
| Age, y | ||||||||||||
| ≤40 | 15.4 | 159.37 ± 73.07 | 170.00 | 0.606 † | 13.4 | 220.00 ± 59.01 | 235.00 | 0.850 † | 16.9 | 124.72 ± 55.94 | 107.92 | 0.410 † |
| 41–50 | 31.8 | 174.34 ± 73.33 | 182.50 | 30.7 | 230.86 ± 52.98 | 237.50 | 32.5 | 135.96 ± 59.08 | 135.00 | |||
| 51–60 | 29.0 | 169.21 ± 76.90 | 175.00 | 30.2 | 230.60 ± 57.25 | 236.25 | 28.1 | 121.85 ± 52.91 | 130.00 | |||
| >60 | 23.8 | 171.61 ± 75.91 | 180.00 | 25.7 | 230.27 ± 49.43 | 232.50 | 22.5 | 123.42 ± 57.80 | 92.50 | |||
| Subsites | ||||||||||||
| Buccal | 41.8 | 229.18 ± 53.93 | 235.00 | <0.001 * | 100.0 | 229.18 ± 53.93 | 235.00 | |||||
| Tongue | 58.2 | 127.28 ± 56.57 | 122.50 | 100.0 | 127.28 ± 56.57 | 122.50 | ||||||
| Cell differentiation | ||||||||||||
| Well | 17.8 | 201.41 ± 73.80 ab | 195.00 | <0.001 † | 27.4 | 235.31 ± 53.89 | 245.00 | 0.065 † | 10.8 | 139.91 ± 65.24 | 155.00 | 0.443 † |
| Moderate | 76.2 | 164.06 ± 74.67 a | 171.67 | 67.6 | 229.63 ± 54.00 | 237.50 | 82.3 | 125.35 ± 55.78 | 118.33 | |||
| Poor | 6.1 | 150.93 ± 55.27 b | 163.33 | 5.0 | 189.72 ± 39.50 | 195.00 | 6.8 | 130.39 ± 51.95 | 120.00 | |||
| AJCC pathological stage | ||||||||||||
| I | 31.8 | 173.39 ± 80.05 | 180.00 | 0.043 † | 35.2 | 235.16 ± 55.89 | 242.50 | 0.190 † | 29.3 | 120.08 ± 55.73 | 90.00 | 0.246 § |
| II | 33.4 | 161.89 ± 71.14 | 170.00 | 26.3 | 236.17 ± 45.71 | 237.50 | 38.6 | 125.53 ± 49.87 | 121.25 | |||
| III | 15.2 | 157.81 ± 75.38 | 175.00 | 10.6 | 210.00 ± 61.10 | 195.00 | 18.5 | 136.25 ± 70.45 | 150.00 | |||
| IV | 19.6 | 187.20 ± 69.41 | 190.00 | 27.9 | 223.35 ± 54.74 | 227.50 | 13.7 | 135.51 ± 55.17 | 130.00 | |||
| T classification | ||||||||||||
| T1 | 33.9 | 172.83 ± 79.29 | 180.00 | 0.044 † | 38.0 | 231.76 ± 58.05 | 242.50 | 0.196 † | 30.9 | 120.79 ± 55.60 | 90.00 | 0.585 † |
| T2 | 43.7 | 166.91 ± 72.17 | 175.00 | 38.0 | 234.60 ± 47.46 | 232.50 | 47.8 | 128.23 ± 52.75 | 125.00 | |||
| T3 | 11.2 | 150.35 ± 74.88 | 170.00 | 6.7 | 200.42 ± 64.31 | 187.50 | 14.5 | 133.66 ± 71.30 | 127.50 | |||
| T4 | 11.2 | 192.19 ± 66.81 | 192.50 | 17.3 | 222.74 ± 52.21 | 230.00 | 6.8 | 136.47 ± 53.79 | 145.00 | |||
| N classification | ||||||||||||
| N0 | 77.3 | 167.84 ± 77.08 | 173.33 | 0.370 § | 74.9 | 232.56 ± 54.80 | 241.25 | 0.325 † | 79.1 | 123.82 ± 56.18 | 113.33 | 0.139 † |
| N1 | 10.7 | 179.37 ± 62.37 | 185.00 | 12.3 | 215.80 ± 46.20 | 197.50 | 9.6 | 145.97 ± 56.82 | 165.00 | |||
| N2 | 11.9 | 174.66 ± 70.75 | 185.00 | 12.8 | 221.28 ± 55.16 | 225.00 | 11.2 | 135.54 ± 57.20 | 130.00 | |||
| Variable | ATG4B | No. (%) | CHR (95% CI) | p Value * | AHR (95% CI) | p Value † |
|---|---|---|---|---|---|---|
| Sex | ||||||
| Female | Low | 7 (20.6) | 1.00 | 1.00 | ||
| High | 27 (79.4) | 33.44 (0.08–13883.49) | 0.254 | 290102.55 (0.00–7.435E + 285) | 0.970 a | |
| Male | Low | 65 (16.5) | 1.00 | 1.00 | ||
| High | 329 (83.5) | 1.52 (0.97–2.38) | 0.067 | 1.74 (1.11–2.73) | 0.016 a | |
| Age, yrs | ||||||
| ≤50 | Low | 32 (15.8) | 1.00 | 1.00 | ||
| High | 170 (84.2) | 2.10 (1.05–4.19) | 0.035 | 1.57 (0.78–3.18) | 0.209 a | |
| >50 | Low | 40 (17.7) | 1.00 | 1.00 | ||
| High | 186 (82.3) | 1.43 (0.79–2.57) | 0.236 | 2.15 (1.17–3.92) | 0.013 a | |
| Cell differentiation | ||||||
| Well | Low | 16 (21.1) | 1.00 | 1.00 | ||
| High | 60 (78.9) | 0.97 (0.32–2.99) | 0.956 | 1.07 (0.34–3.30) | 0.912 b | |
| Moderate, poor | Low | 56 (15.9) | 1.00 | 1.00 | ||
| High | 296 (84.1) | 1.83 (1.12–3.00) | 0.015 | 2.05 (1.26–3.36) | 0.004 b | |
| AJCC pathological stage | ||||||
| I, II | Low | 47 (16.8) | 1.00 | 1.00 | ||
| High | 232 (83.2) | 1.41 (0.74–2.66) | 0.294 | 1.37 (0.72–2.59) | 0.335 c | |
| III, IV | Low | 25 (16.8) | 1.00 | 1.00 | ||
| High | 124 (83.2) | 2.35 (1.25–4.40) | 0.008 | 2.34 (1.25–4.40) | 0.008 c | |
| T classification | ||||||
| T1, T2 | Low | 53 (16.0) | 1.00 | 1.00 | ||
| High | 279 (84.0) | 1.39 (0.82–2.36) | 0.219 | 1.26 (0.75–2.14) | 0.384 d | |
| T3, T4 | Low | 19 (19.8) | 1.00 | 1.00 | ||
| High | 77 (80.2) | 3.21 (1.37–7.51) | 0.007 | 3.10 (1.31–7.31) | 0.010 d | |
| N classification | ||||||
| N0 | Low | 61 (18.4) | 1.00 | 1.00 | ||
| High | 270 (81.6) | 1.44 (0.85–2.45) | 0.179 | 1.68 (0.98–2.88) | 0.060 e | |
| N1, N2 | Low | 11 (11.3) | 1.00 | 1.00 | ||
| High | 86 (88.7) | 2.07 (0.89–4.79) | 0.090 | 1.95 (0.83–4.59) | 0.124 e | |
| Postoperative RT | ||||||
| No | Low | 49 (15.8) | 1.00 | 1.00 | ||
| High | 262 (84.2) | 1.98 (1.06–3.70) | 0.032 | 2.55 (1.36–4.79) | 0.004 a | |
| Yes | Low | 23 (19.7) | 1.00 | 1.00 | ||
| High | 94 (80.3) | 1.62 (0.85–3.10) | 0.143 | 1.43 (0.74–2.75) | 0.283 a |
| Variable | No. (%) | CHR (95% CI) | p-Value | AHR (95% CI) | p-Value | |
|---|---|---|---|---|---|---|
| OSCC | ||||||
| ATG4B | Low | 72 (16.8) | 1.00 | 1.00 | ||
| High | 356 (83.2) | 1.70(1.09–2.66) | 0.020 | 1.87(1.20–2.94) | 0.006 * | |
| Phospho-ATG4B | Low | 44 (10.3) | 1.00 | 1.00 | ||
| High | 384 (89.7) | 1.13(0.69–1.87) | 0.626 | 1.08(0.66–1.79) | 0.752 * | |
| ATG4B (L), pATG4B (L) | 14 (3.3) | 1 | 1 | |||
| Either | 88 (20.6) | 0.79 (0.54–1.16) | 0.224 | 1.89 (0.58–6.18) | 0.290 † | |
| ATG4B (H), pATG4B (H) | 326 (76.2) | 1.40 (0.97–2.02) | 0.072 | 2.47 (0.79–7.75) | 0.122 † | |
| BMSCC | ||||||
| ATG4B | Low | 28 (15.6) | 1.00 | 1.00 | ||
| High | 151 (84.4) | 1.72(0.82–3.57) | 0.149 | 2.70(1.28–5.68) | 0.009 * | |
| Phospho-ATG4B | Low | 6 (3.4) | 1.00 | 1.00 | ||
| High | 173 (96.6) | 1.30(0.32–5.28) | 0.717 | 1.31(0.32–5.34) | 0.708 * | |
| ATG4B (L), pATG4B (L) | 2 (1.1) | 1 | 1 | |||
| Either | 30 (16.8) | 0.72 (0.37–1.39) | 0.324 | 6201.44 (0.00–1.003E + 080) | 0.922 † | |
| ATG4B (H), pATG4B (H) | 147 (82.1) | 1.53 (0.79–2.98) | 0.210 | 8815.48 (0.00–1.425E + 080) | 0.919 † | |
| TSCC | ||||||
| ATG4B | Low | 35 (14.1) | 1.00 | 1.00 | ||
| High | 214 (85.9) | 2.46(1.19–5.06) | 0.015 | 2.19(1.06–4.52) | 0.034 * | |
| Phospho-ATG4B | Low | 43 (17.3) | 1.00 | 1.00 | ||
| High | 206 (82.7) | 1.14(0.68–1.91) | 0.630 | 1.07(0.64–1.81) | 0.788 * | |
| ATG4B (L), pATG4B (L) | 14 (5.6) | 1 | 1 | |||
| Either | 50 (20.1) | 0.81 (0.49–1.33) | 0.401 | 1.96 (0.58–6.65) | 0.278 † | |
| ATG4B (H), pATG4B (H) | 185 (74.3) | 1.48 (0.92–2.37) | 0.104 | 2.57 (0.81–8.15) | 0.109 † | |
| Variable | pATG4B | No. (%) | CHR (95% CI) | p Value * | AHR (95% CI) | p Value † |
|---|---|---|---|---|---|---|
| Sex | ||||||
| Female | Low | 12 (35.3) | 1.00 | 1.00 | ||
| High | 22 (64.7) | 1.12 (0.28–4.47) | 0.874 | 0.88 (0.21–3.58) | 0.852 a | |
| Male | Low | 67 (17.0) | 1.00 | 1.00 | ||
| High | 327 (83.0) | 1.33 (0.86–2.06) | 0.206 | 1.44 (0.93–2.24) | 0.105 a | |
| Age, yrs | ||||||
| ≤50 | Low | 38 (18.8) | 1.00 | 1.00 | ||
| High | 164 (81.2) | 1.63 (0.87–3.09) | 0.130 | 1.74 (0.92–3.29) | 0.089 a | |
| >50 | Low | 41 (18.1) | 1.00 | 1.00 | ||
| High | 185 (81.9) | 1.18 (0.68–2.04) | 0.567 | 1.22 (0.70–2.13) | 0.477 a | |
| Cell differentiation | ||||||
| Well | Low | 7 (9.2) | 1.00 | 1.00 | ||
| High | 69 (90.8) | 23.92 (0.04–14170.39) | 0.330 | 540189.98 (0.00) | 0.978 b | |
| Moderate, poor | Low | 72 (20.5) | 1.00 | 1.00 | ||
| High | 280 (79.5) | 1.40 (0.92–2.13) | 0.119 | 1.36 (0.89–2.07) | 0.157 b | |
| AJCC pathological stage | ||||||
| I, II | Low | 57 (20.7) | 1.00 | 1.00 | ||
| High | 222 (79.6) | 1.04 (0.65–1.68) | 0.863 | 1.15 (0.71–1.87) | 0.558 c | |
| III, IV | Low | 22 (14.8) | 1.00 | 1.00 | ||
| High | 127 (85.2) | 2.55 (1.03–6.34) | 0.044 | 2.59 (1.04–6.45) | 0.040 c | |
| T classification | ||||||
| T1, T2 | Low | 64 (19.3) | 1.00 | 1.00 | ||
| High | 268 (80.7) | 1.21 (0.77–1.90) | 0.402 | 1.30 (0.83–2.04) | 0.260 d | |
| T3, T4 | Low | 15 (15.6) | 1.00 | 1.00 | ||
| High | 81 (84.4) | 2.49 (0.77–8.07) | 0.129 | 2.29 (0.70–7.47) | 0.170 d | |
| N classification | ||||||
| N0 | Low | 67 (20.2) | 1.00 | 1.00 | ||
| High | 264 (79.8) | 1.13 (0.72–1.77) | 0.605 | 1.24 (0.79–1.95) | 0.354 e | |
| N1, N2 | Low | 12 (12.4) | 1.00 | 1.00 | ||
| High | 85 (87.6) | 2.69 (0.84–8.64) | 0.097 | 2.59 (0.80–8.32) | 0.112 e | |
| Postoperative RT | ||||||
| No | Low | 57 (18.3) | 1.00 | 1.00 | ||
| High | 254 (81.7) | 1.17 (0.73–1.86) | 0.517 | 1.18 (0.74–1.90) | 0.488 a | |
| Yes | Low | 22 (18.8) | 1.00 | 1.00 | ||
| High | 95 (81.2) | 2.24 (0.89–5.65) | 0.088 | 2.51 (0.99–6.34) | 0.053 a |
| Variable | No. (%) | CHR (95% CI) | p-Value | AHR (95% CI) | p-Value | |
|---|---|---|---|---|---|---|
| OSCC | ||||||
| ATG4B | Low | 81 (18.9) | 1.00 | 1.00 | ||
| High | 347 (81.1) | 1.26(0.85–1.87) | 0.245 | 1.22(0.82–1.81) | 0.319 * | |
| Phospho-ATG4B | Low | 79 (18.5) | 1.00 | 1.00 | ||
| High | 349 (81.5) | 1.37(0.90–2.08) | 0.137 | 1.45(0.96–2.21) | 0.080 * | |
| ATG4B (L), pATG4B (L) | 20 (4.7) | 1 | 1 | |||
| Either | 120 (28.0) | 0.80 (0.57–1.13) | 0.119 | 1.29 (0.55–3.03) | 0.555 † | |
| ATG4B (H), pATG4B (H) | 288 (67.3) | 1.33 (0.96–1.85) | 0.085 | 1.67 (0.73–3.78) | 0.222 † | |
| BMSCC | ||||||
| ATG4B | Low | 151 (84.4) | 1.00 | 1.00 | ||
| High | 28 (15.6) | 1.15(0.64–2.05) | 0.640 | 1.19(0.66–2.13) | 0.563 * | |
| Phospho-ATG4B | Low | 19 (10.6) | 1.00 | 1.00 | ||
| High | 160 (89.4) | 1.30(0.60–2.82) | 0.514 | 1.38(0.63–3.00) | 0.422 * | |
| ATG4B (L), pATG4B (L) | 19 (10.6) | 1 | 1 | |||
| Either | 132 (73.7) | 1.02 (0.62–1.68) | 0.936 | 1.27 (0.58–2.78) | 0.552 † | |
| ATG4B (H), pATG4B (H) | 28 (15.6) | 1.15 (0.64–2.05) | 0.640 | 1.42 (0.57–3.51) | 0.453 † | |
| TSCC | ||||||
| ATG4B | Low | 42 (16.9) | 1.00 | 1.00 | ||
| High | 207 (83.1) | 1.77(0.97–3.23) | 0.065 | 1.68(0.92–3.08) | 0.095 * | |
| Phospho-ATG4B | Low | 38 (15.3) | 1.00 | 1.00 | ||
| High | 211 (84.7) | 1.93(1.00–3.71) | 0.049 | 1.95(1.02–3.76) | 0.045 * | |
| ATG4B (L), pATG4B (L) | 16 (6.4) | 1 | 1 | |||
| Ether | 48 (19.3) | 0.58 (0.33–1.02) | 0.059 | 1.21 (0.40–3.66) | 0.741 † | |
| ATG4B (H), pATG4B (H) | 185 (74.3) | 1.91 (1.15–3.19) | 0.013 | 2.20 (0.81–6.01) | 0.123 † | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, P.-F.; Chen, H.-C.; Cheng, J.-S.; Tsai, W.-L.; Lee, H.-P.; Wang, S.-C.; Peng, W.-H.; Lee, C.-H.; Ger, L.-P.; Shu, C.-W. Association of ATG4B and Phosphorylated ATG4B Proteins with Tumorigenesis and Prognosis in Oral Squamous Cell Carcinoma. Cancers 2019, 11, 1854. https://doi.org/10.3390/cancers11121854
Liu P-F, Chen H-C, Cheng J-S, Tsai W-L, Lee H-P, Wang S-C, Peng W-H, Lee C-H, Ger L-P, Shu C-W. Association of ATG4B and Phosphorylated ATG4B Proteins with Tumorigenesis and Prognosis in Oral Squamous Cell Carcinoma. Cancers. 2019; 11(12):1854. https://doi.org/10.3390/cancers11121854
Chicago/Turabian StyleLiu, Pei-Feng, Hung-Chih Chen, Jin-Shiung Cheng, Wei-Lun Tsai, Huai-Pao Lee, Shu-Chi Wang, Wei-Hao Peng, Cheng-Hsin Lee, Luo-Ping Ger, and Chih-Wen Shu. 2019. "Association of ATG4B and Phosphorylated ATG4B Proteins with Tumorigenesis and Prognosis in Oral Squamous Cell Carcinoma" Cancers 11, no. 12: 1854. https://doi.org/10.3390/cancers11121854