Carbon Nanotubes under Scrutiny: Their Toxicity and Utility in Mesothelioma Research


Louvain Centre for Toxicology and Applied Pharmacology (LTAP), Institut de Recherche Experimentale et Clinique (IREC), Université catholique de Louvain (UCLouvain), 1200 Brussels, Belgium
*
Author to whom correspondence should be addressed.
Appl. Sci. 2020, 10(13), 4513; https://doi.org/10.3390/app10134513
Submission received: 10 May 2020
/
Revised: 20 June 2020
/
Accepted: 23 June 2020
/
Published: 29 June 2020
(This article belongs to the Special Issue State of the Art of Nanosciences in Belgium)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Research on the toxicity of engineered carbon nanotubes (CNT) was initiated by Belgian academic chemists and toxicologists more than 15 years ago. It is now undisputed that some of these attractive nanomaterials induce serious illness such as fibrosis and cancer. The physico-chemical determinants of CNT-induced adverse effects are now elucidated and include shape, nanoscale diameter, structural defects and scavenger capacity. Generated in vitro and in vivo data on their inflammogenic and fibrogenic activities were combined and translated in worldwide accepted AOP (Adverse Outcome Pathways) available for risk assessment and regulatory policies. The asbestos-like carcinogenic effects of CNT, notably their capacity to induce malignant mesothelioma (MM), remain a cause of concern for public health and strongly curb the craze for CNT in industries. MM still represents a real challenge for clinicians and a highly refractory cancer to existing therapeutic strategies. By comparing mesotheliomagenic CNT (needle-like CNT-N) to non mesotheliomagenic CNT (tangled-like CNT-T), our group generated a relevant animal model that highlights immune pathogenic pathways specifically associated to the carcinogenic process. Evidence indicate that only CNT-N possess the intrinsic capacity to induce a preferential, rapid and sustained accumulation of host immunosuppressive cells that subvert immune surveillance and suppress T lymphocyte anti-mesothelioma immunity. This new concept offers novel horizons for clinical management of mesothelioma and represents an additional tool for predicting mesotheliomagenic activity of newly elaborated CNT or nanoparticles.
1. Discovering the CNT
The architect of carbon nanotubes (CNT) is Sumio Iijima, a Japanese physicist awarded with the prestigious Benjamin Franklin Medal in 2002 “for the discovery and elucidation of the atomic structure and helical character of multi-wall and single-wall carbon nanotubes” [1]. The formation of “carbon needles” of few nanometres in diameter suggested that the manufacture of engineered carbon structures should be possible on very large scales. This technical possibility brought a real enthusiasm in chemistry and several academic teams completed this discovery by proposing complementary procedures to elaborate tailored CNT. Simultaneously, Professor Jànos B. Nagy and leading colleagues from the Université de Namur in Belgium described in Science an unsuspected extrusion of a carbon tubule from a catalytic particle [2].
These success stories had huge impact on the rapidly growing material and nanoscale science field. The physico-chemical characteristics (such as the very high thermal, electrical conductivity, tensile strength and stiffness) attracted the attention of scientists of different domains and the interest of many industrial sectors (mainly in electronics). The production of CNT then emerged from university confinement and was deployed on a larger scale in the early 2000s in spin-offs and newly-built industries as Nanocyl in Belgium. However, this craze was short-lived. The discovery of their toxicity (see below), the difficulty of large-scale synthesis and their high selling price explain slowdown in global demand for CNT during that disillusionment period.
However, industrial production of CNT is now experiencing a revival of interest. Plastic and battery suppliers recently assimilated the major interest of CNT and revived the market. Today, the management of the harmful effects of CNT is no longer be considered as an impassable barrier. Indeed, safe-by-design approaches now offer tailored CNT characterized by weak toxicity and limited health concerns [3,4]. Several recent reports mention that the worldwide market for CNT is estimated to grow from 5 billion U.S. dollars in 2018, to 10 billion by 2023 and 15 billion in 2026. During this forecast period, a CAGR (Compound annual growth rate) of 20% is expected. The Yano Research Institute Ltd. (Tokyo, Japan) released a report predicting a global CNT market of 4000 tons by 2023, representing a compound annual growth of 13% (2250 tons in 2018 when considering the global CNT market) [5].
Chemical vapor deposition (with or without catalysts) represents the main synthetic technique, and accounts for more than 70% of the global production. Currently, efficient production and declined prices for CNT make these nanomaterials more available and affordable [6]. Multi-walled CNT dominate the market and their increasing applications concern battery additives and plastic parts [7,8]. CNT also hold absorbent potential for organic wastewater treatment or emerging contaminant because of their high specific surface area and mesoporous structure [9,10,11]. The recent ability to functionalize CNT or control their growth and assembly [12] gives rise to the development of new biomedical applications, including tissue growth and drug delivery [3,13]. For instance, CNT-based materials permit the elaboration of layered cell architectures for embryonic stem cell and spheroid expansion during tissue regeneration [14,15].
2. From Asbestos to CNT: The Toxicological Aspect
2.1. Morphological Similitudes between CNT and Asbestos
Despite the scientific and industrial enthusiasm born from the discovery of CNT, several toxicologists including the Japanese Jun Kanno [16] rapidly discerned physical and morphological similarities between CNT and asbestos fibers. Indeed, these particles have a long needle shape (term already used by Iijima) and this morphology (longer than 10 µm) makes them foreign bodies difficult to be removed and therefore frustrating for the immune system. This intuitive approach led some toxicologists to test the detrimental effects of CNT in comparison to asbestos.
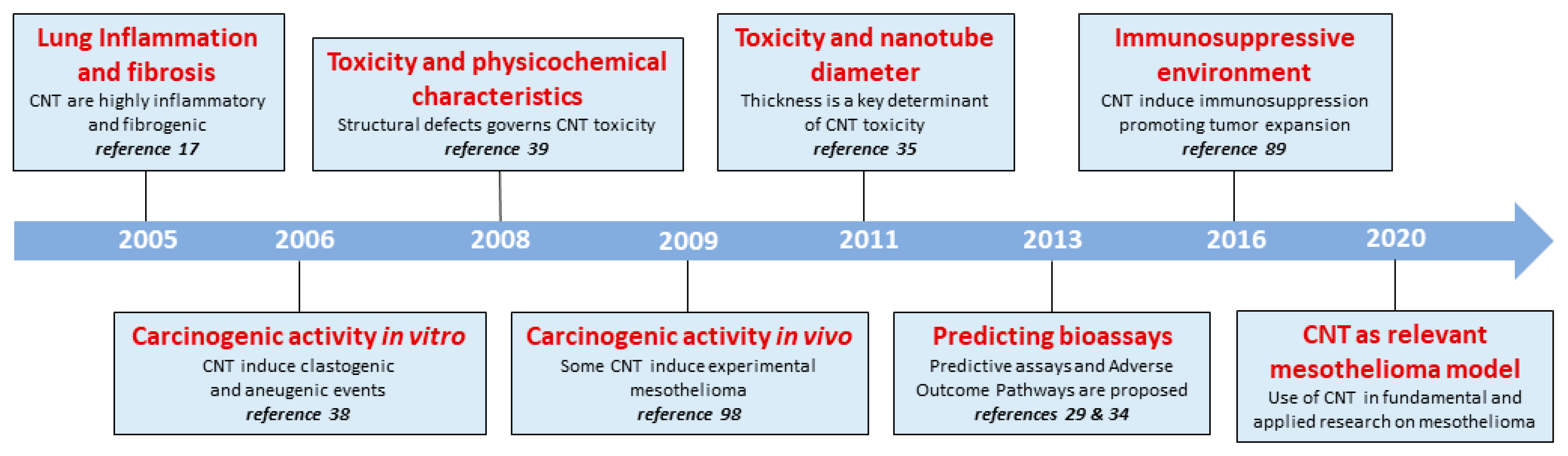
The prevailing idea prior to these experiments was that CNT (consisting predominantly of carbon) were perfectly biocompatible and non-toxic. However, several groups, including ours in 2005 [17] (Figure 1) rapidly reported that CNT injected into the lungs of animals were able to induce inflammatory, fibrogenic and carcinogenic responses [16,18,19,20,21]. These results, like many others thereafter, had a very significant global impact (papers widely cited; sometimes more than 1000 times) and initiated extensive in vitro and in vivo investigations focused on CNT toxicity [22]. Altogether, these findings demonstrated that the respiratory diseases induced by CNT (inflammation, fibrosis and cancer) correspond exactly to the pathologies observed after asbestos exposure [23].
2.2. Inflammogenic and Fibrogenic Effects of CNT: Mechanisms and Adverse Outcome Pathway (AOP)
Several decades were necessary for defining the exact pathogenesis of CNT-induced inflammation and fibrosis (Figure 1). A common scenario is now accepted and can be summarized as follows. Scavenger receptors deployed by sentinel macrophages induce CNT phagocytosis [24,25,26]. Internalized cytotoxic CNT destabilize phagolysosomes in macrophages and lysosomal contents released into their cytoplasm activate a sensing cytosolic complex (named inflammasome) which converts immature cytokines (pro IL–1β) into highly inflammatory mediators (active IL–1β). Our group also demonstrated that the permeabilization of lysosomes induces cell membranolysis and the subsequent release of danger signals (named alarmins) stored in macrophage cytoplasm [25,27,28]. Altogether, these inflammatory mediators orchestrate the persistent accumulation of neutrophils and macrophages. Oxidizing molecules produced by these inflammatory cells strongly damage surrounding tissues [24].
The inflammatory cycles are then followed by a reparative stage, where damaged tissues are renewed. This regeneration involves fibroblast activation by growth factors produced by activated macrophages [29,30]. We have also demonstrated that toxic CNT possess the unique ability to induce fibroblast proliferation, differentiation and collagen production by themselves [31]. When CNT persist into the tissue, remodeling is therefore permanent and degenerates into uncontrolled scar and fibrosis [24].
The global interpretation of these sophisticated mechanisms resulted in the elaboration of several “adverse outcome pathways” (AOP), which detail the exact sequence of molecular and cellular events required for inflammation and fibrosis development after CNT exposure (e.g., AOP #173). AOP are now available to improve current in vitro assays predicting inflammatory and fibrogenic effects of CNT and assess health risks for human [29,32,33] (Figure 1).
2.3. The Key Importance of Physico-Chemical Characteristics
The physicochemical features responsible for the inflammogenic and fibrogenic activities of CNT were defined by using chemically and morphologically modified CNT as well as validated in vitro and in vivo models. Size and shape strongly affect CNT toxicity (Figure 1). Our data showed that long CNT were more potent than short CNT to stimulate fibroblasts or macrophages, indicating that longer particles are more efficient in inducing inflammation and fibrosis [29,34]. CNT diameter was also identified as an important factor since thin CNT are more toxic for the lungs compared to thick CNT [35]. The morphology also determines their capacity to induce inflammation and fibrosis. Tangled CNT are less inflammatory and fibrogenic than straight (needled) CNT [36,37]. Furthermore, the surface reactivity plays a direct role in the ability of CNT to induce inflammatory and fibrotic responses. We demonstrated that the toxic effects of CNT are related to defective sites in the C framework because structural defects introduced in CNT by fracturing procedures increase inflammatory and fibrogenic activity [38,39]. Oxidative stress caused by toxic particles (i.e., silica and asbestos) and/or inflammatory cells (i.e., neutrophils and macrophages) is implicated in inflammation and fibrosis. We proposed, however, that other CNT features than free radical generation govern the toxic potential of CNT. Indeed, CNT exhibit a remarkable radical scavenging capacity and quench rather than generate oxygenated free radicals. This scavenging activity was related to CNT defects and their inflammatory and fibrotic potentials [39] (Figure 1).
3. The Mesotheliomagenic Activity of CNT: A Remaining Issue
3.1. Mesothelioma and Particles
Malignant mesothelioma (MM) is a cancer affecting the mesothelium, a layer of squamous cells covering the serous cavities of the body (pleura, peritoneum, pericardium) and protecting the organs they contain (lungs, peritoneal organs, heart). The most commonly affected tissue is the pleura and peritoneum; this is referred to as pleural and peritoneal malignant mesothelioma [40,41].
MM is an uncommon cancer and most of the cases are due to asbestos exposure [42]. In 1997, all asbestos fibers were classified as carcinogenic to human by IARC (International Agency for Research on Cancer, monograph 100C) because they are inhalable, poorly soluble and can migrate from the lungs to the pleura or peritoneum, directly or through the lymphatic system [42]. These low-degradable fibers trigger MM and other terminal pathologies such as pulmonary fibrosis and bronchial cancer in animal and human [43].
MM can occur more than 40 years after exposure. Considering the use of asbestos until the 1980s and the long latency period of the disease, a peak incidence is expected for 2020 [44]. Unfortunately, patients often reach the final stage of the disease when they are detected making the prognosis of MM very poor (12–15 months) [45,46]. Treatment with conventional therapies is not effective. The classical clinical management is chemotherapy based on platinum salts (alkylating agents) and Pemetrexed (anti-metabolite), which prolongs the patient survival by only 15 months. Debates also remain regarding other multimodal approaches as surgery and radiation [47,48].
Some CNT have been incriminated as being responsible for MM because their physically similarity to asbestos fibers. Several in vivo studies demonstrated that the long and straight multi-walled (MW) CNT-Mitsui-7 are indubitably mesotheliomagenic (see below) [49,50]. In 2014, IARC debated on the carcinogenicity of CNTs (monograph 111) and the consulted experts classified CNT-7 in Group 2b, i.e., as possibly carcinogenic to human [51,52]. All other CNT (single-walled or multi-walled nanotubes) were classified in group 3 (not classifiable as to their carcinogenicity to human) [53].
3.2. Advances in Understanding Mesothelioma Development
The precise cellular and molecular mechanisms explaining asbestos- or CNT-induced MM are difficult to investigate. Indeed, mesothelial cells progressively acquire features common to cancer and tissues representing non-advanced stages of the disease are difficult to obtain. Nevertheless, several pathological mechanisms have already been identified by using cell lines, biopsies and animal models [43,54].
3.2.1. Direct Effects and DNA Damages
Carcinogenicity of asbestos is related to their physical and chemical properties. Once fibers reach the pleural or peritoneal cavity, longer particles accumulate and are directly in contact with mesothelial cells [22,55,56]. Asbestos generate reactive oxygen species (ROS) and induce ROS production by exposed mesothelial cells. ROS trigger genomic instability and mutations by interacting with mesothelial cells [57]. These oxidant molecules also activate various signalling pathways, inducing transformed cell proliferation and survival [22,55,56]. ROS are therefore involved in the initiation, promotion and progression of cancer, which represent the three stages of the carcinogenic process [58].
Asbestos are known to penetrate cell membrane and interact with intracellular molecules resulting in direct (or primary) genotoxicity that include DNA strand breaks, mutations and chromosomal aberrations [57]. Recent studies showed that CNT induce chromosomal disruptions, fragmentations and translocations [59,60] and their nuclear deposition results in clear epigenetic alterations [61,62,63]. The quenching capacity of CNT is also involved in their primary genotoxic effects [39] (Figure 1). However, the literature concerning the direct interaction of asbestos or CNT with DNA is contradictory. The team of Mossman [64] states that this process does not exist, unlike most other chemical carcinogens. The non-clastogenicity of the CNT observed by Sasaki suggest that the CNT may not directly interact with DNA [65].
3.2.2. Inflammatory Responses as a Driver of Malignant Mesothelioma Development
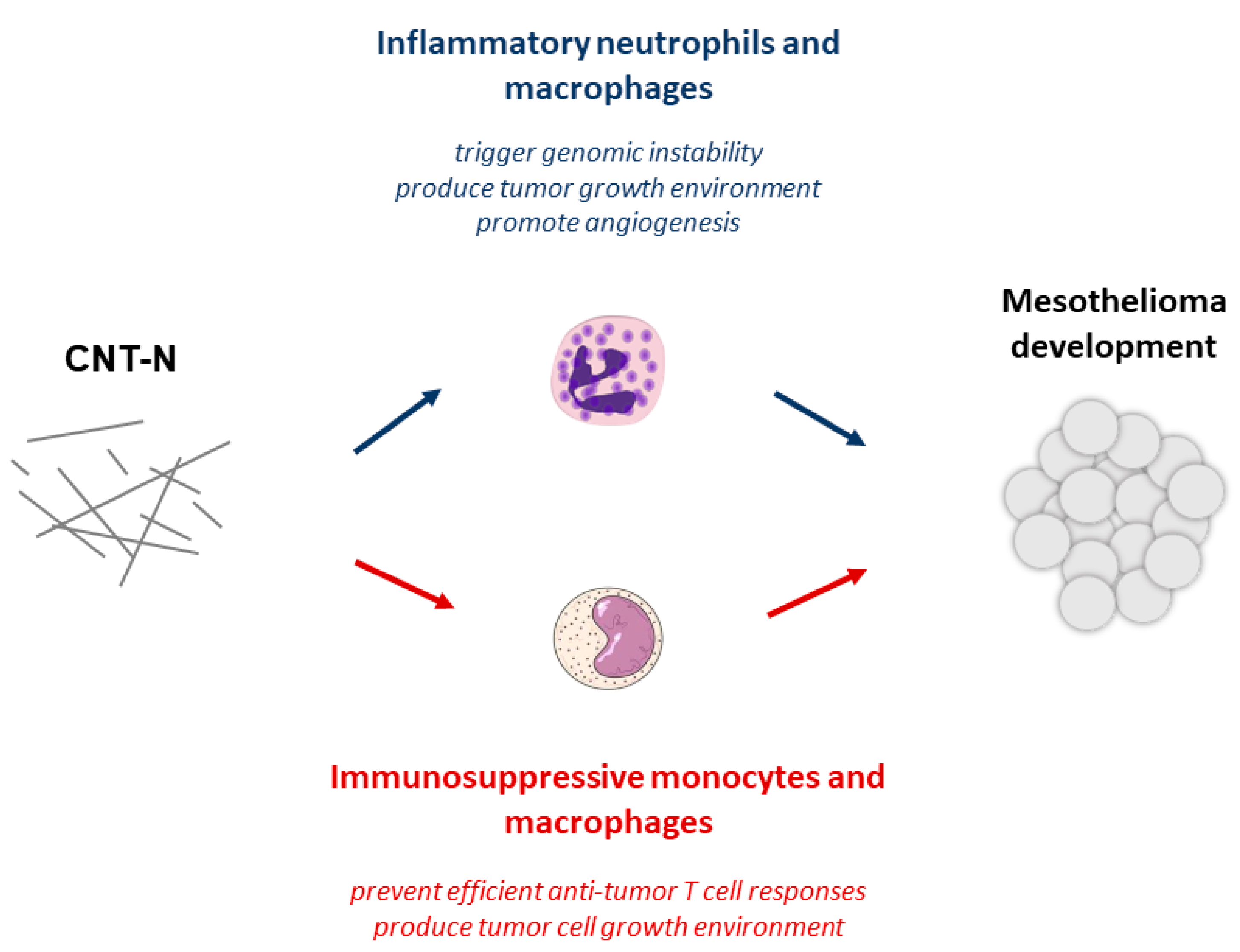
Numerous studies indicate that mesotheliomagenic particles induce, instead, secondary genotoxic damages by promoting inflammation and subsequent free radical release [66,67,68] (Figure 2). Resident macrophage present in mesothelial cell-covered tissues, detect, phagocytize and attempt to degrade inhaled fibers [69]. Whether these particles are long and resistant to degradation by macrophages, they trigger a phenomenon known as frustrated phagocytosis [56,58], which result in the constant and prolonged release of highly reactive inflammatory mediators (see above) [70,71] These mediators notably recruit neutrophils and additional macrophages for further particle clearance [56,58,71,72]. However, this particle removal process is not efficient for persistent fibers and the inflammation becomes consequently chronic [50,56,58].
ROS deriving from frustrated macrophages and neutrophils (Figure 2) cause direct mutations and promote proliferation and invasion of transformed mesothelial cells by modulating cell signalling pathways [55,58,73]. These inflammatory cells also permit mutated mesothelial cells to avoid apoptosis [74]. Indeed, cytokines and alarmins such as TNF-α and HMGB1 produced by macrophages and neutrophils activate the NF-κB (Nuclear Factor Kappa B) signalling pathway in mesothelial cells. This transcription factor then induces the expression of various genes promoting cell survival [75,76]. These inflammatory mediators also convert mesothelial cells to inflammatory cell partner that release growth and differentiation factors for stem (M- and GM-CSF) [77] and endothelial cells (VEGF) [78]. Together, these factors increase the survival of transformed mesothelial cells and allow neoplastic cell migration into the tissue (Figure 2).
3.2.3. Tolerant Microenvironment and Immunosuppressive Cells within Mesothelial Tumors
Overall, these convincing data therefore demonstrate that chronic inflammation contributes to the initiation, promotion and progression of MM. However, several recent studies have suggested additional mechanisms to explain the varied pathological profiles among clinical cases. In recent years, a new concept was developed proposing that the immune evasion permits mesothelioma to evade host anti-tumor responses [79,80,81].
It is now admitted that hematopoietic and lymphocytic immune cells infiltrating MM are reprogrammed by their new microenvironment and play a critical role in the maintenance and progression of cancer. These immunosuppressive or immunoregulatory cells create a tolerant environment by blocking T lymphocytes (dedicated to recognize and eliminate mutated cells) and stimulate tumor growth by promoting angiogenesis, stroma deposition and metastatic tumor formation [82]. Immunosuppressive host cells invading mesothelioma include regulatory T lymphocytes (T regs) [83,84], Myeloid-Derived Suppressor Cells (MDSCs) [85] and Tumor-Associated Macrophages (TAMs) [86,87]. Soluble factors (IL-10, TGF-β and PGE2) and immune checkpoint ligands (PD-1 and CTLA-4) represent the main elements contributing to the establishment of an immunosuppressive microenvironment [79].
The discovery of this tolerant tumor environment resulted in new clinical approaches to control mesothelioma. Immunotherapy aims to boost immunity and block immunosuppressive capacities of tumor cells. The current clinical option to reverse immunosuppressive mechanisms is to inhibit immune checkpoints (anti CTLA-4 and PDL-1 neutralizing antibodies) during cytoreduction therapy (chemotherapy, radiotherapy and surgery) [80,81,88,89,90]. Tumor immune escapes are also operative in animal models of mesothelioma and innovative therapeutic strategies modifying immunosuppressive monocyte and macrophage differentiation are now successfully obtained [77,91].
3.2.4. The Co-Existence of Early Inflammation and Immunosuppression after Mesotheliomagenic Particle-Exposure
Our team was able to demonstrate that immunosuppressive responses were not exclusively generate by tumor cells but also by mesotheliomagenic CNT themselves. Indeed, we demonstrated that mesotheliomagenic CNT induce an early immunosuppressive environment by recruiting immunosuppressive M-MDSCs and macrophages after few days in injected rats (Figure 1 and Figure 2) [92]. This effect is associated with the acute neutrophilic inflammation already well described (see above). The early presence of MDSC was later confirmed by other teams [93,94]. Increased expression of immunosuppressive mediators such as IL-10, TGF-β, NADPH oxidase and prostaglandin synthase was also noted in murine lungs after acute exposure to mesotheliomagenic CNT [95,96]. The rapid development of local and systemic immunosuppressive immune responses reduce the number of circulating T lymphocytes and their ability to proliferate [97,98,99]. This effect is mainly related to IL-10 and TGF-β [96]. Kido and colleagues also noted a rapid increase of IL-10 expression by macrophages after CNT inhalation in rats [100]. IL-10 release by macrophages results in IL-2 deficiency (an essential factor for T cell proliferation) and reduces antitumor activity of T lymphocytes.
Altogether, these results indicate that CNT rapidly induce an immunosuppressive environment, affect T lymphocyte activity and expand transformed mesothelial cells generated by conjoint inflammatory elements (Figure 2). The requirement of these dual environments can explain the diversity of mesothelioma types and long latency period of the disease.
4. An Unexpected Application: The Diversity of CNT to Identify New Pathological Pathways in Malignant Mesothelioma
4.1. Toxic Needled-Like (CNT-N) Versus Non-Toxic Tangled-Like (CNT-T) Carbon Nanotubes
Several animal models have been established to study mesothelioma by using asbestos fibers. Intraperitoneal injection is the preferred administration route because long, straight, fibrous and solid fibers directly reach mesothelial cells of the peritoneal cavity. In addition, the clearance mechanisms of this cavity are similar to those of pleural cavity, which is the predilection site for malignant mesothelioma [22,56]. Not all rodents are equally sensitive to mesotheliomagenic fibers. Indeed, the incidence in rats is higher than in mice, which therefore appears to be more resistant to mesothelioma development [68].
The mesotheliomagenic properties of CNT represent an interesting tool for investigating mesothelioma in animals. We have shown that CNT-induced mesothelioma affects all treated rats (only 30% to 50% of animals injected with asbestos) in a limited period (6 months for CNT instead of 2 years for asbestos) [92,101]. The other key advantage of CNT compared to natural asbestos is the existence of a very wide range of manufactured CNT. These particles can be categorized from a structural and toxicological point of view by using their morphology (tangled CNT T versus needled/straight/rod CNT N). CNT-T are thin enough to fold and self-assemble into short, tangled aggregates, while straight CNT-N are fibrous, resistant and long. Unlike asbestos fibers, which are all considered carcinogenic, several studies demonstrate that only CNT-N are associated with the development of chronic pathologies (including mesothelioma) unlike CNT-T, which are poorly reactive and toxic [92,101,102,103]. The exact reasons for this discrepancy have not yet been fully elucidated. The greater bio-persistence of CNT-N (longer and therefore more difficult to clear) is one possible explanation.
In that respect, Poland and co-authors reported frustrated phagocytosis by macrophages in rodents treated with CNT-N. Accordingly, these needled CNT-N are not completely covered by phagocytes and remain biopersistent in the tissue. In contrast, tangled CNT-T are entirely phagocytosed and taken up by macrophages resulting in tissue translocation, particle biodegradation and accelerated rate of clearance [18,104] Nanotube geometry is also crucial in the development of inflammation, lymphoid infiltration and granuloma formation in animals. Only CNT-N exposure resulted in neutrophilic inflammation and larger lymphoid and fibrotic granulomas contrary to CNT-T [37,105]. Needled CNT-N were more potent than tangled CNT T to elicit inflammatory effects towards macrophages [106,107].
For Sasaki and colleagues, the shape- and length-related structural determinants of CNT are also crucial factors for their potential carcinogenic activity. Straight and fibrous CNT-N were the strongest inducers of chromosomal aberrations in cell cultures [65]. CNT-N elicit a more pronounced primary genotoxicity effect than CNT-T, as assessed by DNA damage and micronuclei formation [108]. These in vitro observations are in accordance with data obtained in vivo. CNT-N cause secondary genotoxicity and mesothelioma in rodents (absolute incidences of 100%) contrary to CNT-T (no genotoxicity and tumor) [92,101,102,103,108,109].
4.2. A New Strategy to Identify Pathogenic Immune Pathways: Gene or Protein Expression by Purified Macrophages after Needled and Tangled CNT Exposure
The paradigm associating physicochemical characteristics and carcinogenicity of CNT represents a major asset for risk management and predictive toxicology. However, these data do not entirely explain the reasons why some CNT induce or not mesothelioma, and do not offer specific mechanisms or targets for clinical management or drug development against mesothelioma. RNA-related profiling methods and next-generation sequencing (NGS) technologies were elaborated to cartography genes expressed by purified cell populations. Bioinformatic analysis of these (big) data often reveal unexpected pathological axis at the level of molecular and cell biology [110].
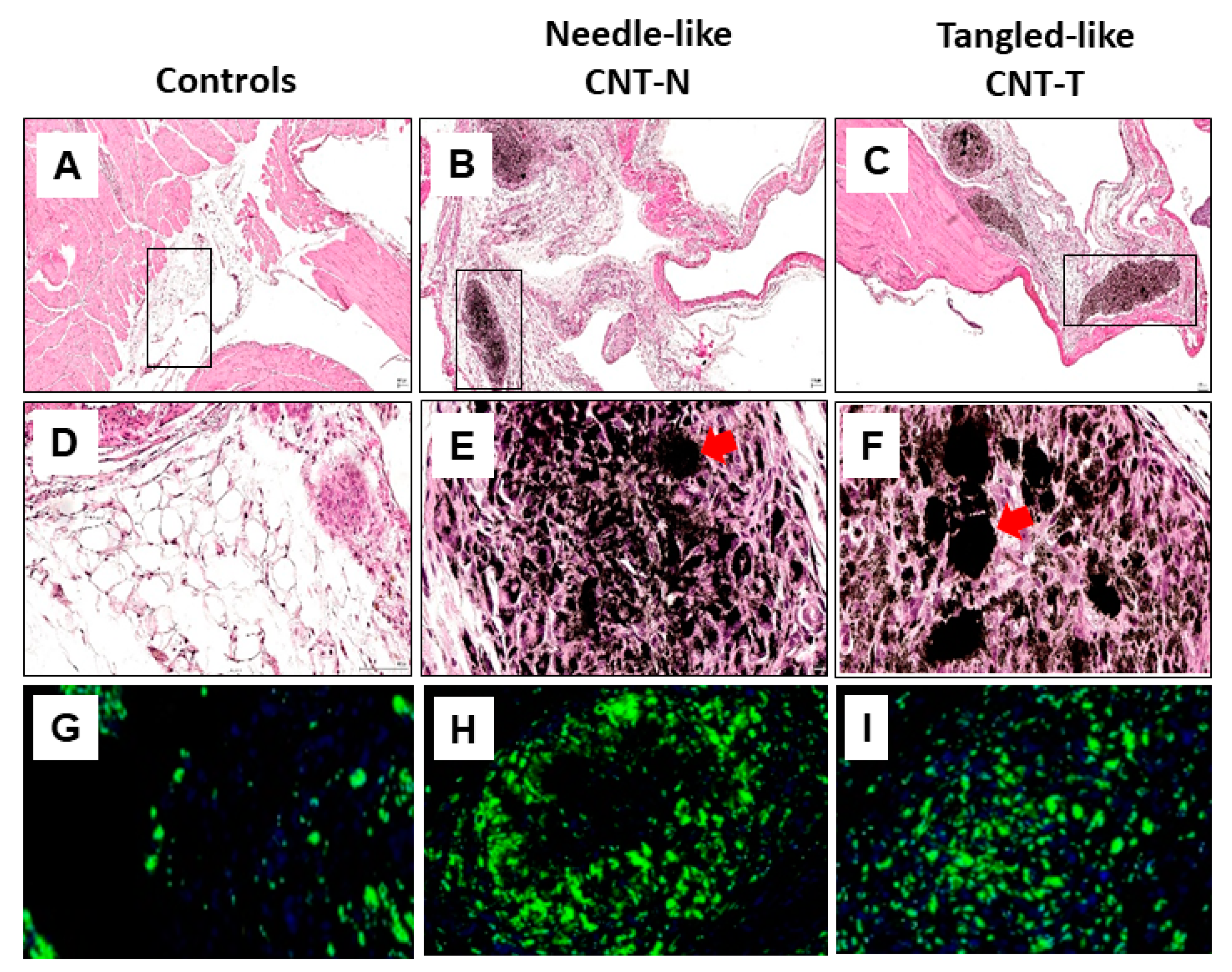
We used this new strategy to shed light on the immune events that specifically regulate mesothelioma development by comparing the effects of CNT on macrophages (Figure 3 and Figure 4). We used our model of early responses (induced by particles and not tumor cells) in rat (sensitive species), after a single intraperitoneal injection of mesotheliomagenic (CNT-N) and non-mesotheliomagenic (CNT-T) CNT [92] (Figure 3). Through histological analysis, we showed that exposure to both types of CNT similarly induces granuloma formation in the connective tissue bordering the peritoneal cavity. CNT-N and CNT-T are assembled in granuloma center and form compact crystalline structures (red arrow, Figure 3). Under both exposure conditions (CNT-T and CNT-N), numerous CD68 positive cells (macrophages) infiltrate damaged tissue and surround CNT. Most of the cells constituting granuloma are macrophages in a greater proportion in tissues exposed to CNT-N and CNT-T compared to controls (Figure 3). Therefore, mesotheliomagenic CNT-N and non-mesotheliomagenic CNT-T alter peritoneal tissues and induce macrophage accumulation in a comparable manner.
Macrophages are crucial immune cells in response to particles (see above). Their versatility is well recognized by immunologists and various distinct functional phenotypes (termed macrophage polarization) directed by specific microenvironmental stimuli and signals have been established and associated to allergic, parasitic and autoimmune diseases for instance [111]. Their cellular origin has been revisited by recent observations, showing that macrophages derive from circulating blood monocytes but also originate from embryonic progenitors and proliferate [112]. The macrophage polarization diversity and our histological observations suggest that the difference between the two comparative models (with and without mesothelioma) is not simply related to the presence or absence of macrophages in affected tissues but probably resides in the ability of macrophages to adopt contrasting immune profiles. These differentially polarized macrophages can explain the inappropriate immune responses leading to mesothelioma development.
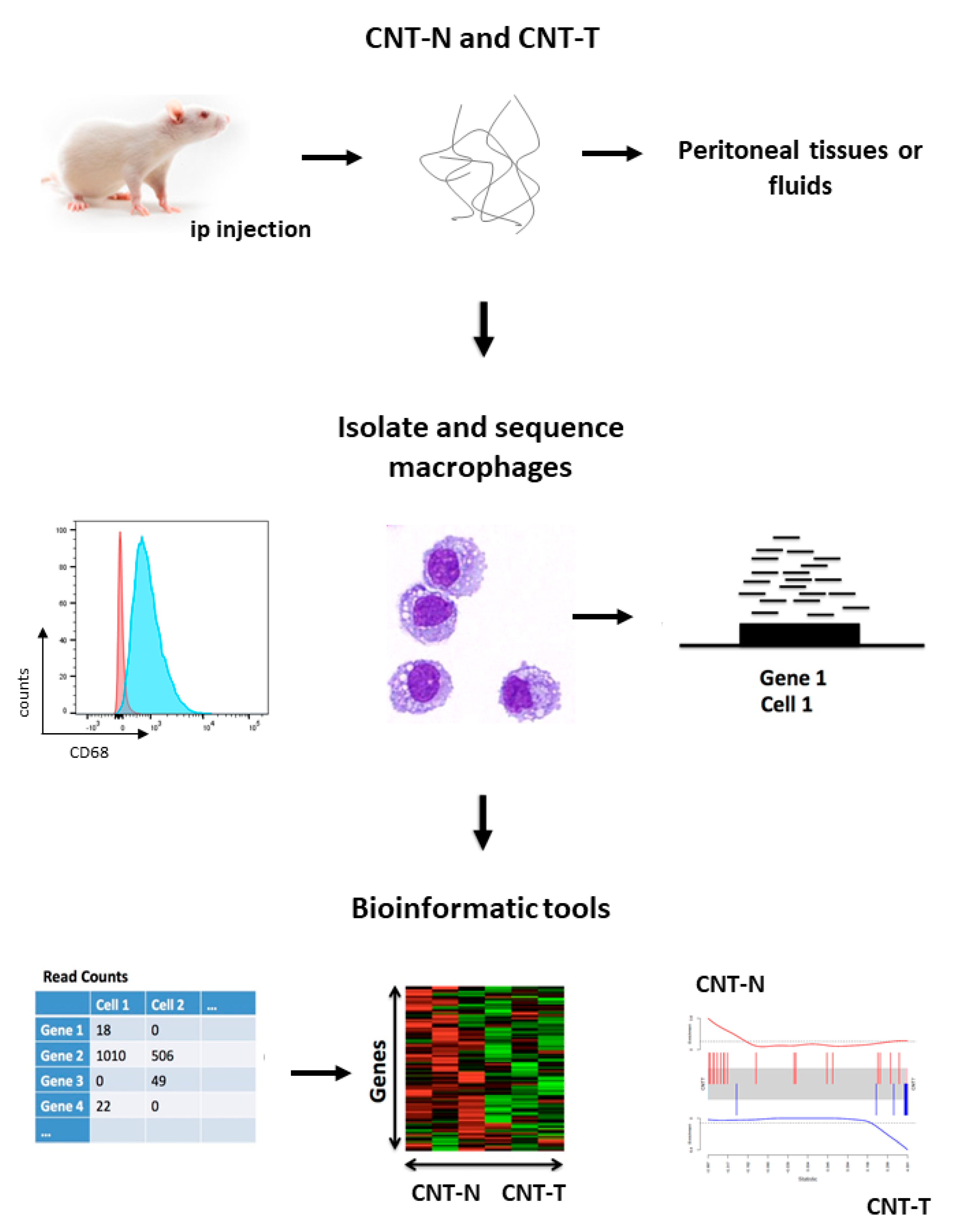
We applied NGS-gene profiling methods to our model to reveal macrophage profile unambiguously related to mesothelioma (Figure 4). Peritoneal macrophages collected from CNT-N or CNT-T-treated rats were purified using flow cytometry cell sorting (e.g; CD68 positive cells). RNA was isolated from bulk macrophage populations and libraries were prepared and sequenced using an Illumina platform. Bioinformatics tools analyzed RNA sequences, generated comparative tables or barcode plots and associated immune and carcinogenic responses for each macrophage subpopulation (Figure 4).
NGS technologies have evolved dramatically in recent years, making individual cell analysis possible. Single-cell RNA sequencing now reveals new characteristics of macrophage subpopulations and is directly relevant for studying and tracing distinct macrophage lineages and polarizations in chronic diseases [113]. The proteomic resources are also concerned by innovative technologies. New generation of mass spectrometry quantitatively analyze protein networks at single cell resolution [114,115]. Interestingly, recent bioinformatic tools integrate conjoint analyses of the transcriptome (RNA) and proteome (protein) in purified macrophages, boosting their characterization and classification [116]. These new technical platforms and experimental strategies greatly help basic science and medical applications and, if used in CNT toxicology, may open new exciting horizons in the physiopathology and for the therapy of mesothelioma.
Author Contributions
F.H., C.A.H., R.L. and M.O. were involved in the conception of the review and wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Actions de Recherche Concertées, Fédération Wallonie-Bruxelles (ARC 19/24-098, CYTAID), Fondation Contre le Cancer (2019-219), Fonds de la Recherche Scientifique (FNRS, PDR T.0119.13), ANSES (Agence nationale française de sécurité sanitaire de l’alimentation, de l’environnement et du travail, MacFibOsis) and European Commission under H2020 project (Contract no. 874707, Eximious). F.H. is a Senior Research Associate with the FNRS, Belgium.
Conflicts of Interest
The authors declare that they have no competing interests.
References
- Qin, L.C.; Zhao, X.; Hirahara, K.; Miyamoto, Y.; Ando, Y.; Iijima, S. The smallest carbon nanotube. Nature 2000, 408, 50. [Google Scholar] [CrossRef]
- Amelinckx, S.; Zhang, X.B.; Bernaerts, D.; Zhang, X.F.; Ivanov, V.; Nagy, J.B. A formation mechanism for catalytically grown helix-shaped graphite nanotubes. Science 1994, 265, 635–639. [Google Scholar] [CrossRef]
- Simon, J.; Flahaut, E.; Golzio, M. Overview of Carbon Nanotubes for Biomedical Applications. Materials 2019, 12, 624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohajeri, M.; Behnam, B.; Sahebkar, A. Biomedical applications of carbon nanomaterials: Drug and gene delivery potentials. J. Cell Physiol. 2018, 234, 298–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Available online: www.idtechex.com/fr/research-report/graphene-2d-materials-and-carbon-nanotubes-markets-technologies-and-opportunities-2018-2028/603; https://www.marketsandmarkets.com/Market-Reports/carbon-nanotubes-139; www.yanoresearch.com/en/press-release/show/press_id/2081-2019 (accessed on 9 May 2020).
- King, S.G.; McCafferty, L.; Tas, M.O.; Snashall, K.; Chen, J.S.; Shkunov, M.; Stolojan, V.; Silva, S.R.P. Low-Cost Catalyst Ink for Simple Patterning and Growth of High-Quality Single- and Double-Walled Carbon Nanotubes. ACS Appl. Mater. Interfaces 2020, 12, 11898–11906. [Google Scholar] [CrossRef]
- Kong, Y.; Nanjundan, A.K.; Liu, Y.; Song, H.; Huang, X.; Yu, C. Modulating Ion Diffusivity and Electrode Conductivity of Carbon Nanotube@Mesoporous Carbon Fibers for High Performance Aluminum-Selenium Batteries. Small 2019, 15, e1904310. [Google Scholar] [CrossRef]
- Luo, Y.; Wang, K.; Li, Q.; Fan, S.; Wang, J. Macroscopic Carbon Nanotube Structures for Lithium Batteries. Small 2020, 16, e1902719. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Cui, C.; Chen, H.; Yu, X.; Qian, W. The Application of Carbon Nanotube/Graphene-Based Nanomaterials in Wastewater Treatment. Small 2020, 16, e1902301. [Google Scholar] [CrossRef]
- Baby, R.; Saifullah, B.; Hussein, M.Z. Carbon Nanomaterials for the Treatment of Heavy Metal-Contaminated Water and Environmental Remediation. Nanoscale Res. Lett. 2019, 14, 341. [Google Scholar] [CrossRef] [Green Version]
- Silva, R.C.F.; Ardisson, J.D.; Cotta, A.A.C.; Araujo, M.H.; Teixeira, A.P.C. Use of iron mining tailings from dams for carbon nanotubes synthesis in fluidized bed for 17alpha-ethinylestradiol removal. Environ. Pollut. 2020, 260, 114099. [Google Scholar] [CrossRef]
- Venkataraman, A.; Amadi, E.V.; Chen, Y.; Papadopoulos, C. Carbon Nanotube Assembly and Integration for Applications. Nanoscale Res. Lett. 2019, 14, 220. [Google Scholar] [CrossRef] [PubMed]
- Maiti, D.; Tong, X.; Mou, X.; Yang, K. Carbon-Based Nanomaterials for Biomedical Applications: A Recent Study. Front. Pharmacol. 2018, 9, 1401. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Dominguez, C.M.; Bauer, J.; Weigel, S.; Schipperges, A.; Oelschlaeger, C.; Willenbacher, N.; Keppler, S.; Bastmeyer, M.; Heissler, S.; et al. Carbon-nanotube reinforcement of DNA-silica nanocomposites yields programmable and cell-instructive biocoatings. Nat. Commun. 2019, 10, 5522. [Google Scholar] [CrossRef] [Green Version]
- Pei, B.; Wang, W.; Dunne, N.; Li, X. Applications of Carbon Nanotubes in Bone Tissue Regeneration and Engineering: Superiority, Concerns, Current Advancements, and Prospects. Nanomaterials 2019, 9, 1501. [Google Scholar] [CrossRef] [Green Version]
- Takagi, A.; Hirose, A.; Nishimura, T.; Fukumori, N.; Ogata, A.; Ohashi, N.; Kitajima, S.; Kanno, J. Induction of mesothelioma in p53+/− mouse by intraperitoneal application of multi-wall carbon nanotube. J. Toxicol. Sci. 2008, 33, 105–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, J.; Huaux, F.; Moreau, N.; Misson, P.; Heilier, J.F.; Delos, M.; Arras, M.; Fonseca, A.; Nagy, J.B.; Lison, D. Respiratory toxicity of multi-wall carbon nanotubes. Toxicol. Appl. Pharmacol. 2005, 207, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Poland, C.A.; Duffin, R.; Kinloch, I.; Maynard, A.; Wallace, W.A.; Seaton, A.; Stone, V.; Brown, S.; Macnee, W.; Donaldson, K. Carbon nanotubes introduced into the abdominal cavity of mice show asbestos-like pathogenicity in a pilot study. Nat. Nanotechnol. 2008, 3, 423–428. [Google Scholar] [CrossRef]
- Shvedova, A.A.; Castranova, V.; Kisin, E.R.; Schwegler-Berry, D.; Murray, A.R.; Gandelsman, V.Z.; Maynard, A.; Baron, P. Exposure to carbon nanotube material: Assessment of nanotube cytotoxicity using human keratinocyte cells. J. Toxicol. Environ. Health A 2003, 66, 1909–1926. [Google Scholar] [CrossRef]
- Lam, C.W.; James, J.T.; McCluskey, R.; Hunter, R.L. Pulmonary toxicity of single-wall carbon nanotubes in mice 7 and 90 days after intratracheal instillation. Toxicol. Sci. 2004, 77, 126–134. [Google Scholar] [CrossRef] [Green Version]
- Shvedova, A.A.; Kisin, E.R.; Mercer, R.; Murray, A.R.; Johnson, V.J.; Potapovich, A.I.; Tyurina, Y.Y.; Gorelik, O.; Arepalli, S.; Schwegler-Berry, D.; et al. Unusual inflammatory and fibrogenic pulmonary responses to single-walled carbon nanotubes in mice. Am. J. Physiol. Lung Cell. Mol. Physiol. 2005, 289, L698–L708. [Google Scholar] [CrossRef] [Green Version]
- Donaldson, K.; Poland, C.A.; Murphy, F.A.; MacFarlane, M.; Chernova, T.; Schinwald, A. Pulmonary toxicity of carbon nanotubes and asbestos—Similarities and differences. Adv. Drug Deliv. Rev. 2013, 65, 2078–2086. [Google Scholar] [CrossRef]
- Mossman, B.T. Mechanistic in vitro studies: What they have told us about carcinogenic properties of elongated mineral particles (EMPs). Toxicol. Appl. Pharmacol. 2018, 361, 62–67. [Google Scholar] [CrossRef]
- Huaux, F. Emerging Role of Immunosuppression in Diseases Induced by Micro- and Nano-Particles: Time to Revisit the Exclusive Inflammatory Scenario. Front. Immunol. 2018, 9, 2364. [Google Scholar] [CrossRef] [Green Version]
- Huaux, F. Innate immunity to inhaled particles: A new paradigm of collective recognition. Curr. Opin. Toxicol. 2018, 10, 84–90. [Google Scholar] [CrossRef]
- Pavan, C.; Delle Piane, M.; Gullo, M.; Filippi, F.; Fubini, B.; Hoet, P.; Horwell, C.J.; Huaux, F.; Lison, D.; Lo Giudice, C.; et al. The puzzling issue of silica toxicity: Are silanols bridging the gaps between surface states and pathogenicity? Part. Fibre Toxicol. 2019, 16, 32. [Google Scholar] [CrossRef] [PubMed]
- Rabolli, V.; Lison, D.; Huaux, F. The complex cascade of cellular events governing inflammasome activation and IL-1beta processing in response to inhaled particles. Part. Fibre Toxicol. 2016, 13, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabolli, V.; Badissi, A.A.; Devosse, R.; Uwambayinema, F.; Yakoub, Y.; Palmai-Pallag, M.; Lebrun, A.; De Gussem, V.; Couillin, I.; Ryffel, B.; et al. The alarmin IL-1alpha is a master cytokine in acute lung inflammation induced by silica micro- and nanoparticles. Part. Fibre Toxicol. 2014, 11, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vietti, G.; Lison, D.; van den Brule, S. Mechanisms of lung fibrosis induced by carbon nanotubes: Towards an Adverse Outcome Pathway (AOP). Part. Fibre Toxicol. 2016, 13, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebrun, A.; Lo Re, S.; Chantry, M.; Izquierdo Carerra, X.; Uwambayinema, F.; Ricci, D.; Devosse, R.; Ibouraadaten, S.; Brombin, L.; Palmai-Pallag, M.; et al. CCR2(+) monocytic myeloid-derived suppressor cells (M-MDSCs) inhibit collagen degradation and promote lung fibrosis by producing transforming growth factor-beta1. J. Pathol. 2017, 243, 320–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vietti, G.; Ibouraadaten, S.; Palmai-Pallag, M.; Yakoub, Y.; Piret, J.P.; Marbaix, E.; Lison, D.; van den Brule, S. Towards predicting the lung fibrogenic activity of MWCNT: Key role of endocytosis, kinase receptors and ERK 1/2 signaling. Nanotoxicology 2016, 10, 488–500. [Google Scholar] [CrossRef]
- Nikota, J.; Banville, A.; Goodwin, L.R.; Wu, D.; Williams, A.; Yauk, C.L.; Wallin, H.; Vogel, U.; Halappanavar, S. Stat-6 signaling pathway and not Interleukin-1 mediates multi-walled carbon nanotube-induced lung fibrosis in mice: Insights from an adverse outcome pathway framework. Part. Fibre Toxicol. 2017, 14, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guseva Canu, I.; Batsungnoen, K.; Maynard, A.; Hopf, N.B. State of knowledge on the occupational exposure to carbon nanotubes. Int. J. Hyg. Environ. Health 2020, 225, 113472. [Google Scholar] [CrossRef]
- Vietti, G.; Ibouraadaten, S.; Palmai-Pallag, M.; Yakoub, Y.; Bailly, C.; Fenoglio, I.; Marbaix, E.; Lison, D.; van den Brule, S. Towards predicting the lung fibrogenic activity of nanomaterials: Experimental validation of an in vitro fibroblast proliferation assay. Part. Fibre Toxicol. 2013, 10, 52. [Google Scholar] [CrossRef] [Green Version]
- Fenoglio, I.; Aldieri, E.; Gazzano, E.; Cesano, F.; Colonna, M.; Scarano, D.; Mazzucco, G.; Attanasio, A.; Yakoub, Y.; Lison, D.; et al. Thickness of multiwalled carbon nanotubes affects their lung toxicity. Chem. Res. Toxicol. 2012, 25, 74–82. [Google Scholar] [CrossRef]
- Murphy, F.A.; Poland, C.A.; Duffin, R.; Donaldson, K. Length-dependent pleural inflammation and parietal pleural responses after deposition of carbon nanotubes in the pulmonary airspaces of mice. Nanotoxicology 2013, 7, 1157–1167. [Google Scholar] [CrossRef]
- Duke, K.S.; Thompson, E.A.; Ihrie, M.D.; Taylor-Just, A.J.; Ash, E.A.; Shipkowski, K.A.; Hall, J.R.; Tokarz, D.A.; Cesta, M.F.; Hubbs, A.F.; et al. Role of p53 in the chronic pulmonary immune response to tangled or rod-like multi-walled carbon nanotubes. Nanotoxicology 2018, 12, 975–991. [Google Scholar] [CrossRef]
- Muller, J.; Huaux, F.; Fonseca, A.; Nagy, J.B.; Moreau, N.; Delos, M.; Raymundo-Pinero, E.; Beguin, F.; Kirsch-Volders, M.; Fenoglio, I.; et al. Structural defects play a major role in the acute lung toxicity of multiwall carbon nanotubes: Toxicological aspects. Chem. Res. Toxicol. 2008, 21, 1698–1705. [Google Scholar] [CrossRef] [PubMed]
- Fenoglio, I.; Greco, G.; Tomatis, M.; Muller, J.; Raymundo-Pinero, E.; Beguin, F.; Fonseca, A.; Nagy, J.B.; Lison, D.; Fubini, B. Structural defects play a major role in the acute lung toxicity of multiwall carbon nanotubes: Physicochemical aspects. Chem. Res. Toxicol. 2008, 21, 1690–1697. [Google Scholar] [CrossRef] [PubMed]
- van Gerwen, M.; Alpert, N.; Flores, R.; Taioli, E. An overview of existing mesothelioma registries worldwide, and the need for a US Registry. Am. J. Ind. Med. 2020, 63, 115–120. [Google Scholar] [CrossRef]
- Carbone, M.; Adusumilli, P.S.; Alexander, H.R., Jr.; Baas, P.; Bardelli, F.; Bononi, A.; Bueno, R.; Felley-Bosco, E.; Galateau-Salle, F.; Jablons, D.; et al. Mesothelioma: Scientific clues for prevention, diagnosis, and therapy. CA Cancer J. Clin. 2019, 69, 402–429. [Google Scholar] [CrossRef] [Green Version]
- Poland, C.A.; Duffin, R. The toxicology of chrysotile-containing brake debris: Implications for mesothelioma. Crit. Rev. Toxicol. 2019, 49, 11–35. [Google Scholar] [CrossRef] [PubMed]
- Gaudino, G.; Xue, J.; Yang, H. How asbestos and other fibers cause mesothelioma. Transl. Lung Cancer Res. 2020, 9, S39–S46. [Google Scholar] [CrossRef] [PubMed]
- Alpert, N.; van Gerwen, M.; Taioli, E. Epidemiology of mesothelioma in the 21st century in Europe and the United States, 40 years after restricted/banned asbestos use. Transl. Lung Cancer Res. 2020, 9, S28–S38. [Google Scholar] [CrossRef]
- Fels Elliott, D.R.; Jones, K.D. Diagnosis of Mesothelioma. Surg. Pathol. Clin. 2020, 13, 73–89. [Google Scholar] [CrossRef] [PubMed]
- Kim, R.Y.; Sterman, D.H.; Haas, A.R. Malignant Mesothelioma: Has Anything Changed? Semin. Respir. Crit. Care Med. 2019, 40, 347–360. [Google Scholar] [CrossRef]
- Cantini, L.; Hassan, R.; Sterman, D.H.; Aerts, J. Emerging Treatments for Malignant Pleural Mesothelioma: Where Are We Heading? Front. Oncol. 2020, 10, 343. [Google Scholar] [CrossRef] [Green Version]
- Disselhorst, M.J.; Baas, P. Chemotherapy options versus “novel” therapies: How should we treat patients with malignant pleural mesothelioma. Transl. Lung Cancer Res. 2020, 9, S77–S85. [Google Scholar] [CrossRef]
- Dong, J.; Ma, Q. Integration of inflammation, fibrosis, and cancer induced by carbon nanotubes. Nanotoxicology 2019, 13, 1244–1274. [Google Scholar] [CrossRef]
- Sinis, S.I.; Hatzoglou, C.; Gourgoulianis, K.I.; Zarogiannis, S.G. Carbon Nanotubes and Other Engineered Nanoparticles Induced Pathophysiology on Mesothelial Cells and Mesothelial Membranes. Front. Physiol. 2018, 9, 295. [Google Scholar] [CrossRef] [Green Version]
- Grosse, Y.; Loomis, D.; Guyton, K.Z.; Lauby-Secretan, B.; El Ghissassi, F.; Bouvard, V.; Benbrahim-Tallaa, L.; Guha, N.; Scoccianti, C.; Mattock, H.; et al. Carcinogenicity of fluoro-edenite, silicon carbide fibres and whiskers, and carbon nanotubes. Lancet Oncol. 2014, 15, 1427–1428. [Google Scholar] [CrossRef]
- IARC Working Group on the Evaluation of Carcinogenic Risks to Humans. Some Nanomaterials and Some Fibres; PMID: 31829532; International Agency for Research on Cancer: Lyon, France, 2017. [Google Scholar]
- Kane, A.B.; Hurt, R.H.; Gao, H. The asbestos-carbon nanotube analogy: An update. Toxicol. Appl. Pharmacol. 2018, 361, 68–80. [Google Scholar] [CrossRef]
- Blanquart, C.; Jaurand, M.C.; Jean, D. The Biology of Malignant Mesothelioma and the Relevance of Preclinical Models. Front. Oncol. 2020, 10, 388. [Google Scholar] [CrossRef]
- Chew, S.H.; Toyokuni, S. Malignant mesothelioma as an oxidative stress-induced cancer: An update. Free Radic. Biol. Med. 2015, 86, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, K.; Murphy, F.A.; Duffin, R.; Poland, C.A. Asbestos, carbon nanotubes and the pleural mesothelium: A review of the hypothesis regarding the role of long fibre retention in the parietal pleura, inflammation and mesothelioma. Part. Fibre Toxicol. 2010, 7, 5. [Google Scholar] [CrossRef] [Green Version]
- Moller, P.; Jacobsen, N.R. Weight of evidence analysis for assessing the genotoxic potential of carbon nanotubes. Crit. Rev. Toxicol. 2017, 47, 867–884. [Google Scholar] [CrossRef] [PubMed]
- Benedetti, S.; Nuvoli, B.; Catalani, S.; Galati, R. Reactive oxygen species a double-edged sword for mesothelioma. Oncotarget 2015, 6, 16848–16865. [Google Scholar] [CrossRef] [Green Version]
- Siegrist, K.J.; Reynolds, S.H.; Porter, D.W.; Mercer, R.R.; Bauer, A.K.; Lowry, D.; Cena, L.; Stueckle, T.A.; Kashon, M.L.; Wiley, J.; et al. Mitsui-7, heat-treated, and nitrogen-doped multi-walled carbon nanotubes elicit genotoxicity in human lung epithelial cells. Part. Fibre Toxicol. 2019, 16, 36. [Google Scholar] [CrossRef] [Green Version]
- Snyder, R.J.; Verhein, K.C.; Vellers, H.L.; Burkholder, A.B.; Garantziotis, S.; Kleeberger, S.R. Multi-walled carbon nanotubes upregulate mitochondrial gene expression and trigger mitochondrial dysfunction in primary human bronchial epithelial cells. Nanotoxicology 2019, 13, 1344–1361. [Google Scholar] [CrossRef] [PubMed]
- Oner, D.; Ghosh, M.; Bove, H.; Moisse, M.; Boeckx, B.; Duca, R.C.; Poels, K.; Luyts, K.; Putzeys, E.; Van Landuydt, K.; et al. Differences in MWCNT- and SWCNT-induced DNA methylation alterations in association with the nuclear deposition. Part. Fibre Toxicol. 2018, 15, 11. [Google Scholar] [CrossRef] [PubMed]
- Emerce, E.; Ghosh, M.; Oner, D.; Duca, R.C.; Vanoirbeek, J.; Bekaert, B.; Hoet, P.H.M.; Godderis, L. Carbon Nanotube- and Asbestos-Induced DNA and RNA Methylation Changes in Bronchial Epithelial Cells. Chem. Res. Toxicol. 2019, 32, 850–860. [Google Scholar] [CrossRef] [PubMed]
- Oner, D.; Ghosh, M.; Coorens, R.; Bove, H.; Moisse, M.; Lambrechts, D.; Ameloot, M.; Godderis, L.; Hoet, P.H.M. Induction and recovery of CpG site specific methylation changes in human bronchial cells after long-term exposure to carbon nanotubes and asbestos. Environ. Int. 2020, 137, 105530. [Google Scholar] [CrossRef] [PubMed]
- Mossman, B.T.; Shukla, A.; Heintz, N.H.; Verschraegen, C.F.; Thomas, A.; Hassan, R. New insights into understanding the mechanisms, pathogenesis, and management of malignant mesotheliomas. Am. J. Pathol. 2013, 182, 1065–1077. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, T.; Asakura, M.; Ishioka, C.; Kasai, T.; Katagiri, T.; Fukushima, S. In vitro chromosomal aberrations induced by various shapes of multi-walled carbon nanotubes (MWCNTs). J. Occup. Health 2016, 58, 622–631. [Google Scholar] [CrossRef] [Green Version]
- Linton, A.; van Zandwijk, N.; Reid, G.; Clarke, S.; Cao, C.; Kao, S. Inflammation in malignant mesothelioma—Friend or foe? Ann. Cardiothorac. Surg. 2012, 1, 516–522. [Google Scholar] [CrossRef]
- Westbom, C.M.; Shukla, A.; MacPherson, M.B.; Yasewicz, E.C.; Miller, J.M.; Beuschel, S.L.; Steele, C.; Pass, H.I.; Vacek, P.M.; Shukla, A. CREB-induced inflammation is important for malignant mesothelioma growth. Am. J. Pathol. 2014, 184, 2816–2827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Testa, J.R.; Berns, A. Preclinical Models of Malignant Mesothelioma. Front. Oncol. 2020, 10, 101. [Google Scholar] [CrossRef] [Green Version]
- Kagan, V.E.; Konduru, N.V.; Feng, W.; Allen, B.L.; Conroy, J.; Volkov, Y.; Vlasova, I.I.; Belikova, N.A.; Yanamala, N.; Kapralov, A.; et al. Carbon nanotubes degraded by neutrophil myeloperoxidase induce less pulmonary inflammation. Nat. Nanotechnol. 2010, 5, 354–359. [Google Scholar] [CrossRef] [PubMed]
- Abdul Rahim, S.N.; Ho, G.Y.; Coward, J.I. The role of interleukin-6 in malignant mesothelioma. Transl. Lung Cancer Res. 2015, 4, 55–66. [Google Scholar] [CrossRef]
- Yang, H.; Bocchetta, M.; Kroczynska, B.; Elmishad, A.G.; Chen, Y.; Liu, Z.; Bubici, C.; Mossman, B.T.; Pass, H.I.; Testa, J.R.; et al. TNF-alpha inhibits asbestos-induced cytotoxicity via a NF-kappaB-dependent pathway, a possible mechanism for asbestos-induced oncogenesis. Proc. Natl. Acad. Sci. USA 2006, 103, 10397–10402. [Google Scholar] [CrossRef] [Green Version]
- Kishimoto, T.; Fujimoto, N.; Ebara, T.; Omori, T.; Oguri, T.; Niimi, A.; Yokoyama, T.; Kato, M.; Usami, I.; Nishio, M.; et al. Serum levels of the chemokine CCL2 are elevated in malignant pleural mesothelioma patients. BMC Cancer 2019, 19, 1204. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.Y.; Rath, E.M.; Linton, A.; Yuen, M.L.; Takahashi, K.; Lee, K. The Current Understanding of Asbestos-Induced Epigenetic Changes Associated with Lung Cancer. Lung Cancer 2020, 11, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Acencio, M.M.; Soares, B.; Marchi, E.; Silva, C.S.; Teixeira, L.R.; Broaddus, V.C. Inflammatory Cytokines Contribute to Asbestos-Induced Injury of Mesothelial Cells. Lung 2015, 193, 831–837. [Google Scholar] [CrossRef] [PubMed]
- Carbone, M.; Yang, H. Mesothelioma: Recent highlights. Ann. Transl. Med. 2017, 5, 238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Affar, E.B.; Carbone, M. BAP1 regulates different mechanisms of cell death. Cell Death Dis. 2018, 9, 1151. [Google Scholar] [CrossRef] [PubMed]
- Dammeijer, F.; Lievense, L.A.; Kaijen-Lambers, M.E.; van Nimwegen, M.; Bezemer, K.; Hegmans, J.P.; van Hall, T.; Hendriks, R.W.; Aerts, J.G. Depletion of Tumor-Associated Macrophages with a CSF-1R Kinase Inhibitor Enhances Antitumor Immunity and Survival Induced by DC Immunotherapy. Cancer Immunol. Res. 2017, 5, 535–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowak, A.K.; Brosseau, S.; Cook, A.; Zalcman, G. Antiangiogeneic Strategies in Mesothelioma. Front. Oncol. 2020, 10, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, G.J.; van Zandwijk, N.; Rasko, J.E.J. The Immune Microenvironment in Mesothelioma: Mechanisms of Resistance to Immunotherapy. Front. Oncol. 2019, 9, 1366. [Google Scholar] [CrossRef] [PubMed]
- Lizotte, P.H.; Ivanova, E.V.; Awad, M.M.; Jones, R.E.; Keogh, L.; Liu, H.; Dries, R.; Almonte, C.; Herter-Sprie, G.S.; Santos, A.; et al. Multiparametric profiling of non-small-cell lung cancers reveals distinct immunophenotypes. JCI Insight 2016, 1, e89014. [Google Scholar] [CrossRef]
- Wong, R.M.; Ianculescu, I.; Sharma, S.; Gage, D.L.; Olevsky, O.M.; Kotova, S.; Kostic, M.N.; Grundfest, W.S.; Hou, D.; Cameron, R.B. Immunotherapy for malignant pleural mesothelioma. Current status and future prospects. Am. J. Respir. Cell Mol. Biol. 2014, 50, 870–875. [Google Scholar] [CrossRef]
- Wong, R.M. Modulating Immunosuppression in the Intrapleural Space of Malignant Pleural Mesothelioma and Predictive Biomarkers to Guide Treatment Decisions. J. Thorac. Oncol. 2016, 11, 1602–1603. [Google Scholar] [CrossRef] [Green Version]
- Noordam, L.; Kaijen, M.E.H.; Bezemer, K.; Cornelissen, R.; Maat, L.; Hoogsteden, H.C.; Aerts, J.; Hendriks, R.W.; Hegmans, J.; Vroman, H. Low-dose cyclophosphamide depletes circulating naive and activated regulatory T cells in malignant pleural mesothelioma patients synergistically treated with dendritic cell-based immunotherapy. Oncoimmunology 2018, 7, e1474318. [Google Scholar] [CrossRef]
- Klampatsa, A.; O’Brien, S.M.; Thompson, J.C.; Rao, A.S.; Stadanlick, J.E.; Martinez, M.C.; Liousia, M.; Cantu, E.; Cengel, K.; Moon, E.K.; et al. Phenotypic and functional analysis of malignant mesothelioma tumor-infiltrating lymphocytes. Oncoimmunology 2019, 8, e1638211. [Google Scholar] [CrossRef] [Green Version]
- Salaroglio, I.C.; Kopecka, J.; Napoli, F.; Pradotto, M.; Maletta, F.; Costardi, L.; Gagliasso, M.; Milosevic, V.; Ananthanarayanan, P.; Bironzo, P.; et al. Potential Diagnostic and Prognostic Role of Microenvironment in Malignant Pleural Mesothelioma. J. Thorac. Oncol. 2019, 14, 1458–1471. [Google Scholar] [CrossRef] [Green Version]
- Hoves, S.; Ooi, C.H.; Wolter, C.; Sade, H.; Bissinger, S.; Schmittnaegel, M.; Ast, O.; Giusti, A.M.; Wartha, K.; Runza, V.; et al. Rapid activation of tumor-associated macrophages boosts preexisting tumor immunity. J. Exp. Med. 2018, 215, 859–876. [Google Scholar] [CrossRef] [PubMed]
- Lievense, L.A.; Bezemer, K.; Aerts, J.G.; Hegmans, J.P. Tumor-associated macrophages in thoracic malignancies. Lung Cancer 2013, 80, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Forde, P.M.; Scherpereel, A.; Tsao, A.S. Use of Immune Checkpoint Inhibitors in Mesothelioma. Curr. Treat. Options Oncol. 2019, 20, 18. [Google Scholar] [CrossRef]
- Remon, J.; Passiglia, F.; Ahn, M.J.; Barlesi, F.; Forde, P.M.; Garon, E.B.; Gettinger, S.; Goldberg, S.B.; Herbst, R.S.; Horn, L.; et al. Immune Checkpoint Inhibitors in Thoracic Malignancies: Review of the Existing Evidence by an IASLC Expert Panel and Recommendations. J. Thorac. Oncol. 2020. [Google Scholar] [CrossRef] [PubMed]
- de Gooijer, C.J.; Borm, F.J.; Scherpereel, A.; Baas, P. Immunotherapy in Malignant Pleural Mesothelioma. Front. Oncol. 2020, 10, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veltman, J.D.; Lambers, M.E.; van Nimwegen, M.; Hendriks, R.W.; Hoogsteden, H.C.; Hegmans, J.P.; Aerts, J.G. Zoledronic acid impairs myeloid differentiation to tumour-associated macrophages in mesothelioma. Br. J. Cancer 2010, 103, 629–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huaux, F.; d’Ursel de Bousies, V.; Parent, M.A.; Orsi, M.; Uwambayinema, F.; Devosse, R.; Ibouraadaten, S.; Yakoub, Y.; Panin, N.; Palmai-Pallag, M.; et al. Mesothelioma response to carbon nanotubes is associated with an early and selective accumulation of immunosuppressive monocytic cells. Part. Fibre Toxicol. 2016, 13, 46. [Google Scholar] [CrossRef] [Green Version]
- Shvedova, A.A.; Kisin, E.R.; Yanamala, N.; Tkach, A.V.; Gutkin, D.W.; Star, A.; Shurin, G.V.; Kagan, V.E.; Shurin, M.R. MDSC and TGFbeta Are Required for Facilitation of Tumor Growth in the Lungs of Mice Exposed to Carbon Nanotubes. Cancer Res. 2015, 75, 1615–1623. [Google Scholar] [CrossRef] [Green Version]
- Shvedova, A.A.; Tkach, A.V.; Kisin, E.R.; Khaliullin, T.; Stanley, S.; Gutkin, D.W.; Star, A.; Chen, Y.; Shurin, G.V.; Kagan, V.E.; et al. Carbon nanotubes enhance metastatic growth of lung carcinoma via up-regulation of myeloid-derived suppressor cells. Small 2013, 9, 1691–1695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, L.A.; Gao, J.; Wal, R.V.; Gigliotti, A.; Burchiel, S.W.; McDonald, J.D. Pulmonary and systemic immune response to inhaled multiwalled carbon nanotubes. Toxicol. Sci. 2007, 100, 203–214. [Google Scholar] [CrossRef]
- Mitchell, L.A.; Lauer, F.T.; Burchiel, S.W.; McDonald, J.D. Mechanisms for how inhaled multiwalled carbon nanotubes suppress systemic immune function in mice. Nat. Nanotechnol. 2009, 4, 451–456. [Google Scholar] [CrossRef] [PubMed]
- Park, E.J.; Cho, W.S.; Jeong, J.; Yi, J.; Choi, K.; Park, K. Pro-inflammatory and potential allergic responses resulting from B cell activation in mice treated with multi-walled carbon nanotubes by intratracheal instillation. Toxicology 2009, 259, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Park, E.J.; Roh, J.; Kim, S.N.; Kang, M.S.; Lee, B.S.; Kim, Y.; Choi, S. Biological toxicity and inflammatory response of semi-single-walled carbon nanotubes. PLoS ONE 2011, 6, e25892. [Google Scholar] [CrossRef] [PubMed]
- Park, E.J.; Roh, J.; Kim, S.N.; Kang, M.S.; Han, Y.A.; Kim, Y.; Hong, J.T.; Choi, K. A single intratracheal instillation of single-walled carbon nanotubes induced early lung fibrosis and subchronic tissue damage in mice. Arch. Toxicol. 2011, 85, 1121–1131. [Google Scholar] [CrossRef] [PubMed]
- Kido, T.; Tsunoda, M.; Kasai, T.; Sasaki, T.; Umeda, Y.; Senoh, H.; Yanagisawa, H.; Asakura, M.; Aizawa, Y.; Fukushima, S. The increases in relative mRNA expressions of inflammatory cytokines and chemokines in splenic macrophages from rats exposed to multi-walled carbon nanotubes by whole-body inhalation for 13 weeks. Inhal. Toxicol. 2014, 26, 750–758. [Google Scholar] [CrossRef]
- Muller, J.; Delos, M.; Panin, N.; Rabolli, V.; Huaux, F.; Lison, D. Absence of carcinogenic response to multiwall carbon nanotubes in a 2-year bioassay in the peritoneal cavity of the rat. Toxicol. Sci. 2009, 110, 442–448. [Google Scholar] [CrossRef]
- Nagai, H.; Okazaki, Y.; Chew, S.H.; Misawa, N.; Yamashita, Y.; Akatsuka, S.; Ishihara, T.; Yamashita, K.; Yoshikawa, Y.; Yasui, H.; et al. Diameter and rigidity of multiwalled carbon nanotubes are critical factors in mesothelial injury and carcinogenesis. Proc. Natl. Acad. Sci. USA 2011, 108, E1330–E1338. [Google Scholar] [CrossRef] [Green Version]
- Rittinghausen, S.; Hackbarth, A.; Creutzenberg, O.; Ernst, H.; Heinrich, U.; Leonhardt, A.; Schaudien, D. The carcinogenic effect of various multi-walled carbon nanotubes (MWCNTs) after intraperitoneal injection in rats. Part. Fibre Toxicol. 2014, 11, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poland, C.A.; Byrne, F.; Cho, W.S.; Prina-Mello, A.; Murphy, F.A.; Davies, G.L.; Coey, J.M.; Gounko, Y.; Duffin, R.; Volkov, Y.; et al. Length-dependent pathogenic effects of nickel nanowires in the lungs and the peritoneal cavity. Nanotoxicology 2012, 6, 899–911. [Google Scholar] [CrossRef] [PubMed]
- Duke, K.S.; Taylor-Just, A.J.; Ihrie, M.D.; Shipkowski, K.A.; Thompson, E.A.; Dandley, E.C.; Parsons, G.N.; Bonner, J.C. STAT1-dependent and -independent pulmonary allergic and fibrogenic responses in mice after exposure to tangled versus rod-like multi-walled carbon nanotubes. Part. Fibre Toxicol. 2017, 14, 26. [Google Scholar] [CrossRef] [PubMed]
- Di Cristo, L.; Bianchi, M.G.; Chiu, M.; Taurino, G.; Donato, F.; Garzaro, G.; Bussolati, O.; Bergamaschi, E. Comparative in Vitro Cytotoxicity of Realistic Doses of Benchmark Multi-Walled Carbon Nanotubes towards Macrophages and Airway Epithelial Cells. Nanomaterials 2019, 9, 982. [Google Scholar] [CrossRef] [Green Version]
- Rydman, E.M.; Ilves, M.; Vanhala, E.; Vippola, M.; Lehto, M.; Kinaret, P.A.; Pylkkanen, L.; Happo, M.; Hirvonen, M.R.; Greco, D.; et al. A Single Aspiration of Rod-like Carbon Nanotubes Induces Asbestos-like Pulmonary Inflammation Mediated in Part by the IL-1 Receptor. Toxicol. Sci. 2015, 147, 140–155. [Google Scholar] [CrossRef] [Green Version]
- Catalan, J.; Siivola, K.M.; Nymark, P.; Lindberg, H.; Suhonen, S.; Jarventaus, H.; Koivisto, A.J.; Moreno, C.; Vanhala, E.; Wolff, H.; et al. In vitro and in vivo genotoxic effects of straight versus tangled multi-walled carbon nanotubes. Nanotoxicology 2016, 10, 794–806. [Google Scholar] [CrossRef]
- Sakamoto, Y.; Hojo, M.; Kosugi, Y.; Watanabe, K.; Hirose, A.; Inomata, A.; Suzuki, T.; Nakae, D. Comparative study for carcinogenicity of 7 different multi-wall carbon nanotubes with different physicochemical characteristics by a single intraperitoneal injection in male Fischer 344 rats. J. Toxicol. Sci. 2018, 43, 587–600. [Google Scholar] [CrossRef] [Green Version]
- Hwang, B.; Lee, J.H.; Bang, D. Single-cell RNA sequencing technologies and bioinformatics pipelines. Exp. Mol. Med. 2018, 50, 96. [Google Scholar] [CrossRef] [Green Version]
- Doran, A.C.; Yurdagul, A., Jr.; Tabas, I. Efferocytosis in health and disease. Nat. Rev. Immunol. 2020, 20, 254–267. [Google Scholar] [CrossRef]
- Guilliams, M.; Thierry, G.R.; Bonnardel, J.; Bajenoff, M. Establishment and Maintenance of the Macrophage Niche. Immunity 2020, 52, 434–451. [Google Scholar] [CrossRef]
- Schyns, J.; Bai, Q.; Ruscitti, C.; Radermecker, C.; De Schepper, S.; Chakarov, S.; Farnir, F.; Pirottin, D.; Ginhoux, F.; Boeckxstaens, G.; et al. Non-classical tissue monocytes and two functionally distinct populations of interstitial macrophages populate the mouse lung. Nat. Commun. 2019, 10, 3964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bottek, J.; Soun, C.; Lill, J.K.; Dixit, A.; Thiebes, S.; Beerlage, A.L.; Horstmann, M.; Urbanek, A.; Heuer, H.; Uszkoreit, J.; et al. Spatial proteomics revealed a CX3CL1-dependent crosstalk between the urothelium and relocated macrophages through IL-6 during an acute bacterial infection in the urinary bladder. Mucosal. Immunol. 2020. [Google Scholar] [CrossRef]
- Bogaert, A.; Fernandez, E.; Gevaert, K. N-Terminal Proteoforms in Human Disease. Trends Biochem. Sci. 2020, 45, 308–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Chen, L.; Lu, X.; Huang, C.; Luo, H.; Jin, J.; Mei, Z.; Liu, J.; Liu, C.; Shi, J.; et al. Data-Independent Acquisition-Based Quantitative Proteomics Analysis Reveals Dynamic Network Profiles during the Macrophage Inflammatory Response. Proteomics 2020, 20, e1900203. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Historical progression of CNT-induced toxicity. Timeline summarizing the discoveries on CNT toxicity obtained by LTAP teams (UCLouvain, Brussels, Belgium) and their scientific collaborators from around the world. For this collective work, we used diverse and relevant in vivo and in vitro models and CNT possessing diverse morphological and physico-chemical properties.
Figure 1.
Historical progression of CNT-induced toxicity. Timeline summarizing the discoveries on CNT toxicity obtained by LTAP teams (UCLouvain, Brussels, Belgium) and their scientific collaborators from around the world. For this collective work, we used diverse and relevant in vivo and in vitro models and CNT possessing diverse morphological and physico-chemical properties.

Figure 2.
A new pathological pathway governs carcinogenesis induced by mesotheliomagenic needled CNT-N. Persistent inflammation and immunosuppression orchestrate carcinogenesis and mesothelioma. Toxic CNT-N induce an inflammatory cascade (in blue) resulting in the influx of inflammatory macrophages and neutrophils. Sustained production of free radicals by these activated immune cells induces irreversible DNA damage. Pro-inflammatory cytokines are also considered as potent polypeptide growth factors for transformed mesothelial cells and angiogenesis. An unexpected conjoint immunosuppression (in red) is induced by mesotheliomagenic CNT. These early responses to CNT-N are characterized by persistent accumulation of immunosuppressive macrophages and monocytes and a sustained production of regulatory cytokines (i.e., IL-10 and TGF-β). These immunoregulatory components are incriminated in carcinogenesis by preventing host immune responses directed against transformed cells and favoring tumor growth.
Figure 2.
A new pathological pathway governs carcinogenesis induced by mesotheliomagenic needled CNT-N. Persistent inflammation and immunosuppression orchestrate carcinogenesis and mesothelioma. Toxic CNT-N induce an inflammatory cascade (in blue) resulting in the influx of inflammatory macrophages and neutrophils. Sustained production of free radicals by these activated immune cells induces irreversible DNA damage. Pro-inflammatory cytokines are also considered as potent polypeptide growth factors for transformed mesothelial cells and angiogenesis. An unexpected conjoint immunosuppression (in red) is induced by mesotheliomagenic CNT. These early responses to CNT-N are characterized by persistent accumulation of immunosuppressive macrophages and monocytes and a sustained production of regulatory cytokines (i.e., IL-10 and TGF-β). These immunoregulatory components are incriminated in carcinogenesis by preventing host immune responses directed against transformed cells and favoring tumor growth.

Figure 3.
Mesotheliomagenic CNT-N and non-mesotheliomagenic CNT-T induce early comparable peritoneal lesions and macrophage accumulation in rats. Wistar rats untreated or injected (i.p) with CNT-N or CNT-T (2 mg) were sacrificed (day 15) and peritoneal tissues (diaphragm) were harvested, fixed in paraformaldehyde and embedded in paraffin. 5 µm sections were stained with classical H&E coloration (A-D-G for controls, B-E-H for CNT-N and C-F-I for CNT-T, magnification ×4 first line, ×40 other lines). The red arrows indicate granulomas containing CNT crystalline structures within the connective tissue (selected from the frame of A-B-C panels). Granulomas mainly comprise macrophages around nanotube aggregates (G-H-I). For macrophage identification, 5 µm sections were incubated with mouse anti-rat CD68 antibody (Abcam monoclonal) and secondary antibody donkey anti-mouse (Jackson ImmunoReserch) coupled with HRP. After incubation with AlexaFluor Tyramide 488, a counterstaining with Hoechst44432 dye was performed. Stained slides were digitalized using a Panoramic 250 FlashIII scanner (3DHistech) at ×20 magnification.
Figure 3.
Mesotheliomagenic CNT-N and non-mesotheliomagenic CNT-T induce early comparable peritoneal lesions and macrophage accumulation in rats. Wistar rats untreated or injected (i.p) with CNT-N or CNT-T (2 mg) were sacrificed (day 15) and peritoneal tissues (diaphragm) were harvested, fixed in paraformaldehyde and embedded in paraffin. 5 µm sections were stained with classical H&E coloration (A-D-G for controls, B-E-H for CNT-N and C-F-I for CNT-T, magnification ×4 first line, ×40 other lines). The red arrows indicate granulomas containing CNT crystalline structures within the connective tissue (selected from the frame of A-B-C panels). Granulomas mainly comprise macrophages around nanotube aggregates (G-H-I). For macrophage identification, 5 µm sections were incubated with mouse anti-rat CD68 antibody (Abcam monoclonal) and secondary antibody donkey anti-mouse (Jackson ImmunoReserch) coupled with HRP. After incubation with AlexaFluor Tyramide 488, a counterstaining with Hoechst44432 dye was performed. Stained slides were digitalized using a Panoramic 250 FlashIII scanner (3DHistech) at ×20 magnification.

Figure 4.
New opportunities to use CNT for delineating specific macrophage immune pathways specifically associated with malignant mesothelioma. Cellular and molecular characterization of macrophage subpopulations by using next-generation sequencing (NGS) technologies. Peritoneal macrophages from CNT-N or CNT-T-treated rats (day 15) were isolated from peritoneal cell suspensions using flow cytometry cell sorting (FACSAria III, BD Biosciences) and APC-antibodies specific for CD68 (mouse anti-rat CD68 antibody, Abcam monoclonal). Cytocentrifuge preparations of purified macrophages were stained with Diff-Quick. RNA was isolated using Qiagen kits and libraries were prepared and sequenced using the Illumina platform. The gene count matrix was transformed in fold-change-related tables or barcode plots.
Figure 4.
New opportunities to use CNT for delineating specific macrophage immune pathways specifically associated with malignant mesothelioma. Cellular and molecular characterization of macrophage subpopulations by using next-generation sequencing (NGS) technologies. Peritoneal macrophages from CNT-N or CNT-T-treated rats (day 15) were isolated from peritoneal cell suspensions using flow cytometry cell sorting (FACSAria III, BD Biosciences) and APC-antibodies specific for CD68 (mouse anti-rat CD68 antibody, Abcam monoclonal). Cytocentrifuge preparations of purified macrophages were stained with Diff-Quick. RNA was isolated using Qiagen kits and libraries were prepared and sequenced using the Illumina platform. The gene count matrix was transformed in fold-change-related tables or barcode plots.

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Orsi, M.; Al Hatem, C.; Leinardi, R.; Huaux, F. Carbon Nanotubes under Scrutiny: Their Toxicity and Utility in Mesothelioma Research. Appl. Sci. 2020, 10, 4513. https://doi.org/10.3390/app10134513
AMA Style
Orsi M, Al Hatem C, Leinardi R, Huaux F. Carbon Nanotubes under Scrutiny: Their Toxicity and Utility in Mesothelioma Research. Applied Sciences. 2020; 10(13):4513. https://doi.org/10.3390/app10134513
Chicago/Turabian StyleOrsi, Micaela, Chafik Al Hatem, Riccardo Leinardi, and François Huaux. 2020. "Carbon Nanotubes under Scrutiny: Their Toxicity and Utility in Mesothelioma Research" Applied Sciences 10, no. 13: 4513. https://doi.org/10.3390/app10134513
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.