A Protocol for Producing Virus-Free Artichoke Genetic Resources for Conservation, Breeding, and Production

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
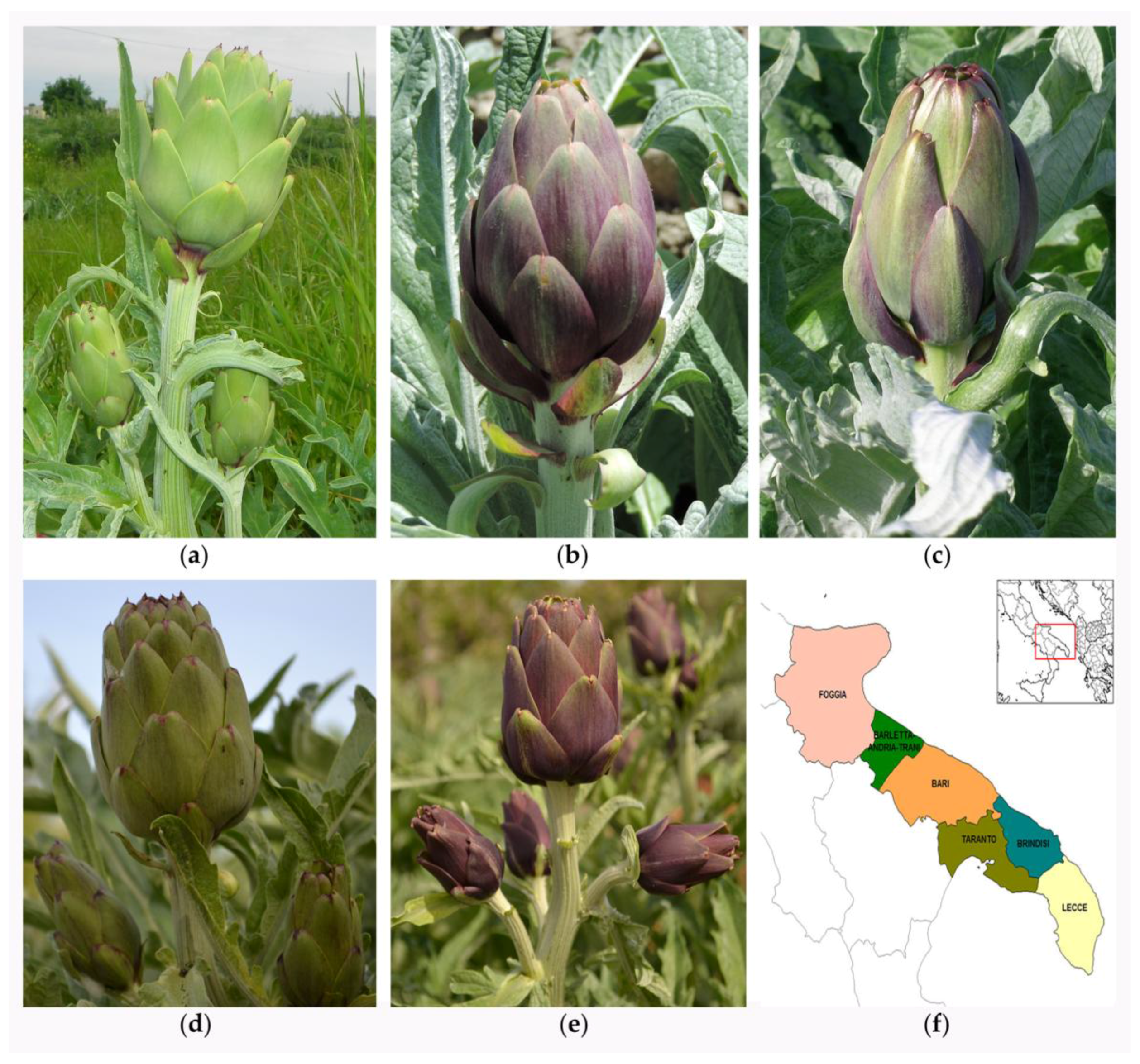
2.1. Plant Material and Assessment of the Sanitary Status
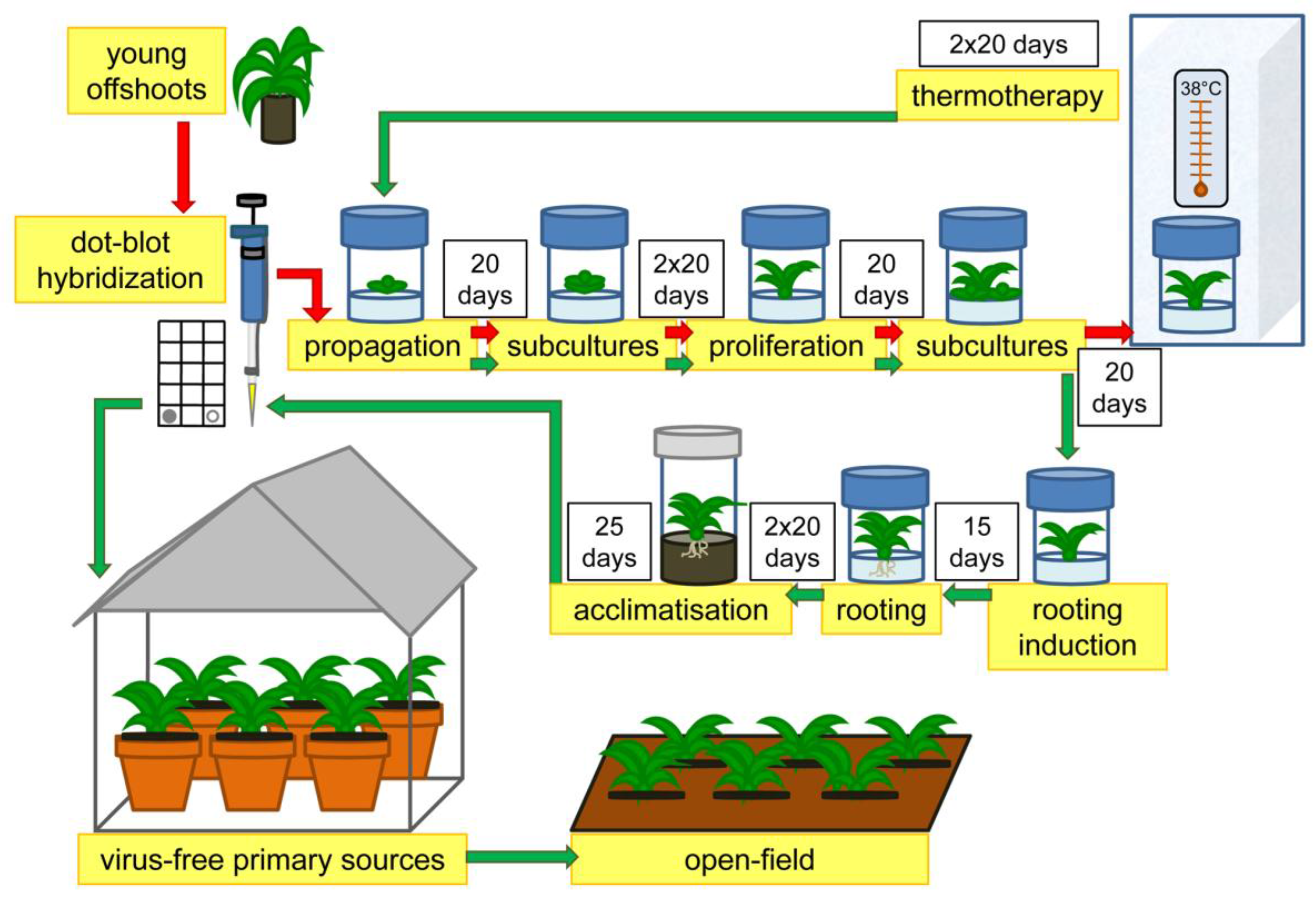
2.2. Plant Sanitation Protocol
3. Results
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Romani, A.; Pinelli, P.; Cantini, C.; Cimato, A.; Heimler, D. Characterization of Violetto di Toscana, a typical Italian variety of artichoke (Cynara scolymus L.). Food Chem. 2006, 95, 221–225. [Google Scholar] [CrossRef]
- Mauro, R.P.; Portis, E.; Acquadro, A.; Lombardo, S.; Mauromicale, G.; Lanteri, S. Genetic diversity of globe artichoke landraces from Sicilian small-holdings: Implications for evolution and domestication of the species. Conserv. Gene 2009, 10, 431–440. [Google Scholar] [CrossRef]
- Mauro, R.P.; Portis, E.; Lanteri, S.; Mauromicale, G. Genotypic and bio-agronomical characterization of an early Sicilian landrace of globe artichoke. Euphytica 2012, 186, 357–366. [Google Scholar] [CrossRef]
- Ciancolini, A.; Rey, N.A.; Pagnotta, M.A.; Crinò, P. Characterization of Italian spring globe artichoke germplasm: Morphological and molecular profiles. Euphytica 2012, 186, 433–443. [Google Scholar] [CrossRef]
- Tavazza, R.; Lucioli, A.; Benelli, C.; Giorgi, D.; D’Aloisio, E.; Papacchioli, V. Cryopreservation in artichoke: Towards a phytosanitary qualified germplasm collection. Ann. Appl. Biol. 2013, 163, 231–241. [Google Scholar] [CrossRef]
- Elia, A.; Santamaria, P. Biodiversity in vegetable crops, a heritage to save: The case of Puglia region. Ital. J. Agron. 2013, 8, 21–34. [Google Scholar] [CrossRef]
- Gallitelli, D.; Mascia, T.; Martelli, G.P. Viruses in Artichoke. Adv. Virus Res. 2012, 84, 289–324. [Google Scholar] [PubMed]
- Virgin, H.W. The virome in mammalian physiology and disease. Cell 2014, 157, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Tollenaere, C.; Susi, H.; Laine, A.L. Evolutionary and epidemiological implications of multiple infection in plants. Trends Plant Sci. 2015, 21, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Mascia, T.; Gallitelli, D. Synergies and antagonisms in virus interactions. Plant Sci. 2016, 252, 176–192. [Google Scholar] [CrossRef] [PubMed]
- Minutillo, S.A.; Mascia, T.; Gallitelli, D. A DNA probe mix for the multiplex detection of ten artichoke viruses. Eur. J. Plant Pathol. 2012, 134, 459–465. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Fortunato, I.M.; Ruta, C.; Castrignano, A.; Saccardo, A. The effect of mycorrhizal symbiosis on the development of micropropagated artichokes. Sci. Hortic. 2005, 106, 472–483. [Google Scholar] [CrossRef]
- Nitsch, J.P.; Nitsch, C. Haploid plants for pollen grains. Science 1969, 63, 85–87. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, G.; McCown, B. Commercially-feasible micropropagation of mountain laurel, Kalmia latifolia, by use of shoot-tip culture. Int. Plant Prop. Soc. Proc. 1980, 30, 421–427. [Google Scholar]
- Hanssen, I.M.; Lapidot, M.; Thomma, P.H.J.B. Emerging viral diseases of tomato crops. Mol. Plant Microbe Interact. 2010, 23, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Hanssen, I.M.; Lapidot, M. Major tomato viruses in the Mediterranean basin. Adv. Virus Res. 2012, 84, 31–66. [Google Scholar] [PubMed]
- Vaira, A.M.; Accotto, G.P.; Vecchiati, M.; Bragaloni, M. Tomato infectious chlorosis virus causes leaf yellowing and reddening of tomato in Italy. Phytoparasitica 2002, 30, 290–294. [Google Scholar] [CrossRef]
- Davino, S.; Tomassoli, L.; Tiberini, A.; Mondello, V.; Davino, M. Outbreak of tomato infectious chlorosis virus in a relevant artichoke producing area of Sicily. J. Plant Pathol. 2009, 91, S4.57–S4.58. [Google Scholar]
- Spanò, R.; Mascia, T.; Minutillo, S.A.; Gallitelli, D. First report of Tomato infectious chlorosis virus from tomato in Apulia, southern Italy. J. Plant Pathol. 2011, 93. [Google Scholar] [CrossRef]
- Spanò, R.; Trisciuzzi, N.; Gallitelli, D. Gravi infezioni di Tomato infectious chlorosis virus su pomodoro in Puglia. Prot. Colt. 2015, 4, 43–45. [Google Scholar]
- López-Pérez, A.J.; Martínez, J.A. In vitro root induction improvement by culture in darkness for different globe artichoke cultivars. In Vitro Cell. Dev. Biol. Plant 2015, 51, 160–165. [Google Scholar] [CrossRef]
- Acquadro, A.; Papanice, M.A.; Lanteri, S.; Bottalico, G.; Portis, E.; Campanale, A.; Finetti-Sialer, M.M.; Mascia, T.; Sumerano, P.; Gallitelli, D. Production and fingerprinting of virus-free clones in a reflowering globe artichoke. Plant Cell Tissue Organ Cult. 2010, 100, 329–337. [Google Scholar] [CrossRef]
- Pecaut, P.; Martin, F. Variation occuring after natural and in vitro multiplication of early Mediterranean cultivars of globe artichoke (Cynara scolymus L.). Agronomie 1993, 13, 909–919. [Google Scholar] [CrossRef]
- Walkey, D.G.A.; Webb, M.J. Virus in plant apical meristem. J. Gen. Virol. 1968, 3, 311–313. [Google Scholar] [CrossRef]
- Appiano, A.; Pennazio, S. Electron microscopy of potato meristem tips infected with potato virus X. J. Gen. Virol. 1972, 14, 273–276. [Google Scholar] [CrossRef] [PubMed]
- Mori, K.; Hosokawa, D. Localization of viruses in apical meristem and production of virus-free plants by means of meristem and tissue culture. Acta Hortic. 1977, 78, 389–396. [Google Scholar] [CrossRef]
- Wang, Q.; Cuellar, W.J.; Rajamäki, M.L.; Hirata, Y.; Valkonen, J.P.T. Combined thermotherapy and cryotherapy for efficient virus eradication: Relation of virus distribution, subcellular changes, cell survival and viral RNA degradation in shoot tips. Mol. Plant Pathol. 2008, 9, 237–250. [Google Scholar] [CrossRef] [PubMed]
- Goshal, B.; Sanfaçon, H. Symptom recovery in virus-infected plants: Revisiting the role of RNA silencing mechanisms. Virology 2015, 479–480, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Santovito, E.; Mascia, T.; Siddiqui, S.A.; Minutillo, S.A.; Valkonen, J.P.T.; Gallitelli, D. Infection cycle of artichoke italian latent virus in tobacco plants: Meristem invasion and recovery from disease symptoms. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Walkey, D.G.A.; Cooper, V.C. Effect of temperature on virus eradication and growth of infected tissue cultures. Ann. Appl. Biol. 1975, 80, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Cooper, V.C.; Walkey, D.G.A. Thermal inactivation of cherry leaf roll virus in tissue cultures of Nicotiana rustica raised from seeds and meristem tips. Ann. Appl. Biol. 1978, 88, 273–278. [Google Scholar] [CrossRef]
- Tang, L.P.; Li, X.M.; Dong, Y.X.; Zhang, X.; Sand Su, Y.H. Microfilament depolymerization is a pre-requisite for stem cell formation during in vitro shoot regeneration in Arabidopsis. Front. Plant. Sci. 2017, 8, 158. [Google Scholar] [CrossRef] [PubMed]
- Benoit, H.; Ducreux, G. Etude de quelques aspects de la multiplication végétative in vitro de l’artichaut (Cynara scolymus L.). Agronomie 1981, 1, 225–230. [Google Scholar] [CrossRef]
- Morzadec, J.M.; Hourmant, A. In vitro rooting improvement of globe artichoke (cv Camus de Bretagne) by GA3. Sci. Hortic. 1997, 72, 59–62. [Google Scholar] [CrossRef]
- Bottalico, G.; Padula, M.; Campanale, A.; Finetti Sialer, M.M.; Saccomanno, F.; Gallitelli, D. Seed transmission of Artichoke Italian latent virus and Artichoke latent virus in globe artichoke. J. Plant Pathol. 2002, 84, 167–168. [Google Scholar]
- Massart, S.; Olmos, A.; Jijakli, H.; Candresse, T. Current impact and future directions of high throughput sequencing in plant virus diagnostics. Virus Res. 2014, 188, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Massart, S.; Candresse, T.; Gil, J.; Lacomme, C.; Predajna, L.; Ravnikar, M.; Reynard, J.S.; Rumbou, A.; Saldarelli, P.; Škorić Vainio, E.J.; et al. A framework for the evaluation of biosecurity, commercial, regulatory, and scientific impacts of plant viruses and viroids identified by NGS technologies. Front. Microbiol. 2017, 8, 45. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Ding, S.W.; Zhang, Y.; Zhu, S. Identification of viruses and viroids by next-generation sequencing and homology-dependentand homology-independent algorithms. Annu. Rev. Phytopathol. 2015, 53, 425–444. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.R.; Constable, F.; Tzanetakis, I.E. Quarantine regulations and the impact of modern detection methods. Annu. Rev. Phytopathol. 2016, 54, 189–205. [Google Scholar] [CrossRef] [PubMed]
- Esquinas-Alcázar, J. Protecting crop genetic diversity for food security: Political, ethical and technical challenges. Nat. Rev. Genet. 2005, 6, 946–953. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Viruses with Isometric Particles | Genus Family | Vector/Mode of Transmission | Disease Symptoms |
| artichoke Aegean ringspot virus (AARSV) | Nepovirus (Subgroup A) Comoviridae | unknown | Yellow blotches and mild mottling. Often symptomless |
| artichoke Italian latent virus (AILV) 1 | Nepovirus (Subgroup B) Comoviridae | Longidorus apulus L. fasciatus Seed coat and expanded cotyledons | Mostly symptomless; patchy chlorotic stunting observed in some cvs |
| artichoke mottled crinkle virus (AMCV) 1 | Tombusvirus Tombusviridae | Contact Soil-borne | Severe deformations, mottling and crinkling of the leaves. Severe distortion of flower capitula. Plant death. New foliage emerging from underground buds develops poorly and often shows bright chrome-yellow discoloration |
| artichoke yellow ringspot virus (AYRSV) | Nepovirus (Subgroup C) Comoviridae | Pollen and seed transmission in tobacco and onion | Bright yellow blotches, ringspots and line patterns sometime followed by necrosis |
| artichoke vein banding virus (AVBV) | Cheravirus Secoviridae | unknown | Chlorotic discolorations along the leaf veins |
| broad bean wilt virus (BBWV) 1 | Fabavirus Comoviridae | Capitophorus horni and other aphid species | Yellow mottle, mosaic, or line patterns |
| cucumber mosaic virus (CMV) 1 | Cucumovirus Bromoviridae | several aphid species | Probably symptomless. Usually found in mixed infection with ArLV and/or TSWV in plants showing severe stunting |
| pelargonium zonate spot virus (PZSV) 1 | Anulavirus Bromoviridae | several thrips species | Moderate stunting and leaf chlorotic mottling. Chlorotic spots and line patterns after inoculation under experimental conditions |
| tobacco streak virus (TSV) | Ilarvirus (Subgroup 1) Bromoviridae | several thrips species | Stunting and leaf deformation |
| tomato black ring virus (TBRV) | Nepovirus (Subgroup B) Comoviridae | Longidorus attenuatus | Mild leaf mottling |
| Viruses with Rod-Shaped Particles | Genus Family | Vector/Mode of Transmission | Disease Symptoms |
| tobacco mosaic virus (TMV) 1 | Tobamovirus | contact | symptomless |
| tobacco rattle virus (TRV) | Tobravirus | Thrichodorus Christiei | Leaf bright yellow discoloration |
| Viruses with Filamentous Particles | Genus Family | Vector/Mode of Transmission | Disease Symptoms |
| artichoke curly dwarf virus (ACDV) | Potexvirus Flexiviridae | unknown | Stunting, leaf distortion and extended vein necrosis, delay in the development of flower capitula |
| Artichoke degeneration virus (ADV). Putative potyvirus | Non-classified | unknown | Leaf mottling curling and crinkling |
| Artichoke latent virus (ArLV) 1 | Macluravirus Potyviridae | Myzus persicae, Brachycaudus cardui, Aphis fabae | Symptomless |
| artichoke latent M virus (ArLMV) | Carlavirus Flexiviridae | unknown | Symptomless. Often in mixed infection with AILV, ArLV, AMCV |
| artichoke latent S virus (ArLSV) | Carlavirus Flexiviridae | aphids | Symptomless |
| bean yellow mosaic virus (BYMV) 1 | Potyvirus Potyviridae | several aphid species | Leaf yellowing, yellow flacking, line patterns, often in mixed infection with ArLV |
| Filamentous viruses and globe artichoke disease | unknown | unknown | Foliar mottling, leaf deformation and crinkling, stunting, and decreased yield. Progressive degeneration |
| potato virus X (PVX) 1 | Potexvirus Flexiviridae | unknown | Mosaic, leaf deformation and narrowing, stunting |
| ranunculus latent virus (RaLV) | Mostly symptomless. Often in mixed infection with cynara 42 virus (Cy42) | ||
| turnip mosaic virus (TuMV) 1 | Potyviridae Potyvirus | several aphid species | Symptomless. Often in mixed infection with ArLV |
| tomato infectious chlorosis virus (TICV) 1 | Crinivirus Closteroviridae | Trialeurodes vaporariorum | Symptomless or mild interveinal yellowing |
| Unnamed putative carlavirus | Non-classified | unknown | Symptomless |
| Viruses with Enveloped Particles | Genus Family | Vector/Mode of Transmission | Disease Symptoms |
| cynara virus (CraV). Putative cytorhabdovirus | unassigned Rhabdoviridae | unknown | Progressive degeneration. Often in mixed infection with AILV, ArLV, BBW, BYMV |
| tomato spotted wilt virus (TSWV) 1 | Tospovirus Bunyaviridae | several thrips species | severe stunting and deformation, generalized chlorosis and bronzing of the apical leaves, distortion of the head stalk, and necrosis of portions of the inner and outer scales |
| Ecotype | Standard Material 1 | Virus Detected 2 | Stabilized Shoots 3 | MR 4 | RC 5 | Primary Sources 6 |
|---|---|---|---|---|---|---|
| Bianco di Taranto | 5 | ArLV | 3 | 2.8 | 68.9% | 6 |
| Francesina | 5 | - | 1 | 3 | 12.5% | 1 |
| Verde di Putignano | 5 | TICV | 2 | 2.7 | 89.7% | 9 |
| Violetto di Putignano | 5 | TICV | 1 | 3.2 | 63.6% | 7 |
| Locale di Mola | 5 | ArLV, AILV | 2 | 7 | 15.4% | 2 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spanò, R.; Bottalico, G.; Corrado, A.; Campanale, A.; Di Franco, A.; Mascia, T. A Protocol for Producing Virus-Free Artichoke Genetic Resources for Conservation, Breeding, and Production. Agriculture 2018, 8, 36. https://doi.org/10.3390/agriculture8030036
Spanò R, Bottalico G, Corrado A, Campanale A, Di Franco A, Mascia T. A Protocol for Producing Virus-Free Artichoke Genetic Resources for Conservation, Breeding, and Production. Agriculture. 2018; 8(3):36. https://doi.org/10.3390/agriculture8030036
Chicago/Turabian StyleSpanò, Roberta, Giovanna Bottalico, Addolorata Corrado, Antonia Campanale, Alessandra Di Franco, and Tiziana Mascia. 2018. "A Protocol for Producing Virus-Free Artichoke Genetic Resources for Conservation, Breeding, and Production" Agriculture 8, no. 3: 36. https://doi.org/10.3390/agriculture8030036