Genome-Wide Association Study Using a Multiparent Advanced Generation Intercross (MAGIC) Population Identified QTLs and Candidate Genes to Predict Shoot and Grain Zinc Contents in Rice

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Plant Growth Condition
2.3. Measurement of Zinc Concentration
2.4. SNP Genotyping and Association Analysis
2.5. RNA Extraction and Real-Time PCR
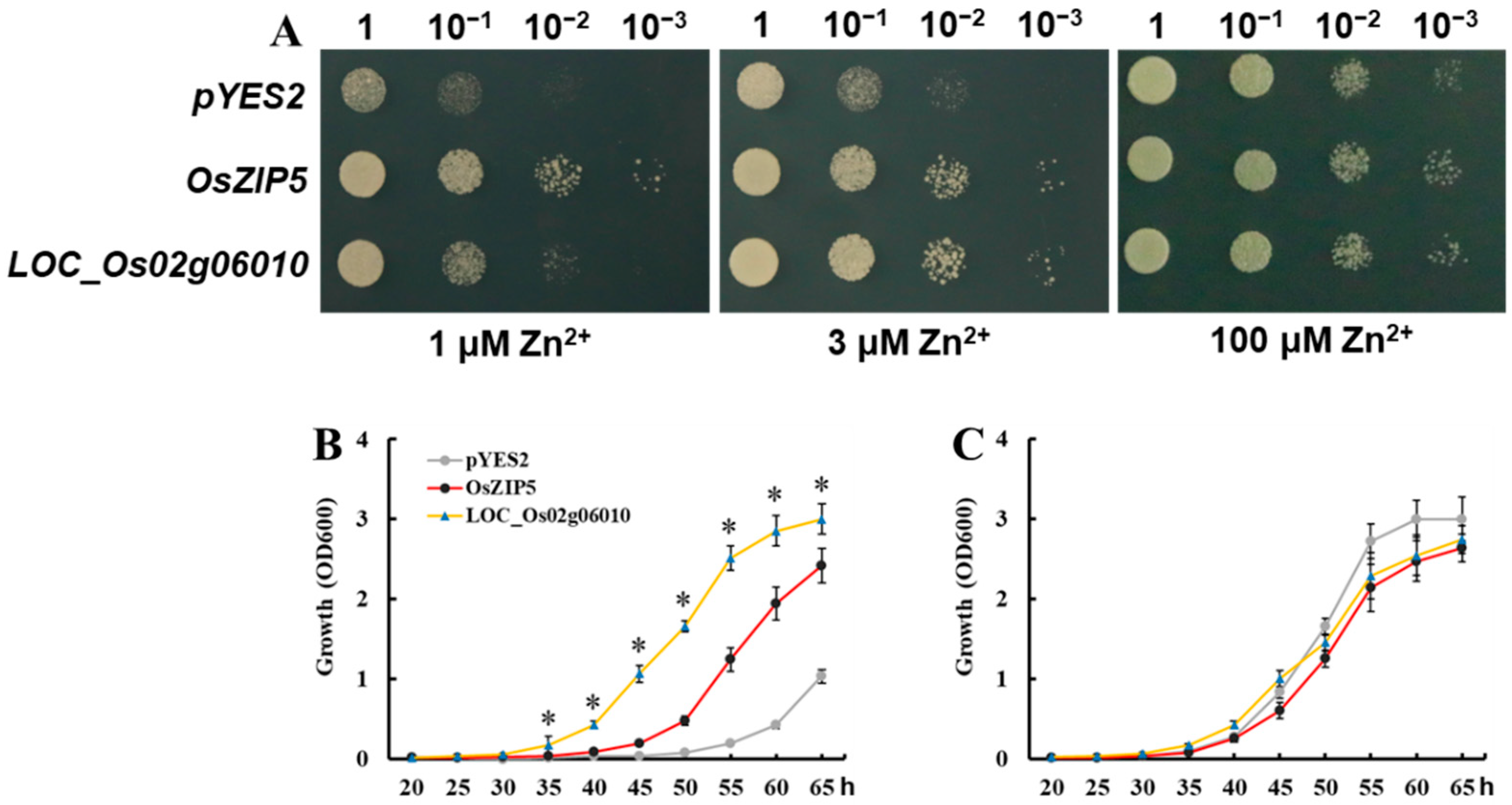
2.6. Expression of Candidate Genes in Yeast
2.7. Sequence Analysis of LOC_Os02g06010 in Four Parental Lines and 30 Other Rice Varieties
3. Results
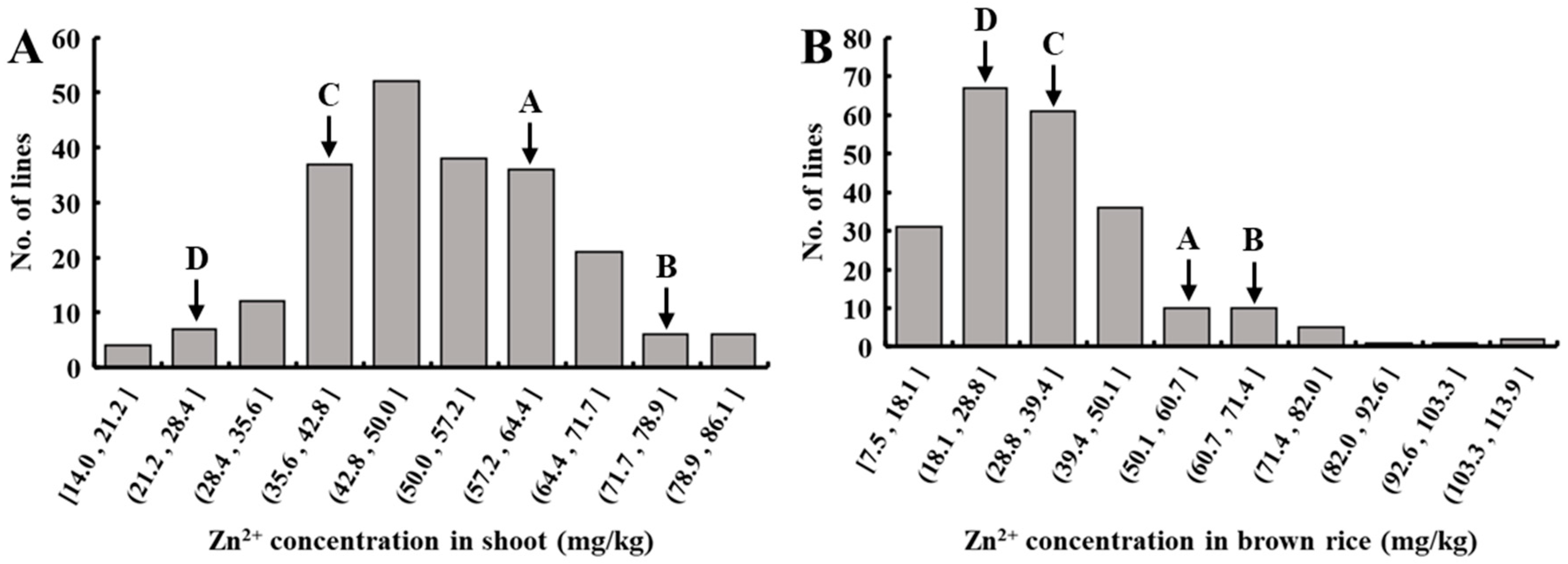
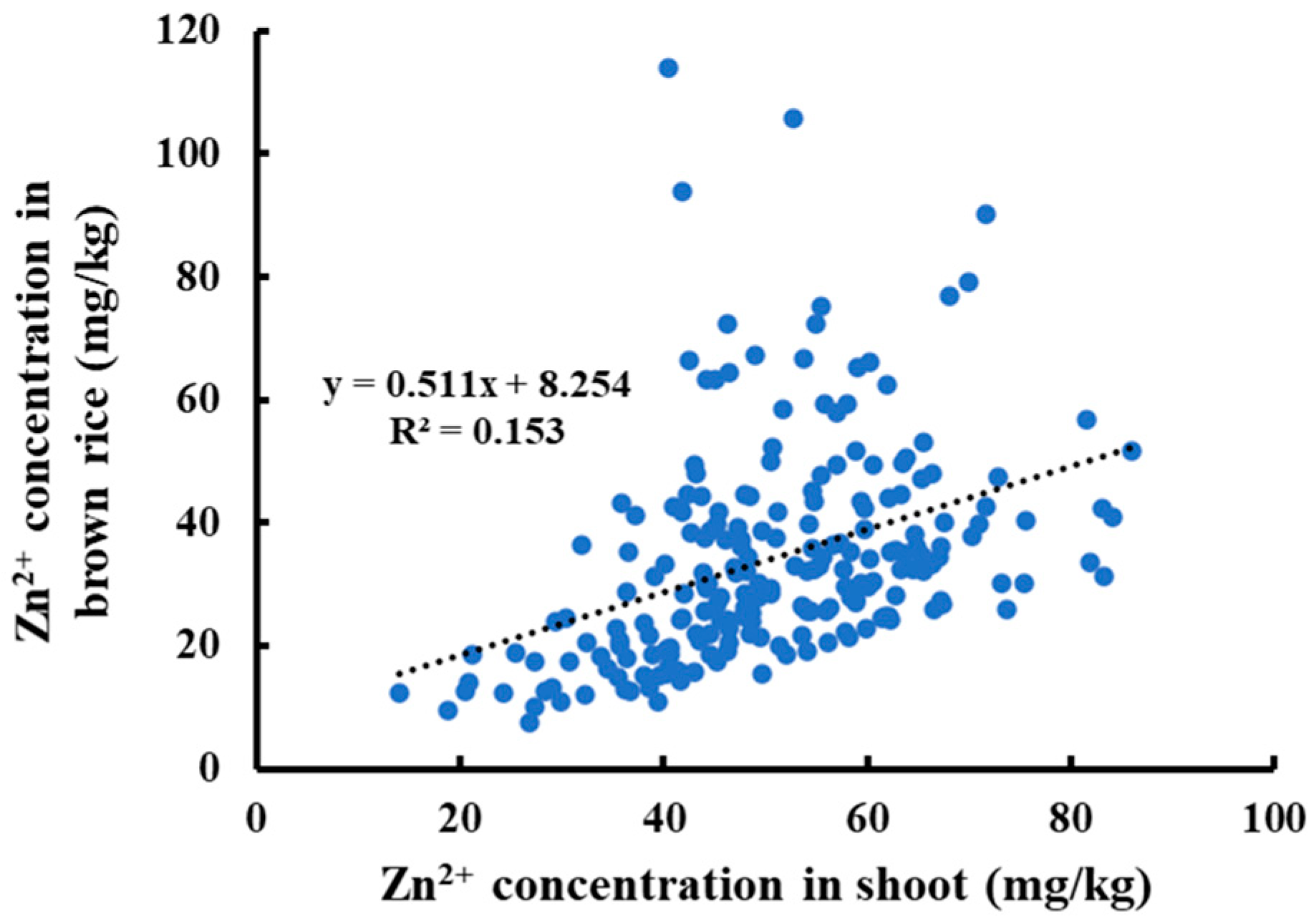
3.1. Distribution of Zn2+ Concentration in Shoots and Grains of MAGIC Population
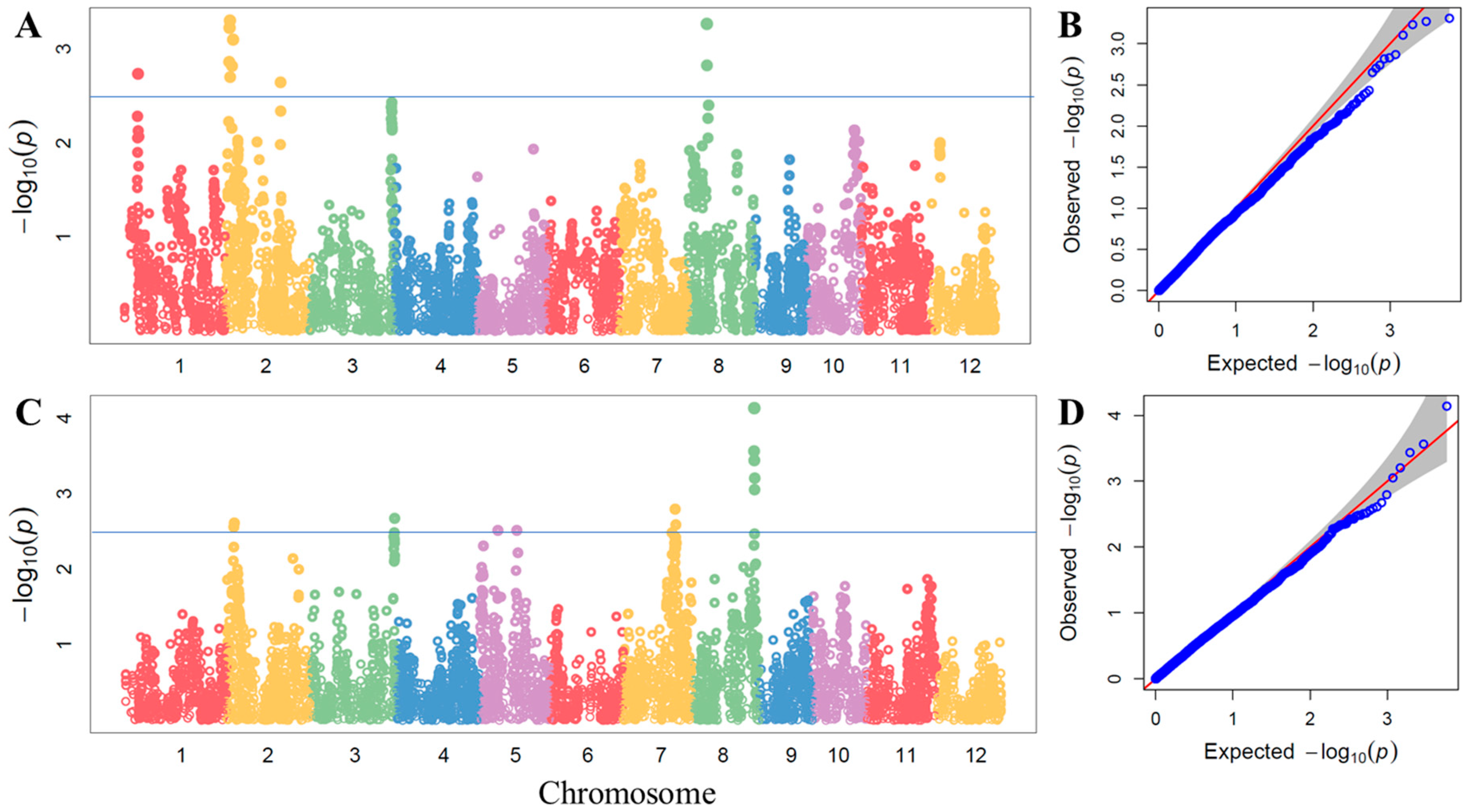
3.2. Mapping of QTLs Associated with Zn2+ Concentration in Shoots and Grains
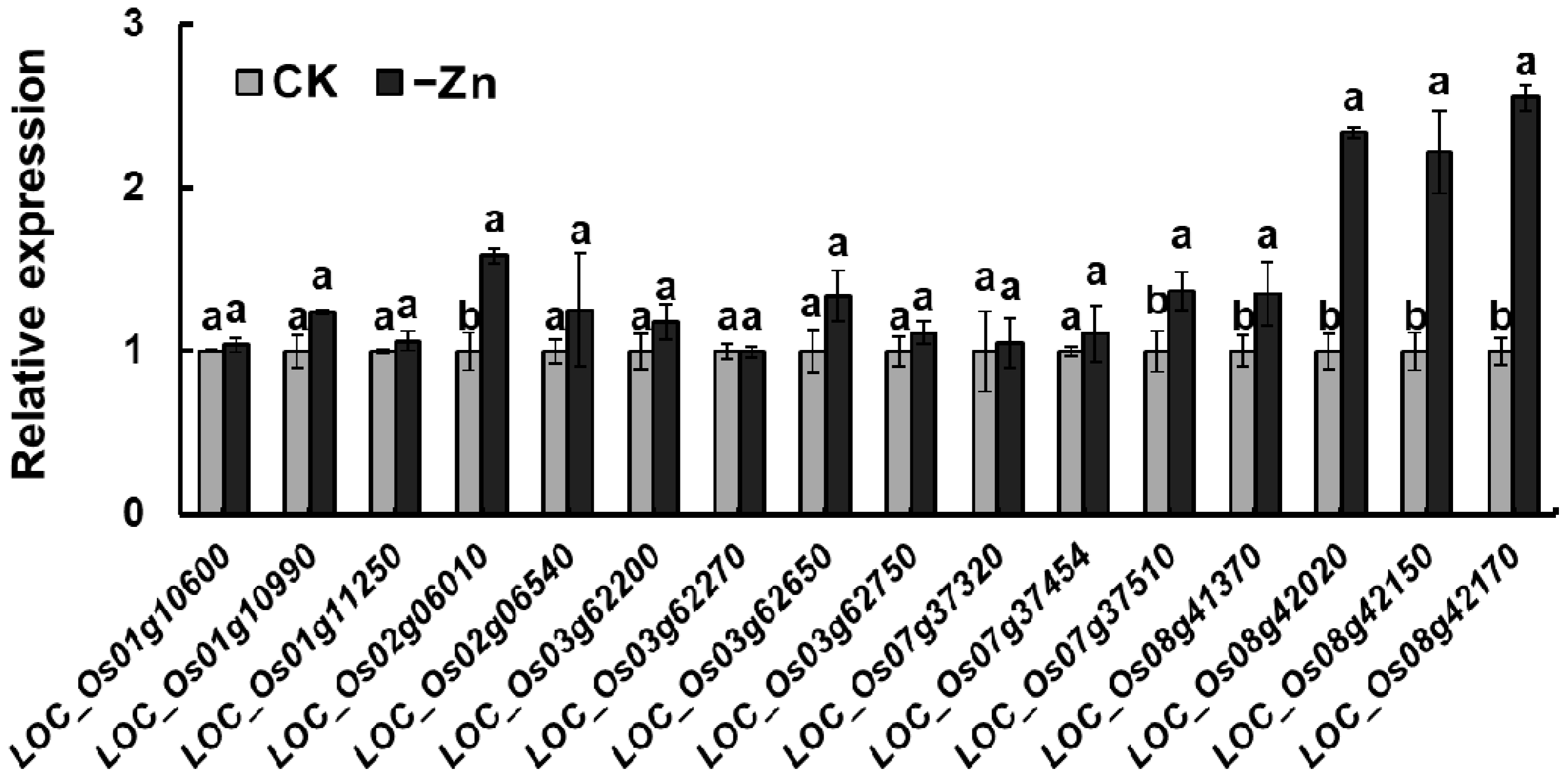
3.3. Identification of Candidate Genes Related to Zn2+ Accumulation in Shoots and Grains
3.4. Expression Pattern and Sequence Analysis of LOC_Os02g06010 in Parental Lines
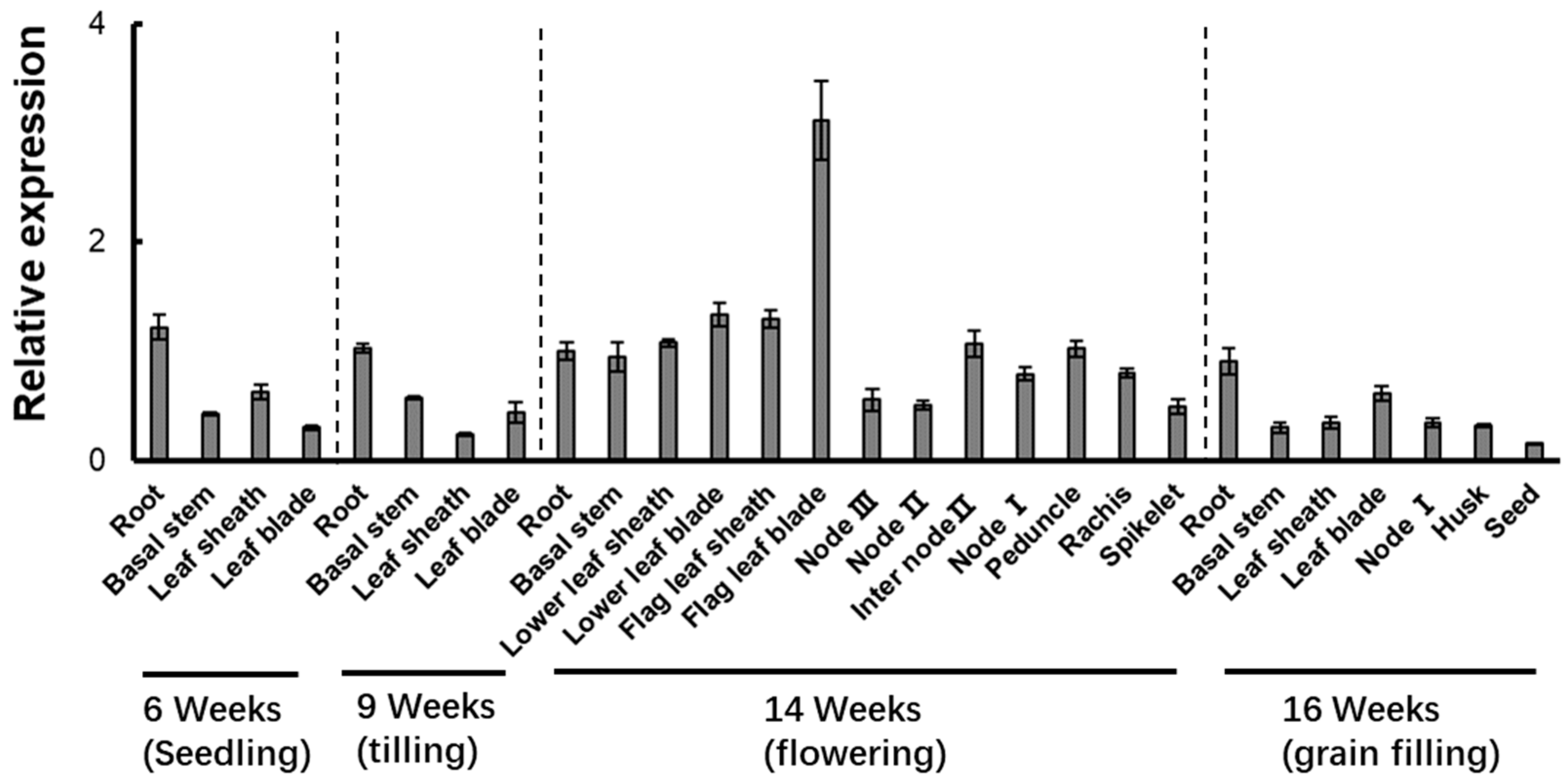
3.5. Expression of LOC_Os02g06010 in Different Organs at Different Growth Stages
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Broadley, M.; Brown, P.; Cakmak, I.; Rengel, Z.; Zhao, F. Function of nutrients. In Marschners Mineral Nutrition of Higher Plants; Academic Press: Cambridge, MA, USA, 2012; pp. 191–248. [Google Scholar]
- Wissuwa, M.; Ismail, A.M.; Yanagihara, S. Effects of zinc deficiency on rice growth and genetic factors contributing to tolerance. Plant Physiol. 2006, 142, 731–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bandyopadhyay, T.; Mehra, P.; Hairat, S.; Giri, J. Morpho-physiological and transcriptome profiling reveal novel zinc deficiency-responsive genes in rice. Funct. Integr. Genom. 2017, 17, 565–581. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, D.D.; Barros, P.M.; Cordeiro, A.M.; Serra, T.S.; Lourenco, T.; Chander, S.; Oliveira, M.M.; Saibo, N.J. Seven zinc-finger transcription factors are novel regulators of the stress responsive gene OsDREB1B. J. Exp. Bot. 2012, 63, 3643. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Jeong, H.J.; Kim, S.A.; Lee, J.; Guerinot, M.L.; An, G. OsZIP5 is a plasma membrane zinc transporter in rice. Plant Mol. Biol. 2010, 73, 507–517. [Google Scholar] [CrossRef] [PubMed]
- Barker, A.; Pilbeam, D. Handbook of Plant Nutrition; CRC Press: Boca Raton, FL, USA, 2015. [Google Scholar]
- Wang, L.C.; Busbey, S.; Bushey, S. Images in clinical medicine. Acquired acrodermatitis enteropathica. New Engl. J. Med. 2005, 352, 1121. [Google Scholar] [CrossRef]
- Zuo, J.; Li, J. Molecular genetic dissection of quantitative trait loci regulating rice grain size. Annu. Rev. Genet. 2014, 48, 99–118. [Google Scholar] [CrossRef]
- Shahzad, Z.; Rouached, H.; Rakha, A. Combating mineral malnutrition through iron and zinc biofortification of cereals. Compr. Rev. Food Sci. Food Saf. 2014, 13, 329–346. [Google Scholar] [CrossRef]
- Bouis, H.E.; Welch, R.M. Biofortification—A sustainable agricultural strategy for reducing micronutrient malnutrition in the global south. Crop Sci. 2010, 50, 20–32. [Google Scholar] [CrossRef] [Green Version]
- Patterson, K.Y.; Holbrook, J.T.; Bodner, J.E.; Kelsay, J.L.; Smith, J.C.; Veillon, C. Zinc, copper, and manganese intake and balance for adults consuming self-selected diets. Am. J. Clin. Nutr. 1984, 40, 1397–1403. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, R.; Ishimaru, Y.; Shimo, H.; Ogo, Y.; Nakanishi, H. The OsHMA2 transporter is involved in root-to-shoot translocation of Zn and Cd in rice. Plant Cell Environ. 2012, 35, 1948–1957. [Google Scholar] [CrossRef]
- Yamaji, N.; Xia, J.; Mitani-Ueno, N.; Yokosho, K.; Ma, J.F. Preferential delivery of zinc to developing tissues in rice is mediated by P-type heavy metal ATPase OsHMA2. Plant Physiol. 2013, 162, 927–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyadate, H.; Adachi, S.; Hiraizumi, A.; Tezuka, K.; Nakazawa, N.; Kawamoto, T.; Katou, K.; Kodama, I.; Sakurai, K.; Takahashi, H.; et al. OsHMA3, a P1B-type of ATPase affects root-to-shoot cadmium translocation in rice by mediating efflux into vacuoles. New Phytol. 2010, 189, 190. [Google Scholar] [CrossRef] [PubMed]
- Hongmei, C.; Sheng, H.; Jing, C.; Naoki, Y.; Ma, J.F. The tonoplast-localized transporter OsHMA3 plays an important role in maintaining Zn homeostasis in rice. J. Exp. Bot. 2019, 70, 2717–2725. [Google Scholar]
- Ramegowda, Y.; Venkategowda, R.; Jagadish, P.; Govind, G.; Hanumanthareddy, R.R.; Makarla, U.; Guligowda, S.A. Expression of a rice Zn transporter, OsZIP1, increases Zn concentration in tobacco and finger millet transgenic plants. Plant Biotechnol. Rep. 2013, 7, 309–319. [Google Scholar] [CrossRef]
- Liu, X.S.; Feng, S.J.; Zhang, B.Q.; Wang, M.Q.; Yang, Z.M. OsZIP1 functions as a metal efflux transporter limiting excess zinc, copper and cadmium accumulation in rice. BMC Plant Biol. 2019, 19, 283. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.R.; Feng, Y.; Chao, Y.E. Genomic analysis and expression pattern of OsZIP1, OsZIP3, and OsZIP4 in two rice (Oryza sativa L.) genotypes with different zinc efficiency. Russ. J. Plant Physiol. 2008, 55, 400–409. [Google Scholar] [CrossRef]
- Sasaki, A.; Yamaji, N.; Mitani-Ueno, N.; Kashino, M.; Ma, J.F. A node-localized transporter OsZIP3 is responsible for the preferential distribution of Zn to developing tissues in rice. Plant J. 2015, 84, 374–384. [Google Scholar] [CrossRef] [Green Version]
- Ishimaru, Y.; Suzuki, M.; Kobayashi, T.; Takahashi, M.; Nakanishi, H.; Mori, S.; Nishizawa, N.K. OsZIP4, a novel zinc-regulated zinc transporter in rice. J. Exp. Bot. 2005, 56, 3207. [Google Scholar] [CrossRef]
- Ishimaru, Y.; Masuda, H.; Suzuki, M.; Bashir, K.; Takahashi, M.; Nakanishi, H.; Mori, S.; Nishizawa, N.K. Overexpression of the OsZIP4 zinc transporter confers disarrangement of zinc distribution in rice plants. J. Exp. Bot. 2007, 58, 2909. [Google Scholar] [CrossRef] [Green Version]
- Youngsup, S.; Ryuichi, T.; Hiromi, N.; Takashi, Y. Sweet potato expressing the rice Zn transporter OsZIP4 exhibits high Zn content in the tuber. Plant Biotechnol. 2016, 33, 99–104. [Google Scholar]
- Tan, L.; Qu, M.; Zhu, Y.; Peng, C.; Wang, J.; Gao, D.; Chen, C. ZINC TRANSPORTER5 and ZINC TRANSPORTER9 function synergistically in Zinc/Cadmium uptake. Plant Physiol. 2020, 183, 1235–1249. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.; Zhu, Y.; Fan, T.; Peng, C.; Wang, J.; Sun, L.; Chen, C. OsZIP7 functions in xylem loading in roots and inter-vascular transfer in nodes to deliver Zn/Cd to grain in rice. Biochem. Biophys. Res. Commun. 2019, 512, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Ricachenevsky, F.K.; Punshon, T.; Lee, S.; Oliveira, B.H.N.; Guerinot, M.L. Elemental profiling of rice FOX lines leads to characterization of a new Zn plasma membrane transporter, OsZIP7. Front. Plant Sci. 2018, 9, 856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gindri, R.G.; Navarro, B.B.; da Cruz Dias, P.V.; Tarouco, C.P.; Ricachenevsky, F.K. Physiological responses of rice (Oryza sativa L.) oszip7 loss-of-function plants exposed to varying Zn concentrations. Physiol. Mol. Biol. Plants 2020, 26, 1349–1359. [Google Scholar] [CrossRef]
- Yang, M.; Li, Y.; Liu, Z.; Tian, J.; Lian, X. A high activity zinc transporter OsZIP9 mediates zinc uptake in rice. Plant J. 2020, 103, 1695–1709. [Google Scholar] [CrossRef]
- Huang, S.; Sasaki, A.; Yamaji, N.; Okada, H.; Mitaniueno, N.; Ma, J.F. The ZIP transporter family member OsZIP9 contributes to root Zn uptake in rice under Zn-limited conditions. Plant Physiol. 2020, 183, 1224–1234. [Google Scholar] [CrossRef]
- Yang, M.; Lu, K.; Zhao, F.-J.; Xie, W.; Ramakrishna, P.; Wang, G.; Du, Q.; Liang, L.; Sun, C.; Zhao, H.; et al. Genome-wide association studies reveal the genetic basis of ionomic variation in rice. Plant Cell 2018, 30, 2720–2740. [Google Scholar] [CrossRef] [Green Version]
- Gao, Z.; Wang, Y.; Chen, G.; Zhang, A.; Qian, Q. The indica nitrate reductase gene OsNR2 allele enhances rice yield potential and nitrogen use efficiency. Nat. Commun. 2019, 10. [Google Scholar] [CrossRef]
- Yan, H.; Xu, W.; Xie, J.; Gao, Y.; He, Z. Variation of a major facilitator superfamily gene contributes to differential cadmium accumulation between rice subspecies. Nat. Commun. 2019, 10, 2562. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, S.; Vgn, T.V.; Kotla, A.; Mangrauthia, S.K.; Neelamraju, S. Expression patterns of QTL based and other candidate genes in Madhukar × Swarna RILs with contrasting levels of iron and zinc in unpolished rice grains. Gene 2014, 546, 430–436. [Google Scholar] [CrossRef]
- Zhang, M.; Pinson, S.R.M.; Tarpley, L.; Huang, X.-Y.; Salt, D.E. Mapping and validation of quantitative trait loci associated with concentrations of 16 elements in unmilled rice grain. Theor. Appl. Genet. 2014, 127, 137–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swamy, B.P.M.; Kaladhar, K.; Anuradha, K.; Batchu, A.K.; Longvah, T.; Sarla, N. QTL Analysis for grain Iron and Zinc concentrations in two O. nivara derived backcross populations. Rice Sci. 2018, 25, 197–207. [Google Scholar] [CrossRef]
- Norton, G.J.; Deacon, C.M.; Xiong, L.; Huang, S.; Meharg, A.A.; Price, A.H. Genetic mapping of the rice ionome in leaves and grain: Identification of QTLs for 17 elements including arsenic, cadmium, iron and selenium. Plant Soil 2010, 329, 139–153. [Google Scholar] [CrossRef]
- Meng, L.; Guo, L.; Ponce, K.; Zhao, X.; Ye, G. Characterization of three rice multiparent advanced generation intercross (MAGIC) populations for quantitative trait loci identification. Plant Genome 2016, 9. [Google Scholar] [CrossRef] [Green Version]
- Meng, L.; Wang, B.; Zhao, X.; Ponce, K.; Qian, Q.; Ye, G. Association mapping of Ferrous, Zinc, and Aluminum tolerance at the seedling stage in indica rice using MAGIC populations. Front. Plant Sci. 2017, 8, 1822. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Ponce, K.; Meng, L.; Chakraborty, P.; Ye, G. QTL identification for salt tolerance related traits at the seedling stage in indica rice using a multi-parent advanced generation intercross (MAGIC) population. Plant Growth Regul. 2020, 92, 365–373. [Google Scholar] [CrossRef]
- Ponce, K.; Zhang, Y.; Guo, L.; Leng, Y.; Ye, G. Genome-wide association study of grain size traits in indica rice multiparent advanced generation intercross (MAGIC) population. Front. Plant Sci. 2020, 11, 395. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Zhan, J.; Li, J.; Lu, X.; Liu, J.; Wang, Y.; Zhao, Q.; Ye, G. Genome-wide association study (GWAS) for mesocotyl elongation in rice (Oryza sativa L.) under multiple culture conditions. Genes 2020, 11, 49. [Google Scholar] [CrossRef] [Green Version]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef]
- Chen, J.; Qi, T.; Hu, Z.; Fan, X.; Zhu, L.; Iqbal, M.F.; Yin, X.; Xu, G.; Fan, X. OsNAR2.1 positively regulates drought tolerance and grain yield under drought stress conditions in rice. Front. Plant Sci. 2019, 10, 197. [Google Scholar] [CrossRef] [Green Version]
- Sperotto, R.A.; Boff, T.; Duarte, G.L.; Santos, L.S.; Grusak, M.A.; Fett, J.P. Identification of putative target genes to manipulate Fe and Zn concentrations in rice grains. J. Plant Physiol. 2010, 167, 1500–1506. [Google Scholar] [CrossRef] [PubMed]
- Bandillo, N.; Raghavan, C.; Muyco, P.A.; Sevilla, M.A.L. Multi-parent advanced generation inter-cross (MAGIC) populations in rice: Progress and potential for genetics research and breeding. Rice 2013, 6, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catherine, C.; Cassin, G.; Couch, D.; Divol, F.; Higuchi, K.; Le Jean, M.; Misson, J.; Schikora, A.; Czernic, P.; Mari, S. Metal movement within the plant: Contribution of nicotianamine and yellow stripe 1-like transporters. Ann. Bot. 2009, 103, 1–11. [Google Scholar]
- Ramesh, S.A.; Shin, R.; Eide, D.J.; Schachtman, D.P. Differential metal selectivity and gene expression of two zinc transporters from rice. Plant Physiol. 2003, 133, 126–134. [Google Scholar] [CrossRef] [Green Version]
- Bashirullah, A.; Cooperstock, R.L.; Lipshitz, H.D. Spatial and temporal control of RNA stability. Proc. Natl. Acad. Sci. USA 2001, 98, 7025–7028. [Google Scholar] [CrossRef] [Green Version]
- Mazumder, B.; Seshadri, V.; Fox, P.L. Translational control by the 3′-UTR: The ends specify the means. Trends Biochem. Sci. 2003, 28, 91–98. [Google Scholar] [CrossRef]
- Venturin, M.; Moncini, S.; Villa, V.; Russo, S.; Bonati, M.T.; Larizza, L.; Riva, P. Mutations and novel polymorphisms in coding regions and UTRs of CDK5R1 and OMG genes in patients with non-syndromic mental retardation. Neurogenetics 2006, 7, 59–66. [Google Scholar] [CrossRef]
- Tsago, Y.; Chen, Z.; Cao, H.; Sunusi, M.; Khan, A.U.; Shi, C.; Jin, X. Rice gene, OsCKX2-2, regulates inflorescence and grain size by increasing endogenous cytokinin content. Plant Growth Regul. 2020, 92, 283–294. [Google Scholar] [CrossRef]
- Jacob, A.G.; Smith, C.W.J. Intron retention as a component of regulated gene expression programs. Hum. Genet. 2017, 136, 1043–1057. [Google Scholar] [CrossRef] [Green Version]
- Shaul, O. How introns enhance gene expression. Int. J. Biochem. Cell Biol. 2017, 91, 145–155. [Google Scholar] [CrossRef]
- Rose, A.B.; Carter, A.; Korf, I.; Kojima, N. Intron sequences that stimulate gene expression in Arabidopsis. Plant Mol. Biol. 2016, 92, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Gallegos, J.E.; Rose, A.B. An intron-derived motif strongly increases gene expression from transcribed sequences through a splicing independent mechanism in Arabidopsis thaliana. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akira, T.; Satoru, M.; Shozo, O.; Junko, K.; Ko, S.; Kenzo, N. Enhancement of foreign gene expression by a dicot intron in rice but not in tobacco is correlated with an increased level of mRNA and an efficient splicing of the intron. Nucleic Acids Res. 1990, 18, 6767–6770. [Google Scholar]
- Lu, J.; Sivamani, E.; Azhakanandam, K.; Samadder, P.; Li, X.; Qu, R. Gene expression enhancement mediated by the 5′ UTR intron of the rice rubi3 gene varied remarkably among tissues in transgenic rice plants. Mol. Genet. Genom. 2008, 279, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Samadder, P.; Sivamani, E.; Lu, J.; Li, X.; Qu, R. Transcriptional and post-transcriptional enhancement of gene expression by the 5′ UTR intron of rice rubi3 gene in transgenic rice cells. Mol. Genet. Genom. 2008, 279, 429–439. [Google Scholar] [CrossRef]
- Wu, C.S.; Chen, D.Y.; Chang, C.F.; Li, M.J.; Hung, K.Y.; Chen, L.J.; Chen, P.W. The promoter and the 5′-untranslated region of rice metallothionein OsMT2b gene are capable of directing high-level gene expression in germinated rice embryos. Plant Cell Rep. 2014, 33, 793–806. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| QTLs | Chr. | Position (bp) | p-Value | Contribution | Gene Symbol | References |
|---|---|---|---|---|---|---|
| Zn2+ concentration in shoot | ||||||
| qSZn1 | 1 | 5911595 | 0.001835 | 4.5% | ||
| qSZn2-1 | 2 | 3312324 | 0.000793 | 5.3% | ||
| qSZn2-2 | 2 | 23649858 | 0.004561 | 3.7% | OsYSL14 | [43] |
| qSZn3 | 3 | 35337103 | 0.003697 | 3.9% | ||
| qSZn8 | 8 | 7947702 | 0.000537 | 5.7% | ||
| Zn2+ concentration in brown rice | ||||||
| qGZn2 | 2 | 3178843 | 0.002451 | 5.7% | ||
| qGZn3 | 3 | 35466303 | 0.002126 | 5.8% | ||
| qGZn5-1 | 5 | 7576955 | 0.003099 | 5.5% | OsZIP7 | [24,25,26] |
| qGZn5-2 | 5 | 15660281 | 0.003082 | 5.5% | ||
| qGZn7 | 7 | 22050741 | 0.001602 | 6.1% | ||
| qGZn8 | 8 | 26287654 | 7.33E-05 | 8.9% | ||
| MSU ID | Annotation |
|---|---|
| qSZn1 | |
| LOC_Os01g10600 | aquaporin protein, putative, expressed |
| LOC_Os01g10990 | integral membrane protein DUF6 containing protein, expressed |
| LOC_Os01g11250 | potassium channel KAT1, putative, expressed |
| qSZn2-1/qGZn2 | |
| LOC_Os02g06010 | integral membrane protein, putative, expressed |
| LOC_Os02g06540 | transporter family protein, putative, expressed |
| qSZn3/qGZn3 | |
| LOC_Os03g62200 | ammonium transporter protein, putative, expressed |
| LOC_Os03g62270 | MATE efflux family protein, putative, expressed |
| LOC_Os03g62650 | ion channel DMI1-like, chloroplast precursor, putative, expressed |
| LOC_Os03g62750 | inner membrane protein, putative, expressed |
| qGZn7 | |
| LOC_Os07g37320 | transporter family protein, putative, expressed |
| LOC_Os07g37454 | urate anion exchanger, putative, expressed |
| LOC_Os07g37510 | organic cation transporter-related, putative, expressed |
| qGZn8 | |
| LOC_Os08g41370 | amino acid permease family protein, putative, expressed |
| LOC_Os08g42020 | zinc ion binding protein, putative, expressed |
| LOC_Os08g42150 | zinc transporter 2 precursor, putative, expressed |
| LOC_Os08g42170 | zinc transporter 2 precursor, putative, expressed |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; Zou, W.; Lu, X.; Bian, J.; He, H.; Chen, J.; Ye, G. Genome-Wide Association Study Using a Multiparent Advanced Generation Intercross (MAGIC) Population Identified QTLs and Candidate Genes to Predict Shoot and Grain Zinc Contents in Rice. Agriculture 2021, 11, 70. https://doi.org/10.3390/agriculture11010070
Liu S, Zou W, Lu X, Bian J, He H, Chen J, Ye G. Genome-Wide Association Study Using a Multiparent Advanced Generation Intercross (MAGIC) Population Identified QTLs and Candidate Genes to Predict Shoot and Grain Zinc Contents in Rice. Agriculture. 2021; 11(1):70. https://doi.org/10.3390/agriculture11010070
Chicago/Turabian StyleLiu, Shilei, Wenli Zou, Xiang Lu, Jianmin Bian, Haohua He, Jingguang Chen, and Guoyou Ye. 2021. "Genome-Wide Association Study Using a Multiparent Advanced Generation Intercross (MAGIC) Population Identified QTLs and Candidate Genes to Predict Shoot and Grain Zinc Contents in Rice" Agriculture 11, no. 1: 70. https://doi.org/10.3390/agriculture11010070