 Xin-jun Du
Xin-jun Du Xiao-yi Wang
Xiao-yi Wang Xuan Dong
Xuan Dong Ping Li
Ping Li Shuo Wang
Shuo Wang- 1State Key Laboratory of Food Nutrition and Safety, Key Laboratory of Food Nutrition and Safety, Ministry of Education, Tianjin University of Science and Technology, Tianjin, China
- 2Tianjin Key Laboratory of Food Science and Health, School of Medicine, Nankai University, Tianjin, China
Strong desiccation tolerance is an outstanding feature of Cronobacter sakazakii and can enable the bacterium to survive in a dry food matrix (such as milk powder) for a long time. Therefore, contamination of food possessing low water activity with C. sakazakii can increase the risk of infection in human beings, particularly in neonates and infants. However, the mechanism underlying the desiccation tolerance property of C. sakazakii is largely unknown. In this study, the desiccation tolerance characteristics of 42 C. sakazakii strains were analyzed. Simultaneously, the sequence types and biofilm formation abilities of the strains were investigated, and their correlations with desiccation tolerance were analyzed. The results showed no significant correlation between desiccation tolerance and sequence type. However, there was a positive correlation between biofilm formation ability and desiccation tolerance. Raman spectroscopy was employed to investigate the biofilm formed by strains with distinct desiccation tolerance levels, and the results showed that the levels of polysaccharide, proteins and carotenoid might play important roles in the resistance to dry environments. In addition, 10 genes involved in osmoprotectant synthesis or transport were selected, and their differential expression in strains with diverse desiccation tolerance levels was compared to investigate whether these genes were responsible for cytoprotection in the dry environment. The results revealed a great difference in gene expression among strains with different desiccation tolerance levels, suggesting that these genes play a regulatory role in the resistance of C. sakazakii to dry environments. Our study provides a useful reference for follow-up studies investigating the mechanism of desiccation tolerance in C. sakazakii.
Introduction
Cronobacter spp., classified in family Enterobacteriaceae, are rod-shaped, motile, non-spore-forming facultative anaerobic Gram-negative opportunistic pathogen (Farmer et al., 1980). The genus of Cronobacter has been classified into the following seven species: Cronobacter sakazakii, Cronobacter malonaticus, Cronobacter condimenti, Cronobacter universalis, Cronobacter dublinensis, Cronobacter turicensis, and Cronobacter muytjensii (Stephan et al., 2014). The organisms have been isolated from a wide variety of foods including cheese, meat, vegetables, grains, herbs and spices, as well as from mammals and invertebrates (Iversen and Stephen, 2003). Due to their survival in food and in the environment, these microorganisms can contact humans and cause many serious infections, such as meningitis, necrotizing enterocolitis, and septicemia. Particularly in low-birth-weight neonates, immuno-compromised neonates and infants less than 4 weeks of age, the mortality rate was about 40–80% (FAO/WHO, 2004, 2008). Additionally, cases of elderly and immunocompromised patients infected by Cronobacter spp. have also been reported (Iversen and Stephen, 2003). Recently, in addition to virulence factors that can be harmful to humans, the resistance of Cronobacter spp. to adverse environments, especially their strong desiccation tolerance and osmotic stress tolerance, has become another focus of research. These properties allow for the long-term persistence of Cronobacter in infant formulas and other continually unfavorable conditions (Breeuwer et al., 2003). In previous studies, members of the genus Cronobacter has exhibited greater desiccation tolerance. The resistance of bacterial pathogens such as Listeria monocytogenes, Escherichia coli O157: H7, Salmonella enterica, and C. sakazakii to dry conditions has been studied, and the results showed that C. sakazakii has a significantly stronger desiccation tolerance than the other organisms (Koseki et al., 2015). However, the mechanism underlying the desiccation tolerance of C. sakazakii is poorly understood.
In other bacteria, the mechanism underlying resistance to adverse environments is much better understood, providing a useful reference for investigating the outstanding desiccation tolerance property of C. sakazakii. A previous study in E. coli showed that when this bacterium is subjected to a hypertonic environment, a primary response of the accumulation of electrolytes, such as potassium, glutamate, etc., took place to increase the internal osmotic pressure of the cell to counteract the high external osmotic pressure, therefore preventing harm caused by the high osmotic environment (Laermann et al., 2013). The secondary response was based on the uptake or de novo synthesis of other protective compounds. In bacteria, uptaking and synthesizing compatible solutes is another important adaptive strategy to high-osmolality environments. These compounds are highly soluble molecules that are uncharged in a physiological pH environment. They are less harmful to cellular processes than electrolytes and are capable of protecting cells from adverse environments for an extended period (Feeney and Sleator, 2011). In addition, some compatible solutes, including saccharides, free amino acids and derivatives, quaternary amines and their sulfonium analogs, polyols, sulfate esters, and small peptides in E. coli and Bacillus subtilis were detected using Nuclear Magnetic Resonance (NMR) and High Performance Liquid Chromatography (HPLC) for osmoregulatory purposes (Kempf and Bremer, 1998). In further studies, an experiment performed in E. coli showed that intracellular trehalose could be induced by hyperosmosis and therefore promoted desiccation tolerance (Welsh and Herbert, 1999). In addition, metagenomics research on microbial responses to dry environments in diverse bacteria, such as trehalose biosynthesis, has clarified the pathways (Le et al., 2016). Furthermore, existing studies have shown that biofilms can play a protective role when bacteria are in adverse conditions. In various dry conditions, algae, fungi, archaea and bacteria mostly survive in the form of a biofilm community, in which they coexist and aggregate (Bar et al., 2002). C. sakazakii can form biofilms, in which cells are embedded in an excreted matrix composed of extracellular polymeric substances (EPSs) (Lebre et al., 2017), which is crucial for the protective effect of biofilms in desiccation tolerance (Bogino et al., 2013; Flemming et al., 2016; Yang et al., 2016; Fei et al., 2017). And the EPSs are primarily composed of polysaccharides, proteins, lipopolysaccharides and extracellular DNA (Bogino et al., 2013; Flemming et al., 2016). A study on Escherichia coli K-12 has found that EPSs involved in desiccation resistance (Danese et al., 2000). Another study on L. monocytogenes has found that preventing the presence of mature biofilms can reduce desiccation survival (Hingston et al., 2013). Although, there are many studies about desiccation tolerance in diverse bacteria, corresponding studies in C. sakazakii have not been comprehensively performed. In a previous study, compatible solutes in C. sakazakii were detected and analyzed by high-performance liquid chromatography (HPLC) (Breeuwer et al., 2003). However, further studies on the protective effect of trehalose and other compatible solutes in C. sakazakii under dry conditions have not been carried out. Previous studies indicated that colanic acid, one of the components of biofilms, and biofilm encapsulation may contribute to the persistence of C. sakazakii in adverse conditions (Iversen et al., 2004; Burgess et al., 2016). In addition, further investigation of the roles of biofilm in desiccation tolerance is still required to understand the mechanism underlying the prominent resistance of C. sakazakii to adverse conditions. Taken together, the current evidence is insufficient to explain the outstanding desiccation tolerance of C. sakazakii.
In order to investigate the mechanism underlying the desiccation tolerance of C. sakazakii, 42 strains were involved in this study and dry-condition-resistance capabilities of these strains were evaluated. Furthermore, sequence types, biofilm-forming abilities, biofilm components, and expression of 10 genes were studied, and their relationship with desiccation tolerance were analyzed and discussed.
Materials and Methods
Bacterial Strains and Genomic DNA Preparation
The 42 Cronobacter sakazakii strains used in this study. Three of them were obtained from the American Type Culture Collection (ATCC BAA-894, ATCC 29004, ATCC 12868) and other 39 were isolated from different food matrices. The isolates were mainly obtained from dairy products, whey protein products and casein products. All of the strains are listed in Table 1.
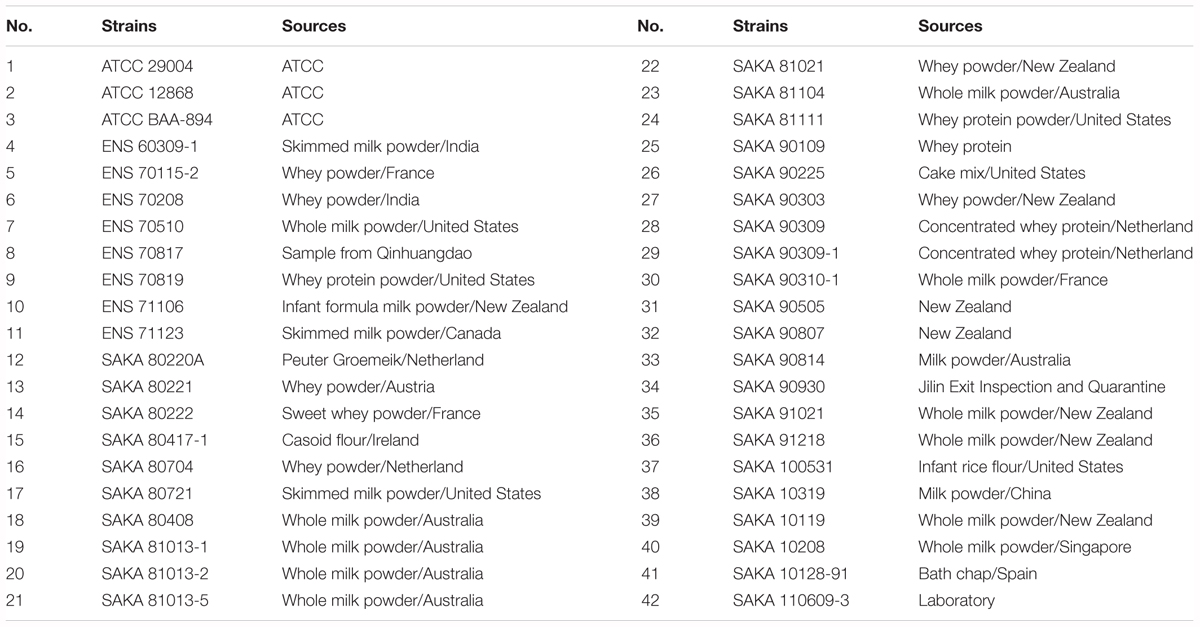
TABLE 1. The C. sakazakii strains used in this study.
Cronobacter sakazakii strains were stored at -80°C in LB medium with 20% glycerol. A single colony from each agar plate was inoculated in 10 mL of liquid LB medium and cultured at 37°C for about 12 h until the bacterial cells in each tube reached 106–108 CFU/mL. Genomic DNA of all strains was extracted by a Bacterial Genome DNA Kit (Tiangen, China) according to the manufacturer’s instructions.
Desiccation Tolerance Analysis
Desiccation tolerance analysis of the strains were performed as in previous reports with some modifications (Breeuwer et al., 2003; Barron and Forsythe, 2007; Hingston et al., 2013). After culture of the strains on plates, a single colony was cultured in 10 mL liquid LB broth until the bacterial cells reached logarithmic phase. The OD600 value of different bacterial culture was measured, and a 100 μL portion of the culture was transferred into each well of a 96-well microtiter plate. Subsequently, the plate was transferred into a sterile dryer with dehydrated silica gel. The dryer was placed in a sterile incubator (DHP-2042BS, Huabei, Tianjin, China), which was controlled at a constant temperature of 37°C. After 6 days of drying, the 96-well microtiter plate was removed, 100 μL/well of fresh medium was added, and the plate was cultured with 200 rpm shaking at 37°C for 3 h. The liquid in each well was transferred to a new 96-well microtiter plate, and the OD600 was detected. Each strain was tested in triplicate, and each test was repeated three times. The inactivation rate was calculated using the following formula to measure the desiccation tolerance of the strains:
OD0 represents the initial OD600 of each strain, and OD1 represents the OD600 of each strain cultured for 3 h after 6-day drying treatment.
Multilocus Sequence Typing (MLST) of C. sakazakii
Seven pairs of primers representing the seven housekeeping genes, including atpD, fusA, glnS, gltB, gyrB, infB, and pps, were synthesized according to the protocol on the MLST website1. The PCR system and conditions followed the protocol provided on the website. PCR products were sequenced by Suzhou Jinweizhi Biotechnology Co., Ltd. (Suzhou, China). To analyze the phylogenetic relationship of all 42 strains, the DNA sequence of the seven housekeeping genes were concatenated together (3,036 bp) in a certain order and used to draw the phylogenetic tree by MEGA 6.0. The relationship of the strains were analyzed by the neighbor-joining statistical method with a Tamura 3-parameter model (Baldwin et al., 2009; Joseph and Forsythe, 2012).
Biofilm Formation Experiment
The experiment of biofilm formation is referred to in previous reports (Auger et al., 2006; Lu et al., 2012).
The overnight culture was transferred to 1 mL fresh LB medium (1:100) and cultured at 37°C until the OD600 value reached 0.6–0.8. 100 μL/well bacterial cells at logarithmic phase were transferred to a 96-well microtiter plate. After 3 days of static culture at 37°C, planktonic cells were removed from the liquid medium. The wells were washed three times with 150 μL of double-distilled water, and the majority of the biofilms were stained with 150 μL of 0.1% crystal violet (CV) for 30 min. Then, the unbound dye was removed, and the plates were washed with double-distilled water. Finally, the CV binding to the biofilm was dissolved in 150 μL of 95% ethanol for 30 min, and the ability to form biofilms was determined by measuring the absorbance at OD590 with a microplate reader. Each strain was detected three times, and fresh medium was used as a control.
Raman Spectroscopy Analysis
Raman spectroscopy was used to examine the components of biofilm formed by strains with differing desiccation tolerance. Four strains, ENS 71106 and SAKA 110609-3 with a stronger desiccation tolerance and ENS 70819 and SAKA 90814 with a weaker desiccation tolerance, were activated and cultured until reaching exponential phase. A 50 μL portion of each strain culture was transferred to a nitrocellulose membrane (Millipore, Ireland). Next, these nitrocellulose membranes were placed on the LB agar plates and cultured at 37°C for 72 h. During this period, the LB agar plates had to be changed every 24 h (Du et al., 2012).
In this study, Raman spectroscopic analyses were performed using a Renishaw inVia Raman system (Renishaw, Gloucestershire, United Kingdom) equipped with a 785 nm near-infrared diode laser and a Leica microscope (Leica Biosystems, Wetzlar, Germany). In addition, a WITec alpha300 Raman microscope (WITec, Ulm, Germany) equipped with a UHTS-300 spectrometer was used to collect Raman spectra. Instrumental control and data collection were performed by WiRE 2.0 software (WITec, Ulm, Germany). Raman spectra were collected in the form of a simultaneous Raman shift range of 1600-600 cm-1 in an extended mode. For measurement in a single area, each full spectral measurement was for a 10 s integration time with 6 spectral accumulations (total integration time, 60 s). The nitrocellulose membranes on which the biofilms were formatted were placed on glass slides and then directly placed under a microscope for focus adjustment and employed for Raman spectral collection. Ten spectra were collected for each biofilm in triplicate. By creating two-dimensional images, principal component analysis (PCA) of the biofilms formed by the four C. sakazakii strains was performed.
Quantitative Real-Time PCR
The two strains with a stronger desiccation tolerance (SAKA 110609-3 and ENS 71106) and the two strains with a weaker desiccation tolerance (ENS 70819 and SAKA 90814) were separately chosen for quantitative real-time PCR experiments. Ten osmotic-related genes were selected, and the expression levels of the genes were evaluated in the four strains pre- and post-dry treatment. After the Cronobacter sakazakii strains were cultured to exponential phase, the bacterial cells were collected by centrifugation at 6,000 rpm for 5 min at 4°C. In preparation of dry-treated samples, the bacteria were kept in a sterile incubator at a constant temperature (42°C) and a relative humidity of 43% for 1 h (Breeuwer et al., 2003). RNA was extracted using a Bacterial RNA Kit (Omega Bio-Tek, Norcross, GA, United States) and reverse transcribed into cDNA using an ImProm-IITM Reverse Transcription System (Takara, Dalian, China). The cDNA was used as the template, and the 16S rDNA gene was used as an internal reference. Three biological replicates were carried out, and the fold changes in gene expression were calculated using the 2-ΔΔCt method (Livak and Schmittgen, 2001). The specific primers for all ten target genes and the reference gene used are listed in Table 2.

TABLE 2. The primers specific for 10 target genes and 16S rRNA used in quantitative real-time PCR analysis.
Results
Desiccation Tolerance Trial
The desiccation tolerance of the strains was performed according to the formula described in the Section “Materials and Methods,” and the results are summarized in Figure 1. After 6 days of treatment in a dry environment, the inactivation rate of the seven strains was less than 55%, indicating stronger desiccation tolerant abilities. Among these strains, the SAKA 110609-3 strain showed the best capability for desiccation tolerance, and the inactivation rate was 49.98%. In contrast, the ENS 70819 strain was the weakest in terms of desiccation resistance, with an inactivation rate of 92.87%. A total of five strains showed more than an 80% inactivation rate, suggesting weak desiccation tolerance. The remaining strains (16 strains) showed moderate desiccation tolerance, with inactivation rates between 60 and 70%.
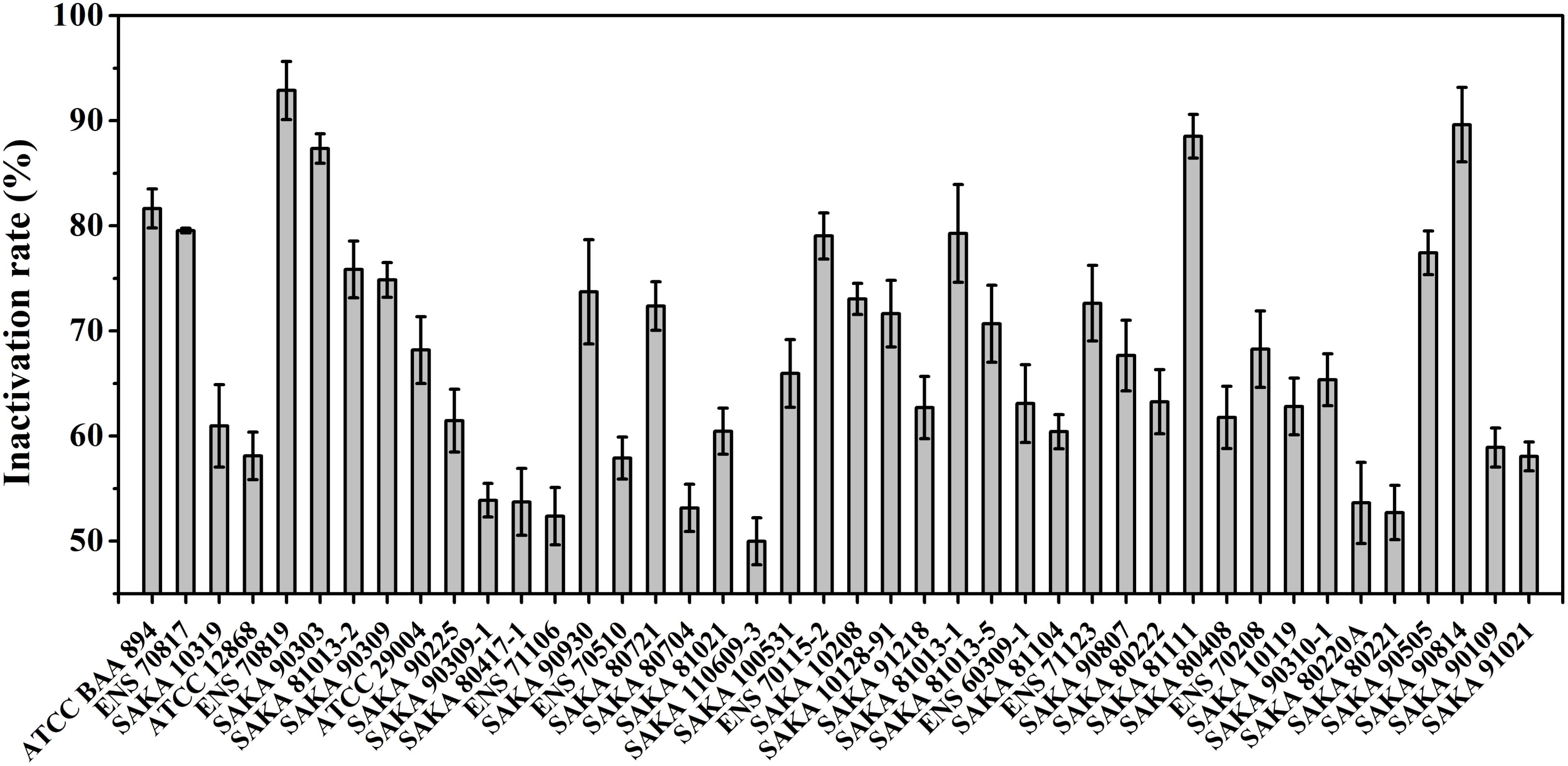
FIGURE 1. Desiccation tolerance analysis of 42 strains.
Multilocus Sequence Typing of C. sakazakii
Multilocus sequence typing assays were performed to analyze C. sakazakii, and the results indicated that 20 different sequence types (STs) were identified in 42 tested strains. In addition, the STs of the strains are listed in Supplementary Table 1. Among the 20 STs, ST4 was the dominant ST (9/42), and four strains each belonged to ST42 and ST58. In addition, three strains were in the ST1 group. Two strains each were classified into ST8, ST14, ST199 and ST327. The remaining strains differed in their STs. Moreover, three strains did not match any STs in the pubMLST database of Cronobacter spp. Concatenated nucleotide sequences (3,036 bp) of the seven loci were used to assess all strains together in one phylogenetic tree (Figure 2). The relationship of the 42 strains were analyzed using the neighbor-joining statistical method with a Tamura 3-parameter model.
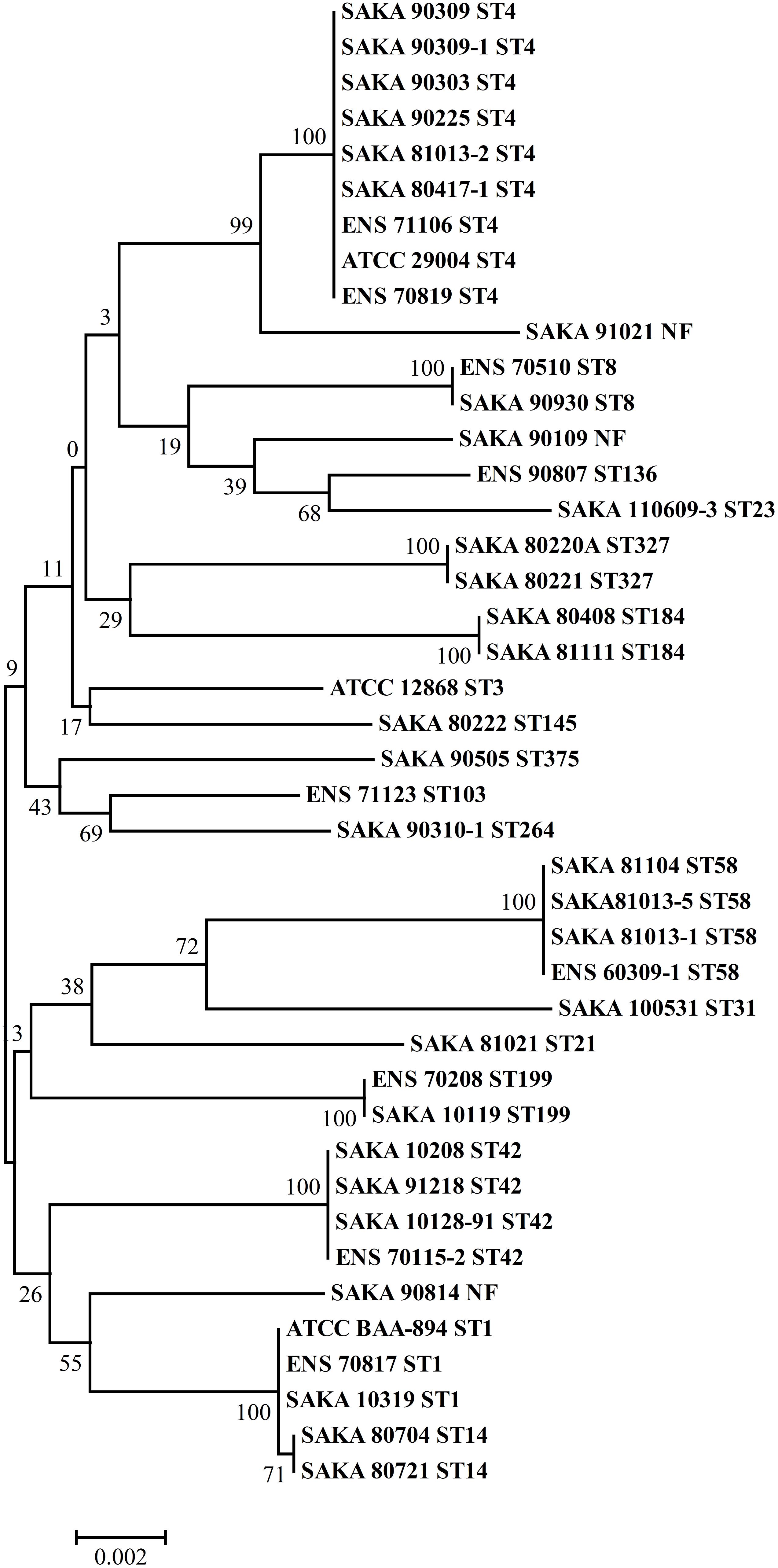
FIGURE 2. Phylogenetic tree of concatenated nucleotide sequences of the seven loci, using the neighbor-joining method with the Tamura 3-parameter model. Bootstrap values are shown for 1,000 replicates.
The correlation between sequence type and desiccation tolerance was analyzed. The strain with the highest desiccation tolerance (SAKA 110609-3) belonged to ST23. In addition, the strain with the weakest desiccation tolerance (ENS 70819) belonged to ST4. Analyzing the strains with an inactivation rate over 80% showed that two strains belonged to ST4 (2/9), and one strain belonged to ST1 (1/9). One of the other two strains belonged to ST184, and the remaining strain was not found in the database. Additionally, 2 ST4 strains (2/9) and 2 ST327 strains (2/2) were found to have an inactivation rate of less than 55%. The other strains were ST14, ST23, and ST8. In particular, 2 of the 9 strains of ST4, the dominant ST in our study, had weak desiccation tolerance (over 80% inactivation). In addition, another 2 of the 9 strains had high desiccation tolerance (the inactivation rate was less than 55%). The ENS 70819 strain was the ST4 strain with the highest inactivation rate, while ENS 70116, with an inactivation rate of 53.6%, was another ST4 strain with more resistance to dry environments than most strains. The great difference in the desiccation tolerance among the ST4 strains showed no significant correlation with ST type in C. sakazakii.
Biofilm Formation Trial
The biofilm-forming ability of the 42 strains was evaluated by CV staining. As shown in Figure 3, the biofilm formation ability of SAKA 110609-3 was the strongest, and the OD590 value reached 0.70. In contrast, the biofilm formation ability of the SAKA 81111 strain was the weakest, with an OD590 value of 0.12. Among these strains, six had higher OD590 values (above 0.60), suggesting that they have a higher biofilm formation ability. Four strains showed a weaker biofilm-formation ability, and the OD590 values were lower than 0.20. The rest of the strains exhibited a moderate biofilm formation ability. The results (Figure 4) show the correlation between biofilm formation ability and desiccation tolerance. Particularly, most of the strains with great desiccation tolerance, such as SAKA 110609-3, ENS 71106, ENS 80221, SAKA 80220A, and SAKA 90309-1 (the lines in red in Figure 4), also exhibited strong biofilm-formation abilities. In contrast, in strains with a higher inactivation rate (more than 80%), such as strains ENS 70819, SAKA 81111, and SAKA 90814 (the lines in blue in Figure 4), the biofilm formation ability was correspondingly weaker than that of most of the tested strains. Accordingly, the stains with moderate desiccation tolerance, such as SAKA 10319, SAKA 80408, SAKA 91218, SAKA 10119, SAKA 90807, and SAKA90210-1, generally showed moderate biofilm formation abilities. Taken these results together, it could be concluded that biofilm formation was positively related with desiccation tolerance.
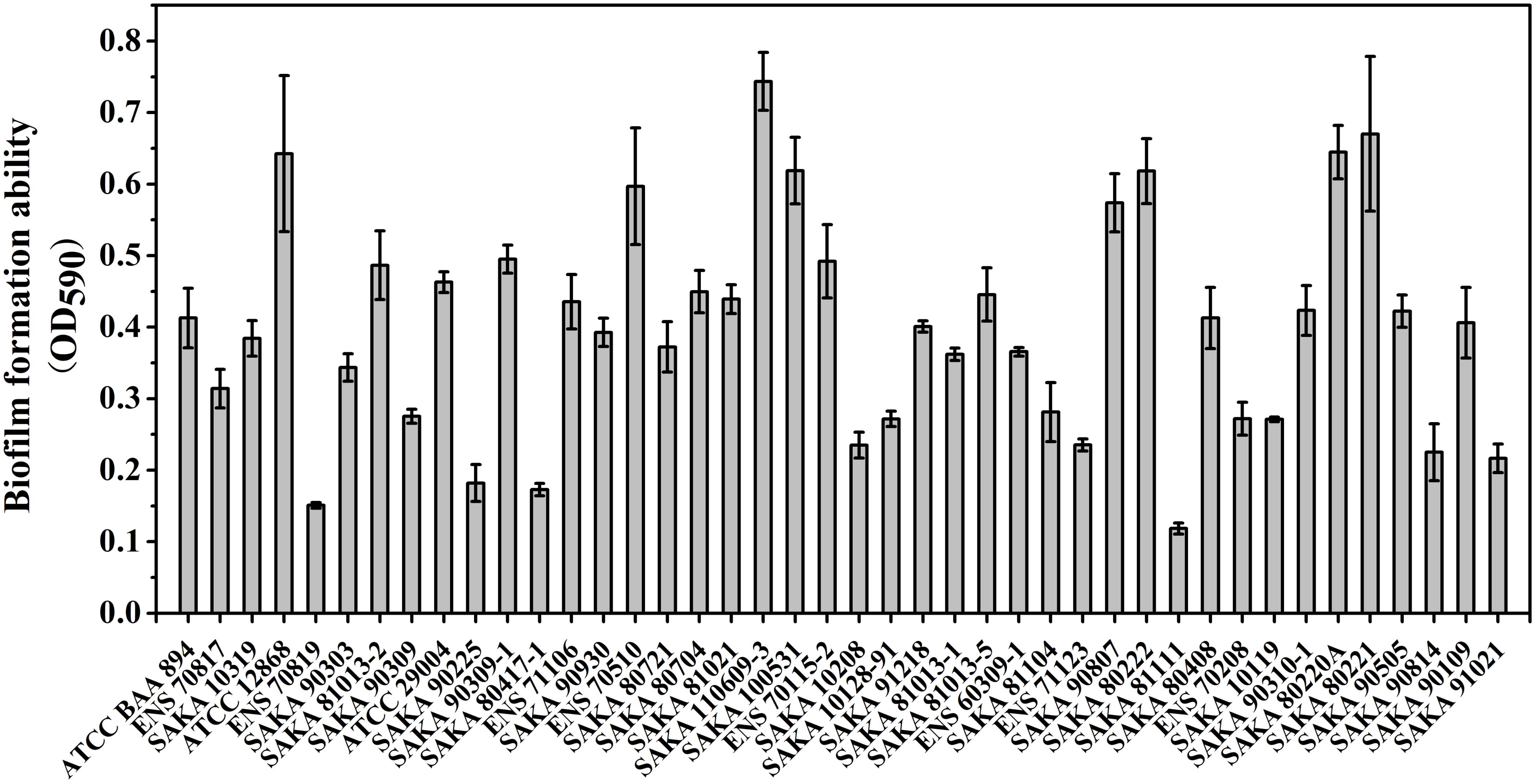
FIGURE 3. Biofilm formation ability of 42 strains.
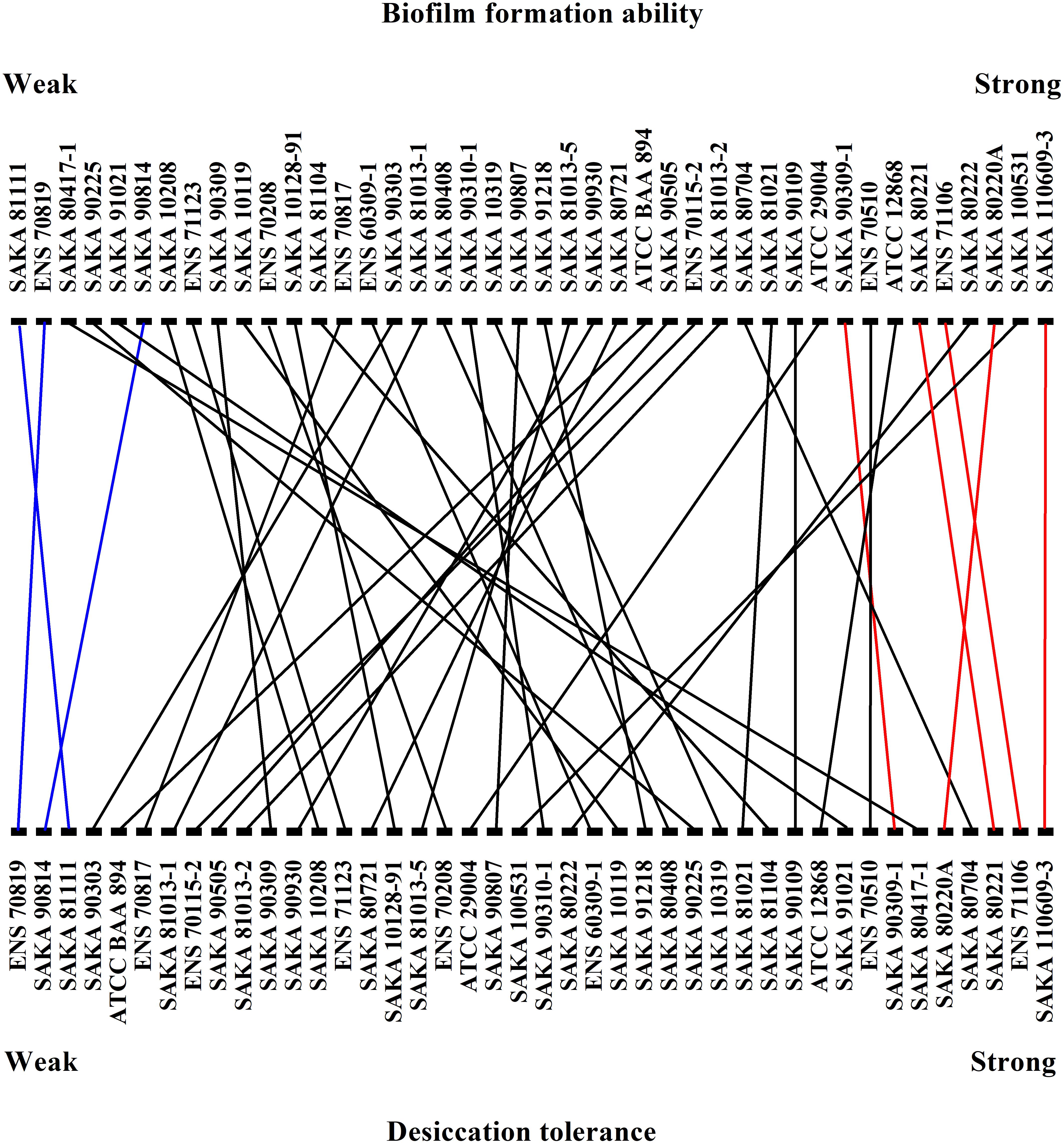
FIGURE 4. Correlation between biofilm formation ability and desiccation tolerance.
Comparison of Biofilm Components in Four Strains With Differing Biofilm-Formation Ability and Desiccation Tolerance
The biofilm components of four strains with different biofilm-forming abilities and different desiccation tolerant abilities (ENS 71106, SAKA 110609-3, ENS 70819 and SAKA 90814) were studied using Raman spectroscopy. Raman peaks at different Raman shifts are shown in Figure 5A. The peaks at 1002, 1157, and 1522 cm-1 in two strains with a strong desiccation tolerance (ENS 71106 and SAKA 110609-3) were significantly higher than those in the other two strains with a weak desiccation tolerance (ENS 70819 and SAKA 90814). In addition, in the two strains with a strong desiccation tolerance, the peak at 1450 cm-1 was slightly higher than in the weak two strains. The band at 1002 cm-1 was characteristic of phenylalanine. In addition, the band at 1157 cm-1 was representative of the C-C and C-N bond stretching of proteins, while the band at 1522 cm-1 was associated with C-C and conjugated C = C bond stretching of carotenoids. The band at 1450 cm-1 was related to fatty acids (De Gelder et al., 2007; Lu et al., 2011). In addition, the peak at 1367 cm-1 matched the characteristics of cellulose (Lu et al., 2011) was lower in the weakest desiccation tolerance strain ENS 70819 than in the other three strains. The peak at 1367 cm-1 representing cellulose was the highest in ENS 110609-3 than in the four strains.
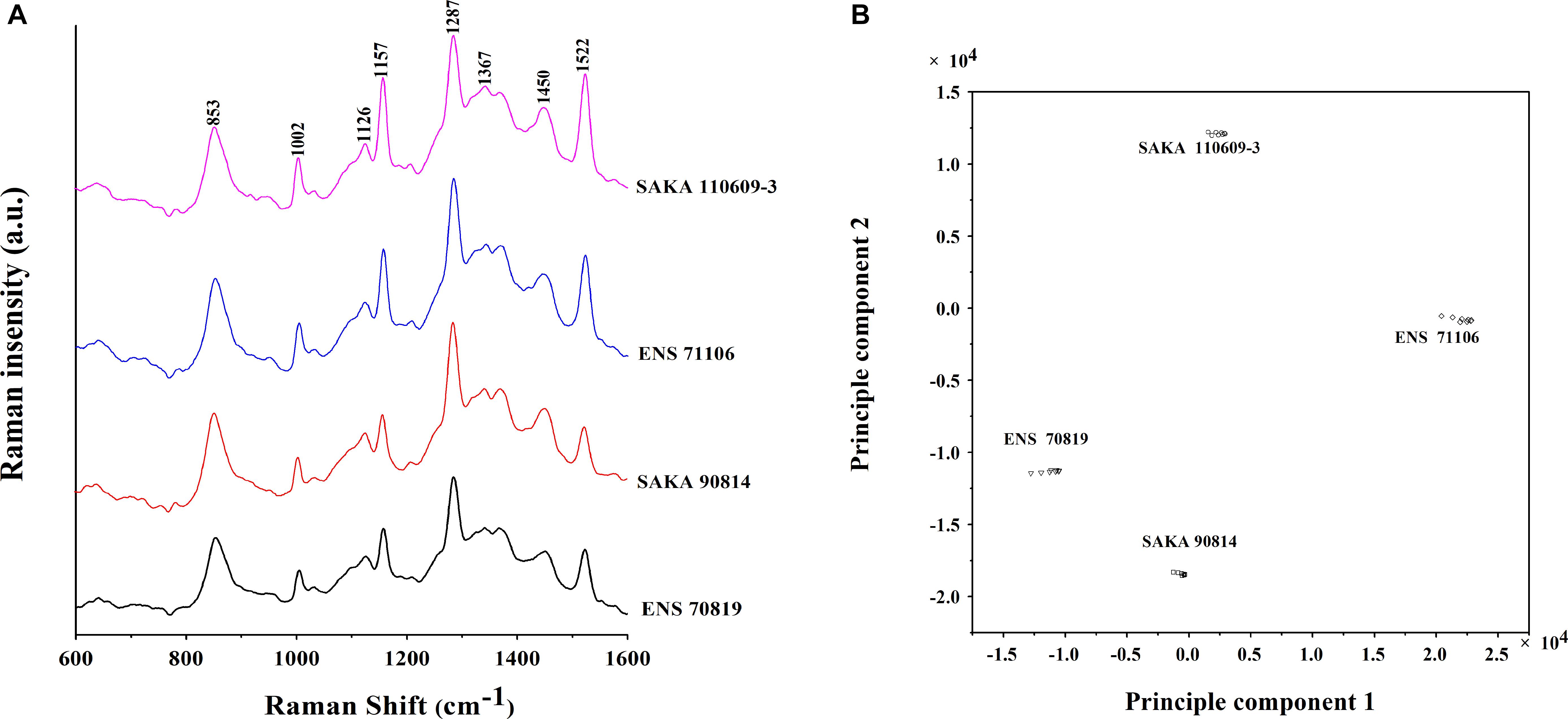
FIGURE 5. Raman spectrum analysis of biofilm formed by the four different strains. (A) Raman spectrum of the biofilms of the four strains (Raman Shift 1600-600 cm-1). (B) Principal component analysis (PCA) model that was based on the Raman spectrum of biofilms of the four strains.
A two-dimensional PCA model was established to distinguish the four strains, and the results showed that there were differences in the main components among ENS 70819, ENS 71106, SAKA 90814 and SAKA 110609-3 (Figure 5B).
Quantitative Real-Time PCR
According to the desiccation tolerance analysis of the 42 strains, two strains with the strongest desiccation tolerance (ENS 71106 and SAKA 110609-3) and the other two strains with the weakest desiccation tolerance (ENS 70819 and SAKA 90814) were selected for quantitative real-time PCR analysis. Ten genes associated with ions or compatible solute transport or synthesis were selected to test their expression changes between pre- and post-dry treatments. The results showed that all 10 genes in two strains with a strong desiccation tolerance were expressed at higher levels in comparison with the two strains with a weak desiccation tolerance (Supplementary Table 2). After drying treatment, proline/glycine betaine transporter (ESA_02131), glycine/betaine ABC transporter ATP-binding protein OpuCA (ESA_01738), glycine/betaine ABC transporter substrate-binding protein OpuCC (ESA_01740), trehalose-6-phosphate synthase OtsA (ESA_01335), and choline dehydrogenase BetA (ESA_02049) showed a significantly upregulated expression (p < 0.05). Compared with strain ENS 70819 with the weakest desiccation tolerance, all genes were upregulated in the two stains with strong desiccation tolerance (SAKA 110609-3 and ENS 71106), and the regulation in strain SAKA 110609-3 was higher than that in strain ENS 71106 (Figure 6A). Expression of the 10 genes was upregulated by 14.94-fold and 4.63-fold on average in SAKA 110609-3 and ENS 71106, respectively, in comparison with ENS 70819. Compared with the other strain with a weak desiccation tolerance (SAKA 90814), the expression levels of the 10 genes in SAKA 110609-3 were also significantly higher (average increase reaching 6.43-fold). However, in the strain with the second strongest desiccation tolerance (ENS 71106), only three genes (ESA_03328, ESA_04389, and ESA_02049) showed significantly higher expression levels in comparison with that of SAKA 90814. The remaining genes (ESA_02131, ESA_01738, ESA_01740, ESA_02641, ESA_01334, ESA_01335, and ESA_02048) showed similar expression levels in strains ENS 71106 and SAKA 90814. Based on these results, it can be concluded that the genes including ESA_03328 (encoding MFS transporter), ESA_04389 (encoding glutathione-regulated potassium-efflux system protein KefB), and ESA_02049 (encoding choline dehydrogenase BetA) are more likely to play key roles in desiccation tolerance than the other genes tested in this study (Figures 6A,B).
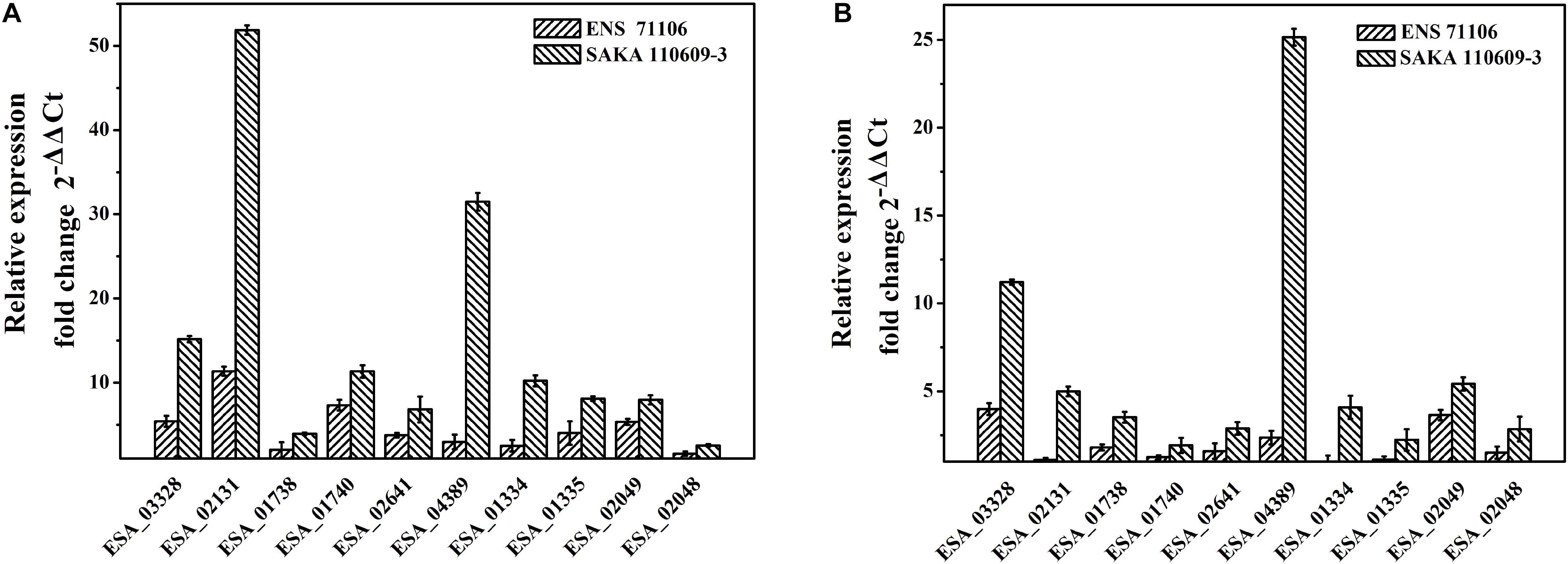
FIGURE 6. Relative expressions of 10 selected genes in strains with strong desiccation tolerance in comparison with strains with weak desiccation tolerance. (A) The expression levels of 10 genes in ENS 71106 and SAKA 110609-3 in comparison with those in ENS 70819. (B) The expression levels of 10 genes in ENS 71106 and SAKA 110609-3 in comparison with those in SAKA 90814.
Discussion
The Relevance Between the Sequence Type (ST) and Desiccation Tolerance
Multilocus sequence typing is a method that can be used to classify various genera of Cronobacter spp. by analyzing the nucleotide sequences of seven housekeeping genes. The STs of 42 strains were obtained by an MLST assay. In this study, ST4 was the predominant type (9/42) in comparison with other ST types. And it is known that the most serious clinical cases of meningitis are relevant to a single ST: C. sakazakii ST4 (Hariri et al., 2013). In this study, ST4 was the predominant type (9/42) in comparison with other ST types. However, C. sakazakii ST4 showed no significantly stronger desiccation tolerance than other STs based on the results obtained in this study. Moreover, even the abilities of the nine ST4 strains to resist dry environments were obviously different (Figure 1). MLST is based on the sequence of seven genes (atpD, fusA, glnS, gltB, gyrB, infB, and pps) responsible for the basic physiological functions to classify different of Cronobacter isolates. However, the function of these genes was not directly related to specific characteristics such as desiccation tolerance. Therefore, it is reasonable that there was no significant correlation between the ST and desiccation tolerance of different strains.
The Relationship Between Biofilm Formation Ability and Desiccation Tolerance
Bacterial biofilms are formed from communities embedded in a self-produced matrix of EPSs (Flemming et al., 2016). In previous studies, biofilm formation was considered an important means of protecting bacteria from long-term (even several years) survival in a dry environment (Barron and Forsythe, 2007). The correlation between desiccation tolerance and biofilm formation ability has been studied and discussed in Salmonella spp. (Finn et al., 2013), L. monocytogenes (Hansen and Vogel, 2011; Hingston et al., 2013) and E. coli (Chen et al., 2004; Barron and Forsythe, 2007), and the results indicate that biofilms can help bacteria avoid harm from adverse conditions. In this study, all 42 tested strains were subjected to CV staining analysis at OD590 to characterize the ability of biofilm formation (Figure 3). According to the results, we found that SAKA 110609-3 had the strongest desiccation tolerance, and its ability to form biofilms was much greater than that of the other strains. Furthermore, most strains with great desiccation tolerance also have a better biofilm formation ability. This result is consistent with similar studies in L. monocytogenes showing that biofilm formation effectively protects bacteria from adverse environmental conditions, including long-term survival in a dry environment (Hingston et al., 2013). Some researchers deemed that colanic acid and encapsulation may contribute to adherence to some surfaces but may also contribute to resistance to dry stress (Flemming et al., 2016). Studies by Scheepe-Leberlolhne and Wagner suggested that colanic acid may be an important contributor to biofilm formation and increased resistance to environmental stresses such as desiccation, heat, and pH in Cronobacter spp. (Scheepe-Leberlolhne and Wagner, 1986).
To further study the biofilm-forming ability among strains with different desiccation tolerance, biofilm component analysis using Raman spectroscopy was performed. Based on the Raman spectroscopy results, the main difference in the components of the four strains was appeared at peaks of 1002, 1157, and 1522 cm-1 for the Raman shift. The band at 1002 cm-1 represents phenylalanine (Manoj and Kumar, 2007), the band at 1157 cm-1 represents C-C and C-N bond stretching of proteins, and the band at 1522 cm-1 represents C-C and conjugated C = C bond stretching of carotenoids (Feng et al., 2014). In existing studies, 80% of the total biofilm composition are polysaccharide and proteins. And the macromolecules of biofilm components, such as proteins, can affect the overall properties of the biofilm (Bogino et al., 2013). In addition, carotenoids distributed on the surface of some C. sakazakii EPSs has been reported (Du et al., 2012). In this study, the significant difference in the biofilm components in the four tested strains were all associated with the proteins and carotenoids, showing an involvement with desiccation tolerance. In addition to polysaccharides, proteins, and carotenoids, fatty acids and cellulose are components of EPSs. In strain ENS 70819 with the weakest desiccation tolerance, the peak heights at 1450 cm-1 and 1367 cm-1 associated with fatty acid and cellulose were lower than those in the other three strains, indicating that fatty acid and cellulose were involved in producing the different biofilm-forming abilities of ENS 70819 and were therefore involved in desiccation tolerance.
Analysis of the Relevance of the 10 Genes With Desiccation Tolerance
It is considered that when C. sakazakii is exposed to a low-water-activity environment, the accumulation of electrolytes increases the internal pressure and counteracts the high external osmotic pressure (Feeney and Sleator, 2011). The Kdp system and Kef system associated with electrolyte accumulation and efflux are considered the primary response regulation. In this study, kdpA and kefB were selected to demonstrate their roles in desiccation tolerance. The Kdp system is one of the transporters of K+, an inducible P-type ATPase with a high affinity and specificity for K+. kdpA is the potassium transporting ATPase subunit A of the kdpABC operon, which consists of three proteins (Heermann et al., 2014). According to the results of quantitative real-time PCR, the gene expression of kdpA (ESA_02641) at the mRNA level was upregulated by 1.55-fold and 2.80-fold in the two strains with strong desiccation tolerance. This result indicated that under dry conditions, K+ accumulation in strains with strong desiccation tolerance is a primary regulating response and is more effective than that in strains with less desiccation tolerance. However, the excess K+ should be effluxed to avoid the toxic effect on cells. It is reported that K+ efflux is required when cells undergo hypo-osmotic shock (Van der Laan et al., 2002). Glutathione-gated potassium-efflux systems (Kef) can control K+ efflux and prevent prolonged exposure of bacterial cells to excess K+ ions. In this study, the glutathione-regulated potassium-efflux system protein KefB was selected to test. As a result of quantitative real-time PCR, the expression of kefB (ESA_04389) was upregulated 1.19-fold and 12.67-fold in two strains with strong desiccation tolerance. In addition, this result suggested that strains with a strong desiccation tolerance have a great regulatory effect on protecting strains in high K+ concentrations.
Potassium and glutamate serve as temporary osmoprotectants, but a high ion concentration in cells for an extensive period can bring great harm to cells. C. sakazakii can accumulate compatible solutes by uptake from the environment or by self-synthesis to adapt to hypertonic conditions (Feeney and Sleator, 2011). Compatible solutes, such as trehalose and glycine betaine, participate in the adaptation strategy in most xerotolerant microorganisms (Santos and Da Costa, 2002). In this study, the expression of two genes in the ProP system (ESA_02131 and ESA_03328) and two other genes (ESA_01738 and ESA_01740) encoding OpuCA and OpuCC were studied. The ProP system, OpuCA and OpuCC are the major osmoprotectant absorption system in C. sakazakii (Sleator et al., 2001; Feeney et al., 2014). The ProP system involves the transport of proline, glycine betaine, and ectoine, and proteins OpuCA and OpuCC are involved in the transport of choline and carnitine. It was shown that the expression of proP (ESA_02131) in C. sakazakii BAA-894 was upregulated under osmotically stressful conditions (Feeney et al., 2014). In the present study, the four genes in the strains with strong desiccation tolerance were upregulated after 1 h of exposure to drying conditions. However, the situations were different in weaker desiccation tolerance strains. In addition to opuCA (ESA_01738), the tested genes in strain ENS 70819 (weak desiccation tolerance) showed a downregulated expression in comparison with the other 3 strains. Taken together, strains with significantly differing desiccation tolerance usually exhibited a different regulatory ability of uptaking and transporting osmoprotection between strains with different desiccation tolerance abilities.
In addition to being transported into cells, a compatible solute such as endogenous trehalose can be synthesized in C. sakazakii cells (Hu et al., 2017). The trehalose biosynthesis pathway in C. sakazakii is similar to the trehalose-6-phosphate synthase/phosphatase (OtsA-OtsB) pathway in E. coli. Glucose is converted to glucose-6-phosphate. Then, glucose-6-phosphate is converted to trehalose-6-phosphate, which is subsequently converted to trehalose. The gene otsA encoding the trehalose-6-phosphate synthetase is strongly stimulated by the accumulation of K+, glutamate and salts of other monovalent cations in the primary response (Strom and Kaasen, 1993). In this study, quantitative real-time PCR results showed that otsA and otsB (ESA_01334) were upregulated in mRNA levels in the three strains except for strain ENS 70819. Breeuwer et al. (2003) found the accumulation of trehalose in dried stationary cells of C. sakazakii, and no accumulation was detected in dried exponential cells (Breeuwer et al., 2003). In this study, the cells were cultured to exponential phase, and the regulatory response of the genes relevant to trehalose synthesis was observed in C. sakazakii using quantitative real-time PCR. Furthermore, the upregulation level of otsB was higher than that of otsA, most likely because during the synthetic process by which trehalose-6-phosphate is converted to trehalose, more trehalose-6-phosphate-phosphatase was required.
Choline is an important compatible solute involved in resisting harm from adverse environments. Proteins BetA and BetB are associated with converting choline to betaine in E. coli. After being transported into cells, choline is converted to betaine aldehyde by choline dehydrogenase (BetA). In addition, betaine aldehyde is transformed to betaine, catalyzed by betaine aldehyde dehydrogenase (BetB) (Landfald and Strøm, 1986). BetA and BetB have been identified in C. sakazakii and the function verified in response to desiccation by comparative proteomic analysis (Hu et al., 2017). In this study, except for strain ENS 70819, the two genes betA (ESA_02049) and betB (ESA_02048) were all upregulated in the other three strains. Furthermore, the upregulation in the strains with a strong desiccation tolerance was more significant than that in the weaker ones, showing that the betaine synthesis has already started after 1 h of drying treatment and that the synthesis in strains with a strong desiccation tolerance was more active.
In summary, the regulatory mechanisms of desiccation resistance, such as K+ accumulation and efflux as well as the uptake and synthesis of compatible solutes, begin after 1 h of drying. Moreover, the expression of related genes at the mRNA level showed differences among C. sakazakii strains with different desiccation tolerance levels. The levels of these genes in strains with strong desiccation tolerance were higher than those in the strains weak desiccation tolerance. The higher levels of these genes may make strains more resistant to dry environments.
Conclusion
In this study, 42 C. sakazakii strains were evaluated for desiccation tolerance and subgrouped by MLST analysis. No significant correlation was observed between the ST and the desiccation tolerance of the strains. By contrast, biofilm formation ability and desiccation tolerance showed a positive correlation. Quantitative real-time PCR analysis indicated that 10 genes played major roles in the primary and secondary responses in desiccation tolerance. This study provides a useful reference for further study of the mechanism underlying the strong resistance of C. sakazakii to dry conditions, at the strain selection level and the gene selection level.
Data Availability Statement
All datasets [GENERATED/ANALYZED] for this study are included in the manuscript and the Supplementary Files.
Author Contributions
X-JD and X-YW contributed to the conception and the design of the study, performed the experiments, and the writing and editing of the manuscript. PL, X-JD, and SW contributed to conceived and designed of the work. XD made a great contribution by performing the experiments, by being involved in the process of the experimental design and the writing of the manuscript. All authors agreed to be accountable for the content of the work contributed to the conception of the study.
Funding
This work was supported by grants from Tianjin Science and Technology Commission (Project No. 18JCZDJC34300) and the Ministry of Science and Technology of the People’s Republic of China (Project No. 2014BAD04B03).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2018.02867/full#supplementary-material
Footnotes
References
Auger, S., Krin, E., Aymerich, S., and Gohar, M. (2006). Autoinducer 2 affects biofilm formation by Bacillus cereus. Appl. Environ. Microbiol. 72, 937–941. doi: 10.1128/AEM.72.1.937-941.2006
Baldwin, A., Loughlin, M., Caubilla-Barron, J., Kucerova, E., Manning, G., Dowson, C., et al. (2009). Multilocus sequence typing of Cronobacter sakazakii and Cronobacter malonaticus reveals stable clonal structures with clinical significance which do not correlate with biotypes. BMC Microbiol. 9:223. doi: 10.1186/1471-2180-9-223
Bar, M., von Hardenberg, J., Meron, E., and Provenzale, A. (2002). Modelling the survival of bacteria in drylands: the advantage of being dormant. Proc. Biol. Sci. 269, 937–942. doi: 10.1098/rspb.2002.1958
Barron, J. C., and Forsythe, S. J. (2007). Dry stress and survival time of Enterobacter sakazakii and other Enterobacteriaceae in dehydrated powdered infant formula. J. Food Prot. 70, 2111–2117. doi: 10.4315/0362-028X-70.9.2111
Bogino, P. C., Oliva Mde, L., Sorroche, F. G., and Giordano, W. (2013). The role of bacterial biofilms and surface components in plant-bacterial associations. Int. J. Mol. Sci. 14, 15838–15859. doi: 10.3390/ijms140815838
Breeuwer, P., Lardeau, A., Peterz, M., and Joosten, H. M. (2003). Desiccation and heat tolerance of Enterobacter sakazakii. J. Appl. Microbiol. 95, 967–973. doi: 10.1046/j.1365-2672.2003.02067.x
Burgess, C. M., Gianotti, A., Gruzdev, N., Holah, J., Knochel, S., Lehner, A., et al. (2016). The response of foodborne pathogens to osmotic and desiccation stresses in the food chain. Int. J. Food Microbiol. 221, 37–53. doi: 10.1016/j.ijfoodmicro.2015.12.014
Chen, J., Lee, S. M., and Mao, Y. (2004). Protective effect of exopolysaccharide colanic acid of Escherichia coli O157:H7 to osmotic and oxidative stress. Int. J. Food Microbiol. 93, 281–286. doi: 10.1016/j.ijfoodmicro.2003.12.004
Danese, N. P., Pratt, A. L., and Kolter, R. (2000). Exopolysaccharide production is required for development of Escherichia coli K-12 biofilm architecture. J. Bacteriol. 182, 3593–3596. doi: 10.1128/JB.182.12.3593-3596.2000
De Gelder, J., De Gussem, K., Vandenabeele, P., and Moens, L. (2007). Reference database of raman spectra of biological molecules. J. Raman Spectrosc. 38, 1133–1147. doi: 10.1002/jrs.1734
Du, X. J., Wang, F., Lu, X., Rasco, B. A., and Wang, S. (2012). Biochemical and genetic characteristics of Cronobacter sakazakii biofilm formation. Res. Microbiol. 163, 448–456. doi: 10.1016/j.resmic.2012.06.002
FAO/WHO. (2004). Enterobacter sakazakii and Other Micro-organisms in Powdered Infant Formula: Meeting Report. Geneva: WHO.
FAO/WHO. (2008). Enterobacter sakazakii (Cronobacter spp.) in Powdered Formulae: Meeting Report. Geneva: WHO.
Farmer, J. J., Asbury, A. M., Hickmann, F. W., and Brenner, D. J. (1980). Enterobacter sakazakii: a new species of Enterobacteriaceae isolated from clinical specimens. Int. J. Syst. Bacteriol. 30, 569–584. doi: 10.1099/00207713-30-3-569
Feeney, A., Johnston, C. D., Lucid, A., O’Mahony, J., Coffey, A., Lucey, B., et al. (2014). The role of the Cronobacter sakazakii prop. Gut Pathog. 6, 46–51. doi: 10.1186/s13099-014-0046-9
Feeney, A., and Sleator, R. D. (2011). An in silico analysis of osmotolerance in the emerging gastrointestinal pathogen Cronobacter sakazakii. Bioeng. Bugs 2, 260–270. doi: 10.4161/bbug.2.5.17238
Fei, P., Jiang, Y., Jiang, Y., Yuan, X., Yang, T., Chen, J., et al. (2017). Prevalence, molecular characterization, and antibiotic susceptibility of Cronobacter sakazakii isolates from powdered infant formula collected from chinese retail markets. Front. Microbiol. 8:2026. doi: 10.3389/fmicb.2017.02026
Feng, S. L., Eucker, T. P., Holly, M. K., Konkel, M. E., Lu, X., and Wang, S. (2014). Investigating the responses of Cronobacter sakazakii to garlic-drived. Appl. Environ. Microb. 80, 959–971. doi: 10.1128/AEM.03460-13
Finn, S., Condell, O., McClure, P., Amezquita, A., and Fanning, S. (2013). Mechanisms of survival, responses and sources of salmonella in low-moisture environments. Front. Microbiol. 4:331. doi: 10.3389/fmicb.2013.00331
Flemming, H. C., Wingender, J., Szewzyk, U., Steinberg, P., Rice, S. A., and Kjelleberg, S. (2016). Biofilms: an emergent form of bacterial life. Nat. Rev. Microbiol. 14, 563–575. doi: 10.1038/nrmicro.2016.94
Hansen, L. T., and Vogel, B. F. (2011). Desiccation of adhering and biofilm Listeria monocytogenes on stainless steel: survival and transfer to salmon products. Int. J. Food Microbiol. 146, 88–93. doi: 10.1016/j.ijfoodmicro.2011.01.032
Hariri, S., Joseph, S., and Forsythe, S. J. (2013). Cronobacter sakazakii ST4 strains and neonatal meningitis, United States. Emerg. Infect. Dis. 19, 175–177. doi: 10.3201/eid1901.120649
Heermann, R., Zigann, K., Gayer, S., Rodriguez-Fernandez, M., Banga, J. R., Kremling, A., et al. (2014). Dynamics of an interactive network composed of a bacterial two-component system, a transporter and K+ as mediator. PLoS One 9:e89671. doi: 10.1371/journal.pone.0089671
Hingston, P. A., Stea, E. C., Knøchel, S., and Hansen, T. (2013). Role of initial contamination levels, biofilm maturity and presence of salt and fat on desiccation survival of Listeria monocytogenes on stainless steel surfaces. Food Microbiol. 36, 46–56. doi: 10.1016/j.fm.2013.04.011
Hu, S., Yu, Y., Wu, X., Xia, X., Xiao, X., and Wu, H. (2017). Comparative proteomic analysis of Cronobacter sakazakii by iTRAQ provides insights into response to desiccation. Food Res. Int. 100, 631–639. doi: 10.1016/j.foodres.2017.06.051
Iversen, C., Lane, M., and Forsythe, S. J. (2004). The growth profile, thermotolerance and biofilm formation of Enterobacter sakazakii grown in infant formula milk. Lett. Appl. Microbiol. 38, 378–382. doi: 10.1111/j.1472-765X.2004.01507.x
Iversen, C., and Stephen, F. (2003). Risk profile of Enterobacter sakazakii, an emergent pathogen associated with infant milk formula. Trends Food Sci. Technol. 14, 443–454. doi: 10.1016/S0924-2244(03)00155-9
Joseph, S., and Forsythe, S. J. (2012). Insights into the emergent bacterial pathogen cronobacter spp., generated by multilocus sequence typing and analysis. Front. Microbiol. 3:397. doi: 10.3389/fmicb.2012.00397
Kempf, B., and Bremer, E. (1998). Uptake and synthesis of compatible solutes as microbial stress responses to high-osmolality environments. Arch. Microbiol. 170, 319–330. doi: 10.1007/s002030050649
Koseki, S., Nakamura, N., and Shiina, T. (2015). Comparison of desiccation tolerance among Listeria monocytoges Escherichia coli O157:H7,Salomonella enterica, and Cronobacter sakazakii in powdered infant formula. J. Food Protect. 78, 104–110. doi: 10.4315/0362-028X.JFP-14-249
Laermann, V., Cudic, E., Kipschull, K., Zimmann, P., and Altendorf, K. (2013). The sensor kinase KdpD of Escherichia coli senses external K+. Mol. Microbiol. 88, 1194–1204. doi: 10.1111/mmi.12251
Landfald, B., and Strøm, A. R. (1986). Choline-glycine betaine pathway confers a high level of osmotic tolerance in Escherichia coli. J. Bacteriol. 165, 849–855. doi: 10.1128/jb.165.3.849-855.1986
Le, P. T., Makhalanyane, T. P., Guerrero, L. D., Vikram, S., Van de Peer, Y., and Cowan, D. A. (2016). Comparative metagenomic analysis reveals mechanisms for stress response in hypoliths from extreme hyperarid deserts. Genome Biol. Evol. 8, 2737–2747. doi: 10.1093/gbe/evw189
Lebre, P. H., De Maayer, P., and Cowan, D. A. (2017). Xerotolerant bacteria: surviving through a dry spell. Nat. Rev. Microbiol. 15, 285–296. doi: 10.1038/nrmicro.2017.16
Livak, K. J., and Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25, 402–408. doi: 10.1006/meth.2001.1262
Lu, X., Al-Qadiri, H. M., Lin, M., and Rasco, B. A. (2011). Application of mid-infrared and raman spectroscopy to the study of bacteria. Food Bioproc. Tech. 4, 919–935. doi: 10.1007/s11947-011-0516-8
Lu, X., Samuelson, D. R., Rasco, B. A., and Konkel, M. E. (2012). Antimicrobial effect of diallyl sulphide on Campylobacter jejuni biofilms. J. Antimicrob. Chemother. 67, 1915–1926. doi: 10.1093/jac/dks138
Manoj, K. M. N., and Kumar, V. (2007). Role of bacterial OmpA and host cytoskeleton in the invasion. Pediatr. Res. 62, 664–669. doi: 10.1203/PDR.0b013e3181587864
Santos, H., and Da Costa, M. S. (2002). Compatible solutes of organisms that live in hot saline environments. Environ. Microbiol. 4, 501–509. doi: 10.1046/j.1462-2920.2002.00335.x
Scheepe-Leberlolhne, M., and Wagner, F. (1986). Optimization and preliminary characterization of an exopolysaccharide synthezised by Enterobacter sakazakii. Biotechnol. Lett. 8, 695–700. doi: 10.1007/BF01032564
Sleator, R. D., Wouters, J., Gahan, C. G., Abee, T., and Hill, C. (2001). Analysis of the role of OpuC, an osmolyte transport system, in salt tolerance and virulence potential of Listeria monocytogenes. Appl. Environ. Microb. 67, 2692–2698. doi: 10.1128/AEM.67.6.2692-2698.2001
Stephan, R., Grim, C. J., Gopinath, G. R., Mammel, M. K., Sathyamoorthy, V., Trach, L. H., et al. (2014). ). Re-examination of the taxonomic status of Enterobacter helveticus, Enterobacter pulveris and Enterobacter turicensis as members of the genus Cronobacter and their reclassification in the genera franconibacter gen. nov. and Siccibacter gen. nov. as Franconibacter helveticus comb. nov., Franconibacter pulveris comb. nov. and Siccibacter turicensis comb. nov., respectively. Int. J. Syst. Evol. Microbiol. 64, 3402–3410. doi: 10.1099/ijs.0.059832-0
Strom, A. R., and Kaasen, I. (1993). Trehalose metabolism in Escherichia coli: stress protection and stress regulation of gene expression. Mol. Microbiol. 8, 205–210. doi: 10.1111/j.1365-2958.1993.tb01564.x
Van der Laan, M., Gassel, M., and Altendorf, K. (2002). Characterization of amino acid substitutions in KdpA, the K+-binding and -translocating subunit of the KdpFABC complex of Escherichia coli. J. Bacteriol. 184, 5491–5494. doi: 10.1128/jb.184.19.5491-5494.2002
Welsh, D. T., and Herbert, R. A. (1999). Osmotically induced intracellular trehalose, but not glycine betaine accumulation promotes desiccation tolerance in Escherichia coli. FEMS Microbiol. Lett. 174, 57–63. doi: 10.1016/S0378-1097(99)00122-6
Keywords: Cronobacter sakazakii, multilocus sequence typing, desiccation tolerance, biofilm formation, desiccation tolerance related genes
Citation: Du X-j, Wang X-y, Dong X, Li P and Wang S (2018) Characterization of the Desiccation Tolerance of Cronobacter sakazakii Strains. Front. Microbiol. 9:2867. doi: 10.3389/fmicb.2018.02867
Received: 28 August 2018; Accepted: 07 November 2018;
Published: 27 November 2018.
Edited by:
Julio Parra-Flores, University of the Bío Bío, ChileReviewed by:
Ariadnna Cruz-Córdova, Hospital Infantil de México Federico Gómez, MexicoBen Davies Tall, United States Food and Drug Administration, United States
Copyright © 2018 Du, Wang, Dong, Li and Wang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Shuo Wang, s.wang@tust.edu.cn; wangshuo@nankai.edu.cn
†These authors have contributed equally to this work