
Abstract
Background
Alport syndrome (ATS) is a progressive hereditary nephropathy characterized by hematuria and proteinuria. It can be associated with extrarenal manifestations. In contrast, thin basement membrane nephropathy (TBMN) is characterized by microscopic hematuria, is largely asymptomatic, and is rarely associated with proteinuria and end-stage renal disease. Mutations have been identified in the COL4A5 gene in ATS and in the COL4A3 and COL4A4 genes in ATS and TBMN. To date, more than 1000 different mutations in COL4A5, COL4A3, and COL4A4 are known.
Methods
In this study mutational analysis by exon sequencing and multiplex ligation-dependent probe amplification was performed in a large European cohort of families with ATS and TBMN.
Results
Molecular diagnostic testing of 216 individuals led to the detection of 47 novel mutations, thereby expanding the spectrum of known mutations causing ATS and TBMN by up to 10 and 6 %, respectively, depending on the database. Remarkably, a high number of ATS patients with only single mutations in COL4A3 and COL4A4 were identified. Additionally, three ATS patients presented with synonymous sequence variants that possible affect correct mRNA splicing, as suggested by in silico analysis.
Conclusions
The results of this study clearly broaden the genotypic spectrum of known mutations for ATS and TBMN, which will in turn now facilitate future studies into genotype–phenotype correlations. Further studies should also examine the significance of single heterozygous mutations in COL4A3 and COL4A4 and of synonymous sequence variants associated with ATS.
Similar content being viewed by others

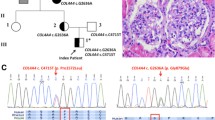

Introduction
Alport syndrome (ATS) is characterized by hematuria and/or proteinuria caused by structural defects of the glomerular basement membrane (GBM), with thinning and thickening due to splitting and multilamellation of the lamina densa [1]. In Western countries, it has a frequency of 1 in 5000 in the general population and is responsible for about 1–2 % of all cases with end-stage renal disease (ESRD) [2–4]. ATS can be associated with extrarenal manifestations, including high-tone sensorineural hearing loss and ocular abnormalities [2, 3, 5–7]. In comparison to ATS, thin basement membrane nephropathy (TBMN) is defined as uniformly thinning of the GBM characterized by persistent glomerular hematuria, minimal proteinuria, and normal renal function [8–11]. As the typical histopathologic features of TBMN are also found in early stages of ATS, the pathomechanisms of both diseases were initially postulated to overlap, which was confirmed in the early 1990s by genetic studies [11, 12]. The frequency of TBMN has been estimated to be as high as 1 % of the world population [13].
ATS and TBMN are genetically heterogeneous diseases caused by mutations in type IV collagen, the major component of basement membranes [6, 14]. Type IV collagen is composed of six homologous α-chains (α1–α6) encoded by the genes COL4A1–COL4A6, respectively. Alterations in any of the COL4A3, COL4A4, and COL4A5 genes can cause ATS. Different forms of inheritance have been described for ATS, including a X-linked form (85 % of the cases; mutations in COL4A5), an autosomal recessive form (most of the other patients; compound-heterozygous or homozygous mutations in COL4A3 or COL4A4), and an autosomal dominant form (rare; dominant mutations in COL4A3 or COL4A4) [2, 3, 5, 6, 15]. There is also evidence for digenic inheritance in ATS in addition to the classic Mendelian inheritance [16]. A relationship between the type and position of the mutation and the age at onset of ESRD, and the association of extrarenal manifestations has been observed in several studies [17–21]. Renal function in ATS patients significantly benefits from early and long-term treatment with angiotensinogen converting enzyme inhibitors (ACEi), as demonstrated in several studies [22, 23]. Therefore, a timely diagnosis of ATS in patients with hematuria and proteinuria has a high impact on the overall prognosis, which highlights the importance of genetic testing in early cases.
In contrast, TBMN is an autosomal dominant disease, and heterozygous mutations have been identified in COL4A3 and COL4A4 [8, 24]. In a few patients with TBMN disease progression to late development of proteinuria and ESRD has been reported. This disease progression seems to be related in part to the development of secondary focal segmental glomerulosclerosis (FSGS) [25, 26].
A correct clinical classification of the disease into ATS or TBMN is often difficult at onset, and here genetic testing of the relevant genes can be of considerable help to clarify the diagnosis. A large number of mutations causing ATS and TBMN, respectively, are collected in the following databases: ARUP® (ARUP® Laboratories), HGMD® (Human Gene Mutation Database, professional release 2015.1, Cardiff, UK), and LOVD (Leiden Open Variation Database, last update December 2013, Leiden, The Netherlands). To date, for COL4A5, 771 and 814 (HGMD®) different mutations causing ATS are listed ARUP® and HGMD®, respectively, and 1168 different mutations causing TBMN are listed in LOVD; for COL4A3, 156 mutations (ATS) and 266 (TBMN) mutations are listed in HGMD® and LOVD, respectively; for COL4A4, 115 (ATS) and 268 (TBMN) different mutations are listed in HGMD® and LOVD, respectively; all are listed in combination with their phenotype. Even though the analytical sensitivity for COL4A3–5 genes is >99 % [27] and many mutations and their associated clinical manifestations have been identified in a very high number of patients, it can be assumed that many mutations of these genes are still unidentified. As mentioned above, a genetic verification is essential for the patient’s prognosis and treatment, and therefore the number of causative mutations and their clinical phenotype must be increased. We here report the results of an extensive mutational analysis in a large European cohort of 216 individuals that led to the identification of 47 novel mutations in type IV collagen genes.
Material and methods
All individuals were clinically examined by human geneticists and adult or pediatric nephrologists. The study cohort consisted of patients mainly treated in Germany, especially at the University Children’s Hospital in Essen, Germany. The clinical classification into ATS or TBMN was performed by the treating physicians in accordance to current guidelines [15] but without any prior knowledge of the results of the mutational analysis. The clinical diagnosis and all available clinical data were re-analyzed for plausibility and corrected if necessary. As clinical data were sometimes incomplete (Tables 2–4), further analysis of clinical data, including intrafamilial data, was not performed in this study. The diagnosis of TBMN was given to patients with persistent isolated hematuria with/without low-grade proteinuria and only thinned GBM (if a biopsy result was available). A diagnosis of ATS was given to patients with glomerular hematuria or renal failure, a kidney biopsy result compatible with ATS (if available), a family history of ATS, and possibly extrarenal manifestations, such as hearing impairment or ocular abnormalities. All male patients with mutations in COL4A5 who were incorrectly diagnosed with TBMN were in retrospect reclassified as patients with ATS in accordance to current guidelines [15]. All female individuals with a mutation in COL4A5 were in retrospect classified as carriers of ATS.
Mutational analysis
Blood samples were collected from all patients after written informed consent. Genomic DNA was extracted from blood using the Gentra Puregene Blood kit (Qiagen, Hilden, Germany). Stepwise direct sequencing was performed depending on the clinical phenotype. Patients with ATS were sequenced as follows: COL4A5, COL4A3, COL4A4; COL4A3 followed by COL4A4 were examined in patients with TBMN. Mutational analysis was continued until a genetic diagnosis was established. Those patients for whom a diagnosis could not be verified or only a synonymous sequence variant (formerly described as silent mutation) could be identified, all currently known COL4A genes were examined. Sanger sequencing was carried out for both strands using the dideoxy chain termination method on an ABI capillary sequencer (model 3730; Applied Biosystems, Foster City, CA). Oligonucleotide primers were designed by the Primer3 program (http://frodo.wi.mit.edu/primer3/input.htm). DNA alignment and sequence variant analysis were carried out using Sequence PilotCE software (JSI medical systems GmbH, Kippenheim, Germany). If no mutation or only a synonymous sequence variant was identified in one of the COL4A3–5 genes, a MLPA (multiplex ligation-dependent probe amplification) assay for COL4A5 was performed using the two commercially available SALSA MLPA kits (P191/P192; MRC-Holland, Amsterdam, The Netherlands). The assay consists of two probe mixes containing probes for 48 of the 51 COL4A5 exons. In patients with ATS and a heterozygous mutation in COL4A3 or COL4A4, we performed a MLPA using the commercially available SALSA MLPA kits P439 (COL4A3) and P444 (COL4A4) (MRC-Holland). The COL4A3–MLPA kit contains probes for 33 of the 52 exons, and the COL4A4-MLPA kit contains 35 of the 48 exons. MLPAs were performed according to the manufacturer’s protocol. Fragment analysis was conducted using GeneMarker software (www.softgenetics.com; Soft Genetics, State College, PA). If possible, segregation analysis in the family was performed. In the relatives only the mutation identified in the index patient was tested by Sanger sequencing.
Evaluation of pathogenicity
The pathogenic potential of all identified mutations and sequence variants was analyzed by MutationTaster (http://www.mutationtaster.org) and PolyPhen-2 (http://genetics.bwh.harvard.edu/pph/). Splicing mutations cannot be analyzed by PolyPhen-2 and were therefore examined by Human Splicing Finder (http://www.umd.be/HSF3/index.html) in addition to MutationTaster. Sequence variants which did not result in an amino acid change but which were located next to the splice site were examined by SpliceView (http://bioinfo4.itb.cnr.it/∼webgene/wwwspliceview_ex.html) in addition to Human Splicing Finder and MutationTaster. Mutations which could not be classified with either of the programs were declared as “not applicable”.
All mutations identified in this study have been checked against ExAC Browser (http://exac.broadinstitute.org), ARUP® (http://www.arup.utah.edu/database/), HGMD® (http://www.hgmd.cf.ac.uk/ac/), and LOVD (http://www.lovd.nl/3.0/home). Furthermore, all novel mutations identified in this study were excluded in 100 ethnically matched control chromosomes.
Results
The total study cohort consisted of 216 individuals (167 patients and 49 relatives) from 91 different families, among whom 111 (51 %) were female and 105 (49 %) were male. Age at diagnostics ranged from 9 days up to 69 years (median 18 years), and 75 patients (35 %) were ≤18 years of age. Consanguinity was known to be present in three (1 %) patients (Table 1, index patients 10320043 and 12300176; Table 2, index patient 11430427). Altogether, five different countries of origin were represented within the cohort: Germany, Austria, Italy, Spain, and Turkey. A kidney biopsy was performed in 35 index patients of different families with ATS and in four index patients of different families with TBMN. Within the families with TBMN the clinical phenotype was consistent in all family members (one exception: Table 1, index patient 11470286). Klinefelter syndrome was confirmed in one patient by chromosomal analysis (Table 1, index patient 10480239).
Mutations in the COL4A3 gene
Twenty-nine patients from 17 different families carried a mutation in COL4A3 (see Table 1, only index patient shown). Of these 17 families, 14 (82 %) and three (18 %) were clinically diagnosed with ATS and TBMN, respectively. Seven (50 %) of the families with ATS were carriers of either a homozygous mutation because of consanguineous parents (Table 1, index patients 10320043 and 12300176) or compound heterozygous mutations in COL4A3. In seven (50 %) families with clinically diagnosed ATS only one heterozygous mutation in COL4A3 was found. The results of the MLPA assay of COL4A3 was unremarkable in these patients (DNA of patient 12410300 was unavailable for this analysis). All families with TBMN (100 %) presented with only one COL4A3 mutation. In patient 10070157 a de-novo mutation was found.
Overall, 16 novel mutations were identified in COL4A3 (Tables 1, 3). Thirteen novel mutations (81 %) were classified as disease causing by both prediction programs (PolyPhen-2, MutationTaster; Table 1) or as disease causing by one and as “not applicable” by the other. Three mutations (19 %) were judged to be benign by one of these prediction programs, but as disease causing by the other. One novel sequence variant was classified by MutationTaster as polymorphism but as probably damaging by PolyPhen-2.
Of the 21 different COL4A3 mutations identified among the study cohort, 13 (62 %) were glycine mutations and four (19 %) were other missense exchanges (Table 4). The remaining mutation types were classified as depicted in Table 4. In one family a synonymous sequence variant was observed in COL4A3 located at the splice donor site (Table 1, index patient 13200065, p.Pro993Pro). The analysis of the pathogenic potential of this sequence variant by Human Splicing Finder, MutationTaster, and SpliceView suggested functional loss of the affected protein caused by aberrant splicing.
Mutations in the COL4A4 gene
Thirty-one patients from 20 different families carried a mutation in COL4A4 (Table 2, only index patient shown). Of these families, six (30 %) and 11 (55 %) were clinically diagnosed with ATS and TBMN, respectively, and samples of three families (15 %) were sent for molecular diagnostics without further clinical information. One patient with ATS (12 %) had a homozygous mutation because of consanguineous parents (Table 2, index patient 11430427). Five of the six families with ATS (83 %) were carriers of only one heterozygous mutation in COL4A4. The results of the MLPA assay of COL4A4 were unremarkable in these patients (DNA of patient 11120210 was unavailable for this analysis). All patients with TBMN (100 %) presented with only one COL4A4 mutation.
Five novel mutations were identified in COL4A4 (Tables 2, 3). Four of these novel mutations (80 %) were classified as disease causing by both prediction programs (PolyPhen-2, MutationTaster; Table 2) or as disease causing by one and not applicable by the other. The remaining novel sequence variant was classified by MutationTaster as polymorphism, but was classified as not applicable by PolyPhen-2. Of the 15 identified different COL4A4 mutations, six (40 %) were glycine mutations and two (13 %) were other missense exchanges (Table 4); the remaining mutations were classified as shown in Table 4.
Mutations in the COL4A5 gene
Eighty-five patients from 54 different families carried a mutation in COL4A5 (Table 5). Of these 54 families, 44 (81 %) were clinically diagnosed with ATS by the referring physician; nine families (17 %) were diagnosed with TBMN by the referring physician; samples of one family (2 %) were sent for molecular diagnostics without further clinical information. Three male patients (age 6, 12, and 16 years, respectively) were falsely diagnosed with TBMN. According to the guidelines, all male patients with mutations in COL4A5 have to be correctly diagnosed with ATS. All female individuals with a mutation in COL4A5 were classified as carriers of ATS [15].
Twenty-six novel mutations were identified in the COL4A5 gene (Tables 3, 5). Twenty novel mutations (77 %) were classified as disease causing by both prediction programs (PolyPhen-2, MutationTaster; Table 5) or classified as disease causing by one and not applicable by the the other. Two novel sequence variants were classified by MutationTaster as polymorphism and were not applicable by PolyPhen-2 (Table 5, upper half, index patient 11200302; Table 5, lower half, index patient 13100284). Four mutations were not applicable by MutationTaster and Polyphen-2.
Of the 43 different COL4A5 mutations identified, 15 (35 %) were glycine mutations, and five (12 %) were other missense exchanges (Table 4). Splice site mutations in COL4A5 were very common (23 %). The remaining mutations were classified as shown in Table 4. In two families synonymous sequence variants were observed in COL4A5 located at the splice donor site (Table 5, upper half, index patient 12290515, p.Lys128Lys; Table 5, lower half, index patient 12290425, p.Lys715Lys). The analysis of the pathogenic potential of these identified sequence variants using Human Splicing Finder, MutationTaster, and SpliceView suggested functional loss of the affected protein caused by aberrant splicing.
Discussion
This study encompasses one of the largest European patient cohorts leading to the identification of 47 novel mutations in the COL4A3, COL4A4, and COL4A5 genes. The results of this study increase the number of known mutations for ATS and TBMN by up to 10 and 6 %, respectively, depending on the underlying database. To date, a mutational hot spot has not been detected and was also not identified in our study.
The identification of novel mutations causing ATS and TBMN is important for screening and for the prognosis of symptomatic and at-risk family members, as well as for the planning of kidney donations within a family [27]. This is especially relevant in children with ATS as an early diagnosis and therapy can postpone ESRD [22, 42]. Blood sampling for genetic testing is a less invasive procedure than a kidney biopsy and, taking the extensive knowledge of genotype–phenotype correlations in COL4A3, COL4A4, and COL4A5 into account, genetic diagnostic testing could possibly replace the kidney biopsy in special cases. Furthermore, it should be noted that especially in the early stages of ATS - particularly in children - typical histopathologic features of TBMN can be found in ATS and, therefore, a correct diagnosis cannot be made at young age in some patients by kidney biopsy alone [11, 12]. The clear clinical overlap between TBMN and early ATS is also reflected by the genetic overlap (heterozygous COL4A3 and COL4A4 mutations in TBMN and autosomal dominant mutations in ATS). This observation complicates genetic counseling, especially in pediatric patients with familial hematuria. More clinical and genetic data are therefore needed to clarify this issue with respect to prevalence, progression, and treatment options. We strongly believe that our findings add considerable data to this important aspect, which is of clinical relevance for the patients and their families.
Interestingly, a high number of index patients with ATS with only one heterozygous mutation in COL4A3 (46 %) or COL4A4 (83 %) were observed in our study cohort. Missed rearrangements and cryptic splice site mutations could be a possible explanation [43]. Furthermore, large deletions or duplications in COL4A3 and COL4A4 are suggested in the literature and could also account for some mutations currently not being detected by Sanger sequencing [19]. MLPA analysis is necessary for the detection of gene deletions or duplications, and we used commercially available kits for COL4A3, COL4A4, and COL4A5 to exclude this kind of mutation. Large deletions and duplications account for approximately 12 % of patients with X-linked ATS, but the frequencies for large deletions and duplications in COL4A3 or COL4A4 are unknown at the present time [18]. Another - albeit unlikely - option are mutations in up to now unknown genes with relevance for ATS alone or in combination with an identified heterozygous mutation. Pathogenic deep intronic mutations have also been reported but cannot be covered by Sanger sequencing [44]. This could be considered as a limitation in our study, and newer sequencing techniques (e.g. targeted next generation sequencing) will allow the analysis of these intronic mutations [16]. Additionally, digenic mutations may have been missed by Sanger sequencing which would not be overlooked by NGS.
Furthermore, it has to be considered that single heterozygous mutations in COL4A3 and COL4A4 are also compatible with an autosomal dominant inheritance in some of the families in our study. However, up to now, the autosomal dominant form of ATS is described as being very rare even though there are new data proposing an autosomal dominant pattern of inheritance in up to 31 % of families with ATS [4, 15].
Only in one patient (Table 1, index patient 10070157) was a de-novo mutation in COL4A3 observed. In COL4A4 and COL4A5 de-novo mutations were not seen, but in some families of our study segregation analysis could not be performed because parental samples were not available. The frequency of de-novo mutations in COL4A5 has been reported to be 10–15 % in the literature, and this is also assumed for the frequency of COL4A3 and COL4A4, respectively [11, 45, 46]. This high rate of de-novo mutations was not observed in our study because the study population mainly consisted of familial cases. The possibility of de-novo mutations should always be considered in patients with signs of ATS or TBMN without a positive family history.
As a special type of mutation three different synonymous sequence variants were detected in this study, all directly located at the splice donor site (Table 1, index patient 13200065, p.Pro993Pro; Table 5, upper half, index patient 12290515, p.Lys128Lys; Table 5, lower half, index patient 12290425, p.Lys715Lys). Although this type of sequence variant is believed to be of neutral effect [47], analysis of the pathogenic potential of the splice sites of the three sequence variants suggested that the nucleotide changes most likely influence the splicing effect. Therefore, these synonymous sequence variants would appear to be a rare cause of ATS. However, because functional analysis of potential splice site variants outside the consensus splice sites and of synonymous sequence variants was not performed, these sequence variants should be classified as sequence variants of unknown significance.
Concerning the three male patients with clinical diagnosis of TBMN and a COL4A5 mutation, the young age of these patients at the time of the study (6, 12, and 16 years, respectively) has to be pointed out. This could be a reason for a yet incomplete development of the clinical symptoms of ATS. The 6-year-old boy exhibited only hematuria at the time of the analysis, while the 12-year-old boy already presented with low-grade proteinuria. Age-related changes of GBM lesions ranging from thin GBM at the age of 9 years, suggestive for TBMN, to thickened GBM at 21 years of age, compatible with the diagnosis of ATS were demonstrated by Cangiotti et al. [12] in a patient with hematuria and slight proteinuria based on renal biopsy. The 16-year-old patient who presented with isolated hematuria had a splice site sequence variant +9. This sequence variant, although predicted as polymorphism, might be responsible for a milder phenotype, as his affected sister, mother, and maternal grandmother also presented only with isolated hematuria. Additionally, Storey et al. also reported the cases of a number of adult patients as well as children with X-linked ATS and normal renal function and/or minor/nonspecific histologic abnormalities mostly seen in early disease [18].
With respect to the families affected by heterozygous autosomal COL4A3 mutations and a clinical phenotype of TBMN, one phenotypic discrepancy in index patient 11470286 (Table 1) has to be pointed out. Although the index patient was clinically diagnosed as having TBMN, the mother had been diagnosed with ATS. Therefore, it is possible that this patient will develop ATS in adulthood under the assumption of an autosomal dominant mode of inheritance. Unfortunately, the genetic result of the mother is not available to confirm the possible mode of inheritance.
Most families with ATS have private mutations [48]. To the contrary, the sequence variant p.Gly624Asp in COL4A5 was observed in eight different families (15 %) in our study. One of the index patients is the 6-year-old boy already mentioned above with the clinical phenotype of TBMN. All other patients presented clinically with ATS. Amongst others, Šlajpah et al. also identified this sequence variant in a family with a progressive form of ATS as well as in families affected with TBMN, suggesting the possibility of significantly different phenotypes associated with the same gene mutation [49]. Additionally, Demosthenous et al. observed a wide phenotypical spectrum of this sequence variant ranging from isolated microhematuria to severe ATS [50]. p.Gly624Asp is reported to be located either on a recurrently mutated position; alternatively, it might be an example of a trans-national founder mutation [51]. As p.Gly624Asp is described as probably being a hypomorphic missense mutation mimicking the phenotypes of autosomal recessive heterozygous COL4A3 and COL4A4 mutations leading to microhematuria and TBMN, screening of COL4A5 should be considered in patients with TBMN and inconspicuous mutational analysis of COL4A3 and COL4A4 mutation [52].
In conclusion, our findings significantly increase the number of known COL4A3-5 mutations and thereby strengthen our knowledge with respect to the genetic background of TBMN and ATS. Further studies should specifically elucidate the occurrence and significance of single heterozygous mutations in COL4A3 and COL4A4 associated with Alport syndrome and investigate the possibility of an autosomal dominant mode of inheritance.
References
Noone D, Licht C (2013) An update on the pathomechanisms and future therapies of Alport syndrome. Pediatr Nephrol 28:1025–1036
Marcocci E, Uliana V, Bruttini M, Artuso R, Silengo MC, Zerial M, Bergesio F, Amoroso A, Savoldi S, Pennesi M, Giachino D, Rombola G, Fogazzi GB, Rosatelli C, Martinhago CD, Carmellini M, Mancini R, Di Costanzo G, Longo I, Renieri A, Mari F (2009) Autosomal dominant Alport syndrome: molecular analysis of the COL4A4 gene and clinical outcome. Nephrol Dial Transplant 24:1464–1471
Nagel M, Nagorka S, Gross O (2005) Novel COL4A5, COL4A4, and COL4A3 mutations in Alport syndrome. Hum Mutat 26:60
Fallerini C, Dosa L, Tita R, Del Prete D, Feriozzi S, Gai G, Clementi M, La Manna A, Miglietti N, Mancini R, Mandrile G, Ghiggeri GM, Piaggio G, Brancati F, Diano L, Frate E, Pinciaroli AR, Giani M, Castorina P, Bresin E, Giachino D, De Marchi M, Mari F, Bruttini M, Renieri A, Ariani F (2014) Unbiased next generation sequencing analysis confirms the existence of autosomal dominant Alport syndrome in a relevant fraction of cases. Clin Genet 86:252–257
Hou P, Chen Y, Ding J, Li G, Zhang H (2007) A novel mutation of COL4A3 presents a different contribution to Alport syndrome and thin basement membrane nephropathy. Am J Nephrol 27:538–544
Cook C, Friedrich CA, Baliga R (2008) Novel COL4A3 mutations in African American siblings with autosomal recessive Alport syndrome. Am J Kidney Dis 51:e25–e28
Longo I, Scala E, Mari F, Caselli R, Pescucci C, Mencarelli MA, Speciale C, Giani M, Bresin E, Caringella DA, Borochowitz ZU, Siriwardena K, Winship I, Renieri A, Meloni I (2006) Autosomal recessive Alport syndrome: an in-depth clinical and molecular analysis of five families. Nephrol Dial Transplant 21:665–671
Baek JI, Choi SJ, Park SH, Choi JY, Kim CD, Kim YL, Kim UK (2009) Identification of novel variants in the COL4A4 gene in Korean patients with thin basement membrane nephropathy. Indian J Med Res 129:525–533
Fujinaga S, Kaneko K, Ohtomo Y, Murakami H, Takemoto M, Takada M, Shimizu T, Yamashiro Y (2006) Thin basement membrane nephropathy associated with minimal change disease in a 15-year-old boy. Pediatr Nephrol 21:277–280
Voskarides K, Pierides A, Deltas C (2008) COL4A3/COL4A4 mutations link familial hematuria and focal segmental glomerulosclerosis. glomerular epithelium destruction via basement membrane thinning? Connect Tissue Res 49:283–288
Tryggvason K, Patrakka J (2006) Thin basement membrane nephropathy. J Am Soc Nephrol 17:813–822
Cangiotti AM, Sessa A, Meroni M, Montironi R, Ragaiolo M, Mambelli V, Cinti S (1996) Evolution of glomerular basement membrane lesions in a male patient with Alport syndrome: ultrastructural and morphometric study. Nephrol Dial Transplant 11:1829–1834
Savige J, Rana K, Tonna S, Buzza M, Dagher H, Wang YY (2003) Thin basement membrane nephropathy. Kidney Int 64:1169–1178
Gubler MC (2007) Diagnosis of Alport syndrome without biopsy? Pediatr Nephrol 22:621–625
Savige J, Gregory M, Gross O, Kashtan C, Ding J, Flinter F (2013) Expert guidelines for the management of Alport syndrome and thin basement membrane nephropathy. J Am Soc Nephrol 24:364–375
Mencarelli MA, Heidet L, Storey H, van Geel M, Knebelmann B, Fallerini C, Miglietti N, Antonucci MF, Cetta F, Sayer JA, van den Wijngaard A, Yau S, Mari F, Bruttini M, Ariani F, Dahan K, Smeets B, Antignac C, Flinter F, Renieri A (2015) Evidence of digenic inheritance in Alport syndrome. J Med Genet 52:163–174
Bekheirnia MR, Reed B, Gregory MC, McFann K, Shamshirsaz AA, Masoumi A, Schrier RW (2010) Genotype-phenotype correlation in X-linked Alport syndrome. J Am Soc Nephrol 21:876–883
Storey H, Savige J, Sivakumar V, Abbs S, Flinter FA (2013) COL4A3/COL4A4 mutations and features in individuals with autosomal recessive Alport syndrome. J Am Soc Nephrol 24:1945–1954
Jais JP, Knebelmann B, Giatras I, De Marchi M, Rizzoni G, Renieri A, Weber M, Gross O, Netzer KO, Flinter F, Pirson Y, Verellen C, Wieslander J, Persson U, Tryggvason K, Martin P, Hertz JM, Schroder C, Sanak M, Krejcova S, Carvalho MF, Saus J, Antignac C, Smeets H, Gubler MC (2000) X-linked Alport syndrome: natural history in 195 families and genotype-phenotype correlations in males. J Am Soc Nephrol 11:649–657
Gross O, Netzer KO, Lambrecht R, Seibold S, Weber M (2002) Meta-analysis of genotype-phenotype correlation in X-linked Alport syndrome: impact on clinical counselling. Nephrol Dial Transplant 17:1218–1227
Jais JP, Knebelmann B, Giatras I, De Marchi M, Rizzoni G, Renieri A, Weber M, Gross O, Netzer KO, Flinter F, Pirson Y, Dahan K, Wieslander J, Persson U, Tryggvason K, Martin P, Hertz JM, Schroder C, Sanak M, Carvalho MF, Saus J, Antignac C, Smeets H, Gubler MC (2003) X-linked Alport syndrome: natural history and genotype-phenotype correlations in girls and women belonging to 195 families: a “European Community Alport Syndrome Concerted Action” study. J Am Soc Nephrol 14:2603–2610
Study Group Members of the Gesellschaft fur Padiatrische N, Gross O, Licht C, Anders HJ, Hoppe B, Beck B, Tonshoff B, Hocker B, Wygoda S, Ehrich JH, Pape L, Konrad M, Rascher W, Dotsch J, Muller-Wiefel DE, Hoyer P, Knebelmann B, Pirson Y, Grunfeld JP, Niaudet P, Cochat P, Heidet L, Lebbah S, Torra R, Friede T, Lange K, Muller GA, Weber M (2012) Early angiotensin-converting enzyme inhibition in Alport syndrome delays renal failure and improves life expectancy. Kidney Int 81:494–501
Proesmans W, Van Dyck M (2004) Enalapril in children with Alport syndrome. Pediatr Nephrol 19:271–275
Rana K, Tonna S, Wang YY, Sin L, Lin T, Shaw E, Mookerjee I, Savige J (2007) Nine novel COL4A3 and COL4A4 mutations and polymorphisms identified in inherited membrane diseases. Pediatr Nephrol 22:652–657
Pierides A, Voskarides K, Athanasiou Y, Ioannou K, Damianou L, Arsali M, Zavros M, Pierides M, Vargemezis V, Patsias C, Zouvani I, Elia A, Kyriacou K, Deltas C (2009) Clinico-pathological correlations in 127 patients in 11 large pedigrees, segregating one of three heterozygous mutations in the COL4A3/COL4A4 genes associated with familial haematuria and significant late progression to proteinuria and chronic kidney disease from focal segmental glomerulosclerosis. Nephrol Dial Transplant 24:2721–2729
Voskarides K, Damianou L, Neocleous V, Zouvani I, Christodoulidou S, Hadjiconstantinou V, Ioannou K, Athanasiou Y, Patsias C, Alexopoulos E, Pierides A, Kyriacou K, Deltas C (2007) COL4A3/COL4A4 mutations producing focal segmental glomerulosclerosis and renal failure in thin basement membrane nephropathy. J Am Soc Nephrol 18:3004–3016
Hertz JM, Thomassen M, Storey H, Flinter F (2015) Clinical utility gene card for: Alport syndrome - update 2014. Eur J Hum Genet. doi:10.1038/ejhg.2014.254
Wang YY, Rana K, Tonna S, Lin T, Sin L, Savige J (2004) COL4A3 mutations and their clinical consequences in thin basement membrane nephropathy (TBMN). Kidney Int 65:786–790
Heidet L, Arrondel C, Forestier L, Cohen-Solal L, Mollet G, Gutierrez B, Stavrou C, Gubler MC, Antignac C (2001) Structure of the human type IV collagen gene COL4A3 and mutations in autosomal Alport syndrome. J Am Soc Nephrol 12:97–106
Chatterjee R, Hoffman M, Cliften P, Seshan S, Liapis H, Jain S (2013) Targeted exome sequencing integrated with clinicopathological information reveals novel and rare mutations in atypical, suspected and unknown cases of Alport syndrome or proteinuria. PLoS One 8:e76360
Lemmink HH, Nillesen WN, Mochizuki T, Schroder CH, Brunner HG, van Oost BA, Monnens LA, Smeets HJ (1996) Benign familial hematuria due to mutation of the type IV collagen alpha4 gene. J Clin Invest 98:1114–1118
Knebelmann B, Deschenes G, Gros F, Hors MC, Grunfeld JP, Zhou J, Tryggvason K, Gubler MC, Antignac C (1992) Substitution of arginine for glycine 325 in the collagen alpha 5 (IV) chain associated with X-linked Alport syndrome: characterization of the mutation by direct sequencing of PCR-amplified lymphoblast cDNA fragments. Am J Hum Genet 51:135–142
Martin P, Heiskari N, Zhou J, Leinonen A, Tumelius T, Hertz JM, Barker D, Gregory M, Atkin C, Styrkarsdottir U, Neumann H, Springate J, Shows T, Pettersson E, Tryggvason K (1998) High mutation detection rate in the COL4A5 collagen gene in suspected Alport syndrome using PCR and direct DNA sequencing. J Am Soc Nephrol 9:2291–2301
Beicht S, Strobl-Wildemann G, Rath S, Wachter O, Alberer M, Kaminsky E, Weber LT, Hinrichsen T, Klein HG, Hoefele J (2013) Next generation sequencing as a useful tool in the diagnostics of mosaicism in Alport syndrome. Gene 526:474–477
Boye E, Flinter F, Zhou J, Tryggvason K, Bobrow M, Harris A (1995) Detection of 12 novel mutations in the collagenous domain of the COL4A5 gene in Alport syndrome patients. Hum Mutat 5:197–204
Plant KE, Green PM, Vetrie D, Flinter FA (1999) Detection of mutations in COL4A5 in patients with Alport syndrome. Hum Mutat 13:124–132
Massella L, Gangemi C, Giannakakis K, Crisafi A, Faraggiana T, Fallerini C, Renieri A, Muda AO, Emma F (2013) Prognostic value of glomerular collagen IV immunofluorescence studies in male patients with X-linked Alport syndrome. Clin J Am Soc Nephrol 8:749–755
Hashimura Y, Nozu K, Kaito H, Nakanishi K, Fu XJ, Ohtsubo H, Hashimoto F, Oka M, Ninchoji T, Ishimori S, Morisada N, Matsunoshita N, Kamiyoshi N, Yoshikawa N, Iijima K (2014) Milder clinical aspects of X-linked Alport syndrome in men positive for the collagen IV alpha5 chain. Kidney Int 85:1208–1213
Wang F, Zhao D, Ding J, Zhang H, Zhang Y, Yu L, Xiao H, Yao Y, Zhong X, Wang S (2012) Skin biopsy is a practical approach for the clinical diagnosis and molecular genetic analysis of X-linked Alport’s syndrome. J Mol Diagn 14:586–593
Hertz JM, Juncker I, Marcussen N (2008) MLPA and cDNA analysis improves COL4A5 mutation detection in X-linked Alport syndrome. Clin Genet 74:522–530
Knebelmann B, Breillat C, Forestier L, Arrondel C, Jacassier D, Giatras I, Drouot L, Deschenes G, Grunfeld JP, Broyer M, Gubler MC, Antignac C (1996) Spectrum of mutations in the COL4A5 collagen gene in X-linked Alport syndrome. Am J Hum Genet 59:1221–1232
Gross O, Friede T, Hilgers R, Gorlitz A, Gavenis K, Ahmed R, Durr U (2012) Safety and efficacy of the ACE-inhibitor Ramipril in Alport syndrome: the double-blind, randomized, placebo-controlled, multicenter phase III EARLY PRO-TECT Alport trial in pediatric patients. ISRN Pediatr 2012:436046
Miner JH, Baigent C, Flinter F, Gross O, Judge P, Kashtan CE, Lagas S, Savige J, Blatt D, Ding J, Gale DP, Midgley JP, Povey S, Prunotto M, Renault D, Skelding J, Turner AN, Gear S (2014) The 2014 International Workshop on Alport Syndrome. Kidney Int 86:679–684
King K, Flinter FA, Nihalani V, Green PM (2002) Unusual deep intronic mutations in the COL4A5 gene cause X linked Alport syndrome. Hum Genet 111:548–554
Tryggvason K, Zhou J, Hostikka SL, Shows TB (1993) Molecular genetics of Alport syndrome. Kidney Int 43:38–44
Anton-Martin P, Aparicio Lopez C, Ramiro-Leon S, Santillan Garzon S, Santos-Simarro F, Gil-Fournier B (2012) Alport syndrome: de novo mutation in the COL4A5 gene converting glycine 1205 to valine. Clin Med Insights Pediatr 6:41–49
Strachan T, Read AP (1999) Human molecular genetics. Wiley-Liss, New York
Kashtan CE (1999) Alport syndrome. An inherited disorder of renal, ocular, and cochlear basement membranes. Medicine (Baltimore) 78:338–360
Slajpah M, Gorinsek B, Berginc G, Vizjak A, Ferluga D, Hvala A, Meglic A, Jaksa I, Furlan P, Gregoric A, Kaplan-Pavlovcic S, Ravnik-Glavac M, Glavac D (2007) Sixteen novel mutations identified in COL4A3, COL4A4, and COL4A5 genes in Slovenian families with Alport syndrome and benign familial hematuria. Kidney Int 71:1287–1295
Demosthenous P, Voskarides K, Stylianou K, Hadjigavriel M, Arsali M, Patsias C, Georgaki E, Zirogiannis P, Stavrou C, Daphnis E, Pierides A, Deltas C, Hellenic Nephrogenetics Research C (2012) X-linked Alport syndrome in Hellenic families: phenotypic heterogeneity and mutations near interruptions of the collagen domain in COL4A5. Clin Genet 81:240–248
Deltas C, Pierides A, Voskarides K (2012) The role of molecular genetics in diagnosing familial hematuria(s). Pediatr Nephrol 27:1221–1231
Pierides A, Voskarides K, Kkolou M, Hadjigavriel M, Deltas C (2013) X-linked, COL4A5 hypomorphic Alport mutations such as G624D and P628L may only exhibit thin basement membrane nephropathy with microhematuria and late onset kidney failure. Hippokratia 17:207–213
Barker DF, Denison JC, Atkin CL, Gregory MC (2001) Efficient detection of Alport syndrome COL4A5 mutations with multiplex genomic PCR-SSCP. Am J Med Genet 98:148--160
Acknowledgments
We sincerely thank the patients and the physicians for participation in this study.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Approval for this study was obtained from the Ethics Committee of the Ludwig-Maximilians University, Munich, Germany. Written informed consent was obtained.
Conflict of interest
The authors declare no conflict of interest.
Additional information
Stefanie Weber and Katja Strasser contributed equally to this work.
Rights and permissions
About this article

Cite this article
Weber, S., Strasser, K., Rath, S. et al. Identification of 47 novel mutations in patients with Alport syndrome and thin basement membrane nephropathy. Pediatr Nephrol 31, 941–955 (2016). https://doi.org/10.1007/s00467-015-3302-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00467-015-3302-4