
Differential attraction of drosophilids to banana baits inoculated with Saccharomyces cerevisiae and Hanseniaspora uvarum within a Neotropical forest remnant
- Published
- Accepted
- Received
- Academic Editor
- Therese Markow
- Subject Areas
- Biodiversity, Ecology, Entomology, Microbiology, Mycology
- Keywords
- Food preference, Yeast, Drosophila, Drosophila assemblage, Atlantic Rainforest, Tripunctata, Willistoni, Melanogaster, Exotic drosophila, Guarani
- Copyright
- © 2017 Batista et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2017. Differential attraction of drosophilids to banana baits inoculated with Saccharomyces cerevisiae and Hanseniaspora uvarum within a Neotropical forest remnant. PeerJ 5:e3063 https://doi.org/10.7717/peerj.3063
Abstract
Background
Yeasts are a necessary requisite in the diet of most Drosophila species that, in turn, may vector their dispersal in natural environments. Differential attractiveness experiments and the isolation of yeasts consumed by Drosophila may be informative for characterizing this association. Hanseniaspora uvarum is among the most common yeast species isolated from Drosophila crops, with high attractiveness to drosophilids. Saccharomyces cerevisiae has been widely used to collect flies, and it allows broad sampling of almost all local Drosophila species. Pronounced differences in the field concerning Drosophila attractivity to baits seeded with these yeast species have been previously reported. However, few explicit generalizations have been set. Since late fifties, no field experiments of Drosophila attractivity were carried out in the Neotropical region, which is facing shifts in abiotic and biotic factors. Our objective is to characterize preference behavior that mediates the interaction in the wild among Neotropical Drosophila species and yeasts associated with them. We want to set a broad generalization about drosophilids attracted to these yeasts. Here we present the results of a differential attractiveness experiment we carried out in a natural Atlantic Rainforest fragment to assess the preferences of Drosophila species groups to baits inoculated with H. uvarum and S. cerevisiae.
Methods
Both yeast species were cultured in GYMP broth and separately poured in autoclaved mashed banana that was left fermenting. In the field, we collected drosophilids over five arrays of three different baits: non-inoculated autoclaved banana and banana inoculated with each yeast. In the laboratory the drosophilids were sorted to five sets according to their external morphology and/or genitalia: tripunctata; guarani; willistoni; exotic; and the remaining flies pooled in others.
Results and Conclusions
Uninoculated banana baits attracted virtually no flies. We found significant departures from random distribution over the other two baits (1:1 proportion) for all sets, except the pooled others. Flies of the sets willistoni and exotic preferred H. uvarum over S. cerevisiae, while the remaining sets were more attracted to S. cerevisiae. Previously, various authors reported similar patterns in attraction experiments with S. cerevisiae and H. uvarum. It is also noteworthy that both yeast species have been isolated from natural substrates and crops of Drosophila species. Taken together, these results suggest that the preferences among Drosophila species groups may be reflecting deep and stable relations with yeast species in natural environments. They can be summarized as: forest dwelling species from subgenus Drosophila (such as tripunctata and guarani groups) are attracted to banana baits seeded with S. cerevisiae; while exotic (as D. melanogaster) and subgenus Sophophora species are preferentially attracted to baits seeded with H. uvarum.
Introduction
Drosophila-yeast association can be considered a diffuse mutualism (Starmer & Lachance, 2011; Buser et al., 2014), in which yeasts are a necessary requisite in the diet of most Drosophila species (Powell, 1997) that, in turn, may vector their dispersal in natural environments (Ganter, 1988; Christiaens et al., 2014). Moreover, Drosophila adults and larvae regulate yeast composition and density in natural substrates (Stamps et al., 2012), while different yeast species affect Drosophila breeding (Barker, 1992) and feeding preferences (Becher et al., 2012) as well as bionomic features (Anagnostou, Dorsch & Rohlfs, 2010).
Traditionally, this association is characterized by isolating yeasts from Drosophila crops (Phaff et al., 1956) and natural substrates (Carson, Knapp & Phaff, 1956; Starmer, 1981; Barker, Starmer & Vacek, 1987); also, by investigating Drosophila species attraction to baits inoculated with different yeast species in the field (Da Cunha, Dobzhansky & Sokoloff, 1951; Klaczko, Powell & Taylor, 1983) and in the laboratory (Barker et al., 1981; Becher et al., 2012; Palanca et al., 2013). These papers show the essential role yeasts play for the attractiveness of fruit baits and fermenting substrates (see also: Walsh et al., 2011; Hamby et al., 2012; Kleiber et al., 2014).
Hanseniaspora uvarum (= Kloeckera apiculata) is among the most common yeast species isolated from Drosophila crops in different parts of the world (828/2222 yeast OTUs of 15 Drosophila populations reported by Chandler, Eisen & Kopp (2012)). Its prevalence is about 50% in association with species of D. melanogaster group in North America (78/163 reported by Camargo & Phaff (1957); and 173/344 by Chandler, Eisen & Kopp (2012)). In the Neotropical region, its prevalence associated with D. willistoni from the Amazon is close to 50% (85/174 isolates, see Morais et al., 1995) and almost 40% with D. willistoni populations from the Atlantic Rainforest (146/394 isolates, see Da Cunha, Shehata & De Oliveira, 1957). Additionally, banana baits seeded with H. uvarum have been used since the early fifties in attractiveness experiments due to its easy growth on bananas and high attractiveness of Drosophila specimens (Da Cunha, Dobzhansky & Sokoloff, 1951).
Despite evidences of different substrates attracting distinct assortments of resident Drosophila (Dobzhansky & Pavan, 1950; Del Pino et al., 2015), mashed banana fermented with Saccharomyces cerevisiae has been used since the dawn of Drosophila research to collect and to raise flies (Loeb & Northrop, 1916; Dobzhansky, 1936; Reed, 1938; and others; see Spencer, 1950 for a review of the early use of banana baits seeded with baker’s yeast). This kind of baits allows a broad sampling with almost all resident Drosophila species (see Da Cunha, Dobzhansky & Sokoloff, 1951). Therefore, it has been viewed as a control treatment concerning bait attractiveness in the field or an all-purpose bait.
Previous studies have examined Drosophila attractivity to baits seeded with H. uvarum and S. cerevisiae in the field (Da Cunha, Dobzhansky & Sokoloff, 1951; Da Cunha, Shehata & De Oliveira, 1957; Klaczko, Powell & Taylor, 1983). Pronounced differences concerning the abundance of Drosophila species collected over baits with these yeasts are described. However, due to technical complexity in their experimental design (for example, the use of various baits with different yeast species simultaneously), few explicit generalizations could be set when comparing the attractiveness of these yeast species.
The biodiversity of Drosophila in the Neotropical region is rich (Val, Vilela & Marques, 1981), especially in the Atlantic Rainforest biome, where half of the species remains to be described (Medeiros & Klaczko, 2004). Furthermore, phylogenetic (Yotoko et al., 2003; Hatadani et al., 2009; Izumitani et al., 2016) and morphological (Throckmorton, 1975) differences among Drosophila species groups are so great that they may be considered—and have been used as—a valid taxonomic classification for characterizing patterns of abundance and distribution in especially rich environments (Dobzhansky & Pavan, 1950; Dobzhansky & Da Cunha, 1955).
Since the late 1950s (Da Cunha, Shehata & De Oliveira, 1957), no experiments examining Drosophila differential attractivity in the Neotropical region were carried out. Moreover, due to climate change (Lemes, Melo & Loyola, 2014) and forest fragmentation (Ribeiro et al., 2009) environmental conditions in the Atlantic Rainforest biome are becoming more heterogeneous, with pronounced shifts for local fauna (Batista, Ananina & Klaczko, 2012; Batista & Klaczko, 2013) and flora (Carvalho, Braga & Nascimento, 2016). Furthermore, new occurrence of invasive drosophilid species, such as Zaprionus indianus (see Vilela, 1999) and D. suzukii (see Deprá et al., 2014; Vilela & Mori, 2014), have probably affected ecological interactions among taxa from this biome.
Our objective is to characterize preference (breeding and feeding) behavior that mediates the interaction in the wild among Neotropical Drosophila species and yeasts naturally associated with them. Previous studies have repeatedly shown differences of Drosophila attractivity to baits seeded with H. uvarum and S. cerevisiae. However, so far no clear generalization has been made for the attractivity in the wild. Thus, as a first step in this endeavor, we want to assess the preferences of Drosophila species (groups), from a Neotropical forest fragment, to baits inoculated with either of two yeast species: H. uvarum, one of the yeast species most commonly associated with Drosophila; and S. cerevisiae, the most commonly used yeast species for collecting these flies. Our working hypothesis is that these two yeast species attract sets of flies with different proportions of Drosophila species.
Material and Methods
The Drosophila attraction experiment was carried out within a forest fragment of the Atlantic Rainforest located at Itatiba, SP, Brazil (23°00.07′S, 46°52.917′W; altitude: 740 m) on October 22, 2014 (Permanent Field Permit for Collecting Zoological Material from IBAMA, ICMBio, Ministério do Meio Ambiente—MMA, number: 17238-1). This forest fragment is located 88 km northern Serra da Cantareira, SP, Brazil where Da Cunha, Shehata & De Oliveira (1957) carried out their experiments. Floristic and climatologic properties of both localities are similar, since they belong to the same orogenic formation—Serra da Mantiqueira (Ross, 2013).
We started our experiment around 06h30 a.m., when we randomly exposed baits in the field, and swept entomological nets over baits every 15 min until noon. Then, between 04h00 p.m. and 06h00 p.m., the same procedure was repeated. This strategy was adopted, to minimize possible effects of aggregation behavior and daily temperature variation.
Two different yeast species (commercial S. cerevisiae and H. uvarum—strain ACL-35D; deposited under code UFMG-CM-Y4001 in the Collection of Microorganisms and Cells of the Federal University of Minas Gerais, Brazil) were cultured for 48 h in 200 ml GYMP broth (2% glucose, 0.5% yeast extract, 1% malt extract and 0.2% sodium phosphate monobasic monohydrate). This procedure usually in our laboratory produces suspensions with concentration of 107–108 cells/ml. Then, 200 ml yeast suspensions were poured and stirred with a sterile spoon over approximately 1.44 kg of autoclaved mashed banana; which were left to ferment for about 20 h. On October 22, 2014, temperature varied between 16.5°C and 30.5°C, and the average daily temperature was 23.5°C (see https://www.agritempo.gov.br/agritempo/index.jsp?lang=en, meteorological station CEPAGRI—Campinas, SP). We used field proceedings similar to those described by Da Cunha, Shehata & De Oliveira (1957), when they used mashed banana seeded with H. uvarum and S. cerevisiae with positive yeast growth confirmed by the fermentation of the banana bait and a noticeable bouquet.
Three kinds of banana baits (non-inoculated autoclaved banana and autoclaved banana inoculated with each of the two yeast species) were randomly placed, in order to avoid bias related to position effects, at the edges of an equilateral triangle inscribed in a circle of about 3.5 m of diameter. Each set was distant 10 m from the next set. We collected over five sets of three baits with a total of 15 baits. We collected drosophilids over each type of baits separately, stored them in separate vials, and brought them alive to the laboratory to be analyzed.
Flies were sorted to five groups (see Table 1): tripunctata (D. tripunctata species group); guarani (D. guarani species group); willistoni (D. willistoni species group); exotic (D. immigrans, D. melanogaster species group and Zaprionus indianus); and the remaining flies pooled in others (D. calloptera, D. cardini species group and other non-identified drosophilids). We used Drosophila species group identification as proposed by Freire-Maia & Pavan (1949).
| Group | Control | H. uvarum | S.cerevisiae | Total | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Species | ♀ | ♂ | ∑ | ♀ | ♂ | ∑ | ♀ | ♂ | ∑ | |
| Tripunctata group | ||||||||||
| D. bandeirantorum | 0 | 0 | 0 | 0 | 3 | 3 | 0 | 1 | 1 | 4 |
| D. bifilum | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 |
| D. cuaso | 1 | 1 | 2 | 0 | 0 | 0 | 0 | 1 | 1 | 3 |
| D. fragilis | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 2 | 2 | 3 |
| D. mediopunctata | 1 | 2 | 3 | 2 | 2 | 4 | 4 | 13 | 17 | 24 |
| D. paraguayensis | 0 | 0 | 0 | 5 | 7 | 12 | 13 | 38 | 51 | 63 |
| D. paramediostriata | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 1 |
| D. nappae | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 3 | 3 |
| D. trifilum | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 2 | 2 |
| Non-identified | 1 | 0 | 1 | 6 | 1 | 7 | 15 | 0 | 15 | 23 |
| Group total | 6 | 28 | 93 | 127 | ||||||
| Guaranigroup | ||||||||||
| D. griseolineata | 0 | 2 | 2 | 5 | 19 | 24 | 8 | 32 | 40 | 66 |
| D. maculifrons | 0 | 0 | 0 | 1 | 0 | 1 | 3 | 7 | 10 | 11 |
| Non-identified | 0 | 0 | 0 | 4 | 0 | 4 | 2 | 0 | 2 | 6 |
| Group total | 2 | 29 | 52 | 83 | ||||||
| Willistonigroup | ||||||||||
| D. nebulosa | 0 | 0 | 0 | 11 | 2 | 13 | 2 | 0 | 2 | 15 |
| D. willistoni | 2 | 0 | 2 | 19 | 16 | 35 | 3 | 2 | 5 | 42 |
| Group total | 2 | 48 | 7 | 57 | ||||||
| Exoticspecies | ||||||||||
| D. immigrans | 0 | 0 | 0 | 3 | 1 | 4 | 0 | 0 | 0 | 4 |
| D. melanogaster | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 1 |
| D. suzukii | 0 | 0 | 0 | 0 | 6 | 6 | 1 | 1 | 2 | 8 |
| D. simulans | 0 | 1 | 1 | 5 | 8 | 13 | 3 | 1 | 4 | 18 |
| Zaprionus indianus | 0 | 0 | 0 | 2 | 3 | 5 | 0 | 0 | 0 | 5 |
| Group total | 1 | 29 | 6 | 36 | ||||||
| Others | ||||||||||
| D. atrata | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 2 | 2 |
| D. polymorpha | 0 | 0 | 0 | 1 | 2 | 3 | 1 | 0 | 1 | 4 |
| Drosophilids | 0 | 0 | 0 | 4 | 0 | 4 | 0 | 0 | 0 | 4 |
| Group total | 0 | 7 | 3 | 10 | ||||||
| Total | 11 | 141 | 161 | 313 | ||||||
Wild male flies were identified to species level by dissecting their genitalia; and for collected females, the genitalia of their laboratory reared F1 males were analyzed. The specimen genitalia of tripunctata group flies was compared to drawings reported by: Frota-Pessoa (1954), Val (1982), Vilela & Pereira (1985), Vilela & Pereira (1986), Bächli, Vilela & Ratcov (2000) and Vilela, Valente & Basso-da-Silva (2004). Species of calloptera, cardini and guarani groups were compared to drawings reported by: Val (1982) and Vilela & Bächli (1990). Specimens that belong to melanogaster and willistoni groups were compared to drawings reported by: Salles (1948) and Malogolowkin (1952). We used external morphology for classification of D. immigrans, D. suzukii and Z. indianus specimens. When the genitalia were lost, flies were identified by external morphology and labeled as non-identified in the respective group.
Breeding and egg-laying preferences may motivate choice behavior of Drosophila females. Therefore, bias in sex ratio would be expected over a particular bait, if females would choose that substrate for oviposition. We performed a chi-square test for characterizing differences in sex ratio between baits. Since no bias in group sex ratio was found (see Table 2), we analyzed the sum of females and males collected over baits. After that, we compared the attractiveness of each set of baits testing the observed numbers of flies within each group collected over S. cerevisiae and H. uvarum against an expected 1:1 proportion with a chi-square.
| Groups | X2 | d.f. | p-value |
|---|---|---|---|
| Tripunctata | 2.22 | 1 | >0.1ns |
| Guarani | 0.82 | 1 | >0.3ns |
| Willistoni | 0.21 | 1 | >0.7ns |
| Exotic | 2.15 | 1 | >0.3ns |
Notes:
- X2
-
result of chi-square tests
- d.f.
-
degree of freedom
- p-value
-
associated probability
- ns
-
non-significant
Results
Table 1 shows the 313 specimens of drosophilids collected. Uninoculated banana baits (controls) attracted virtually no flies (11 versus 302 in the other ones; less than 4% of the total). Thus, the results of flies from uninoculated banana baits were no further analyzed.
We collected 141 flies (47% of the 302 flies attacted to yeast inoculated baits) over baits with H. uvarum and 161 (53% of the total 302) over baits with S. cerevisiae. Species with largest numbers among the 20 species identified were Drosophila griseolineata (n = 66), D. paraguayensis (n = 63) and D. willistoni (n = 42). The D. tripunctata group was the most diverse with ten species, followed by D. melanogaster group with three species (pooled as exotic in Table 1). Although no significant difference was detected between total number of flies collected over baits seeded with the two diffent yeasts (X2 = 0.66; d.f. = 1; p > 0.30), the composition of flies was clearly different (Fig. 1).

Figure 1: Drosophilids attracted to yeast-inoculated baits.
Percentage of Drosophilidae groups (in green tripunctata; in yellow guarani; in red willistoni; in purple exotic; and in blue others) collected over baits inoculated with Hanseniaspora uvarum (A) and with Saccharomyces cerevisiae (B).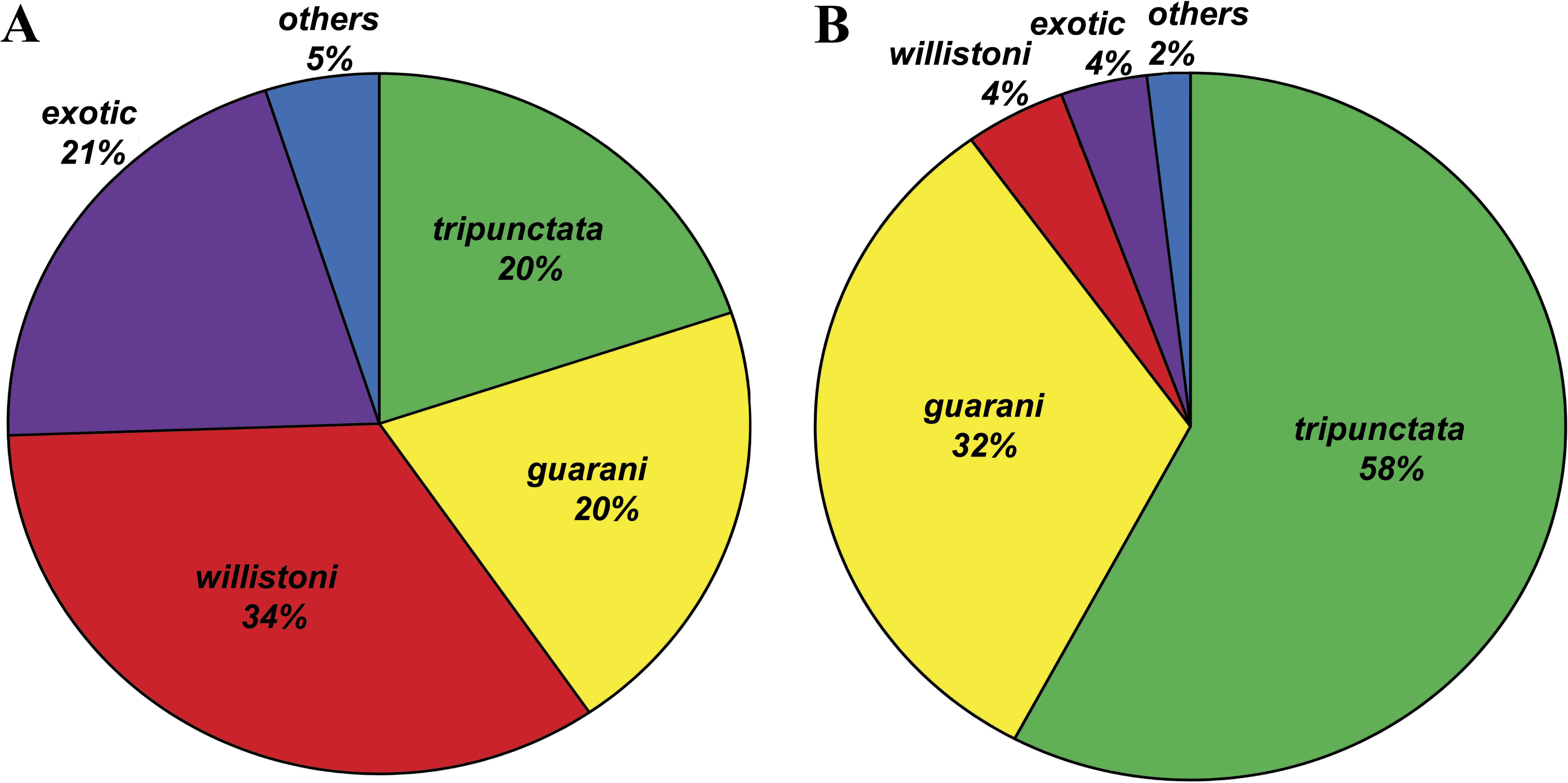{kind=link}
Flies of the tripunctata and guarani groups (subgenus Drosophila) showed similar pattern and were significantly more attracted to baits inoculated with S. cerevisiae (93 in 121 = 77%, X2 = 34.9, d.f. = 1, p < 0.001; and 52 in 81 = 64%, X2 = 6.5, p < 0.01) than to H. uvarum (23% and 36%, respectively). However, flies of willistoni (7 in 55 = 13%; X2 = 30.6, d.f. = 1, p < 0.001), exotic (6 in 35 = 17%; X2 = 15.1, p < 0.01) and other (3 in 10 = 30%; X2 = 1.6, p > 0.2—non-significant) groups were less collected over S. cerevisiae than over H. uvarum (87%, 83%, and 70%, respectively). After Bonferroni multiple tests correction, all test significance results remain qualitatively unchanged.
Discussion
Parts of plants or fungi with a particular microbiota are substrates used by several Drosophila species for feeding, mating, oviposition and breeding (Powell, 1997). Although fresh fruits are attractive for D. suzukii (see Keesey, Knaden & Hansson, 2015), most Drosophila species are attracted to decayed fruits. Furthermore, variation in microbiota density associated to its decaying age (in number of days) affects the attraction and abundance of several cosmopolitan species, such as D. melanogaster, D. simulans and D. immigrans, to decayed oranges (Nunney, 1996).
The low attractiveness of baits with non-fermented banana reinforces the fact of the presence of yeast are necessary for baits attractiveness (Klaczko, Powell & Taylor, 1983). Although no direct count of yeast colonies was carried out in the banana baits, the obvious banana fermentation (increased volume and typical bouquet) were compeling evidences of yeast growth. Additionally, we collected approximately the same magnitude of specimens over banana baits with each of the two yeasts, indicating both yeasts grew and the species composition differences could be attributed to differences in odor profiles between H. uvarum and S. cerevisiae (see Scheidler et al., 2015).
We collected more flies of subgenus Sophophora such as D. melanogaster and D. suzukii over baits with H. uvarum (68 in a total of 81 = 84%) than over S. cerevisiae (13/81 = 16%), while flies of the tripunctata group (subgenus Drosophila) were more attracted to baits inoculated with S. cerevisiae (93 in 121 = 77%) than to H. uvarum (23%).
Species that belong to subgenus Sophophora, such as D. melanogaster and flies of D. obscura group, showed preferences for baits inoculated with apiculate yeast H. uvarum over other yeasts, such as S. cerevisiae in laboratory populations (Hoang, Kopp & Chandler, 2015) and natural populations (Da Cunha, Dobzhansky & Sokoloff, 1951; Klaczko, Powell & Taylor, 1983). However, other species from subgenus Drosophila, such as D. occidentalis were more collected over baits with S. cerevisiae than over baits with apiculate yeasts in San Jacinto Mountains, CA, USA (Klaczko, Powell & Taylor, 1983).
In the tropical region, Dobzhansky & Da Cunha (1955) and Da Cunha, Shehata & De Oliveira (1957) carried out experiments of differential attractiveness in the Amazon (Belém, PA, Brazil and Tapajós, PA, Brazil) and in the Atlantic Rainforest (Rio Doce, MG, Brazil and Serra da Cantareira, SP, Brazil). These authors observed that flies of Sophophora subgenus, such as D. willistoni, D. nebulosa, and D. simulans, were more collected over baits with H. uvarum than over baits with Candida krusei (=Pichia kudriavzevii) and S. cerevisiae or close relatives. However, H. uvarum baits were poorly attractive to some species from subgenus Drosophila such as D. calloptera, D. guaramunu and flies from D. tripunctata group, which were collected over baits with yeast from genera Candida, Pichia and Saccharomyces.
Choice behavior may be triggered by females that are choosing oviposition sites. If so, it is expected to collect more females over one kind of bait. However, no differences between female and male collected over baits were observed (see Table 2). Furthermore, only nine isofemales out of 69 (five collected over H. uvarum and four over S. cerevisiae) did not produced any progeny, so we have not much evidence of oviposition choice with this sample. Finally, further experiments evaluating differences in the choice behavior between females virgin and non virgin in the field as well as the relationship between yeast preference and components of biological fitness will be carried out. Moreover, the experimental design we used cannot rule out conspecific attraction (Lihoreau et al., 2016).
H. uvarum and S. cerevisiae have been already isolated from fruits and tree bark, respectively, in Amazonian and Atlantic Rainforests (Morais et al., 1995; Pimenta et al., 2009; Barbosa et al., 2016) as well as from crops of several Drosophila species (see Da Cunha, Shehata & De Oliveira, 1957; Morais et al., 1992; Morais, Pagnocca & Rosa, 2006; Batista et al., 2016). Species such as D. paraguayensis (see Batista et al., 2016) and D. maculifrons (see Da Cunha, Shehata & De Oliveira, 1957), which belong to D. tripunctata and D. guarani groups respectively, both of the Drosophila subgenus, had S. cerevisiae isolated from their crops. In contrast, H. uvarum group was the most prevalent yeast isolated from crops of Sophophora subgenus species such as: D. willistoni (see Da Cunha, Shehata & De Oliveira, 1957); D. melanogaster group (see Camargo & Phaff, 1957; Morais et al., 1995; Chandler, Eisen & Kopp, 2012); and D. suzukii (see Hamby et al., 2012).
Several evidences suggest the natural association between yeasts and Drosophila in the wild. Pimenta et al. (2009) states that Drosophila may be a major vector of yeasts in Atlantic Rainforest. Our data shows that species of D. tripunctata and D. guarani groups are preferentially attracted to S. cerevisiae, reflecting their natural association in the wild; while species of subgenus Sophophora such as D. melanogaster, which is preferentially attracted to baits with H. uvarum may be naturally associated with apiculate yeasts. Therefore, differences in dispersion and distribuition of the yeast species might be related to the variation observed for Drosophila species.
It is noteworthy that our results are consistent with those obtained more than half a century ago by researchers such as Da Cunha even if working with different objectives (see above) suggesting that the preferences among Drosophila species group found may be reflecting deep and stable relations with yeast species in natural forests in spite of all the environmental changes that have occurred. These results represent a first step to understand differences in feeding preferences among Drosophila species and their consequences for biological fitness. Naturally, additional studies characterizing yeast species associated with Drosophila species in natural remnants of Atlantic Rainforest, as well as the differences between male and female behavior, different physiological states, and on the molecular basis of Drosophila species olfactory system may further our understanding of the associations we now report.
Supplemental Information
Raw data
List of drosophilids (females—sheet 1 and males—sheet 2) collected over uninoculated banana baits and banana baits with Hanseniaspora uvarum and Saccharomyces cerevisiae.