
Opportunities drive the global distribution of protected areas
- Published
- Accepted
- Received
- Academic Editor
- Patricia Gandini
- Subject Areas
- Biogeography, Conservation Biology, Ecology, Natural Resource Management
- Keywords
- Protected areas, National parks, Conservation paradigms, Representativeness, Opportunity, Preferentiality
- Copyright
- © 2017 Baldi et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2017. Opportunities drive the global distribution of protected areas. PeerJ 5:e2989 https://doi.org/10.7717/peerj.2989
Abstract
Background
Protected areas, regarded today as a cornerstone of nature conservation, result from an array of multiple motivations and opportunities. We explored at global and regional levels the current distribution of protected areas along biophysical, human, and biological gradients, and assessed to what extent protection has pursued (i) a balanced representation of biophysical environments, (ii) a set of preferred conditions (biological, spiritual, economic, or geopolitical), or (iii) existing opportunities for conservation regardless of any representation or preference criteria.
Methods
We used histograms to describe the distribution of terrestrial protected areas along biophysical, human, and biological independent gradients and linear and non-linear regression and correlation analyses to describe the sign, shape, and strength of the relationships. We used a random forest analysis to rank the importance of different variables related to conservation preferences and opportunity drivers, and an evenness metric to quantify representativeness.
Results
We find that protection at a global level is primarily driven by the opportunities provided by isolation and a low population density (variable importance = 34.6 and 19.9, respectively). Preferences play a secondary role, with a bias towards tourism attractiveness and proximity to international borders (variable importance = 12.7 and 3.4, respectively). Opportunities shape protection strongly in “North America & Australia–NZ” and “Latin America & Caribbean,” while the importance of the representativeness of biophysical environments is higher in “Sub-Saharan Africa” (1.3 times the average of other regions).
Discussion
Environmental representativeness and biodiversity protection are top priorities in land conservation agendas. However, our results suggest that they have been minor players driving current protection at both global and regional levels. Attempts to increase their relevance will necessarily have to recognize the predominant opportunistic nature that the establishment of protected areas has had until present times.
Introduction
Historically and throughout the world, societies have set aside land from its conventional uses in order to protect particular natural or cultural values (McNeely, Harrison & Dingwall, 1994). In this way, hilltops, old-growth forests, or seashores maintained their biodiversity, scenic attributes, or provision of ecological services. In the last century, simultaneously with the rising pressures over land resources (Vitousek et al., 1997; Ellis et al., 2013), protected areas have greatly increased in number and total area. From just a small handful of locations at the end of the 19th century to thousands nowadays, protection encompasses 15.4% of the world’s continental surface (1.4 × 108 km2), excluding Antarctica (Fig. 1) (Juffe-Bignoli et al., 2014).
The current distribution of protected areas responds to a deliberate process guided by a complex interplay of motivations related to perceived societal benefits (McNeely, Harrison & Dingwall, 1994; Pressey, 1994; Margules & Pressey, 2000; Watson et al., 2014). The strength of different motivations changed through history and across territories (Wirth, 1962; Sellars, 1997; Erize, 2003; Mace, 2014). Many of the protected areas established in the late 19th and early 20th centuries responded to practical interests such as favoring tourism or preserving iconic landscape features. However, since the second half of the 20th century, protection has been influenced by a widespread agreement on the importance of maintaining nature in general and biodiversity in particular. Therefore, part of the present-day expansion of protected areas aims to include areas of high species richness, endemism hotspots, or underrepresented ecological or biophysical conditions. Ultimately, we classify these motivations as preferential or representative. The former corresponds to the preservation of specific biological, spiritual, economic, or geopolitical values offered by some territory. The latter corresponds to the protection of a balanced sample of the multiple biophysical environments hosted by a territory, a country, or the whole globe (Pressey, 1994; Lovejoy, 2006) (Table 1). These two groups of motivations interact with different opportunistic forces that shape conservation, as protected areas are frequently deployed in areas that face little human interventions and have comparatively low opportunity-costs, at least at the time of their establishment (Joppa & Pfaff, 2009; Aycrigg et al., 2013; Durán et al., 2013). Consequently, protection has been biased towards unproductive or isolated areas (e.g., cold, dry, with poor soils), leaving other territories inadequately protected despite their potential conservation value (e.g., temperate, subhumid areas) (Pressey, 1994; McNeely & Schutyser, 2003; Hoekstra et al., 2005; Joppa & Pfaff, 2009).
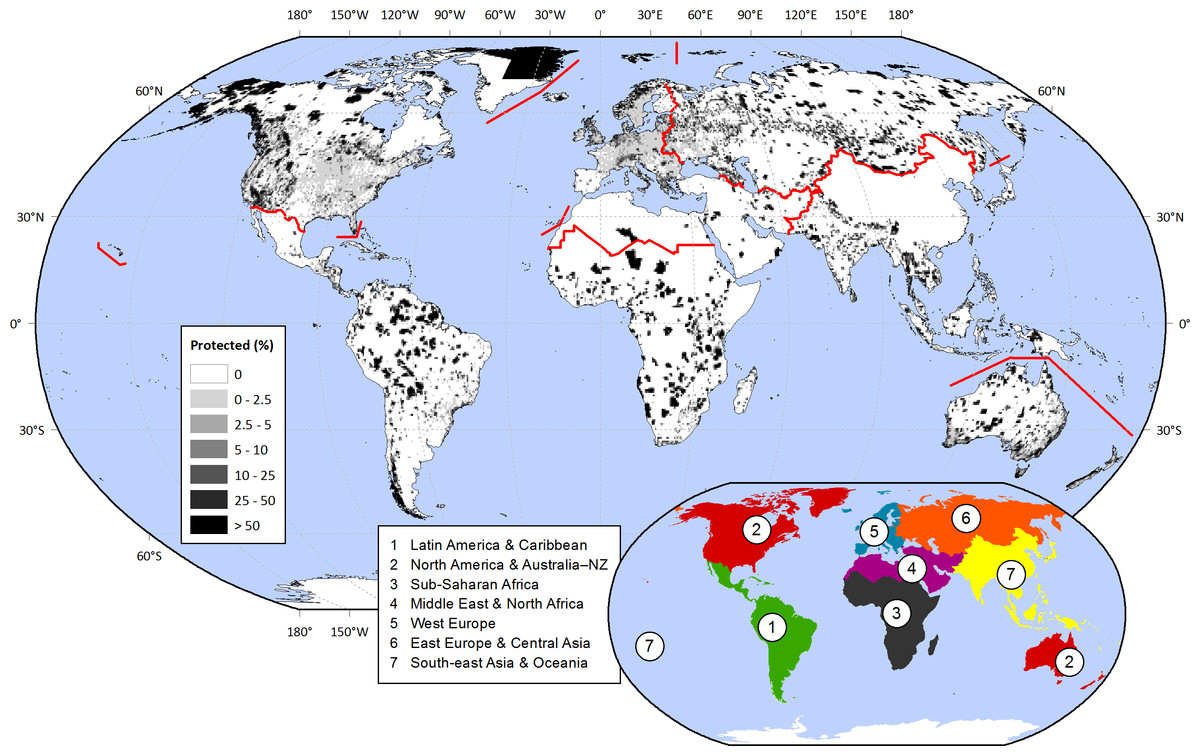
Figure 1: Protected areas fraction on a 0.5° × 0.5° cell basis.
IUCN & UNEP-WCMC (2013) data was summarized within 0.5° × 0.5° contiguous cells, considering IUCN categories I–IV (1994). The regions under analysis are depicted in the inset map, and with red lines in the main map. Regional protected fractions are shown in Table S2.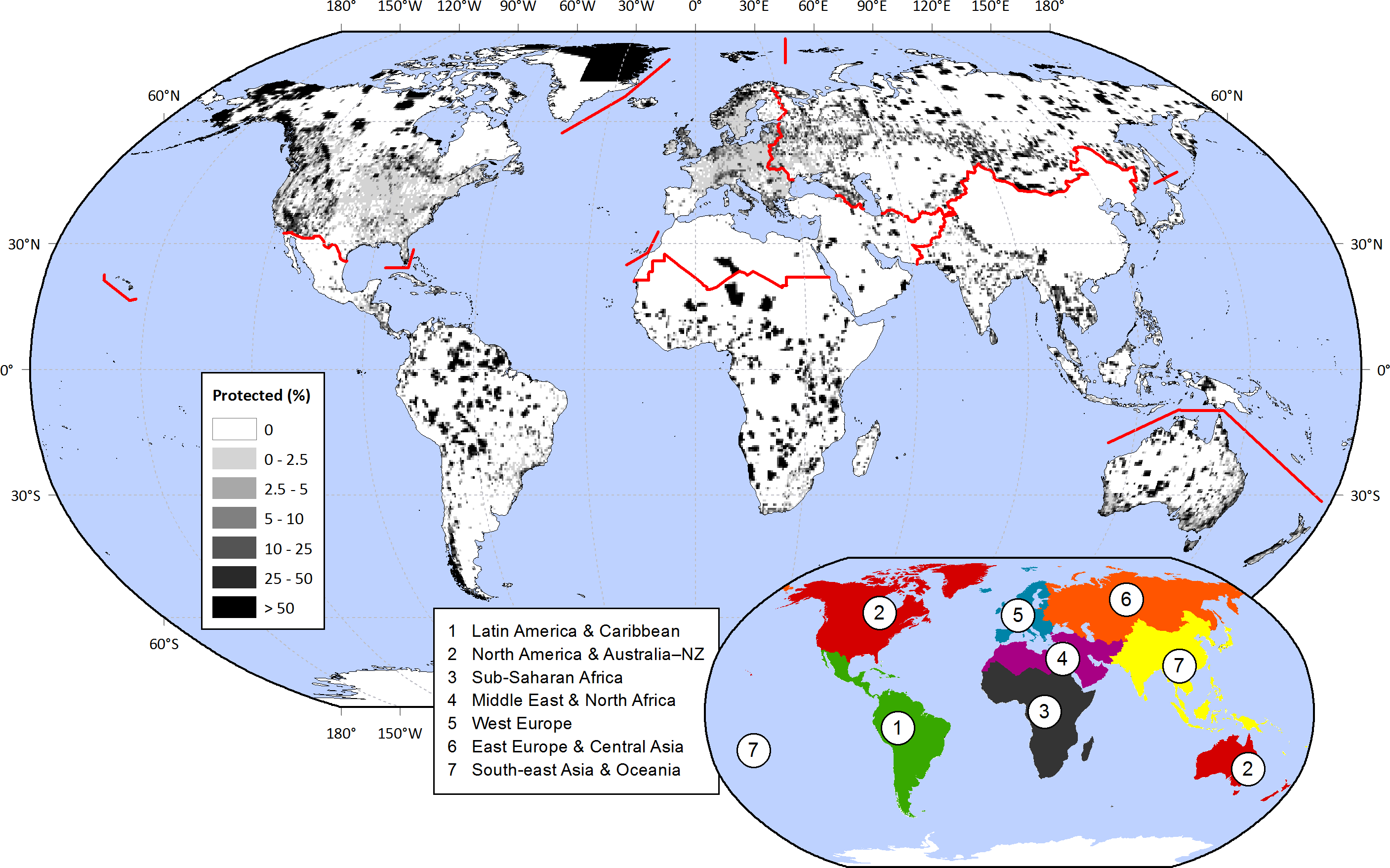{kind=link}
Most research about the spatial distribution of protected areas has focused on evaluating the effectiveness of existing networks to encompass biodiversity (Scott, 1993; Brooks et al., 2004; Rodrigues et al., 2004a; Rodrigues et al., 2004b) and biogeographical (ecoregions, biomes, realms) (McNeely, Harrison & Dingwall, 1994; Jenkins & Joppa, 2009; Barr et al., 2011; Watson et al., 2014) or anthropogenical units (Martin et al., 2014). However, few studies have addressed the relative importance that different forces may have had on the deployment of protected areas (Joppa & Pfaff, 2009). Here, we characterized the current distribution of terrestrial protected areas explicitly designated for nature protection—i.e., categorized as I–IV under IUCN guidelines (1994)—in relation to biophysical, human, and biological variables (Table 2). By associating these variables to representative motivations, preferential motivations, and opportunistic forces (Table 1), we assessed the relative impact of these drivers at regional and global levels. While motivations and opportunistic forces are likely to coexist, the predominance of any of them should result in a singular spatial pattern of land protection: (i) If representative motivations prevail, two alternative patterns can be expected, depending on whether protection targets a uniform fraction or on a uniform absolute area of biophysical environments. A uniform fraction leads to a prevalence of the most abundant environments (hereafter, “fraction representativeness”) (Fig. 2A). Alternatively, a uniform absolute area leads to a balanced contribution of common and rare environments (hereafter, “quota representativeness”) (Fig. 2B); (ii) If preferential motivations prevail, protection should be geographically biased towards areas with high biological, spiritual, economic, or geopolitical values (e.g., species diversity, frontiers) (Fig. 2C); (iii) Finally, biases would also arise if opportunistic forces prevail, with protected areas having greater chances of being established where productive potential (e.g., agriculture) and/or human presence are low (Fig. 2C). Our analyses included linear and non-linear regressions, correlations, random forests, and evenness metrics, taking advantage of available spatial datasets.
| Group and name | Origin | Description | Examples |
|---|---|---|---|
| Preferential motivations | |||
| Cultural and spiritual | Anthropocentric and non-utilitarian. Early formation of unified societies (e.g., feudal). | Protection is established on remarkable natural and/or cultural sceneries, as their aesthetic appreciation—through direct contact—ensures the fulfillment of basic human needs and thus the well-being of individuals and societies (Bhagwat & Rutte, 2006; Loreau, 2014). | Current Uluru-Kata Tjuta NP (Australia, 1958), Forêt de Fontainebleau (France, 1861) |
| Gaming and wildlife managing | Anthropocentric and utilitarian. Early. | Protection limits hunting wildlife with the aim of maintaining healthy animal populations (especially ‘singular’ species) and—in the case of gaming—providing recreation to a restricted part of society (Szafer, 1973). | Białowieża Forest (Poland/Belarus, <1541), Pongola Game Reserve (South Africa, 1894) |
| National imaginary | Anthropocentric and non-utilitarian. Consolidation of modern states. | Similar to the Cultural and spiritual motivation, but with a planned governmental aim of shaping a national pride and identity through natural or cultural icons (Paül Carril, Santos Solla & Pazos Otón, 2015). | Iguazú/Iguaçu NP (Argentina/Brazil, 1935/1939) |
| Frontier protection and peace preservation | Anthropocentric and utilitarian. Post-independence. | Protection is established close to international borders, as these can be conceived as areas where assert sovereignty or as neutral zones fostering or dedicated to cooperative and peaceful economic activities (Zbicz & Green, 1997). | Waterton-Glacier International Peace Park (Canada/USA, 1932) |
| Ecosystem goods and services provision | Anthropocentric and utilitarian. Early, but the service concept was popularized since 1900. | Protection is established on territories able to supply over time critical environmental goods and services (timber, water, pollination, soil protection, carbon sequestration) (Costanza et al., 1997). | Malleco National Reserve (Chile, 1907) |
| Tourism, leisure and recreation | Anthropocentric and utilitarian. Beginning of the 20th century. | Similar to the cultural or spiritual motivation, but with the aim of providing popular entertainment and enjoyment and bringing significant economic benefits to local to regional economies (McKercher, 1996; Mulholland & Eagles, 2002). Could be considered a particular ecosystem service. | Abel Tasman NP (New Zealand, 1942), Nikkō NP (Japan, 1934) |
| Biological conservation | Biocentric and non-utilitarian. Beginning of the 20th century, but actively after 1960. | Protection is established on territories of high species richness, rates of endemism, or of unique species assemblies (Myers et al., 2000; Terborgh & Winter, 1983). | Virunga NP (Congo DR, 1925), Komodo NP (Indonesia, 1980) |
| Representative motivations | |||
| Fraction | Idem biological conservation | Protection is focused on the representation of ecosystems (biota and processes) due to their intrinsic values (Kareiva & Marvier, 2003; Pressey, 1994), or as pristine scenarios where knowledge of the Earth system can be improved (Bourlière, 1962). Under this motivation, protection targets a uniform fraction of the biophysical environments of a given territory (McNeely, Harrison & Dingwall, 1994; SCBD, 2010), assuming a close relationship between biophysical and ecosystem diversities (Belbin, 1993; Holdridge, 1947). | |
| Quota | Idem biological conservation | Idem fraction representativeness, but protection targets a uniform absolute area of biophysical environments. | |
| Opportunistic forces | |||
| Anthropocentric. Beginning of the 20th century. | Protection is established on where opportunity exists, mostly where it is economically feasible, i.e., territories that have a low economic value for traditional and profitable land uses (Margules & Pressey, 2000). | Northeast Greenland NP (Denmark, 1974) | |
Methods
Data sources
The location of protected areas was obtained from the “World Database on Protected Areas”, Annual Release 2013 (IUCN & UNEP-WCMC, 2013). We considered only terrestrial areas explicitly designated for nature protection, i.e., strict nature reserves, wilderness areas, national parks, natural monuments or features, and habitat/species management areas—categories I–IV (IUCN, 1994). We compiled a database of 15 biophysical, human, and biological variables (Table 2). These variables can be directly related to individual motivations and opportunistic forces. For example, the metric “distance to frontiers” can be linked to the preferential motivation of “frontier protection and peace preservation.” We excluded the Antarctica from all analyses.
| Variable | Calculation and source | Summarizing method | Group and name |
|---|---|---|---|
| Temperature | Mean annual values in °C, from the “Ten Minute Climatology data base” (New et al., 2002), representing averaged monthly figures for the 1961–1990 period. | Mean | Representativeness motivations (fraction and quota) |
| Precipitation | Amount of annual precipitation in mm. Same source as temperature | ||
| Precipitation to potential evapotranspiration ratio (PPT:PET) | Mean annual values describing water availability (unitless). Same source as temperature. Potential evapotranspiration is retrieved from the Penman-Monteith equation (Allen et al., 2004) and calculated on a monthly basis. | ||
| Elevation | From “Shuttle Radar Topography Mission” (SRTM) digital elevation model (USGS, 2004). Spatial resolution: 90 m. In m above sea level. | ||
| Terrain slope | From “Shuttle Radar Topography Mission” (SRTM) digital elevation model (USGS, 2004). In degrees. | ||
| Soil fertility | Represented by top-soil total exchangeable bases (TEB, 0–30 cm), in cmolc * kg−1. From ISRIC-WISE—Global data set of derived soil properties (v.3.0) (Batjes, 2006). Spatial resolution: 30 arc-min. | ||
| Tourism attractiveness | “Panoramio” photos (http://www.panoramio.com) to population counts ratio, in photos * inh−1. Modified from the “World touristiness map” (http://www.bluemoon.ee). Panoramio photos were downloaded in December 2013 and processed with Python v.2.7. Population came from the same source referred previously. | Preferential motivations: Cultural and spiritual; National imaginary; Tourism, leisure and recreation | |
| Distance to frontiers | Considering exclusively cells within countries with terrestrial political frontiers. Euclidean distance in km from vector data from “Natural Earth” (http://www.naturalearthdata.com). Cartographic scale: 1:50 m. | Preferential motivations: Frontier protection and peace preservation | |
| Biomass | Biomass carbon stored in above and belowground living vegetation circa 2000 (Ruesch & Gibbs, 2008), in Mg ha−1. Spatial resolution: 1 km. | Maximum, representing attainable conditions | Preferential motivations: Ecosystem goods and services provision |
| Animal richness | Number of breeding bird, amphibian, and mammal species from Jenkins, Pimm & Joppa (2013). Spatial resolution: 10 km. | Mean | Preferential motivations: Biological conservation |
| Vascular plant richness | Number of vascular plant species from Kreft & Jetz (2007) (combined multipredictor model). Spatial resolution: 110 km. | ||
| Population | Inhabitants from the “Gridded Population of the World v.3 (GPWv3): Population Grids” for the years 1990–1995 (CIESIN-CIAT, 2005). Spatial resolution: 2.5 arc-min. | Sum | Opportunistic forces |
| Isolation | From the 2000 map “Travel Time to Major Cities” (Nelson, 2008). Representing the distance to large cities (>50,000 inh) in minutes by using a cost-distance algorithm. Spatial resolution: 0.5 arc-min. | Minimum, representing human context of the surrounds of protected areas | |
| Distance to coasts | Considering ocean coasts. Potentially related to the proximity to docking ports. Euclidean distance in km from vector data from “Natural Earth.” | Mean | |
| Cropland suitability | Land suitability for low input level rain-fed crops, considering cereals, soybean, and oil palm (FAO/IIASA, 2011). Calculated as the maximum suitability of the included species, per pixel (unitless). Spatial resolution: 5 arc-min. |
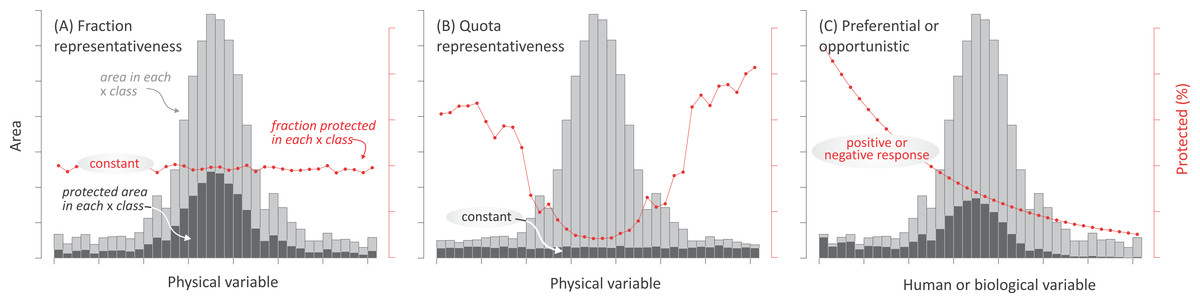
Figure 2: Expected protection patterns according to different forces.
Expected geographic patterns of protected areas according to the three groups of forces. In (A) and (B) “fraction” and “quota” representativeness motivations, in (C) preferential motivation and opportunistic forces. Encircled text refers to the expected and tested behavior. Three measurements are shown in the histograms: the area in each class of the independent variables (light gray bars), the area under protection in each j class (intervals in the histograms) of the independent variable (dark gray bars), and the fraction under protection of the j class of the i independent variable (red dots and lines). Only the last two measures were used in the statistical analyses.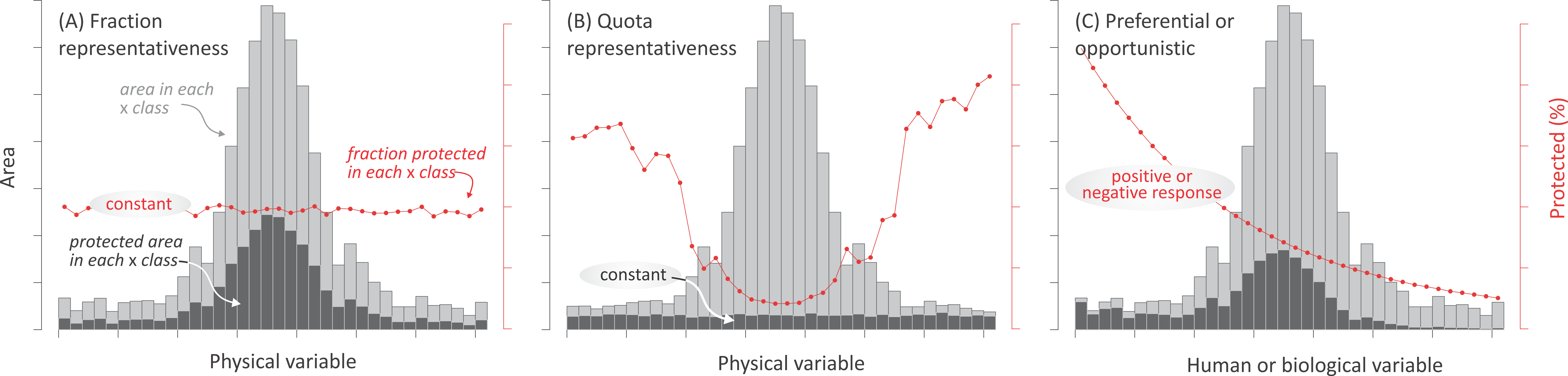{kind=link}
Sampling procedure
We explored the distribution of protected areas at global and regional levels, considering “Latin America & Caribbean,” “North America & Australia–NZ” (New Zealand), “Sub-Saharan Africa,” “Middle East & North Africa,” “West Europe,” “East Europe & Central Asia,” and “South-east Asia & Oceania” (Fig. 1). This regional division relied on cultural, historical, and biogeographical factors (adapted from McNeely, Harrison & Dingwall, 1994; Inglehart & Welzel, 2005; Ellis & Ramankutty, 2008). In order to analyze the links between protection and biophysical, human, and biological conditions (Table 2), we summarized all data into 66,555 cells of 0.5° × 0.5° (Table S1), excluding those with a terrestrial fraction <5%. Compared to other approaches in which each protected area is treated as a single sample, this grid-based approach offered the advantages of (i) providing a unified spatial resolution for all variables, (ii) encompassing the full range of global biophysical, human, and biological conditions, and (iii) avoiding the averaging of these conditions within very large protected areas. Additionally, (iv) this approach provided a clearer representation of the geographical context of protected areas by characterizing the full grid cell in which they are embedded and not just the protected territory (99.55% of the cells incorporates unprotected conditions).
Data analysis
After summarizing all data within grid cells, we generated 120 histograms —i.e., (7 regions + globe) * 15 independent variables—, containing three sets of information: (i) the absolute area under protection in each j class (interval in the histograms) of the i independent variable —AREA.PROT—, (ii) the fraction under protection of the class j of the i independent variable —FRAC.PROT—, and (iii) the area in each class of the i independent variables —AREA—; considering a weighted arithmetic mean according to a maximum cell area within each j class. For each independent variable, we set a particular class width considering data distribution at the global level. In order to avoid long tails in the histograms, lower and upper j classes were grouped using the percentile values 0.025 and 0.975 of the i independent variable. At the regional level, we maintained the width of classes in order to facilitate comparisons. We conducted all statistical analyses with the AREA.PROT and FRAC.PROT information separately, while AREA information was shown only for descriptive purposes. For all tests, we carried out the modeling with the values of ≥8 histogram intervals (if not, we divided histogram classes up to accomplish this rule).
We assessed the reciprocal associations between the i independent variables through a Kendall’s τ non-parametric test (Whittaker, 1987). All calculations were run in RStudio v.0.98.507 (packages Segmented, Scatterplot3d) and Python v.2.7 (packages Scikit-learn, Pandas, Numpy). In order to explore the relative significance of “fraction” and “quota” representativeness motivations (Figs. 2A–2B), we analyzed the existence of a relationship between the FRAC.PROT or AREA.PROT values and the six biophysical variables (Fig. 2) by means of a modification of the “Shannon evenness” () (Hill, 1973), calculated as:
(1) (2) (3)where xij represents the FRAC.PROT and AREA.PROT in the j class of the i independent variables, and n the number of classes on the histogram. Vij is calculated to transform FRAC.PROT and AREA.PROT into probabilities. In , the numerical effects of an uneven number of classes as well as of despair xij magnitudes are canceled. The index ranges between 0 and 1, with a value of 1 when xij is constant along the i gradient.
While the modified Shannon evenness index indicates the presence of a relationship, regression analyses characterize the behavior of a relationship in terms of shape, sign, and eventually multivariate strength. In this sense, we regressed the FRAC.PROT on the i independent variables related to preferential motivations (e.g., animal richness) and opportunistic forces (e.g., cropland suitability) (Fig. 2C). We assessed first and second order polynomials, exponentials, one phase associations, semi-logarithmic (X axis logarithmic, Y linear), and piecewise models (Faraway, 2006), selected models through the Akaike’s information criterion (Akaike, 1974), and calculated a pseudo-R 2 by correlating observed and predicted values from each model as a goodness-of-fit measurement.
We then ranked the relative importance of these variables by means of a random forest algorithm —a machine-learning technique (Breiman, 2001). Random forest estimates the variable importance by looking at how much the mean square error (MSE) increases when the out-of-bag data (observations which are not used for building the current tree, OOB) for that variable are permuted while all others are left unchanged (Liaw & Wiener, 2002). For each unpruned (fully grown) tree, the MSE on the OOB portion of the data is recorded, and then the same is done after permuting each independent variable. Differences between MSE and OOB are averaged over all trees and normalized by their standard deviation. The allocated variable importance can differ substantially with the selection of number of trees to grow (ntree), the minimum size of the terminal nodes (nodesize), or the number of input variables at each split (mtry) (Grömping, 2009; Genuer, Poggi & Tuleau-Malot, 2010). This last parameter has been described as the most critical one; if mtry = 1, the splitting variable would be determined completely randomly; whereas a mtry =p (maximum number of variables) would eliminate the previously described first aspect of randomness, and the possibility of some independent variables—related to the dependent variable but correlated to a stronger regressor—to become the basis of splitting. A usually recommended value on a regression is mtry =p∕3 because a lower correlation between individual trees improves prediction accuracy (Liaw & Wiener, 2002). However, as the mtry values depend on the model and the correlation between independent variables (Breiman, 2001; Grömping, 2009), we set mtry1 values that minimize the OOB-MSE of the model (and a ntree = 500, and a nodesize = 1). The variable importance was used here with an explanatory and interpretative, rather than predictive, aim (Grömping, 2009). We excluded biophysical variables from the random forest since their importance would not reflect the importance of the representativeness motivation, but quite the opposite.

Figure 3: Global distribution of protected areas.
Global distribution of protected areas along biophysical, human, and biological gradients. See graphic explanations in Fig. 2. Variables represented in (A–F) are related to the representativeness motivations, those in (G–K) are related to the preferential motivations, and those in (L–O) are related to the opportunistic forces. Lower and upper j classes were grouped using the percentile values 0.025 and 0.975 of the i independent variable. Blue asterisks denote that histograms were generated with the log10 transformed independent variable, and thus do not correspond to the untransformed data used for statistical analyses. Region-specific histograms are shown in Fig. S2.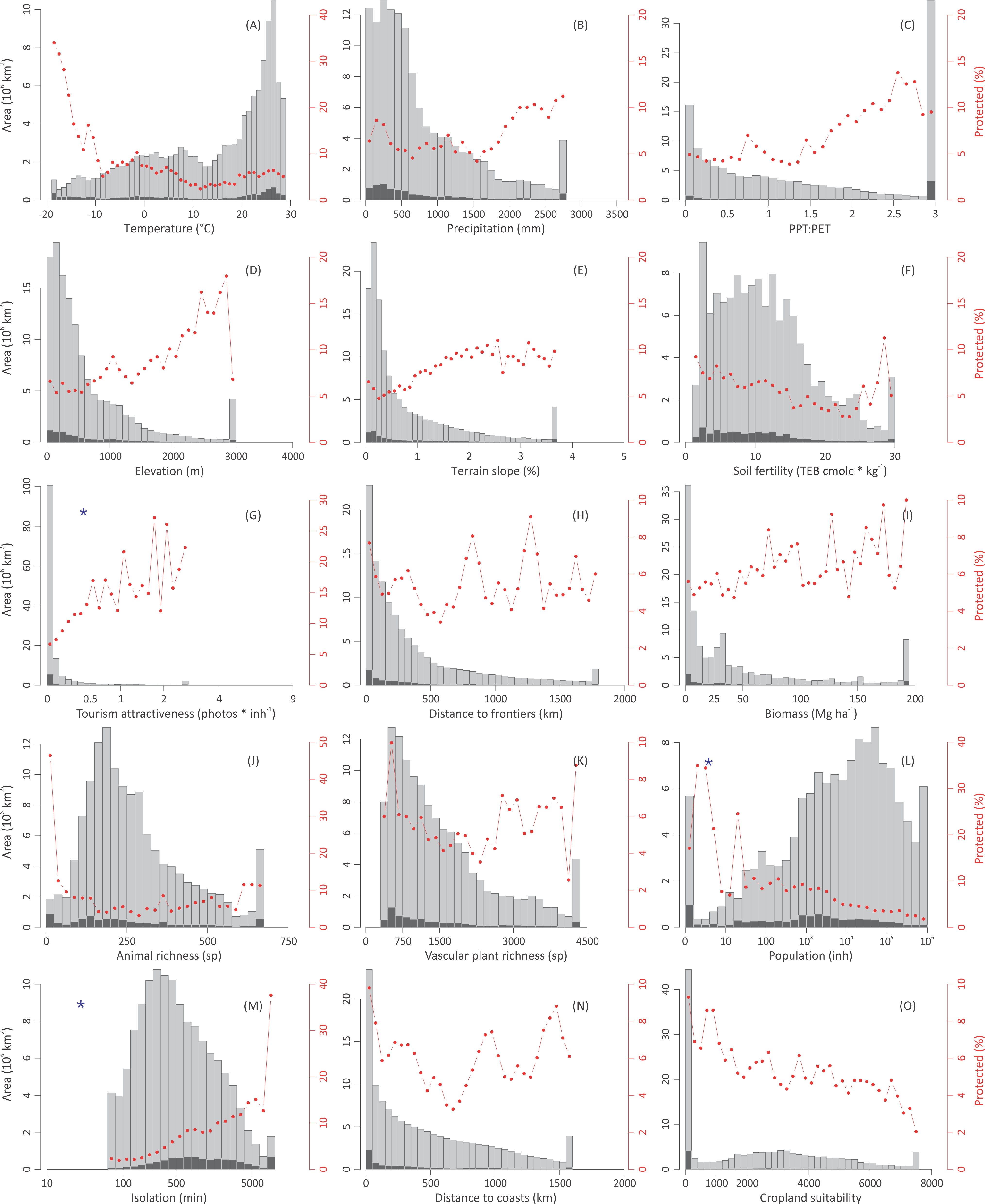{kind=link}
| Global | Latin America & Caribbean | North America & Australia–NZ | Sub-Saharan Africa | Middle East & North Africa | West Europe | East Europe & Central Asia | South-east Asia & Oceania | ||
|---|---|---|---|---|---|---|---|---|---|
| Preferential | Tourism attractiveness | 12.72 | 0.82 | 6.93 | 6.28 | 9.29 | 4.87 | 4.06 | 3.62 |
| Distance to frontiers | 3.42 | 10.77 | 5.94 | 6.60 | 4.09 | 5.52 | 7.76 | 19.68 | |
| Biomass | 7.31 | 5.40 | 12.25 | 5.58 | 3.46 | 4.81 | 8.40 | 17.87 | |
| Animal richness | 9.19 | 6.15 | 0.14 | 13.52 | 36.13 | 10.93 | 7.18 | 11.35 | |
| Vascular plant richness | 2.54 | 5.16 | 0.15 | 5.03 | 10.79 | 10.12 | 7.28 | 15.88 | |
| Average | 7.33 | 5.66 | 6.32 | 6.93 | 10.08 | 6.43 | 6.86 | 13.70 | |
| Opportunistic | Population | 19.93 | 19.72 | 15.96 | 11.93 | 17.17 | 14.59 | 6.04 | 6.73 |
| Isolation | 34.61 | 47.40 | 14.18 | 34.12 | 5.33 | 12.65 | 21.10 | 8.92 | |
| Distance to coasts | 7.22 | 2.83 | 4.81 | 12.37 | 11.36 | 3.76 | 16.99 | 13.07 | |
| Cropland suitability | 3.07 | 1.76 | 39.63 | 4.58 | 2.38 | 32.74 | 21.19 | 2.89 | |
| Average | 16.21 | 17.93 | 18.65 | 15.75 | 9.06 | 15.94 | 16.33 | 7.90 |
Results and Discussion
Globally, opportunistic forces prevailed over preferential and representative motivations in predicting current protection patterns, as protection notably increased towards areas that are isolated, lightly populated, and have low cropland suitability (these three variables are highly correlated, Fig. 3 and Fig. S1). According to the random forest analysis, on average, the importance of the variables related to opportunistic forces doubled in significance those related to preferential motivations (Table 3). These results support previous global explorations that highlighted the importance of opportunistic forces at an ecoregional- (Loucks et al., 2008) or a national- basis (Joppa & Pfaff, 2009). At a regional level, opportunistic forces predominated in North America & Australia–NZ (driven by cropland suitability) and Latin America & Caribbean (driven by isolation) (Table 3 and Figs. S2 and S3). These results challenge Loucks et al’s 2008 realm-based assessment, which showed that globally the number of endemic species was the best variable predicting protected area coverage. Opportunistic forces can lose strength with time (e.g., by road expansion or improvements in crop resistance to biophysical constraints), weakening the legal status of a protected area, a phenomenon of significant magnitude in North America & Australia–NZ, and emergent at the global level (Mascia & Pailler, 2011).
Beyond the imprint of opportunistic forces, protection appeared to respond to preferential motivations that provide benefits to individuals or societies (economic, geopolitical, spiritual). In particular, we found that the tourism attractiveness of an area (Table 2) was positively related to its level of protection (Fig. 3, Figs. S2 and S3), achieving a top importance in the ranking of variables (Table 3). Probably, tourism and protection are involved into positive feedbacks, as protection itself attracts visitors interested in remarkable natural or cultural landscapes, and visitors drive protection to preserve this quality. Tourism engages local communities and regional and national governments in the preservation of these landscapes, offering economic revenues that eventually exceed those obtained from traditional land uses (Mulholland & Eagles, 2002; Siikamäki et al., 2015). As examples, visitors generate annually US$ 1.5 × 109 in the highly populated UK’s Lake District National Park (helping to maintain the landscape naturalness, UK National Parks, 2015). Under a contrasting economic/environmental context, visitors generate annually US$ 2.1 × 107 in the parks inhabited by mountain gorillas in Congo DR, Rwanda, and Uganda (Maekawa et al., 2013). While disentangling the type of existing relationship between tourism and protection is difficult, it is important to note that the most exceptional natural and cultural landscapes around the world are protected under different IUCN categories.
In the last three decades, the inclusion of new species into protection networks as well as the balancing of geographical asymmetries (Stattersfield et al., 1998; Myers et al., 2000; Olson & Dinerstein, 2002) occupied a central place in national and international conservation agendas. However, these motivations are only weakly reflected in the current distribution of protected areas, perhaps because they lead to protection when land is economically unproductive or remote, but not when land is productive and accessible (Margules & Pressey, 2000). Interestingly, new areas created specifically to protect unrepresented environments or species tend to be of small size (Marinaro, Grau & Aráoz, 2012). As a measure on how biological conservation is weakly related to protection, we found that the more intensely protected lands were the poorer in animal and vascular plant species both globally and regionally (Fig. 3 and Fig. S2). Animal and vascular plant richness were positively correlated to cropland suitability at the global level (Kendall’s τ = 0.26 for animals and 0.40 for plants, Fig. S1) revealing how the conflict between this biocentric preference and traditional or profitable land uses exacerbates the current biodiversity crisis (Rodrigues et al., 2004a; Rodrigues et al., 2004b; Hoekstra et al., 2005; Venter et al., 2014). The single exception to these findings appeared in South-east Asia & Oceania (Fig. S2), most likely due to the considerable protected systems in highly-diverse countries like Bhutan, Thailand, Cambodia, and Sri Lanka (Fig. 1) (though see Sodhi et al., 2004). Loucks et al. (2008) found a negative relationship between species richness and protection only at the global level and for the Neotropical realm, but not for the remaining five realms, a discrepancy with our results probably related to their ecoregional approach vs. our grid-based approach.
Representativeness remains nowadays unachieved, as shown by the large biases in the distribution of protected areas along biophysical gradients, with an overrepresentation of lands with extreme climates (polar, arid or very humid), high elevations, complex topographies, and unfertile soils (e.g., Northeast Greenland NP, Denmark; Tassili n’Ajjer NP, Algeria; Fig. 3 and Fig. S2). Even under this context, protection followed closer a fraction- rather than a quota representativeness (1.7-times higher, Fig. 4) according to the modified Shannon evenness index, implying that the current network of protected areas encompasses the most abundant environments. The regions that followed a fraction representativeness more closely were Sub-Saharan Africa and, to a lesser extent, East Europe & Central Asia, while the highest quota representativeness was accomplished by West Europe (Fig. 4). The representativeness levels were lower and similar in the remaining regions, despite the strong differences in their total protected fraction, which ranged from 2.1% in Middle East & North Africa to 11.4% in North America & Australia–NZ (Fig. S2 and Table S2).
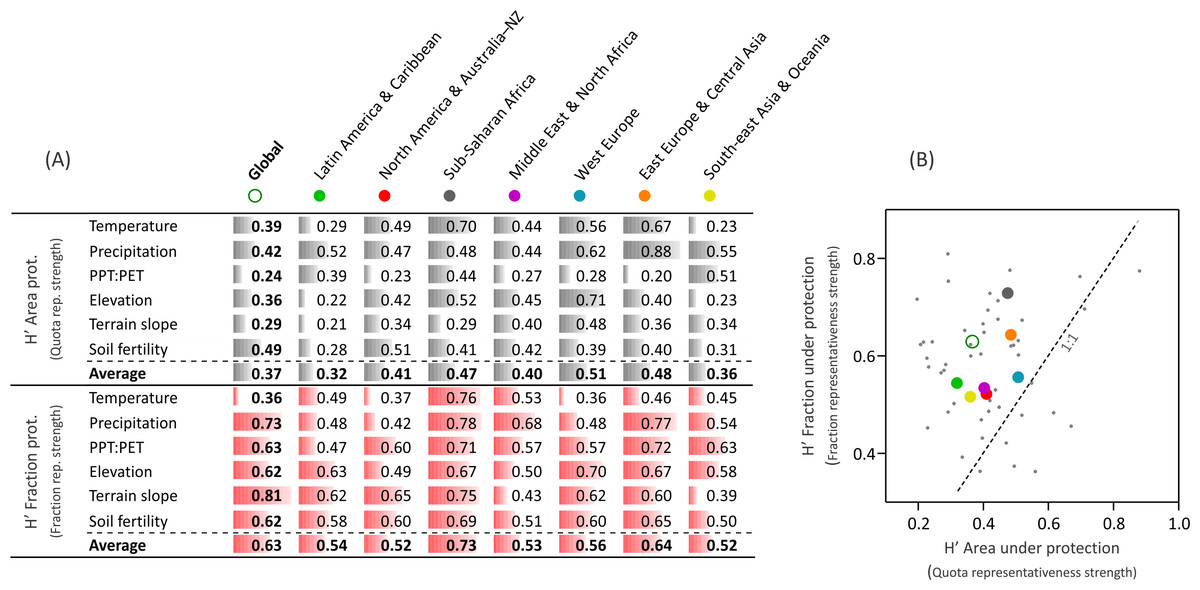
Figure 4: Representativeness according to a modified Shannon evenness.
Modified Shannon evenness (H′) for the biophysical variables. The index ranges between 0 and 1, with a value of 1 when xij is constant along the i gradient. (A) H′ values of the area under protection (in light gray), related to the “quota representativeness” motivation; and H′ values of the fraction under protection (in red), related to the “fraction representativeness”. (B) plot of all H’ values of the 48 biophysical variables * globe/regions combinations (small gray dots), and averaged H′ values for the globe and the seven regions (large colored dots).{kind=link}
Independently of which type of representativeness prevailed, the analysis of protection along biophysical gradients offers the chance to assess the achievement of national and international protection targets and agreements. Among them, the influential Convention on Biological Diversity stipulates that >17% of terrestrial “areas” (i.e., biogeographical units) needed to be included in protected systems by 2020 (SCBD, 2010). Our analyses based on continuous biophysical gradients (which purposely avoid predefined geographical units) show that this protection target is far from being uniformly achieved across the whole array of global environments if we consider exclusively protected areas categorized as I–IV (IUCN, 1994). Regionally, only North America & Australia–NZ in terms of relief, and West Europe in terms of temperature accomplished this protection target (Table S3). The analyses of land protection along biophysical gradients implemented in this study could be used as well to model and to plan future environmental representativeness under a scenario of climate change (Davis & Shaw, 2001; Scott, Malcolm & Lemieux, 2002). The role of unconsidered protected areas (categorized as V “Protected Landscape/Seascape” and VI “Protected area with sustainable use of natural resources” by IUCN—1994) in representativeness remains to be explored.
The predominance of fraction representativeness in the conservation agendas (and in the literature) implies that the environments or geographic units of small extent are unintentionally penalized. The concept of quota representativeness introduced here overcomes this problem, broadening what an equal representation should be. In fact, quotas are often considered in political organization, as many countries have formal electoral rules which warrant a minimum participation of minorities (e.g., ethnic, gender) or an equal contribution of subnational to national administrative entities regardless of their population size (Bird, 2014). This complementary quota conservation approach would ultimately overcome the long-lasting conservation dilemma of hotspot/species-richness vs. coldspot/species-poorness (Myers et al., 2000; Kareiva & Marvier, 2003), as each environment has per se an equal importance (including its encompassed biological distinctiveness and evolutionary strategies).
Regional differences in the weight of alternative motivations and opportunistic forces likely reflect the interactions among direct drivers (e.g., conservation agendas), that—following Lambin, Geist & Lepers (2003)—can be conceptualized as:
Motivations and opportunistic forces =f (policies and economy, social organization, moral rules); with
-
policies and economy =f(agendas, economic/financial contexts, property rights, state-owned lands, infrastructure, governance);
-
social organization =f(urban-rural interactions, ONG and philanthropists actions);
-
moral rules =f(importance of religion, priority to environmental protection, deference to authority, trust and tolerance, economic/physical security);
with the functions f having variable forms at the time of the establishment of protection. Even though direct drivers have been previously linked to the protected fraction on a country-basis (McDonald & Boucher, 2011), very few studies formulated or assessed their interactions with motivations (Marinaro, Grau & Aráoz, 2012), identifying an adequate fraction representation with strong economies, “modern” societies or states, or extensive and long lasting protection networks. However, North America & Australia–NZ and West Europe, representing these conditions with a pioneering and profuse history of protection (Table S2), were surpassed in the fraction representativeness by other regions, and—at the same time—surpassed others in terms of the weight of opportunistic forces (especially cropland suitability) and preferential motivations (especially tourism attractiveness). The strength of tourism attractiveness in these two regions (Table 3 and Figs. S2, and S3) may reflect the combination of an affluent population capable of devoting resources to “luxury” goods and services (in this case, conservation; Marinaro, Grau & Aráoz, 2012) and a growing need to access natural settings by highly urbanized societies (Pyle, 2003).
With an opposite socioeconomic context, Sub-Saharan Africa reached the top of the representativeness ranking, perhaps due to the historical indirect effect of colonial regimes, unconstrained by the local social organization and with conservation agendas decoupled from local population needs and wills (Naughton-Treves, Holland & Brandon, 2005). In this regard, Sub-Saharan Africa showed the highest fraction of protected areas established before the formation of modern states (Table S2). Historical factors can be ascribed as well to the protection and consolidation of international frontiers (Table 1), as asserting sovereignty and the possibilities of armed conflicts had a high relative weight in national politics in many new countries around the world (under autocratic governments or young democracies) (Zbicz & Green, 1997; Hegre, 2003). This motivation appeared to be especially influential in Latin America & Caribbean and South-east Asia & Oceania, where there is a large concentration of protected areas within the first hundreds of kilometers from borders (Fig. S2) and where there is a large fraction of post-independence protected areas (Table S2). How the change of these direct drivers might affect the relative strength of motivations and opportunistic forces remains to be explored, especially considering the transition of the promotion and deployment of protected areas from national governments to philanthropists and non-governmental organizations, or the empowerment of indigenous peoples or local rural populations (McNeely & Schutyser, 2003; Naughton-Treves, Holland & Brandon, 2005).
We should issue certain caveats from our analyses. First, the time dimension has not been explored, yet it could reveal important shifts in the strength of protection motivations and opportunistic forces (Joppa & Pfaff, 2009; Marinaro, Grau & Aráoz, 2012) and in the impact of evolving conservation paradigms (Mace, 2014). Second, the sampling approach implies a spatial integration of data into grid cells, and thus the results can mask heterogeneous biophysical or human conditions. For example, our analyses do not reveal the fact that some small protected areas that abut urban or productive areas were established under locally rough topographies and/or poor soils (e.g., Tijuca NP, Brazil; Sanjay Gandhi NP, India). An assessment focused on individual protected areas rather than on cells would solve this problem and would allow exploring the spatial dependencies in relation with the geometry of protected areas, as small and large areas may have different origins and geographical contexts (Andrew, Wulder & Coops, 2011). Third, our results are most probably affected by multicollinearity, as the explanatory variables of the distribution of protected areas are, by nature and in nature, correlated (e.g., cropland suitability derives—among other variables—from temperature). The applied random forest technique handles this phenomenon by means of the random selection of input variables at each node creation (mtry), but can not remove it completely (Breiman, 2001; Graham, 2003). Fourth, our study subjectively groups countries into regions and defines explanatory variables (biophysical, human, and biological) as proxies of individual motivations and opportunistic forces. Regarding the spatial grouping, even though the regions shared cultural, historical, and biogeographical traits, the proximate causes of protection (as defined above) and their consequences vary considerably within regions (e.g., Venezuela protecting 18.9% of its territory vs. Argentina protecting just 1.7%). Regarding the proxy variables, even though we considered the most up-to-date and accurate global information as far as we know, their selection could be modified, expanded, or improved with new or more suited options. For example, Durán et al. (2013) evaluated the representation within the Chilean protected network of different ecosystem services, including under this category the primary production, the carbon storage, the species richness, and the agricultural production. In this sense, our theoretical/methodological schemes can be subject to modifications and criticisms, and the precision and stability of our findings should be verified following alternative approaches.
Conclusions
Present-day protected areas are mostly located in zones of relatively low productive value or population pressure, and to a lesser extent in areas of high tourism attractiveness. The search for geographical or biophysical representativeness and biodiversity conservation has had a relatively minor effect in shaping the distribution of land protection, in spite of their explicit priority in the debates and agendas of national and international conservation agencies. These geographical patterns will probably persist or increase (McNeely & Schutyser, 2003) under the concurrent expansion of protected networks (Jenkins & Joppa, 2009) and the increasing pressure on land resources (Foley et al., 2007; Ellis & Ramankutty, 2008). In this sense, representativeness and biodiversity conservation will only be strengthened if coupled with opportunistic forces. Operatively, this coupling requires a more explicit identification and spatial representation of conservation motivations (e.g., what are protection needs and targets of societies) and opportunities (e.g., where is it feasible to meet these needs and targets given current geographical and social conditions) (Andrew, Wulder & Coops, 2012; Martin et al., 2014). At last, if humans are increasingly considered as modelers and dependents of nature at regional and global levels (Van den Born et al., 2001; Lambin & Meyfroidt, 2011), future conservation policies will need to consider the role of goods and services like water provision, or tourism values (Durán et al., 2013) and the basic human need to interact with nature, which increases happiness and health, and fosters an environmentally sustainable behavior (Loreau, 2014; Zelenski & Nisbet, 2014).