Unique morphogenetic signatures define mammalian neck muscles and associated connective tissues
- Institut Pasteur, France
- CNRS UMR 3738, France
- Brno University of Technology, Czech Republic
- University of Utah, United States
- Aix-Marseille Université, CNRS UMR 7288, IBDM, France
Abstract
In vertebrates, head and trunk muscles develop from different mesodermal populations and are regulated by distinct genetic networks. Neck muscles at the head-trunk interface remain poorly defined due to their complex morphogenesis and dual mesodermal origins. Here, we use genetically modified mice to establish a 3D model that integrates regulatory genes, cell populations and morphogenetic events that define this transition zone. We show that the evolutionary conserved cucullaris-derived muscles originate from posterior cardiopharyngeal mesoderm, not lateral plate mesoderm, and we define new boundaries for neural crest and mesodermal contributions to neck connective tissue. Furthermore, lineage studies and functional analysis of Tbx1- and Pax3-null mice reveal a unique developmental program for somitic neck muscles that is distinct from that of somitic trunk muscles. Our findings unveil the embryological and developmental requirements underlying tetrapod neck myogenesis and provide a blueprint to investigate how muscle subsets are selectively affected in some human myopathies.
https://doi.org/10.7554/eLife.40179.001Introduction
The neck is composed of approximately 80 skeletal muscles in humans that allow head mobility, respiration, swallowing and vocalization processes, containing essential elements such as the trachea, esophagus, larynx, and cervical vertebrae. These processes are ensured by a robust network of muscles at the head-trunk interface, a transition zone subjected to a spectrum of human muscle diseases such as dropped head syndrome, oculopharyngeal myopathy, myotonic dystrophy, Duchenne-type dystrophy and congenital muscular disorders (Emery, 2002; Martin et al., 2011; Randolph and Pavlath, 2015). Defining the embryology of these distinct muscle groups is critical to understand the mechanisms underlying the susceptibility of specific muscles to muscular dystrophies. While myogenesis at the cranial and trunk levels has been studied extensively, the developmental mechanisms at the basis of neck muscle formation are poorly documented and often debated (Ericsson et al., 2013).
In vertebrates, head and trunk muscles arise from different mesodermal origins and their development depends on distinct myogenic programs. At the cranial level, the cardiopharyngeal mesoderm (CPM) resides in pharyngeal arches and gives rise to branchiomeric muscles and the second heart field. CPM specification is initiated by the activation of genes such as Mesp1, Islet1 and Tbx1, while Pax7 subsequently marks muscle stem cells (Diogo et al., 2015; Kelly et al., 2004; Nathan et al., 2008; Saga et al., 1996; Sambasivan et al., 2009). In contrast, Pax3 and Pax7 are expressed in somitic mesoderm giving rise to trunk and limb muscles, with Pax3 then being downregulated in most muscles during fetal stages, while Pax7 maintains the stem cell pool (Kassar-Duchossoy et al., 2005; Relaix et al., 2005; Tajbakhsh et al., 1997). After the differential specification of cranial and trunk progenitors, the bHLH myogenic regulatory factors (MRFs) Myf5, Mrf4, Myod and Myog regulate myogenic cell fate and differentiation (reviewed in [Comai and Tajbakhsh, 2014; Noden and Francis-West, 2006]).
In early embryos, Tbx1 is required for robust activation of MRF genes and proper branchiomeric muscle formation (Grifone et al., 2008; Kelly et al., 2004; Kong et al., 2014; Sambasivan et al., 2009). In Tbx1 mutant embryos, the first pharyngeal arch is hypoplastic and posterior pharyngeal arches do not form resulting in variably penetrant defects of masticatory muscles and absence of muscles derived from more posterior arches (Kelly et al., 2004; Lescroart et al., 2015; Theis et al., 2010). In humans, TBX1 is a major gene involved in 22q11.2 deletion syndrome (DiGeorge/velo-cardio-facial syndrome), a congenital disease characterized by cardiovascular defects and craniofacial malformations (Papangeli and Scambler, 2013). In contrast, Pax3 acts upstream of MRF genes in somites and Pax3 mutants have defects of epaxial and hypaxial muscle formation while double Pax3/Pax7-null embryos lack trunk/limb muscles (Brown et al., 2005; Relaix et al., 2005; Tajbakhsh et al., 1997; Tremblay et al., 1998).
The neck constitutes a transition zone characterizing land vertebrates (tetrapods). The major muscle groups in the neck consist of: epaxial back muscles; ventral hypaxial musculature; pharyngeal, laryngeal and esophagus striated muscles located medioventrally; and cucullaris-derived muscles. The cucullaris is a generic term defining putative homologous muscles that are evolutionarily conserved and connect the head and trunk in jawed vertebrates (gnathostomes). In amniotes, the cucullaris represents the embryonic anlage that gives rise to trapezius and sternocleidomastoid muscles which are innervated by the accessory nerve XI (Diogo, 2010; Edgeworth, 1935; Ericsson et al., 2013; Kuratani, 2008; Kuratani et al., 2018; Lubosch, 1938; Tada and Kuratani, 2015).
While the somitic origin of epaxial/hypaxial neck muscles and CPM origin of pharyngeal, laryngeal and esophagus striated muscles are well defined (Gopalakrishnan et al., 2015; Noden, 1983; Tabler et al., 2017), the embryological origin of cucullaris-derived muscles has remained a subject of controversy (Couly et al., 1993; Edgeworth, 1935; Greil, 1913; Huang et al., 1997; Huang et al., 2000; Matsuoka et al., 2005; Noden, 1983; Piatt, 1938; Piekarski and Olsson, 2007). This muscle group was reported to originate either from lateral plate mesoderm (LPM) or CPM populations adjacent to the first three somites in chick and axolotl (Nagashima et al., 2016; Sefton et al., 2016; Theis et al., 2010). However, retrospective lineage analysis indicated that the murine trapezius and sternocleidomastoid muscles are clonally related to second heart-field-derived myocardium and laryngeal muscles, consistent with a CPM origin (Lescroart et al., 2015). Moreover, cucullaris development follows a branchiomeric program and cucullaris-derived muscles were shown to be absent in Tbx1-null mice (Kelly et al., 2004; Lescroart et al., 2015; Sefton et al., 2016; Theis et al., 2010). Nevertheless, the source of the cucullaris is still equivocal due to the location of its embryonic anlagen at the interface of cranial, somitic and lateral plate mesodermal populations.
Skeletal elements and muscle-associated connective tissue (MCT) also have distinct embryological origins along the rostro-caudal axis. The connective tissue of branchiomeric and tongue muscles originate from neural crest cells (NCCs) of cranial origin (Evans and Noden, 2006; Köntges and Lumsden, 1996; Noden, 1983; Noden, 1988; Ziermann et al., 2018b). Cranial NCCs also give rise to skeletal components and tendons in the head. In contrast, the skeleton and connective tissue originate from somitic mesoderm in the trunk and from LPM in limbs (Nassari et al., 2017). The neck and shoulder girdle contain skeletal elements and connective tissues of distinct NCC, LPM or somitic origins (Durland et al., 2008; Matsuoka et al., 2005; McGonnell et al., 2001; Nagashima et al., 2016; Tabler et al., 2017; Valasek et al., 2010). It has been suggested that NCCs form both connective tissue and endochondral cells at the attachment sites of neck muscles to shoulders in mouse (Matsuoka et al., 2005). However, studies in non-mammalian animals have contested a NCC contribution to the pectoral girdle (Epperlein et al., 2012; Kague et al., 2012; Ponomartsev et al., 2017).
Therefore, the neck region consists of muscle, skeletal and connective tissue elements of mixed cellular origins, underscoring the difficulty in assigning embryonic identities for these structures. In addition, the genetic requirements for the formation of non-somitic and somitic neck muscles remain to be defined. To resolve these issues, we used genetic lineage and loss-of-function mice combined with histology, µCT and 3D reconstructions to map the embryological origins of all neck muscles and associated connective tissues. In doing so, we show that cucullaris-derived muscles originate from a posterior CPM population and are differentially affected in Tbx1-null mice. Moreover, we identify a unique genetic network involving both Mesp1 and Pax3 genes for somite-derived neck muscles and we define a new limit of neural crest contribution to neck connective tissue and shoulder components.
Results
Distinct myogenic programs define neck muscle morphogenesis
To investigate the embryological origin of neck muscles in the mouse, we mapped CPM- and somite-derived myogenic cells using lineage-specific Cre drivers including Mef2c-AHF, Islet1, Mesp1 and Pax3 (Figure 1). The Mef2c-AHF (anterior heart field) enhancer is activated in the second heart field and myogenic progenitors of CPM origin (Lescroart et al., 2010; Verzi et al., 2005). Islet1 and Mesp1 genes are both expressed in early CPM and are essential for cardiac development. The Mesp1 lineage also marks some anterior somitic derivatives (Cai et al., 2003; Harel et al., 2009; Saga et al., 2000; Saga et al., 1999). In contrast, Pax3 is activated in all somitic progenitors and is a key actor during trunk and limb muscle formation (Relaix et al., 2005; Tajbakhsh et al., 1997; Tremblay et al., 1998). Given that the majority of Mef2c-AHF derivatives are myogenic cells (Lescroart et al., 2015; Lescroart et al., 2010; Verzi et al., 2005), we analyzed this lineage using Rosa26R-lacZ/+ (R26R) reporter mice. Islet1, Mesp1 and Pax3 genes are also expressed in cells contributing to skeletal components, connective tissues or neurons. To focus on the myogenic lineage, we used Pax7nGFP-stop/nlacZ (Pax7GPL) reporter mice, which mark cells with nuclear β-galactosidase (β-gal) activity following Cre recombination (Sambasivan et al., 2013).
Figure 1 with 3 supplements see all
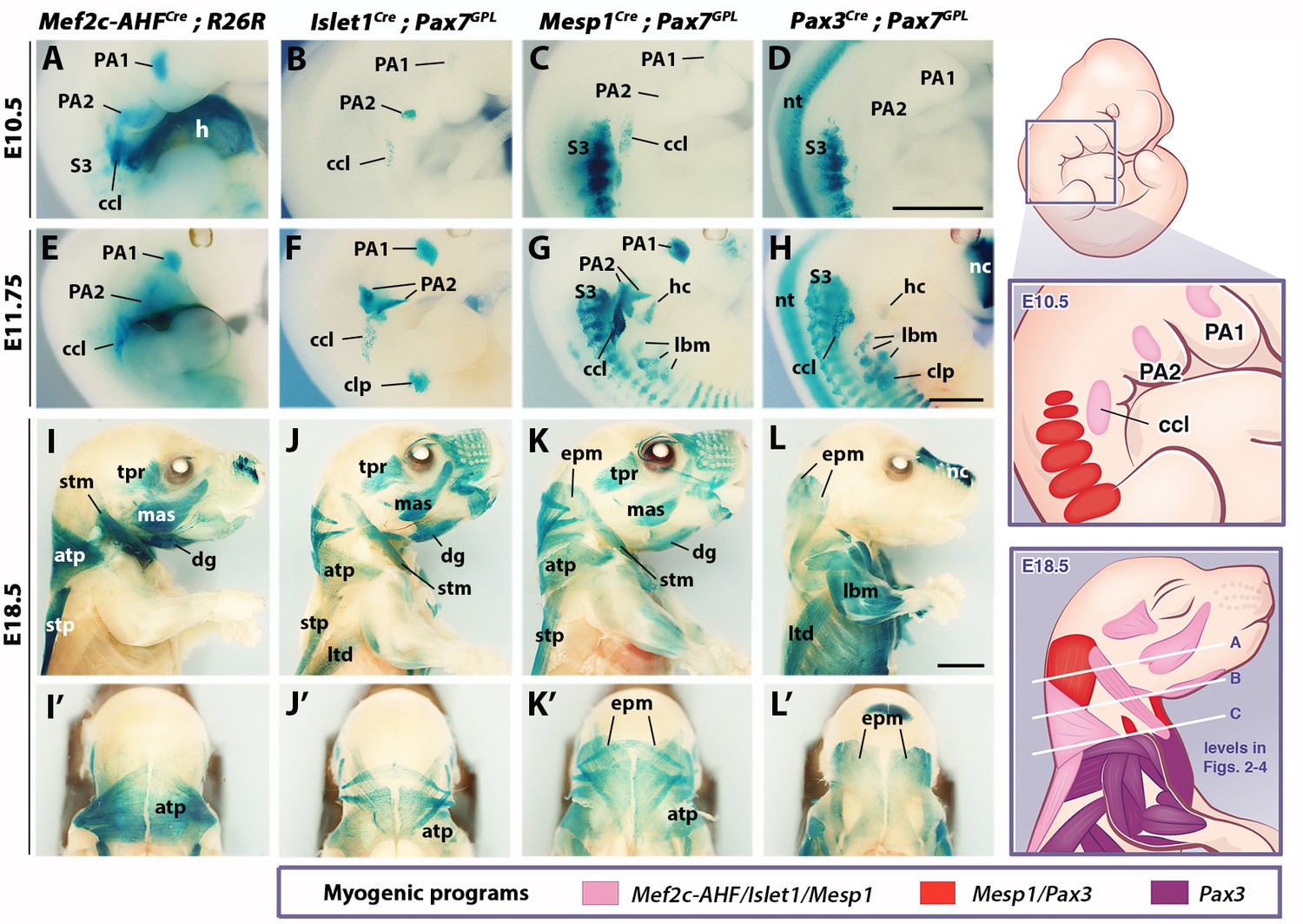
Genetic lineage tracing of neck muscle progenitors.
Whole-mount X-gal stainings of Mef2c-AHFCre;R26R, Islet1Cre;Pax7GPL, Mesp1Cre;Pax7GPL and Pax3Cre;Pax7GPL mice at E10.5 (A–D), E11.75 (E–H) and E18.5 (I–L’) (n = 3 for each condition). See associated Figure 1—supplements 1–3. (A–H) Note labeling of mesodermal core of pharyngeal arches (PAs) and cucullaris anlage (ccl) by Mef2c-AHF, Islet1 and Mesp1 lineage reporters; β-gal+ cells in anterior somites of Mesp1Cre embryos and in the clp anlagen of Islet1Cre embryos. Pax3 lineage marked somitic mesoderm. (I–L’) Mef2c-AHF, Islet1 and Mesp1 lineages marked branchiomeric (mas, tpr, dg) and cucullaris muscles (stm, atp and stp). Pax3Cre and Mesp1Cre labeled somitic epaxial neck muscles (epm). atp, acromiotrapezius; ccl, cucullaris anlage; clp, cutaneous maximus/latissimus dorsi precursor; dg, digastric; epm, epaxial musculature; h, heart; hc, hypoglossal cord; lbm, limb muscle anlagen and limb muscles; ltd, latissimus dorsi; mas, masseter; nc, nasal capsule; nt, neural tube; PA1-2, pharyngeal arches 1–2; S3, somite 3; stm, sternocleidomastoid; stp, spinotrapezius; tpr; temporal. Scale bars: in D for A-D and in H for E-H, 1000 µm; in L for I-L’, 2000 µm.
We first examined embryos after myogenic specification (E10.5 and E11.75), and fetuses when muscles are patterned (E18.5). In Mef2c-AHFCre;R26R embryos, β-gal-positive cells were observed in the mesodermal core of pharyngeal arches at the origin of branchiomeric muscles, in second heart field derivatives, and in the cucullaris anlage (Figure 1A,E). A spatiotemporal analysis of the cucullaris using Myf5Cre;Pax7GPL and Myf5Cre;R26mTmG embryos (Figure 1—figure supplement 1) showed that Myf5-derived muscle progenitors located at the level of the posterior pharyngeal arches, and adjacent to somites S1-S3 (Figure 1—figure supplement 1A’), were innervated by the accessory nerve XI (Figure 1—figure supplement 1G–G”). These cells gave rise to the trapezius and sternocleidomastoid muscles (Figure 1—figure supplement 1A–F’) thus confirming the identity of the cucullaris anlage in mouse (Tada and Kuratani, 2015).
In Islet1Cre;Pax7GPL and Mesp1Cre;Pax7GPL embryos, labeling was also observed in pharyngeal arch derivatives and the cucullaris (Figure 1B–C,F–G), the latter showing less contribution from the Islet1 lineage. On sections, a subset of the Myod-positive cells in the cucullaris originated from Islet1-derived cells (Figure 1—figure supplement 2A). Surprisingly, Pax3Cre;Pax7GPL embryos also showed lacZ expression in the cucullaris at E11.75, although no expression was detected at E10.5 (Figure 1D,H). Given that Pax3 and Pax7 are also expressed in neural crest cells (Relaix et al., 2004), and that these Pax3/Pax7-derived cells were excluded from the Myod-positive myogenic population at E12.5 after muscle specification (Figure 1—figure supplement 2B), they are likely to be of NCC origin. As expected, Pax3 lineage tracing also labeled the somite-derived myotomes, hypaxial migrating progenitors that form the hypoglossal cord (origin of tongue and infrahyoid muscles), and limb muscle progenitors. Furthermore, the hypaxial anlage, which is located at the proximal limb bud and gives rise to the cutaneous maximus and latissimus dorsi muscles, was Pax3-derived (Figure 1D,H; Figure 1—figure supplement 1D’) (Prunotto et al., 2004; Tremblay et al., 1998). Unexpectedly, this anlage and the latissimus dorsi muscle were also labeled in Islet1Cre;Pax7GPL but not in Mesp1Cre;Pax7GPL embryos (Figure 1F–G,J–K). On sections at E12.5, Islet1 expression was observed in Pax3-derived cells after the emergence of myogenic cells from the proximal limb bud (Figure 1—figure supplement 2C). In addition, the Mesp1 lineage contributed to anterior somitic derivatives during early embryonic development as previously reported (Loebel et al., 2012; Saga et al., 1999); strong lacZ expression was observed in the hypoglossal cord and somites S1-S6. Labeling decreased in more posterior myotomes and in forelimb muscle progenitors compared to Pax3Cre;Pax7GPL embryos (Figure 1C–D,G–H).
Lineage tracings with Mef2c-AHFCre, Islet1Cre and Mesp1Cre marked branchiomeric (temporal, masseter, digastric, mylohyoid and pharyngeal) and cucullaris-derived neck muscles (acromiotrapezius, spinotrapezius and sternocleidomastoid), all of which were excluded from the Pax3 lineage (Figure 1I–L, Figure 1—figure supplement 2D–G’). These findings support previous studies showing that cucullaris muscle development is controlled by a branchiomeric myogenic program (Kelly et al., 2004; Lescroart et al., 2015; Sefton et al., 2016; Theis et al., 2010). In addition, both Mesp1 and Pax3 lineages labeled somitic neck muscles (Figure 1K–L’, Figure 1—figure supplement 2F–G’).
Analysis of different somite-derived neck muscles on sections showed that Mesp1 and Pax3 lineages gave rise to the great majority of the Pax7-positive myogenic population (Figure 1—figure supplement 3), demonstrating the high recombination efficiency obtained with the Cre lines. The results indicate that neck somitic muscles originate from myogenic cells that have expressed both Mesp1 and Pax3 genes.
To further investigate the contributions of Mesp1 and Pax3 lineages to neck muscles, we examined sections using the R26tdTomato reporter co-immunostained with the myofibre marker Tnnt3 at three representative levels (A, B and C levels in Figure 1; see also Figure 2—figure supplement 1). At anterior levels, while Pax3 lineage contribution was limited to somite-derived neck muscles, the Mesp1 lineage marked branchiomeric muscles (mylohyoid, pharyngeal, laryngeal, esophagus), cucullaris-derived muscles (acromiotrapezius and sternocleidomastoid) and somite-derived neck muscles (Figure 2A–H, Figure 1—figure supplement 2F–G’, Figure 2—figure supplement 2A–H’). The epaxial and hypaxial neck muscles showed equivalent Tomato expression in both Mesp1Cre;R26tdTomato and Pax3Cre;R26tdTomato mice. These observations further indicate that Mesp1 and Pax3 lineages contribute equivalently to neck muscles derived from anterior somites.
Figure 2 with 2 supplements see all
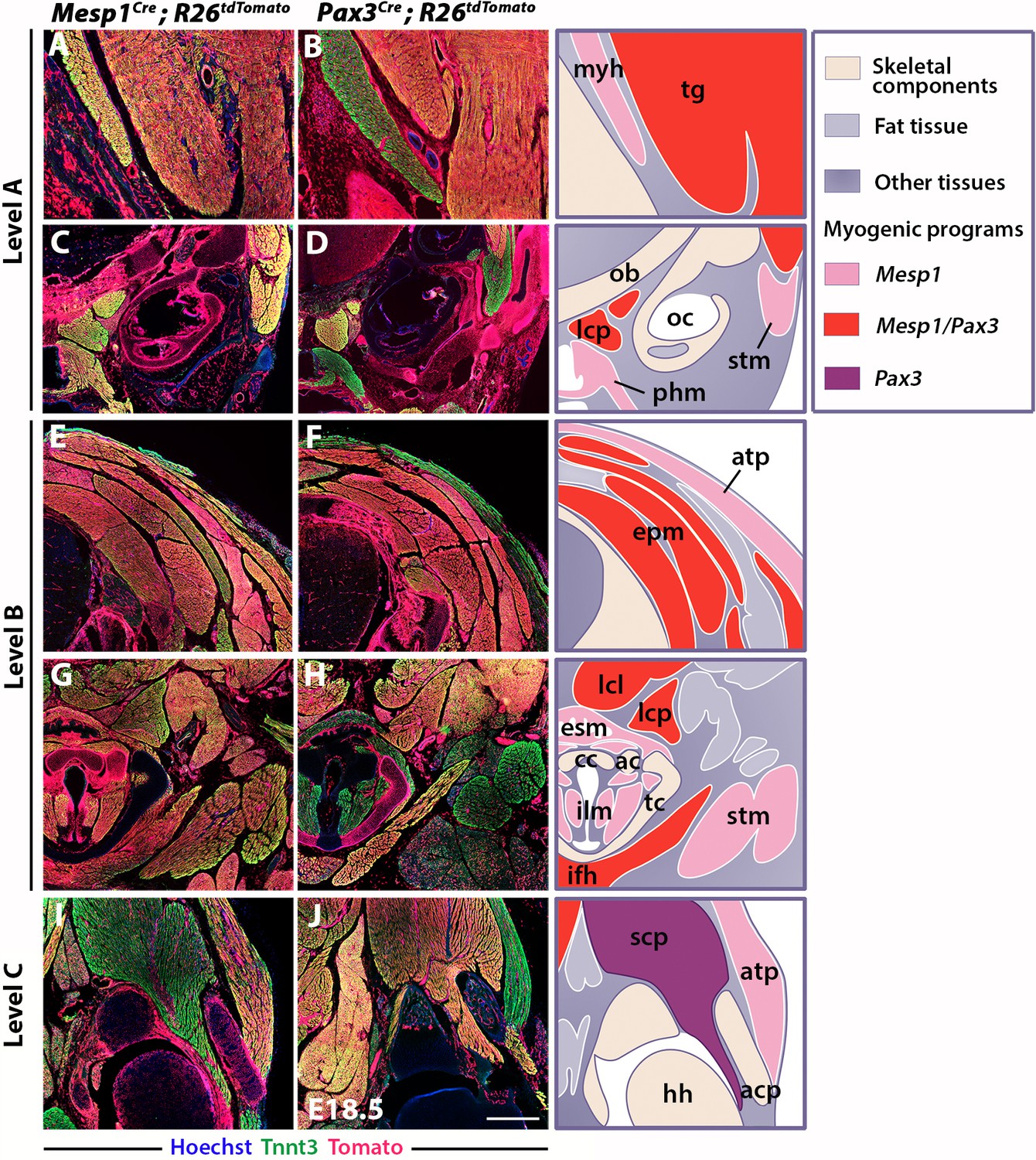
Differential contributions of Mesp1 and Pax3 lineages to neck and shoulders.
Immunostainings on coronal cryosections of E18.5 Mesp1Cre;R26tdTomato and Pax3Cre;R26tdTomato mice for the myofibre Tnnt3 and Tomato markers at levels indicated in Figure 1. Higher magnifications of selected areas in (A–J) are shown in Figure 2—figure supplement 2; (n = 2 for each condition). See also the atlas of neck musculature in Figure 2—figure supplement 1. (A–H) Mesp1Cre labeled all neck muscles including branchiomeric (myh, esm, phm and ilm), cucullaris (stm, atp), somitic epaxial (epm) and hypaxial (tg, lcp, lcl, ifh) muscles. Pax3Cre marked somitic muscles. (I–J) At shoulder level, Mesp1-derived cells did not contribute to posterior somitic myofibres including scapular muscles (scp) compared to that observed in Pax3Cre embryos. ac, arytenoid cartilage; acp, scapular acromion process; atp, acromiotrapezius; cc, cricoid cartilage; epm, epaxial musculature; esm, esophagus striated muscle; hh, humeral head; ifh, infrahyoid muscles; ilm, intrinsic laryngeal muscles; lcl, longus colli; lcp, longus capitis; myh, mylohyoid; ob, occipital bone; oc, otic capsule; phm, pharyngeal muscles; stm, sternocleidomastoid; scp, scapular musculature; tc, thyroid cartilage; tg, tongue. Scale bars: in J for A-B 200 µm, for C-J 400 µm.
At the shoulder level, we observed less Mesp1 contribution to more posterior somitic muscles (Figure 2I–J). In contrast to that observed at anterior levels, little or no Tomato expression was detected in myofibres of scapular muscles in Mesp1Cre;R26tdTomato mice (Figure 2—figure supplement 2I–J’). Therefore, Mesp1 lineage contribution was restricted to epaxial and hypaxial neck muscles, in contrast to pectoral and trunk muscles that originate from the Pax3 lineage (Figures 1–2) (Table 1). These observations lead us to propose that three distinct myogenic programs are involved in the formation of neck and pectoral musculature at the head-trunk interface. The branchiomeric and cucullaris-derived muscles depend on a common myogenic program involving Mef2c-AHF, Islet1 and Mesp1 lineages; the somitic neck muscles that originate from anterior somites derive from both Mesp1 and Pax3 lineages; the pectoral muscles derived from more posterior somites depend on the activation of Pax3 only (Table 1).
Table 1
Contribution of Mef2c-AHF, Islet1, Mesp1 and Pax3 lineages to neck and pectoral musculature.
https://doi.org/10.7554/eLife.40179.009| Mef2c/Islet1/Mesp1-derived muscles | Mesp1/Pax3-derived muscles | Pax3- derived muscles |
|---|---|---|
| Mylohyoid Digastric muscles Pharyngeal muscles Intrinsic laryngeal muscles Esophagus striated muscle Sternocleidomastoid Acromiotrapezius Spinotrapezius | Epaxial neck muscles (splenius, semispinalis, levator scapula, rhomboid occipitalis, suboccipital and postvertebral muscles) Hypaxial neck muscles (tongue muscles*, infrahyoid muscles, longus capitis, longus colli) | Scapular muscles (supraspinatus, Infraspinatus, subscapularis) Pectoralis Latissimus dorsi† Cutaneous maximus† |
| Branchiomeric myogenic program | Anterior-most somite myogenic program | More posterior somite myogenic program |
-
*Including intrinsic and extrinsic tongue muscles of somitic origin
†Also derived from an Islet1 lineage
Dual neural crest and mesodermal origins of neck connective tissues
To define the cellular origin of neck muscle-associated connective tissue (MCT), we traced the contribution of different embryonic populations using Mesp1Cre;R26tdTomato and Pax3Cre;R26tdTomato mice as well as Wnt1Cre and Prx1Cre reporters that label NCC and postcranial LPM derivatives, respectively (Burke and Nowicki, 2003; Danielian et al., 1998; Durland et al., 2008). Both NCC and LPM populations were reported to contribute to trapezius MCT (Durland et al., 2008; Matsuoka et al., 2005). Moreover, it was suggested that the postcranial LPM is a source for cucullaris-derived muscles (Theis et al., 2010). A direct comparison of NCC and LPM derivatives allowed us to clarify the contribution of these two populations to cucullaris formation (Figures 3–4).
Figure 3 with 4 supplements see all
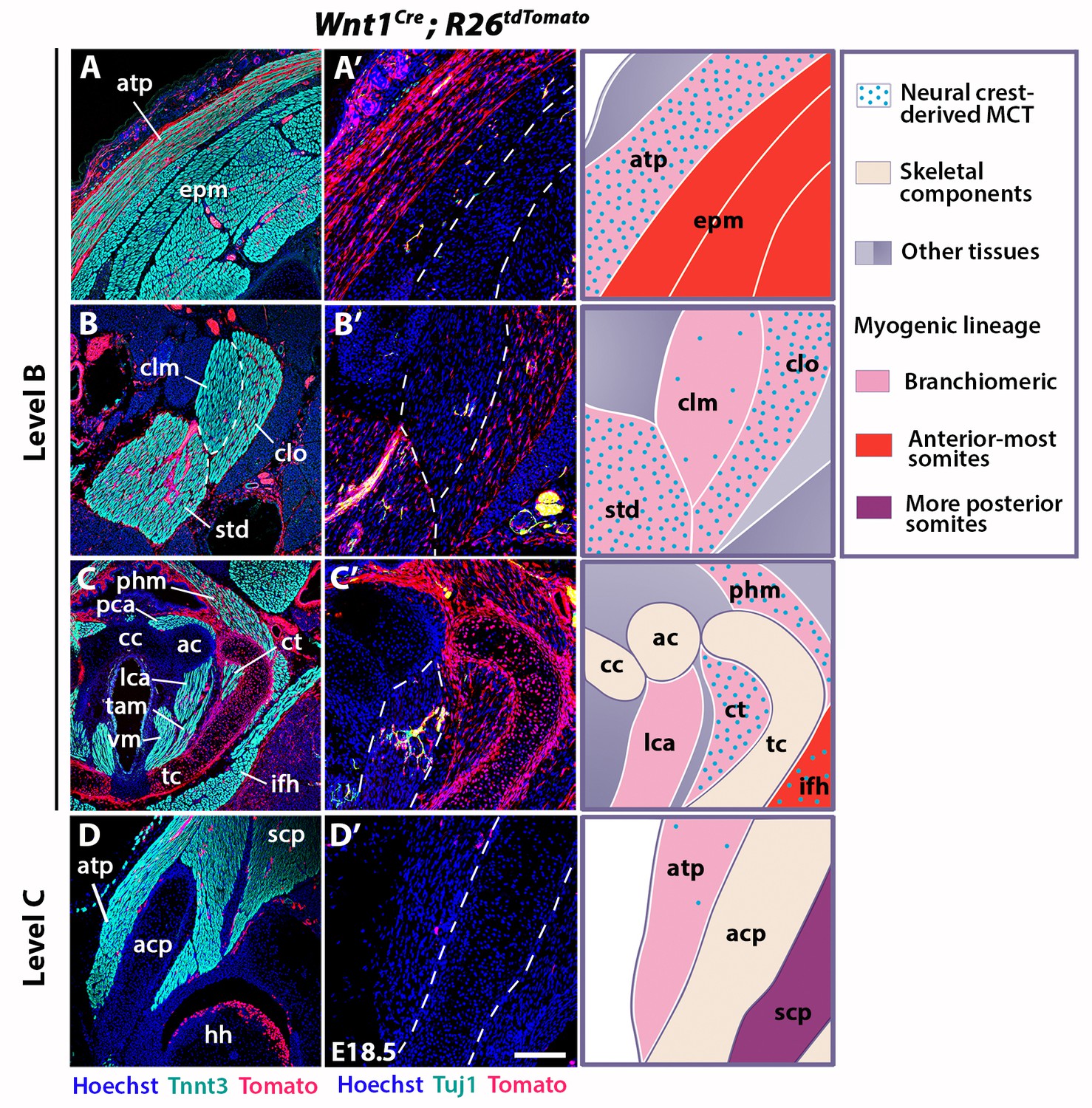
Neural crest contribution to neck muscle-associated tissue.
Immunostainings on coronal cryosections of E18.5 Wnt1Cre;R26tdTomato mice at levels indicated in Figure 1. Tnnt3/Tomato immunostainings are shown in (A–D) and immunostainings for Tuj1/Tomato on selected areas of (A–D) are shown with higher magnifications in (A’–D’). See associated Figure 3—figure supplement 1–4; (n = 2). (A–A’) Note high Wnt1 contribution in the acromiotrapezius but not in epaxial muscles where Wnt1-derived cells marked neuronal cells. (B–C’) Wnt1-derived cells marked differentially the distinct muscles composing the sternocleidomastoid and laryngeal musculatures. (D–D’) At shoulder level, the Wnt1 cells did not contribute to attachment of acromiotrapezius to scapula. ac, arytenoid cartilage; acp, scapular acromion process; atp, acromiotrapezius; cc, cricoid cartilage; clm, cleidomastoid; clo, cleido-occipitalis; ct, cricothyroid; epm, epaxial musculature; hh, humeral head; ifh, infrahyoid muscles; lca, lateral cricoarytenoid; MCT, muscle-associated connective tissue; pca, posterior cricoarytenoid; phm, pharyngeal muscles; scp, scapular musculature; std, sternomastoid; tam, thyroarytenoid muscle; tc, thyroid cartilage; vm, vocal muscle. Scale bars: in D’ for A-D 400 µm for A’-D’ 200 µm.
Figure 4 with 2 supplements see all
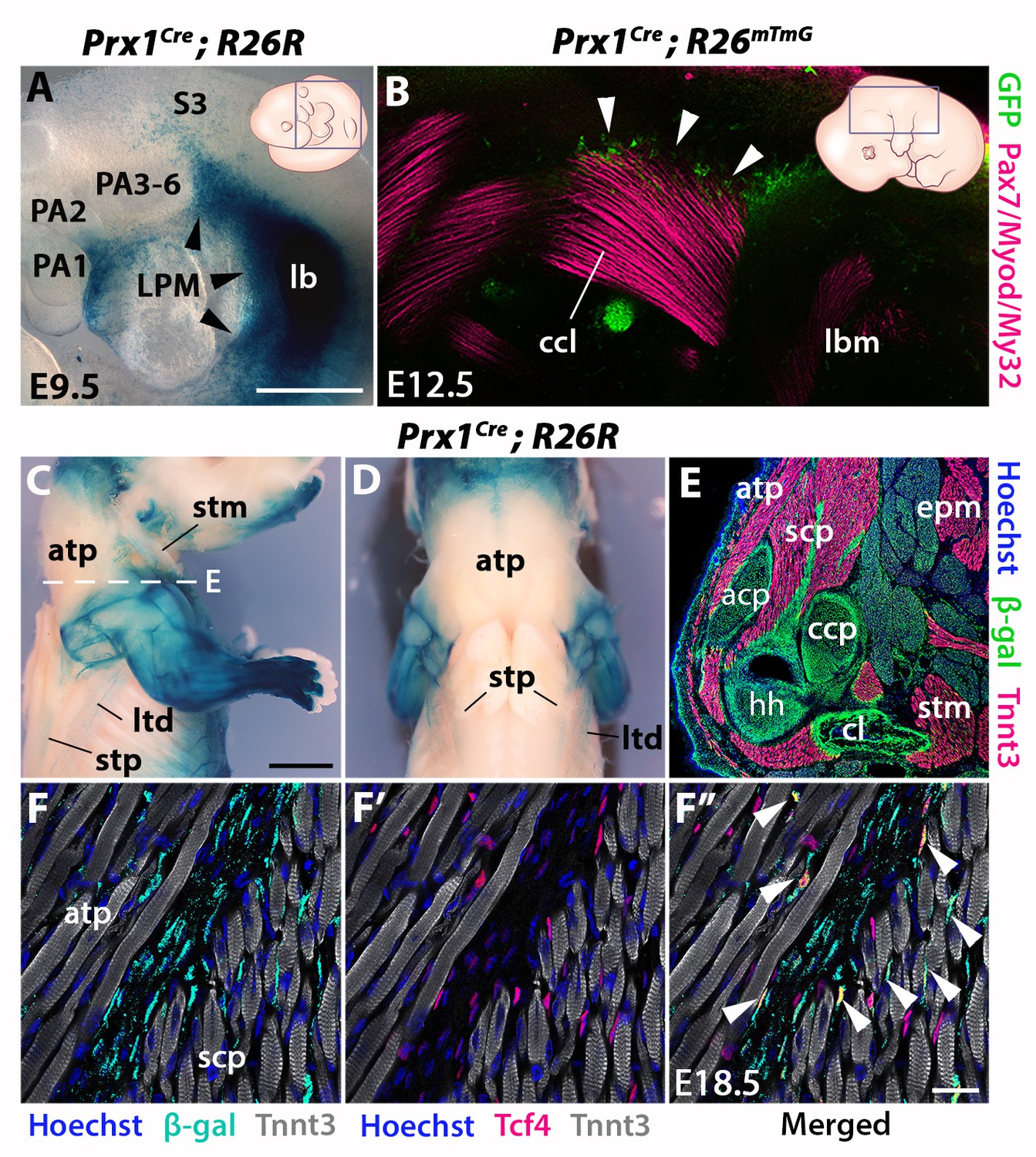
Prx1-LPM lineage contribution to neck and pectoral girdle.
See also Figure 4—figure supplement 1, 2. (A–D) X-gal stainings of Prx1Cre;R26R reporter mice at E9.5 (n = 3) (A) and E18.5 (n = 3) (C–D), and immunostaining for GFP and the Pax7/Myod/My32 myogenic markers in Prx1Cre;R26mTmG E12.5 embryo (n = 2) (B). Note Prx1-derived cells in postcranial LPM (A, black arrowheads) and Prx1-derived cells among, but not in, cucullaris myofibres (B–D). (E–F’’) Immunostaining for β-gal, Tnnt3 and Tcf4 on coronal cryosections of E18.5 Prx1Cre;R26R mice (n = 2) showed β-gal+ cells constituting the pectoral girdle (E, level C in Figure 1) and in MCT fibroblasts (F-F’’, white arrowheads), but not in trapezius myofibres. acp, scapular acromion process; atp, acromiotrapezius; ccl, cucullaris anlage; ccp, scapular coracoid process; cl, clavicle; epm, epaxial musculature; hh, humeral head; lb, forelimb bud; lbm, limb muscle anlagen; LPM, lateral plate mesoderm; ltd, latissimus dorsi; PA1-6, pharyngeal arches 1–6; S3, somite 3; scp, scapular muscles; stm, sternocleidomastoid; stp, spinotrapezius. Scale bars: in A for A, B 500 µm; in C for C-D 2000 µm, for E 500 µm; in F’’ for F-F’’ 20 µm.
We first investigated the distribution of neck muscles and NCCs using Myf5nlacZ/+, Mef2c-AHFCre;R26R, Pax3Cre;R26R and Wnt1Cre;R26R embryos (Figure 3—figure supplement 1). At E10.5, the cucullaris anlage was positioned at the level of posterior pharyngeal arches where Wnt1-derived-positive cells were detectable (Figure 1A–C, Figure 1—figure supplement 1A’, Figure 3—figure supplement 1A–B). Subsequently, the cucullaris progenitors expanded caudally from E11.5 to E13.5. The posterior limit of the cranial NCC domain also extended posteriorly; however, the Wnt1-labeled cells did not cover the posterior portion of cucullaris-derived muscles (Figure 3—figure supplement 1C–H). At E14.5, the acromiotrapezius and spinotrapezius attained their definitive position in Myf5nlacZ/+ and Mef2c-AHFCre;R26R embryos (Figure 3—figure supplement 1I–J). Wnt1-derived cells were observed in the anterior acromiotrapezius muscle, but not in the spinotrapezius that was situated in a Pax3-derived domain (Figure 3—figure supplement 1K–L). Analysis of whole-mount embryos indicated that the cranial NCCs did not contribute to connective tissue of posterior cucullaris derivatives, in contrast to what was reported previously (Matsuoka et al., 2005).
To further analyze NCC contribution to the cervical region at the cellular level, we performed immunostainings on sections for Tomato and Tnnt3 in E18.5 Wnt1Cre;R26tdTomato fetuses (Figure 3, Figure 3—figure supplement 2). Given that the Wnt1 lineage is a source of both neuronal and connective tissue derivatives, we associated Tomato immunostaining with Tuj1 that marks neuronal cells and with Tcf4 that labels MCT fibroblasts (Figure 3, Figure 3—figure supplements 2–3). At the cranial level, the MCT of branchiomeric (masseter, mylohyoid), tongue and acromiotrapezius muscles was derived from Wnt1- and Pax3-lineages but not from the mesodermal Mesp1 lineage (Figure 3—figure supplement 2A–B’, Figure 3—figure supplement 3A,F, Figure 3—figure supplement 4A–D,G). The acromiotrapezius showed a high contribution from Wnt1-derived cells while the underlying epaxial muscles had considerably less labeled cells that were limited to the neuronal Tuj1-positive population (Figure 3A–A’). The Wnt1 lineage gave rise to Tcf4-positive fibroblasts in the acromiotrapezius, but not in epaxial neck muscles, where fibroblasts were derived from the Mesp1 lineage (Figure 3—figure supplements 3B–C and 4E). These observations are in accordance with a NCC origin of branchiomeric, anterior trapezius and tongue connective tissue as reported previously (Matsuoka et al., 2005).
However, the NCC contribution to connective tissue in the sternocleidomastoid subset of cucullaris-derived muscles appeared more heterogeneous than that observed in the acromiotrapezius. In rodents, the sternocleidomastoid is composed of three individual muscles (cleidomastoid, sternomastoid and cleido-occipitalis portions); a differential NCC contribution to MCT was observed in these muscles. While Wnt1-derived NCCs were widely present in the sternomastoid and cleido-occipitalis, the NCC contribution was limited in the cleidomastoid (Figure 3B–B’). Indeed, Tcf4-positive fibroblasts in the cleido-occipitalis originated from the Wnt1 lineage, whereas the majority of MCT fibroblasts in the cleidomastoid were derived from the Mesp1 lineage (Figure 3—figure supplements 3D–E and 4F).
A differential contribution of NCCs to connective tissue was also seen within the laryngeal and infrahyoid musculature. Extensive Wnt1 lineage contributions to MCT was observed in laryngeal muscles (thyroarytenoid and cricothyroid) that connect to the thyroid cartilage, which is of NCC origin (Figure 3C–C’) (Tabler et al., 2017). In contrast, the laryngeal muscles (cricoarytenoid and vocal muscles) that link mesoderm-derived laryngeal cartilages (cricoid, arytenoid and medio-caudal portion of the thyroid) did not contain NCC-derived connective tissue (Figures 2G–H and 3C–C’) (Tabler et al., 2017). In these muscles, the Wnt1-derived cells were neuronal, as observed in the esophagus, whereas the MCT fibroblasts were derived from the Mesp1 lineage (Figure 3C–C’, Figure 3—figure supplements 2D–D’ and 4H). As another example, Wnt1-derived cells contributed to a greater extent to MCT in infrahyoid muscles (thyrohyoid muscles) that connect the hyoid and thyroid cartilage that are of NCC origin, compared to infrahyoid muscles (omohyoid and sternohyoid muscles) that link posteriorly pectoral structures of mesodermal origin (Figure 3—figure supplement 2C,C’, H; Figure 3—figure supplement 3G–H). These observations suggest that MCT composition within laryngeal and infrahyoid muscles correlates in part with the embryonic origin of the skeletal components to which they attach (Figure 2G–H, Figure 3C–C’, Figure 3—figure supplement 2C–C’, H).
Given our findings that connective tissues of neck muscles have differential contributions of NCC and mesodermal populations, we analyzed the caudal connections of the cucullaris-derived muscles to the pectoral girdle (Figure 3D, Figure 3—figure supplement 2E–H). The acromiotrapezius attaches dorsally to the nuchal ligament and ventrally to the scapular acromion process in continuity with the scapular spine. While Wnt1-derived cells were present dorsally (Figure 3A, Figure 3—figure supplement 2E), this contribution diminished gradually and was undetectable at the insertion on the scapula (Figure 3D–D’, Figure 3—figure supplement 2F). Similarly, the sternocleidomastoid muscle showed limited NCC contribution to the attachment sites of the clavicle and sternum (Figure 3—figure supplement 2G–H). In contrast to what was previously described (Matsuoka et al., 2005), we did not observe NCC contribution to the shoulder endochondral tissue nor to the nuchal ligament (Figure 3—figure supplement 2E–H). Taken together, these observations define a novel boundary for neural crest contribution to neck/pectoral components. The posterior contribution limit of neural crest to branchiomeric MCT occurs at the level of laryngeal muscles that connect to NCC skeletal derivatives. Moreover, NCCs do not participate in connecting posterior cucullaris and infrahyoid muscles to their skeletal elements.
To assess the cellular origin of cucullaris connective tissue at posterior attachment sites, we next traced the contribution of lateral plate mesoderm (LPM) to the neck/shoulder region using Prx1Cre reporter mice (Durland et al., 2008; Logan et al., 2002) (Figure 4, Figure 4—figure supplements 1–2). Analysis of E9.5 embryos showed that Prx1-derived cells contribute to the forelimb bud and cells adjacent to the anterior-most somites, but not to pharyngeal arches (Figure 4A). At E12.5, the postcranial Prx1-derived domain clearly defined the lateral somitic frontier along the rostrocaudal axis (Durland et al., 2008) and did not include the cucullaris anlage (Figure 4—figure supplement 1, white arrowheads). Whole-mount immunostainings for the myogenic markers Pax7/Myod/My32 and for GFP in Prx1Cre;R26mTmG embryos showed that Prx1-derived cells were present in the dorsal part of the cucullaris but did not contribute to myofibres (Figure 4B, white arrowheads). At E18.5, the Prx1 lineage marked the limb, scapular and abdominal regions, whereas only a few Prx1-derived cells were detected in the cucullaris-derived sternocleidomastoid, acromiotrapezius and spinotrapezius muscles (Figure 4C–D). On sections, immunostaining for β-gal and Tnnt3 showed that Prx1-derived LPM contributed to limb/shoulder MCT and to skeletal components of the pectoral girdle (Figure 4E, Figure 4—figure supplement 2A–B). In contrast, fewer Prx1-derived cells were detected in the acromiotrapezius and little or no contribution was observed in the epaxial muscles (Figure 4E, Figure 4—figure supplement 2B–C). In addition, only a limited number of Prx1-derived cells gave rise to Tcf4-positive fibroblasts in the trapezius muscles, but they contributed more extensively to the fibroblast population in scapular muscles (Figure 4F–F’’, white arrowheads, Figure 4—figure supplement 2D–D”). Notably, β-gal expression for this lineage was not detected in trapezius myofibres thereby confirming the results obtained at E12.5 (Figure 4B–F”, Figure 4—figure supplements 1–2).
Therefore, these observations reveal a dual NCC/LPM origin of trapezius connective tissue, with a decrease of NCC contribution at posterior attachment sites. Moreover, our analysis shows that the postcranial LPM does not give rise to cucullaris myofibres in contrast to what was suggested previously (Theis et al., 2010), thus providing further evidence for a branchiomeric origin of the cucullaris.
Divergent functions of Tbx1 and Pax3 in neck development
Given the key role for Tbx1 and Pax3 genes in the specification of the CPM and somites respectively, we analyzed the effect of inactivation of these genes on neck muscle formation, compared to the muscle phenotypes observed at cranial and trunk levels.
Analysis has been performed by immunostainings on sections and 3D reconstructions of the neck and pectoral girdle using high-resolution micro-computed tomographic (µCT) scans of control, Tbx1-/- and Pax3-/- fetuses (Figures 5–6).
Figure 5
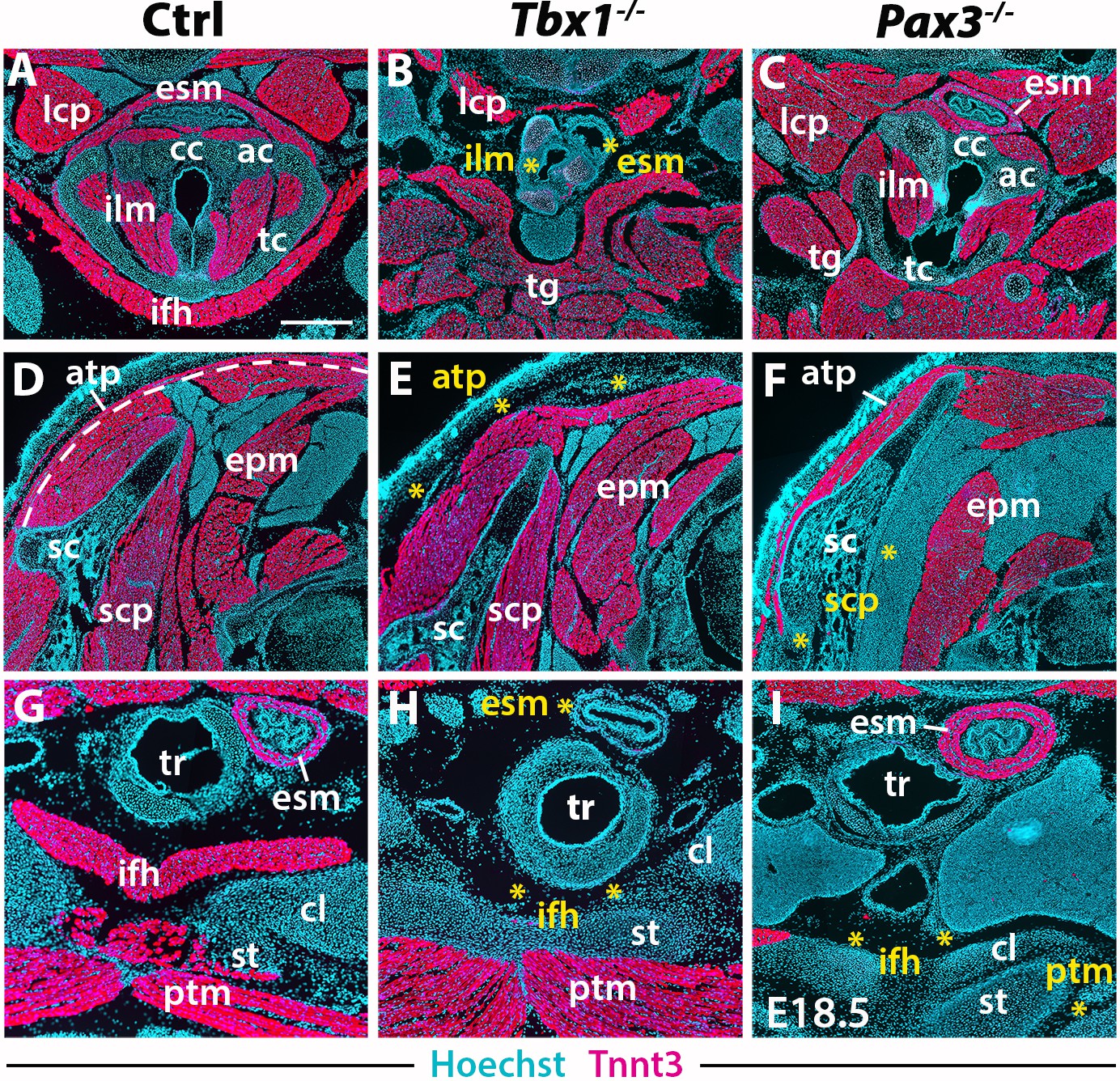
Neck muscle phenotypes in Tbx1 and Pax3 mutants.
(A–I) Immunostainings for Tnnt3 on coronal cryosections of control, Tbx1-null and Pax3-null fetuses at E18.5 (n = 3 each condition). Yellow asterisks indicate missing muscles. Note absence of branchiomeric laryngeal (ilm), esophagus (esm) and trapezius (atp) muscles and severe alteration of somitic infrahyoid muscles (ifh) in Tbx1 mutants. Scapular (scp) and pectoral (ptm) muscles are missing in Pax3 mutants. ac, arytenoid cartilage; atp, acromiotrapezius; cc, cricoid cartilage; cl, clavicle; epm, epaxial musculature; esm, esophagus striated muscle; ifh, infrahyoid muscles; ilm, intrinsic laryngeal muscles; lcp, longus capitis; ptm, pectoralis muscles; sc, scapula; scp, scapular muscles; st, sternum; tc, thyroid cartilage; tg, tongue. Scale bars: in A for A-I 500 µm.
Figure 6

3D reconstructions of neck musculoskeletal system in Tbx1 and Pax3 mutants.
See interactive 3D PDFs in Supplementary file 1–3; control n = 1; mutants n = 2. (A–C) Branchiomeric and cucullaris-derived muscles marked by Mef2c-AHF/Islet1/Mesp1 lineages are indicated in pink. (D–F) Anterior somitic muscles (Mesp1, Pax3 lineages), in red. (G–I) Scapular muscles from more posterior somites (Pax3 lineage), in violet. atp, acromiotrapezius; cc, cricoid cartilage; dg, digastric muscles; epm, epaxial musculature; ifh, infrahyoid muscles; ilm, intrinsic laryngeal muscles; lcl, longus colli; lcp, longus capitis; myh, mylohyoid; sc, scapula; scp, scapular muscles; stm, sternocleidomastoid; tc, thyroid cartilage; tg, tongue.
In the early embryo, Tbx1 is expressed in pharyngeal mesoderm and is required for proper branchiomeric muscle formation (Grifone et al., 2008; Kelly et al., 2004). While Tbx1 is expressed in other cranial populations including the pharyngeal ectoderm and endoderm (Arnold et al., 2006; Huynh et al., 2007), the gene is known to be required cell autonomously during CPM myogenesis (Kong et al., 2014; Zhang et al., 2006). Analysis of Tbx1 mutants revealed unexpected features in cucullaris and hypaxial neck muscle formation. As previously described (Gopalakrishnan et al., 2015; Kelly et al., 2004), anterior branchiomeric muscles (digastric and mylohyoid) showed phenotypic variations, whereas posterior branchiomeric muscles (esophagus and intrinsic laryngeal muscles) and the acromiotrapezius were severely affected or undetectable (Figure 5B,E,H; Figure 6B) (Table 2). However, detailed examination of the cucullaris-derived muscles revealed a heterogeneous dependence on Tbx1 function that was not reported previously (Lescroart et al., 2015; Theis et al., 2010). Unexpectedly, the sternocleidomastoid muscle was present bilaterally but smaller (Figure 6B); the different portions (cleido-occipitalis, cleidomastoid and sternomastoid) were unilaterally or bilaterally affected in a stochastic manner. Moreover, while the epaxial neck and scapular muscles were unaffected (Figure 5E, Figure 6E–H), the hypaxial neck muscles derived from anterior somites were altered. Indeed, the tongue and longus capitis were reduced and the infrahyoid and longus colli muscles were severely affected or undetectable (Figure 5B,H, Figure 6E,H; see interactive 3D PDFs in Supplementary file 1–2).
Table 2
Summary of the neck muscle phenotype observed in Tbx1- and Pax3-null fetuses.
https://doi.org/10.7554/eLife.40179.020| Tbx1-null | Pax3-null | |
|---|---|---|
| Branchiomeric muscles (Mef2c-AHF/Islet1/Mesp1 lineage) Mylohyoid Digastric muscles Intrinsic laryngeal muscles Esophagus striated muscle Sternocleidomastoid Acromiotrapezius | +/- +/- − − +/- − | ++ ++ + ++ + + |
| Anterior-most somite muscles (Mesp1/Pax3 lineage) Epaxial musculature Longus capitis Longus colli Infrahyoid muscles Tongue muscles* | ++ +/- − − + | + ++ ++ +/- + |
| More posterior somite muscles (Pax3 lineage) Scapular muscles Pectoralis | ++ ++ | − − |
-
++,normal; +, altered morphology; +/-, affected; -, severely affected or undetectable
*Including intrinsic and extrinsic tongue muscles of somitic origin
Analysis of Pax3 mutants showed that the neck and pectoral muscles were differentially affected. As expected, branchiomeric and epaxial muscles developed normally but displayed morphological differences adapted to malformations noted in some skeletal components (Figure 5C,F; Figure 6C,I). However, whereas hypaxial trunk/limb muscles were severely affected or undetectable in Pax3 mutants (Figure 5F,I; Figure 6F,I) (Tajbakhsh et al., 1997; Tremblay et al., 1998), surprisingly the majority of hypaxial neck muscles derived from both Mesp1 and Pax3 lineages were present. Tongue muscles were reduced in size but patterned, the infrahyoid were hypoplastic, whereas the longus capitis and longus colli were unaffected (Figure 5C; Figure 6F,I; see interactive 3D PDF in Supplementary file 3). The phenotypes of the different muscle groups observed in Tbx1 and Pax3 mutants are summarized in Table 2 (see also Figure 7—figure supplement 1).
Taken together, these observations revealed that hypaxial muscles in the neck were less affected in Pax3 mutants than more posterior hypaxial muscles, pointing to distinct requirements for Pax3 function during neck and trunk muscle formation. In addition, Tbx1 mutants exhibited more severe phenotypes in hypaxial neck muscles, thus highlighting distinct roles for this gene in branchiomeric and hypaxial neck myogenesis.
Discussion
The embryological origins of neck muscles and connective tissues at the head-trunk interface have been poorly defined largely due to their localization at a transition zone that involves multiple embryonic populations. Using a combination of complementary genetically modified mice and 3D analysis that identifies muscles in the context of their bone attachments, we provide a detailed map of neck tissue morphogenesis and reveal some unexpected features regarding the muscle and connective tissue network.
Branchiomeric origin of cucullaris-derived muscles
The mammalian neck consists of somitic epaxial/hypaxial muscles, branchiomeric muscles and cucullaris-derived muscles (Table 1). The latter constitute a major innovation in vertebrate history, connecting the head to the pectoral girdle in gnathostomes and allowing head mobility in tetrapods (Ericsson et al., 2013). Recent studies in different organisms including shark, lungfish and amphibians suggest that the cucullaris develops in series with posterior branchial muscles and that its developmental origin and innervation is conserved among gnathostomes (Diogo, 2010; Ericsson et al., 2013; Naumann et al., 2017; Noda et al., 2017; Sefton et al., 2016; Tada and Kuratani, 2015; Ziermann et al., 2018a; Ziermann et al., 2017). However, multiple embryological origins including CPM, LPM and somites have been reported for the cucullaris, underscoring the difficulty in deciphering the morphogenesis of this and other muscles in the head-trunk transition zone (Huang et al., 2000; Nagashima et al., 2016; Sefton et al., 2016; Theis et al., 2010).
Our study shows that the cucullaris anlage is innervated by the accessory nerve XI and develops contiguously with the mesodermal core of posterior arches and anterior-most somites 1–3. Our lineage analysis reveals that cucullaris development depends on a branchiomeric myogenic program involving Mef2c-AHF, Islet1 and Mesp1 lineages in keeping with previous results (Table 1) (Lescroart et al., 2015; Sefton et al., 2016; Theis et al., 2010). However, our detailed functional analysis and 3D reconstructions lead us to modify the view of the genetic requirements of cucullaris-derived muscles (Lescroart et al., 2015; Theis et al., 2010). Notably, these muscles are differentially affected in Tbx1-null fetuses; the acromiotrapezius does not form while the sternocleidomastoid is present but reduced. Therefore, Tbx1 is differentially required for sternocleidomastoid and trapezius formation, suggesting that distinct subprograms regulate cucullaris development.
We also demonstrate that the cucullaris anlage is excluded from the postcranial Prx1-derived expression domain, which delineates the trunk LPM field (Figure 4). The Prx1 lineage instead gives rise to connective tissue, thereby excluding a contribution from LPM to cucullaris-derived myofibres. Thus, our results, combined with innervation studies, retrospective clonal analyses and grafting experiments in chick and axolotl (Lescroart et al., 2015; Nagashima et al., 2016; Sefton et al., 2016; Tada and Kuratani, 2015), suggest that the postcranial extension of the CPM lateral to the first three somites in tetrapod embryos is a source of cucullaris myogenic cells (Figure 7A). The discordance with previous studies regarding the origin of the cucullaris is likely due to its proximity to both anterior somites and LPM (Figure 7A–B), and consequently, to potential contamination of embryonic sources in grafting experiments (Couly et al., 1993; Huang et al., 1997; Huang et al., 2000; Noden, 1983; Piekarski and Olsson, 2007; Theis et al., 2010).
Figure 7 with 1 supplement see all
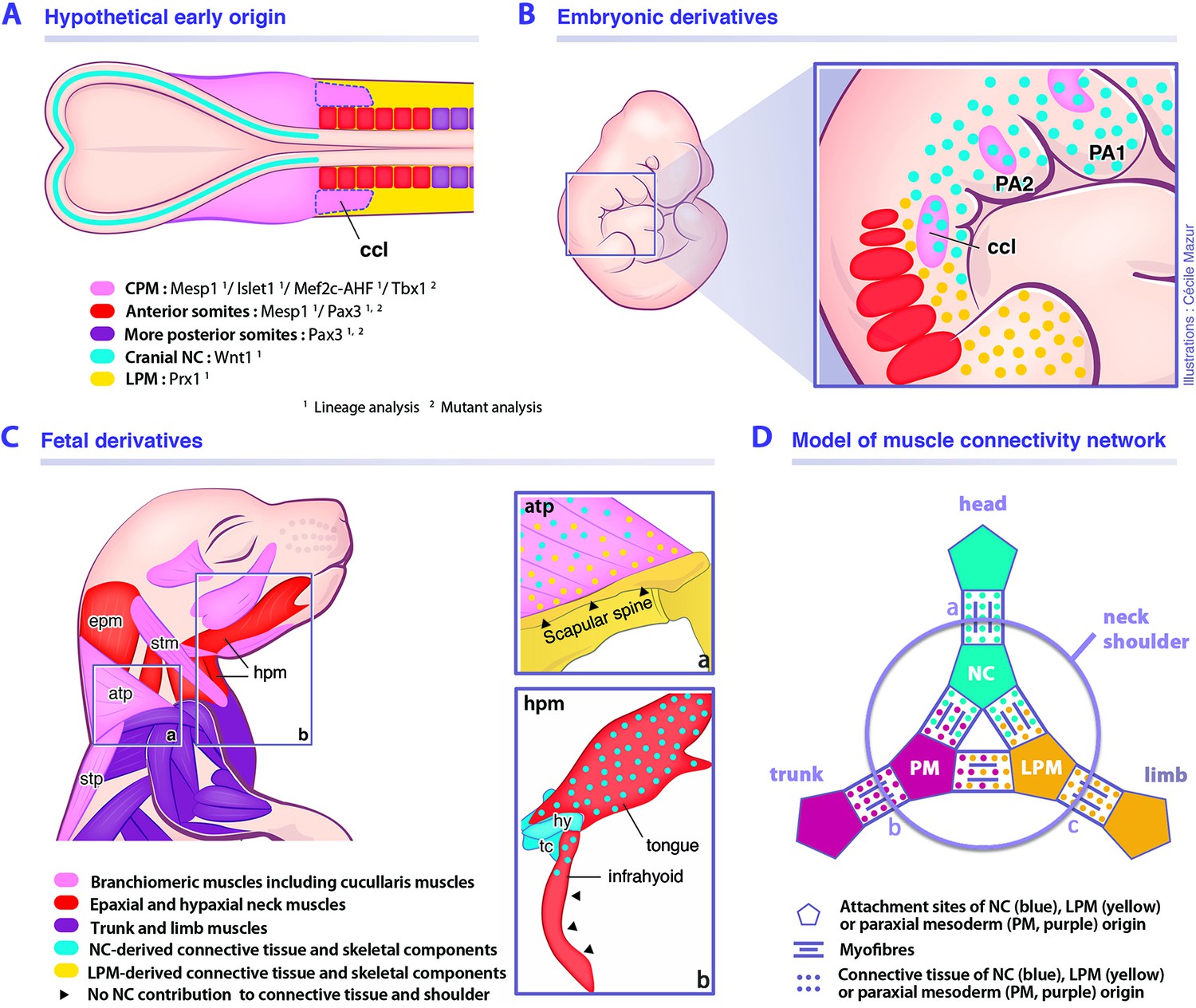
Model for musculoskeletal and connective tissue relationships during murine neck development.
See also Figure 7—figure supplement 1. (A, C) CPM (pink), anterior somites (red) and more posterior somites (violet) muscles are defined by three distinct myogenic programs. (B) Note that the cucullaris develops in a NC domain (blue dots), but is excluded from the postcranial LPM (yellow dots). (C) Dual NC/LPM origin of trapezius connective tissue is indicated in (a). NC contribution to connective tissue extends to tongue and anterior infrahyoid musculature (b). (D) Mixed origins of muscle connective tissues at the head-trunk-limb interface. Example of representative muscles: (a) masseter, (b) spinalis dorsi, (c) deltoid. atp, acromiotrapezius; ccl, cucullaris; CPM, cardiopharyngeal mesoderm; epm, epaxial neck musculature; hpm, hypaxial neck musculature; hy, hyoid bone; LPM, postcranial lateral plate mesoderm; NC, neural crest; PA1-2, pharyngeal arches 1–2; PM, paraxial mesoderm; stm, sternocleidomastoid; stp, spinotrapezius; tc, thyroid cartilage.
A unique genetic program for somite-derived neck muscles
Our study also points to a unique Mesp1/Pax3 genetic program in anterior somites for epaxial/hypaxial neck muscle formation (Table 1). While it was shown that the Mesp1 lineage gives rise to tongue muscles (Harel et al., 2009), we demonstrate that it also contributes to all neck muscles. In chordates, Mesp genes appear to play a conserved role in cardiogenesis and axis segmentation. In mouse, Mesp1 inactivation causes early embryonic death from abnormal heart development, and Mesp1/Mesp2 double-knockout embryos lack non-axial mesoderm (Moreno et al., 2008; Saga, 1998; Saga et al., 2000; Satou et al., 2004; Sawada et al., 2000). During early murine development, Mesp1 shows two waves of activation; initially in the nascent mesoderm destined for extraembryonic, cranial and cardiac mesoderm at the onset of gastrulation; later during somitogenesis, transient Mesp1 expression is limited to anterior presomitic mesoderm (Saga, 1998; Saga et al., 1996; Saga et al., 2000; Saga et al., 1999). Our lineage analysis shows that Mesp1 extensively labels the anterior mesoderm, including the CPM and anterior somites 1–6, while contribution decreases in more posterior somites (Figure 1) (Loebel et al., 2012; Saga et al., 2000; Saga et al., 1999). Previous fate mapping experiments have shown that the mesoderm of late-streak stage embryos contributes to both CPM and anterior somites (Parameswaran and Tam, 1995). It appears that the first wave of Mesp1 expression defines not only the CPM field but also includes the mesoderm destined for anterior somites. In contrast, the Mesp1-labeled cells observed in more posterior somites using the sensitive Pax7GPL reporter may result from the transient wave of Mesp1 expression in the presomitic mesoderm during axis segmentation. Furthermore, we show that Mesp1-derived anterior somites give rise to all epaxial/hypaxial neck muscles in contrast to trunk/limb muscles originating from more posterior somites marked by Pax3. The boundary of Mesp1 lineage contribution to muscles corresponds to the neck/pectoral interface. Our findings indicate that the anterior somitic mesoderm employs a specific transition program for neck muscle formation involving both Mesp1 and Pax3 genes implicated in CPM and somitic myogenesis, respectively (Figure 7A–C).
Whereas little is known about the function of Mesp genes in chordates, there is evidence that Mesp1 might be differentially required during anterior versus posterior somitic formation. In mouse, different Mesp1 enhancer activities have been observed between CPM/anterior somites and posterior somites indicating that the regulation of Mesp1 expression might differ in the two embryonic compartments (Haraguchi et al., 2001). In zebrafish, quadruple mutants of Mesp genes (Mesp-aa/-ab/-ba/-bb) lack anterior somite segmentation while the positioning of posterior somite boundaries is unaffected, suggesting distinct requirements for Mesp genes in anterior and posterior somites (Yabe et al., 2016). Interestingly, during early ascidian development, Mesp is expressed in B7.5 founder cells that give rise to both CPM and anterior tail muscles (ATM) (Satou et al., 2004). In Ciona, the CPM precursors at the origin of heart and atrial siphon (pharyngeal) muscles depend on the ascidian homologs of Mesp1, Islet1 and Tbx1 (reviewed in [Diogo et al., 2015]), indicating that a conserved genetic network promotes chordate myogenesis in the anterior embryonic domain.
Our lineage analysis also reveals an unexpected contribution of Islet1-derived cells to the formation of cutaneous maximus and latissimus dorsi muscle progenitors (Table 1) (Prunotto et al., 2004; Tremblay et al., 1998). Islet1 is activated in a subset of CPM progenitors giving rise to branchiomeric muscles and second heart field myocardium (Cai et al., 2003; Harel et al., 2009; Nathan et al., 2008). At the trunk level, while Islet1 is widely expressed in the nervous system and in the LPM forming the hindlimb bud (Cai et al., 2003; Yang et al., 2006), to our knowledge its expression in somitic myogenic cells has not been reported. The cutaneous maximus and latissimus dorsi muscles are missing in both Pax3 and Met mutants (Prunotto et al., 2004; Tajbakhsh et al., 1997; Tremblay et al., 1998). Therefore, the formation of the latissimus dorsi and cutaneous maximus muscles depends on a specific developmental program implicating Pax3, Islet1 and Met genes. Given that the latissimus dorsi and cutaneous maximus participated in the gain in mobility of the forelimbs towards the shoulder girdle in tetrapods, our findings provide insights into their genetic and evolutionary origins.
Our detailed analysis of Tbx1- and Pax3-null mice on sections and in 3D reconstructions now provides a clarified view of neck muscle morphogenesis (Table 2). In both Tbx1 and Pax3 mutants, whereas the epaxial neck musculature is unaffected, the hypaxial muscles originating from anterior somites are perturbed with a more severe phenotype observed in Tbx1 mutants (Table 2). Whereas no Tbx1 expression has been reported in early myotomes in somites, Tbx1 transcripts appear in hypaxial limb and tongue precursors after myogenic specification (Grifone et al., 2008; Kelly et al., 2004; Zoupa et al., 2006). Tbx1-null embryos show normal myotomal and limb muscle morphology while the hypoglossal cord is hypoplastic, resulting in reduced tongue musculature (Table 2) (Grifone et al., 2008; Kelly et al., 2004). Therefore, we cannot exclude the possibility that Tbx1 is activated and plays a role after specification of neck hypaxial muscles (Okano et al., 2008; Zoupa et al., 2006). The hypaxial muscle defects might also be secondary to a failure of caudal pharyngeal outgrowth (Kelly et al., 2004). While Tbx1 acts cell autonomously in mesodermal progenitors (Kong et al., 2014; Zhang et al., 2006), its expression in pharyngeal endoderm might imply an indirect role in CPM myogenesis (Arnold et al., 2006). Defects in signaling from pharyngeal endoderm may explain the hypoglossal cord deficiency and the potential non-autonomous role for Tbx1 in neck hypaxial myogenesis. Detailed analysis of muscle formation in conditional Tbx1 mutants is needed to resolve the relative roles of Tbx1 in neck myogenesis.
It has been shown that hypaxial muscles are perturbed to a greater extent than epaxial muscles in Pax3 mutants (Tajbakhsh et al., 1997; Tremblay et al., 1998), suggesting a different requirement for Pax3 in these muscle groups, possibly through differential gene regulation (Brown et al., 2005). An unexpected outcome of our analysis was that hypaxial neck muscles (derived from Mesp1 and Pax3 lineages) are less perturbed in Pax3-null mutants than hypaxial trunk/limb muscles (Pax3 lineage only) that are severely altered or undetectable (Table 2). Our results indicate that Pax3 is not essential for the formation of neck muscles derived from anterior somites in contrast to hypaxial muscles originating from more posterior somites. These observations support our model that a distinct genetic program governs somitic neck muscles compared to more posterior trunk muscles.
Connectivity network of the neck and shoulders
Assessing the non-muscle contribution to the neck region is a major challenge due to the extensive participation of diverse cell types from different embryological origins. Previous studies in amphibians, chick and mouse reported that branchiomeric and hypobranchial connective tissue originates from NCCs (Hanken and Gross, 2005; Köntges and Lumsden, 1996; Matsuoka et al., 2005; Noden, 1983; Olsson et al., 2001; Ziermann et al., 2018b). It has been shown that the neural crest provides connective tissue for muscles that link the head and shoulders, whereas mesodermal cells give rise to connective tissue for muscles connecting the trunk and limbs (Matsuoka et al., 2005).
Our findings demonstrate that not all branchiomeric muscles are composed of neural crest-derived connective tissue, thereby redefining a new limit for NCC contribution to the neck and shoulders. Unexpectedly, we noted that the contribution of the neural crest lineage is limited in infrahyoid and posterior branchiomeric muscles that connect skeletal components of mesodermal origin. Indeed, it appears that the connective tissue of muscles that link exclusively mesodermal skeletal derivatives is of mesodermal origin. In contrast, the connective tissue of cucullaris-derived muscles is of a mixed origin, first developing in a cranial NCC domain at early stages, then expanding to incorporate connective tissue from both neural crest and LPM populations (Figure 7B). While NCCs are present in the anterior acromiotrapezius, sternocleidomastoid and infrahyoid muscles, contribution gradually decreases at posterior attachment sites and is undetectable at scapular level. In parallel, the LPM gives rise to shoulder skeletal components and to connective tissue at the attachment sites of associated musculature including trapezius muscles (Figure 7C). Therefore, the dual NCC/LPM origin of the trapezius connective tissue correlates with the embryonic origin of skeletal components to which it is connected.
Wnt1Cre and Sox10Cre NCC reporter mice were used to show that endochondral cells connecting the cucullaris-derived muscles on the scapula, clavicle and sternum share a common NCC origin with the connective tissue (Matsuoka et al., 2005). However, NCCs are not found in pectoral components of fish, axolotl and chick, while contribution to neurocranium is conserved, suggesting that NCC involvement in shoulder formation would be specific to mammals (Epperlein et al., 2012; Kague et al., 2012; Piekarski et al., 2014; Ponomartsev et al., 2017). In contrast to this view, our lineage analysis reveals that the neural crest lineage shows limited contribution to cucullaris connective tissue and does not form endochondral cells at the posterior attachment sites (Figure 7C). Differences in genetic lineage tracers and reagents might explain these discordant results (Matsuoka et al., 2005).
Taken together, our findings indicate that the gradient of neural crest and mesodermal contributions to neck connective tissue depends on the embryonic source of attachment sites. Therefore, it reveals that connective tissue composition in the neck region correlates with the cellular origin of associated skeletal components, independently of the myogenic source or ossification mode, forming a strong link between muscles and bones of the head, trunk and limb fields (Figure 7D).
Evolutionary and clinical perspectives
Our findings demonstrate that the hybrid origin of the skeletal, connective tissue and muscle components of the neck is defined during early embryogenesis. The close proximity of neural crest, CPM, LPM and somitic populations is unique along the body plan and underscores the difficulty in defining their relative contributions to structures in the neck (Figure 7A–B). Our results refine the relative contributions of the neural crest and mesodermal derivatives in mouse, thereby providing a coherent view of embryonic components at the head-trunk interface in gnathostomes. Our study highlights the limited NCC contribution to posterior branchiomeric and infrahyoid muscle connective tissue, that is instead of mesodermal origin. This reinforces recent notions suggesting that the cranial NCCs and the postcranial rearrangement of mesodermal populations at the head-trunk interface had been central for the establishment of the neck during gnathostome evolution (Adachi et al., 2018; Kuratani et al., 2018; Lours-Calet et al., 2014; Nagashima et al., 2016; Sefton et al., 2016). The contribution of anterior mesoderm in the origin of the neck needs to be elucidated in future studies of gnathostomes.
Our study reveals that neck muscles develop in a complex domain that is distinct from the head and trunk (Figure 7A–D), and that might be a contributing factor to pathologies that affect subsets of neck muscles in specific myopathies (Emery, 2002; Randolph and Pavlath, 2015). In human, TBX1 has been identified as a major candidate gene for 22q11.2 deletion syndrome (Papangeli and Scambler, 2013). Laryngeal malformations, esophageal dysmotility and shortened neck are frequent in patients. Moreover, the neck deficiencies might not be exclusively due to cervical spine abnormalities but also to neck muscle defects (Hamidi et al., 2014; Leopold et al., 2012; Marom et al., 2012). Therefore, our analysis of Tbx1-null mutants provides a better understanding of the etiology of the 22q11.2 deletion syndrome and has direct implications in establishing clinical diagnosis in cases where patients present failure in neck-associated functions.
Materials and methods
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Strain, strain background (Mus musculus) | B6D2F1/JRj | Janvier | ||
| Genetic reagent (M. musculus) | Mef2c-AHFCre | PMID:16188249 | MGI:3639735 | Dr. Brian L Black (Cardiovascular Research Institute, University of California, USA) |
| Genetic reagent (M. musculus) | Islet1Cre | PMID:11299042 | MGI:2447758 | Dr. Thomas M Jessell (Howard Hughes Medical Institute, Columbia University, USA) |
| Genetic reagent (M. musculus) | Mesp1Cre | PMID:10393122 | MGI:2176467 | Pr. Yumiko Saga (National Institute of Genetics, Japan) |
| Genetic reagent (M. musculus) | Pax3Cre | PMID:22394517 | MGI:3573783 | Dr. Jonathan A. Epstein (Perelman Shool of Medicine, University of Pennsylvania, USA) |
| Genetic reagent (M. musculus) | Myf5Cre | PMID:17418413 | MGI:3710099 | Dr. Mario R Capecchi (Institute of Human Genetics, University of Utah, USA) |
| Genetic reagent (M. musculus) | Wnt1Cre | PMID:9843687 | MGI:J:69326 | Pr. Andrew P. McMahon (Keck School of Medicine of the University of Southern California, USA) |
| Genetic reagent (M. musculus) | Prx1Cre | PMID:12112875 | MGI: J:77872 | Dr. Clifford J Tabin (Department of genetics, Harvard Medical School, USA) |
| Genetic reagent (M. musculus) | Pax7GPL | PMID:19531352 | MGI:3850147 | Dr. Shahragim Tajbakhsh (Department of Developmental and Stem Cell Biology, Institut Pasteur, France) |
| Genetic reagent (M. musculus) | Rosa26R-lacZ | PMID:9916792 | MGI:1861932 | Pr. Philippe Soriano (Icahn School of Medicine at Mt. Sinai, USA) |
| Genetic reagent (M. musculus) | R26mTmG | PMID:17868096 | MGI:3716464 | Pr. Philippe Soriano (Icahn School of Medicine at Mt. Sinai, USA) |
| Genetic reagent (M. musculus) | R26tdTomato | PMID:20023653 | MGI:3809524 | Dr. Hongkui Zeng (Allen Institute for Brain Science, USA) |
| Genetic reagent (M. musculus) | Myf5nlacZ/+ | PMID:8918877 | MGI:1857973 | Dr. Shahragim Tajbakhsh (Department of Developmental and Stem Cell Biology, Institut Pasteur, France) |
| Genetic reagent (M. musculus) | Tbx1-null | PMID:11242110 | MGI:2179190 | Dr. Virginia Papaioannou (Department of Genetics and Development, Columbia University Medical Center, USA) |
| Antibody | Chicken polyclonal anti-β-gal | Abcam | Cat. #: ab9361 | IF (1:1000) |
| Antibody | Rabbit polyclonal anti-β-gal | MP Biomedicals | Cat. #: MP 559761 | IF (1:750) |
| Antibody | Chicken polyclonal anti-GFP | Aves Labs | Cat. #: 1020 | IF (1:500) |
| Antibody | Chicken polyclonal anti-GFP | Abcam | Cat. #: 13970 | IF (1:1000) |
| Antibody | Mouse monoclonal IgG1 anti-Islet1 | DSHB | Cat. #: 40.2D6 | IF (1:1000) |
| Antibody | Mouse monoclonal IgG1 anti-My32 | Sigma | Cat. #: M4276 | IF (1:400) |
| Antibody | Mouse monoclonal IgG1 anti-Myod | Dako | Cat. #: M3512 | IF (1:100) |
| Antibody | Mouse monoclonal IgG1 anti-Pax7 | DSHB | Cat. #: AB_528428 | IF (1:20) |
| Antibody | Rabbit polyclonal anti-Tcf4 | Cell Signalling | Cat. #: C48H11 | IF (1:150) |
| Antibody | Mouse monoclonal IgG1 anti-Tnnt3 | Sigma | Cat. #: T6277 | IF (1:200) |
| Antibody | Rabbit polyclonal anti-Tomato | Clontech | Cat. #: 632496 | IF (1:500) |
| Antibody | Mouse monoclonal IgG2a anti-Pax7 | Ozyme | Cat. #: BLE801202 | IF (1:1000) |
| Software, algorithm | GE phoenix datos|x 2.0 | GE Sensing and Inspection Technologies GmbH | ||
| Software, algorithm | 3D PDF maker | SolidWorks Corporation | ||
| Software, algorithm | Zen | Zeiss | ||
| Chemical compound, drug | X-gal | Fisher | Cat. #: 10554973 | |
| Chemical compound, drug | paraformaldehyde | Electron Microscopy Sciences | Cat. #: 15710 | |
| Chemical compound, drug | Triton X-100 | Sigma | Cat. #: T8787 | |
| Chemical compound, drug | Tween 20 | Sigma | Cat. #: P1379 | |
| Chemical compound, drug | Histoclear II | National Diagnostics | Cat. #: HS-202 |
Animals
Animals were handled as per European Community guidelines and the ethics committee of the Institut Pasteur (CTEA) approved protocols (APAFIS#6354–20160809 l2028839). Males carrying the Cre driver gene, Mef2c-AHFCre (Verzi et al., 2005), Islet1Cre (Srinivas et al., 2001), Mesp1Cre (Saga et al., 1999), Pax3Cre (Engleka et al., 2005), Myf5Cre (Haldar et al., 2007), Wnt1Cre (Danielian et al., 1998), Prx1Cre (Logan et al., 2002), were crossed to reporter females from previously described lines including Pax7GPL (Sambasivan et al., 2013), Rosa26R-lacZ (R26R) (Soriano, 1999), R26mTmG (Muzumdar et al., 2007) and R26tdTomato (Madisen et al., 2010). Myf5nlacZ/+ KI mice and mice carrying the Tbx1tm1pa allele (referred to as Tbx1-null) were previously described (Jerome and Papaioannou, 2001; Kelly et al., 2004; Tajbakhsh et al., 1996). To generate experimental Pax3-null fetuses, Pax3WT/Cre males and females were intercrossed (Engleka et al., 2005) (n = 5 Tbx1 and Pax3 mutants analysed including n = 2 by µCT scanning). Mice were crossed and maintained on a B6D2F1/JRj background and genotyped by PCR. Mouse embryos and fetuses were collected between E9.5 and E18.5, with noon on the day of the vaginal plug considered as E0.5.
X-gal and immunofluorescence stainings
Request a detailed protocolWhole-mount samples were analysed for beta-galactosidase activity with X-gal (0.6 mg/ml) in 1X PBS buffer (D1408, Sigma, St. Louis, MO) containing 4 mM potassium ferricyanide, 4 mM potassium ferrocyanide, 0.02% NP-40 and 2 mM MgCl2 as previously described (Comai et al., 2014). For immunostaining on cryosections, foetuses were fixed 3 hr in 4% paraformaldehyde (PFA) (15710, Electron Microscopy Sciences, Hatfield, PA) 0.5% Triton X-100 (T8787, Sigma) at 4°C, washed overnight at 4°C in PBS 0.1% Tween 20 (P1379, Sigma), cryopreserved in 30% sucrose in PBS and embedded in OCT for 12–16 µm sectioning with a Leica cryostat (CM3050 S, Leica, Wetzlar, Germany). Cryosections were dried for 30 min and washed in PBS. For immunostaining on paraffin sections, samples were fixed overnight in 4% PFA, dehydrated in graded ethanol series and penetrated with Histoclear II (HS-202, National Diagnostics, Atlanta, GA), embedded in paraffin and oriented in blocks. Paraffin blocks were sectioned at 10–12 µm using a Leica microtome (Reichert-Jung 2035). Sections were then deparaffinized and rehydrated by successive immersions in Histoclear, ethanol and PBS. Samples were then subjected to antigen retrieval with 10 mM Citrate buffer (pH 6.0) using a 2100 Retriever (Aptum Biologics, Rownhams, UK).
Rehydrated sections were blocked for 1 hr in 10% normal goat serum, 3% BSA, 0.5% Triton X-100 in PBS. Primary antibodies were diluted in blocking solution and incubated overnight at 4°C. Primary antibodies included the following: β-gal (1/1000, chicken polyclonal, ab9361, Abcam, Cambridge, UK; 1/750, rabbit polyclonal, MP 559761, MP Biomedicals, Illkirch, France), GFP (1/500, chick polyclonal, 1020, Aves Labs, Tigard, OR; 1/1000, chick polyclonal, 13970, Abcam), Islet1 (1/1000, mouse monoclonal IgG1, 40.2D6, DSHB), My32 (1/400, mouse monoclonal IgG1, M4276, Sigma), Myod (1/100, mouse monoclonal IgG1, M3512, Dako, Santa Clara, CA), Pax7 (1/20, mouse monoclonal IgG1, AB_528428), Tcf4 (1/150, rabbit polyclonal, C48H11, Cell Signalling, Leiden, Netherlands), Tnnt3 (1/200, monoclonal mouse IgG1, T6277, Sigma), Tomato (1/500, rabbit polyclonal, 632496, Clontech, Shiga, Japan; 1/250, chick polyclonal, 600-901-379, Rockland, Pottstown, PA) and Tuj1 (1/1000, monoclonal mouse IgG2a, BLE801202, Ozyme, Montigny-le-Bretonneux, France). After 3 rounds of 15 min washes in PBS 0.1% Tween 20, secondary antibodies were incubated in blocking solution 2 hr at RT together with 1 μg/ml Hoechst 33342 to visualize nuclei. Secondary antibodies consisted of Alexa 488, 555 or 633 goat anti-rabbit, anti-chicken or anti-mouse isotype specific (1/500, Jackson Immunoresearch, Cambridgeshire, UK). After 3 rounds of 15 min washes in PBS 0.1% Tween 20, slides were mounted in 70% glycerol for analysis.
For whole-mount immunofluorescence staining, embryos were dissected in PBS, fixed in 4% PFA, washed in PBS and stored at −20°C in 100% methanol. After rehydration in PBS, whole mount immunostainings were performed incubating the primary and secondary antibodies for 3 days each. Samples were cleared using benzyl alcohol/benzyl benzoate (BABB) clarification method (Yokomizo et al., 2012).
µCT scan analysis
Request a detailed protocolFor µCT scan analysis, the fetuses were treated with the phosphotungstic acid (PTA) contrast agent to well reveal skeletal and muscle structures. After dissection of the cervical region (including the mandible and scapular components, see Figure 2—figure supplement 1), the fetuses were fixed in 4% PFA for 24 hr at 4°C. Samples were then additionally fixed and dehydrated by exchanging the fixative and washing solutions to incrementally increasing ethanol concentrations (30%, 50%, 70%) with 2 days in each concentration to minimize the shrinkage of tissues. To start the contrasting procedure, the embryos were firstly incubated in ethanol-methanol-water mixture (4:4:3) for 1 hr and then transferred for 1 hr into 80% and 90% methanol solution. The staining procedure was then performed for 10 days in 90% methanol 1.5% PTA solution (changed every day with fresh solution) to ensure optimal penetration of the contrast agent. Staining was followed by rehydration of the samples in methanol-grade series (90%, 80%, 70%, 50% and 30%) and stored in sterile distilled water. The samples were placed in polypropylene tubes and embedded in 1% agarose gel to avoid movement artefacts during measurements. μCT scanning was performed using laboratory system GE Phoenix v|tome|x L 240 (GE Sensing and Inspection Technologies GmbH, Hamburg, Germany), equipped with a nanofocus X-ray tube with maximum power of 180 kV/15 W and a flat panel detector DXR250 with 2048 × 2048 pixel2, 200 × 200 µm2 pixel size. The μCT scan was carried out at 60 kV acceleration voltage and 200 µA tube current with voxel size of 5.7 μm for all samples. The beam was filtered by a 0.2 mm aluminium filter. The 2200 projections were taken over 360° with exposure time of 900 ms. The tomographic reconstructions were done using the software GE phoenix datos|x 2.0 (GE Sensing and Inspection Technologies GmbH) and data segmentations and visualizations were performed by combination of software VG Studio MAX 2.2 (Volume GraphicsGmbH, Heidelberg, Germany) and Avizo 7.1 (Thermo Fisher Scientific, Waltham, MA) according to (Tesařová et al., 2016). The interactive 3D PDFs were set up using 3D PDF maker software.
Imaging
Request a detailed protocolImages were acquired using the following systems: a Zeiss Axio-plan equipped with an Apotome, a Zeiss stereo zoom microscope V16 or a Zeiss LSM 700 laser-scanning confocal microscope with ZEN software (Carl Zeiss, Oberkochen, Germany). For whole-mount rendering, acquired Z-stacks were 3D reconstructed using Imaris software. All images were assembled in Adobe Photoshop (Adobe Systems, San Jose, CA).
Data availability
All data generated or analysed during this study are included in the manuscript and supporting files.
References
-
A new view of patterning domains in the vertebrate mesodermDevelopmental Cell 4:159–165.https://doi.org/10.1016/S1534-5807(03)00033-9
-
Molecular and cellular regulation of skeletal myogenesisCurrent Topics in Developmental Biology 110:1–73.https://doi.org/10.1016/B978-0-12-405943-6.00001-4
-
The triple origin of skull in higher vertebrates: a study in quail-chick chimerasDevelopment 117:409–429.
-
BookMuscles of Vertebrates - Comparative Anatomy, Evolution, Homologies and DevelopmentEnfield, New Hampshire: Science Publishers.https://doi.org/10.1201/9781439845622
-
Visualizing the lateral somitic frontier in the Prx1Cre transgenic mouseJournal of Anatomy 212:590–602.https://doi.org/10.1111/j.1469-7580.2008.00879.x
-
Evolution and development of the vertebrate neckJournal of Anatomy 222:67–78.https://doi.org/10.1111/j.1469-7580.2012.01530.x
-
Spatial relations between avian craniofacial neural crest and paraxial mesoderm cellsDevelopmental Dynamics 235:1310–1325.https://doi.org/10.1002/dvdy.20663
-
A Cranial Mesoderm Origin for Esophagus Striated MusclesDevelopmental Cell 34:694–704.https://doi.org/10.1016/j.devcel.2015.07.003
-
Entwicklungsgeschichte des Kopfes und des Blutgefässsystemes von Ceratodus forsteri. II. Die epigenetischen Erwerbungen während der Stadien 39–48. Denkschr Med-Naturwiss Ges JenaDenkschriften Der Medicinisch-NaturwissenschaftlichenGesellschaft Zu Jena 9:935–1492.
-
Properties of branchiomeric and somite-derived muscle development in Tbx1 mutant embryosDevelopmental Dynamics 237:3071–3078.https://doi.org/10.1002/dvdy.21718
-
Cervical spine abnormalities in 22q11.2 deletion syndromeThe Cleft Palate-Craniofacial Journal 51:230–233.https://doi.org/10.1597/12-318
-
Distinct origins and genetic programs of head muscle satellite cellsDevelopmental Cell 16:822–832.https://doi.org/10.1016/j.devcel.2009.05.007
-
The fate of the first avian somiteAnatomy and Embryology 195:435–449.https://doi.org/10.1007/s004290050063
-
DiGeorge syndrome phenotype in mice mutant for the T-box gene, Tbx1Nature Genetics 27:286–291.https://doi.org/10.1038/85845
-
Pax3/Pax7 mark a novel population of primitive myogenic cells during developmentGenes & Development 19:1426–1431.https://doi.org/10.1101/gad.345505
-
The del22q11.2 candidate gene Tbx1 regulates branchiomeric myogenesisHuman Molecular Genetics 13:2829–2840.https://doi.org/10.1093/hmg/ddh304
-
Rhombencephalic neural crest segmentation is preserved throughout craniofacial ontogenyDevelopment 122:3229–3242.
-
Evolutionary developmental studies of cyclostomes and the origin of the vertebrate neckDevelopment, Growth & Differentiation 50:S189–S194.https://doi.org/10.1111/j.1440-169X.2008.00985.x
-
Laryngeal abnormalities are frequent in the 22q11 deletion syndromeInternational Journal of Pediatric Otorhinolaryngology 76:36–40.https://doi.org/10.1016/j.ijporl.2011.09.025
-
Handbuch Der Vergleichenden Anatomie Der WirbeltiereMuskeln des Kopfes: Viscerale Muskulatur, Handbuch Der Vergleichenden Anatomie Der Wirbeltiere, Berlin, Urban & Schwarzenberg.
-
Head and neck manifestations of 22q11.2 deletion syndromesEuropean Archives of Oto-Rhino-Laryngology 269:381–387.https://doi.org/10.1007/s00405-011-1745-1
-
Dropped head syndrome: diagnosis and managementEvidence-Based Spine-Care Journal 2:41–47.https://doi.org/10.1055/s-0030-1267104
-
Non-myogenic Contribution to Muscle Development and Homeostasis: The Role of Connective TissuesFrontiers in Cell and Developmental Biology 5:22.https://doi.org/10.3389/fcell.2017.00022
-
The development of the cucullaris muscle and the branchial musculature in the Longnose Gar, (Lepisosteus osseus, Lepisosteiformes, Actinopterygii) and its implications for the evolution and development of the head/trunk interface in vertebratesEvolution & Development 19:263–276.https://doi.org/10.1111/ede.12239
-
The embryonic origins of avian cephalic and cervical muscles and associated connective tissuesAmerican Journal of Anatomy 168:257–276.https://doi.org/10.1002/aja.1001680302
-
Interactions and fates of avian craniofacial mesenchymeDevelopment 103:121–140.
-
The differentiation and morphogenesis of craniofacial musclesDevelopmental Dynamics 235:1194–1218.https://doi.org/10.1002/dvdy.20697
-
The 22q11 deletion: DiGeorge and velocardiofacial syndromes and the role of TBX1Wiley Interdisciplinary Reviews: Developmental Biology 2:393–403.https://doi.org/10.1002/wdev.75
-
Morphogenesis of the cranial muscles of Amblystoma punctatumJournal of Morphology 63:531–587.https://doi.org/10.1002/jmor.1050630306
-
A muscle stem cell for every muscle: variability of satellite cell biology among different muscle groupsFrontiers in Aging Neuroscience 7:190.https://doi.org/10.3389/fnagi.2015.00190
-
Divergent functions of murine Pax3 and Pax7 in limb muscle developmentGenes & Development 18:1088–1105.https://doi.org/10.1101/gad.301004
-
MesP1: a novel basic helix-loop-helix protein expressed in the nascent mesodermal cells during mouse gastrulationDevelopment 122:2769–2778.
-
Genetic rescue of segmentation defect in MesP2-deficient mice by MesP1 gene replacementMechanisms of Development 75:53–66.https://doi.org/10.1016/S0925-4773(98)00077-X
-
MesP1 is expressed in the heart precursor cells and required for the formation of a single heart tubeDevelopment 126:3437–3447.
-
Mesp1 expression is the earliest sign of cardiovascular developmentTrends in Cardiovascular Medicine 10:345–352.https://doi.org/10.1016/S1050-1738(01)00069-X
-
Embryonic founders of adult muscle stem cells are primed by the determination gene Mrf4Developmental Biology 381:241–255.https://doi.org/10.1016/j.ydbio.2013.04.018
-
The ascidian Mesp gene specifies heart precursor cellsDevelopment 131:2533–2541.https://doi.org/10.1242/dev.01145
-
Zebrafish Mesp family genes, mesp-a and mesp-b are segmentally expressed in the presomitic mesoderm, and Mesp-b confers the anterior identity to the developing somitesDevelopment 127:1691–1702.
-
Generalized lacZ expression with the ROSA26 Cre reporter strainNature Genetics 21:70–71.https://doi.org/10.1038/5007
-
Cre reporter strains produced by targeted insertion of EYFP and ECFP into the ROSA26 locusBMC Developmental Biology 1:4.
-
Isl1Cre reveals a common Bmp pathway in heart and limb developmentDevelopment 133:1575–1585.https://doi.org/10.1242/dev.02322
-
Cephalic muscle development in the Australian lungfish, Neoceratodus forsteriJournal of Morphology 279:494–516.https://doi.org/10.1002/jmor.20784
-
Tbx1 is expressed at multiple sites of epithelial-mesenchymal interaction during early development of the facial complexThe International Journal of Developmental Biology 50:504–510.https://doi.org/10.1387/ijdb.052116mz
Article and author information
Author details
Elizabeth M Sefton

Shahragim Tajbakhsh
Funding
Institut Pasteur
- Eglantine Heude
- Alexandre Grimaldi
- Shahragim Tajbakhsh
Agence Nationale de la Recherche
- Eglantine Heude
- Alexandre Grimaldi
- Shahragim Tajbakhsh
Centre National de la Recherche Scientifique
- Eglantine Heude
- Estelle Jullian
- Noritaka Adachi
- Alexandre Grimaldi
- Robert G Kelly
- Shahragim Tajbakhsh
French Muscular Dystrophy Association
- Eglantine Heude
- Alexandre Grimaldi
- Robert G Kelly
- Shahragim Tajbakhsh
Central European Institute of Technology
- Marketa Tesarova
- Tomas Zikmund
- Jozef Kaiser
March of Dimes Foundation
- Elizabeth M Sefton
- Gabrielle Kardon
National Institutes of Health
- Elizabeth M Sefton
- Gabrielle Kardon
Fondation pour la Recherche Médicale
- Estelle Jullian
- Noritaka Adachi
- Robert G Kelly
Fondation Leducq
- Estelle Jullian
- Noritaka Adachi
- Robert G Kelly
Yamada Science Foundation
- Noritaka Adachi
- Robert G Kelly
Bourses du Gouvernement Français
- Noritaka Adachi
project CEITEC 2020
- Marketa Tesarova
- Tomas Zikmund
- Jozef Kaiser
The Ministry of Education, Youth and Sports of the Czech Republic under the National Sustainability Programme II
- Marketa Tesarova
- Tomas Zikmund
- Jozef Kaiser
CEITEC Nano Research Infrastructure (MEYS CR, 2016–2019)
- Marketa Tesarova
- Tomas Zikmund
- Jozef Kaiser
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Acknowledgements
We thank Drs. Claudio Cortes and Françoise Helmbacher for providing transgenic mice and Mirialys Gallardo for technical assistance. We also thank Cécile Mazur for illustrations.
Ethics
Animal experimentation: Animals were handled as per European Community guidelines and the ethics committee of the Institut Pasteur (CTEA) approved protocols. (APAFIS#6354-20160809l2028839)
Version history
- Received: July 17, 2018
- Accepted: November 17, 2018
- Accepted Manuscript published: November 19, 2018 (version 1)
- Version of Record published: December 28, 2018 (version 2)
Copyright
© 2018, Heude et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 3,213
- views
-
- 598
- downloads
-
- 50
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Unique morphogenetic signatures define mammalian neck muscles and associated connective tissues
eLife 7:e40179.
https://doi.org/10.7554/eLife.40179
Further reading
-
- Developmental Biology
Inhibitory G alpha (GNAI or Gαi) proteins are critical for the polarized morphogenesis of sensory hair cells and for hearing. The extent and nature of their actual contributions remains unclear, however, as previous studies did not investigate all GNAI proteins and included non-physiological approaches. Pertussis toxin can downregulate functionally redundant GNAI1, GNAI2, GNAI3, and GNAO proteins, but may also induce unrelated defects. Here, we directly and systematically determine the role(s) of each individual GNAI protein in mouse auditory hair cells. GNAI2 and GNAI3 are similarly polarized at the hair cell apex with their binding partner G protein signaling modulator 2 (GPSM2), whereas GNAI1 and GNAO are not detected. In Gnai3 mutants, GNAI2 progressively fails to fully occupy the sub-cellular compartments where GNAI3 is missing. In contrast, GNAI3 can fully compensate for the loss of GNAI2 and is essential for hair bundle morphogenesis and auditory function. Simultaneous inactivation of Gnai2 and Gnai3 recapitulates for the first time two distinct types of defects only observed so far with pertussis toxin: (1) a delay or failure of the basal body to migrate off-center in prospective hair cells, and (2) a reversal in the orientation of some hair cell types. We conclude that GNAI proteins are critical for hair cells to break planar symmetry and to orient properly before GNAI2/3 regulate hair bundle morphogenesis with GPSM2.
-
- Computational and Systems Biology
- Developmental Biology
Organisms utilize gene regulatory networks (GRN) to make fate decisions, but the regulatory mechanisms of transcription factors (TF) in GRNs are exceedingly intricate. A longstanding question in this field is how these tangled interactions synergistically contribute to decision-making procedures. To comprehensively understand the role of regulatory logic in cell fate decisions, we constructed a logic-incorporated GRN model and examined its behavior under two distinct driving forces (noise-driven and signal-driven). Under the noise-driven mode, we distilled the relationship among fate bias, regulatory logic, and noise profile. Under the signal-driven mode, we bridged regulatory logic and progression-accuracy trade-off, and uncovered distinctive trajectories of reprogramming influenced by logic motifs. In differentiation, we characterized a special logic-dependent priming stage by the solution landscape. Finally, we applied our findings to decipher three biological instances: hematopoiesis, embryogenesis, and trans-differentiation. Orthogonal to the classical analysis of expression profile, we harnessed noise patterns to construct the GRN corresponding to fate transition. Our work presents a generalizable framework for top-down fate-decision studies and a practical approach to the taxonomy of cell fate decisions.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}