








-
Fruit crops are an essential part of agriculture and fruits contribute greatly to the world economy. Fruits are also rich sources of food and nutrients that are beneficial to human health. As one of the most important fruit crops, grape is widely cultivated worldwide, and its related foodstuffs, such as grape juice, raisins and wine, are favored globally by consumers. For instance, the most widely consumed fruit wine worldwide is grape wine[1]. However, cultivars of wine grape (Vitis vinifera L.) are peculiarly prone to damage by cold stress[2]. Conventional breeding is commonly used to develop cold-resistant grapevines by using cold-hardy species such as Amur grape (V. amurensis) that can survive low temperature −30 to −40 °C[3] as breeding material[4−5]. Though conventional breeding contributes significantly to grapevine improvement, it is accompanied by genetic diversity and loss of fitness, and is also inefficient to obtain multiple desired characteristics simultaneously, as it largely depends on natural allelic variations[6−7]. Moreover, grapevine has a relatively long juvenile stage and breeding is therefore a time-consuming practice. Hence, novel approaches are required to improve the traits or/and production of grapevine more rapidly and efficiently.
The clustered regularly interspaced short palindromic repeat (CRISPR)/CRISPR-associated protein 9 (Cas9) has emerged as a simple but efficient genome editing technology[8] and shown great promise for crop improvement[9−11]. In recent years, the CRISPR toolbox has swiftly expanded, and new approaches such as base editing and prime editing have been developed and applied to crop improvement[10]. CRISPR/Cas9-mediated genome editing in grapevine was first reported in 2016[12−13]. Since then, genome editing research based on CRISPR/Cas9 technology in this species has been boosted. In this review, we describe CRISPR/Cas platforms designed for plant genome editing and summarize research progress on genome editing based on CRISPR/Cas9 systems in grapevine. The challenges and future prospects of applications of CRISPR/Cas technologies in grapevine are also discussed.
-
CRISPR/Cas systems were initially identified in bacteria and archaea function as adaptive immune systems to protect prokaryotes from invading nucleic acid molecules[14]. CRISPR/Cas systems can be classified into two different classes based on effector protein organization[15,16]. The class 1 systems are characterized as multi-protein effectors, whereas the class 2 systems utilize single-protein effectors[15,16]. The class 2 systems can be divided into three types, with the type II system requiring the Cas9 protein as the effector[17,18]. Native Cas9 nuclease has a bi-lobed architecture: a recognition (REC) lobe and a nuclease (NUC) lobe. The REC could bind to the guide RNA (gRNA) and therefore determines the specific function of Cas9, while the NUC contains two nuclease domains, namely RuvC and HNH[19−21]. Once the Cas9-gRNA complex binds to the target sequence through Watson-Crick pairing between gRNA and target DNA, Cas9 nuclease can cleave target DNA strands along with a protospacer adjacent motif (PAM), resulting in a double-strand break (DSB) (Fig. 1). The produced DSBs can be repaired by non-homologous end joining (NHEJ) or the homology-directed repair (HDR) pathway[22,23]. NHEJ could induce indel (insertion or deletion) mutations, which usually lead to the shift of an open reading frame. By taking advantage of the error-prone NEHJ pathway, CRISPR/Cas9 system is commonly adopted for gene knockout. In contrast, HDR pathway stimulated by homologous donor templates is an accurate repair mechanism and can be used to accomplish point mutations or fragment knock-in[24] (Fig. 1). The reports on CRISPR/Cas9-mediated genome editing in plants first appeared in 2013[25,26], and artificial single gRNA (sgRNA) was used in these studies instead of the naturally occurring gRNA formed by the fusion of bacterial CRISPR RNA (crRNA) and trans-activating crRNA (tracr-RNA) (Fig. 1). Due to its simplicity and high efficiency, the engineered CRISPR/Cas9 system has since then been widely applied to genome editing in plants.
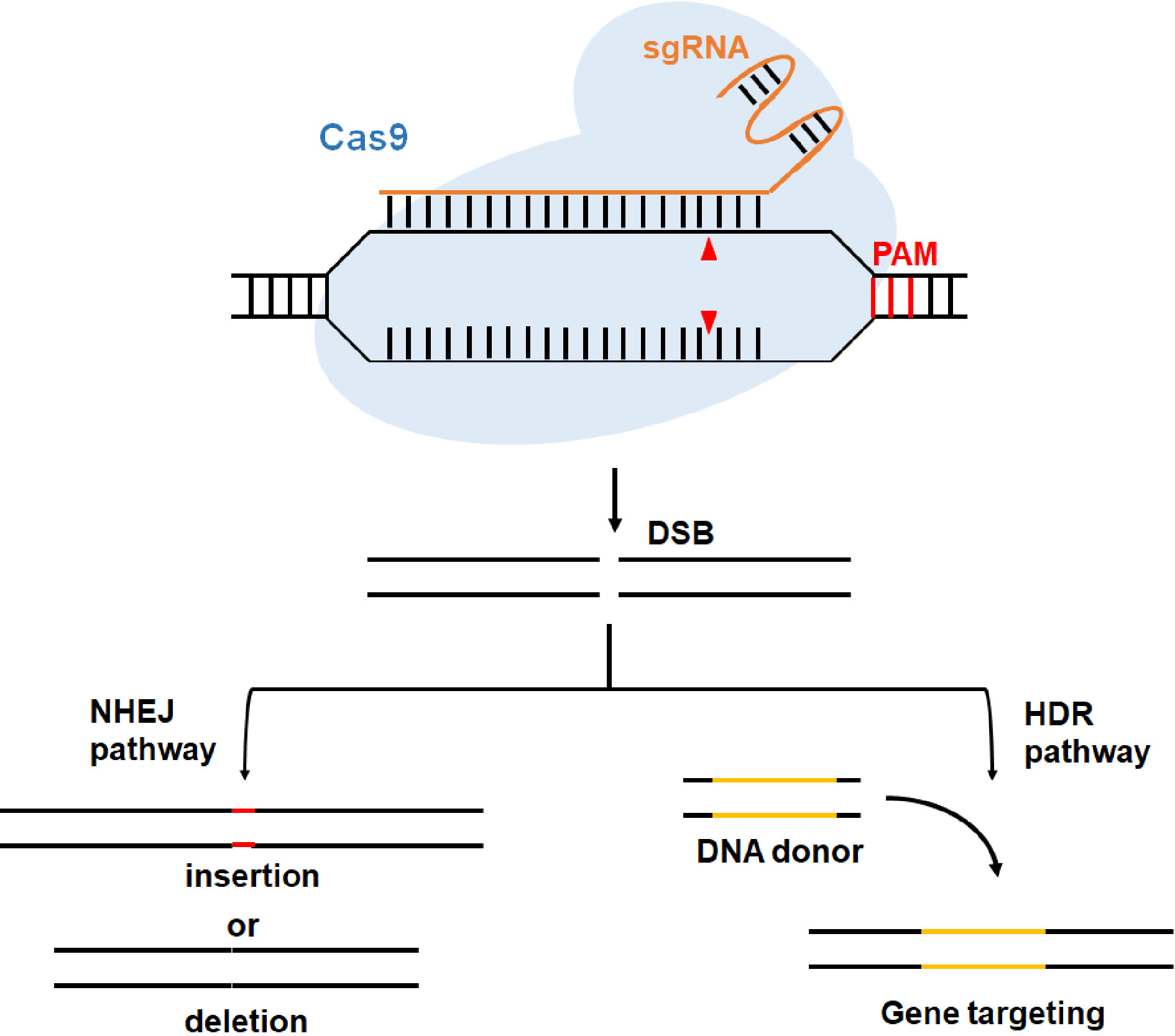
Figure 1. Schematic diagram of genome editing induced by CRISPR/Cas9 system. Cas9 protein cleaves the target sequence (the cut sites are indicated by red triangles) that is complementary with the single guide RNA (sgRNA) and produces double strand breaks (DSB). The DSB could be repaired through non-homologous end joining (NHEJ) or the homology-directed repair (HDR) pathway, resulting in indel (insertion or deletion) or gene targeting mutation.
Modifications of native Cas9 protein result in different Cas9 variants. The Cas9 nickase (Cas9n), with a mutation in RuvC or HNH domain, could cleave a single DNA strand to produce a nick, rather than DSB. The use of Cas9n combined with a pair of sgRNAs could improve the specificity of genome editing and reduce off-target effects[27,28]. The catalytically inactive Cas9, called dead Cas9 (dCas9), loses the ability of nuclease but is capable of binding to the target site[29]. The binding of the dCas9-sgRNA complex to the promoter of a gene could repress gene expression[29]. Moreover, the dCas9 can be fused with activator or repressor domains to develop CRISPR activator (CRISPRa) or CRISPR interference (CRISPRi) systems for gene activation or silencing[29,30]. In addition, the CRISPR/dCas9 system can also serve as modular platform for epigenetic modification and visualization of targeted genomic locations when fused with epigenetic factors and fluorescent protein, respectively[31,32].
A big concern for the use of the CRISPR/Cas9 system is the possible off-target effect. The best studied and most widely used Cas9 protein is isolated from Streptococcus pyogenes, and SpCas9 has been shown to generate off-target mutations during genome editing[33−35]. To minimize off-target effects, several high-fidelity Cas9 variants have been developed[36−40]. For example, the SpCas9-HF1, a quadruple substitution SpCas9 variant (N497A/R661A/Q695A/Q926A), exhibited nearly no detectable off-target effects without sacrificing the on-target activities[36]. In addition, PAM sequence is also an important factor that limits the editing scope of SpCas9. The PAMs are essential for recognition of target sites by Cas9-sgRNA complexes[8]. In fact, SpCas9 protein contains a PAM-interacting domain and recognizes canonical NGG PAMs. However, in rice, the non-canonical NAG PAMs were also reported to be efficiently recognized by SpCas9[41]. Furthermore, the modified SpCas9 variants VQR-SpCas9 (D1135V/R1335Q/T1337R) and EQR-SpCas9 (D1135E/R1335Q/T1337R), which recognize NGA and NGCG PAMs, respectively, have been used for rice genome editing[42,43]. Recently, another two newly engineered SpCas9 variants, namely SpCas9-NG and xCas9, have been applied in plants for targeted mutagenesis of the targets with NG PAMs[44−46]. The use of these SpCas9 variants greatly expands the target scope in genome editing. Alternatively, orthologous Cas9 proteins from Streptococcus canis (ScCas9) and Staphylococcus aureus (SaCas9) were also reported to cleave targets with altered PAMs in plants[47−50]. More recently, an engineered SpCas9 variant named SpRY was developed to target almost all PAMs (NRN and NYN), resulting in unconstrained genome editing[51]. The CRISPR toolbox based on the SpRY has been developed now for plant genome engineering[52], which is expected to promote plant genome editing in a PAM-less fashion. In contrast to SpCas9, Cas12a (also known as Cpf1) was reported to recognize a T-rich PAM at 5' of the target site and lead to a staggered DSB, which is distant from the PAM, with higher specificity[53,54]. More importantly, Cas12a only requires a small crRNA to cleave the target DNA, and the RNase III activity of Cas12a allows for pre-crRNA processing[53], both of which make it powerful for the editing of multiple targets simultaneously.
-
Although HDR-mediated gene targeting could accomplish precise genome modifications, the editing efficiency achieved with HDR is very low, which restricts its applications in plants[55]. The development of base editors enables more efficient and precise genome editing.
Cytosine base editor (CBE) consists of a Cas9n (D10A), a cytidine deaminase and an uracil DNA glycosylase inhibitor (UGI) (Fig. 2). Cytidine deaminase mediates deamination of cytidines (C) and produces uridines (U), which are protected by UGI from base-excision repair and finally changed into thymines (T) during DNA replication[56]. CBE was first described to generate C:G>T:A conversions in human cells[56], and following the development of human CBE, multiple CBE systems have been developed and optimized for plant base editing[57−59]. In the initial CBE (called BE3) system, rat cytidine deaminase APOBEC1 (rAPOBEC1) was used with an editing window of seven nucleotides from position 3 to 9 within the protospacer[56]. Additionally, Petromyzon marinus cytidine deaminase 1 (PmCDA1), human activation-induced cytidine deaminase (hAID) and human APOBEC3A (hAPOBEC3A) were also employed to develop CBEs for efficient base editing in plants[57,58,60]. Recently, two CBE variants, A3Bctd-VHM-BE3 and A3Bctd-KKR-BE3, were developed by using rationally designed hAPOBEC3B and showed no DNA off-target edits in rice[61].

Figure 2. Schematic overview of base editing and prime editing. In the cytosine base editor (CBE) system, a Cas9 nickase (Cas9n) is fused with a cytidine deaminase and a uracil DNA glycosylase inhibitor (UGI). The CBE could mediate C-to-T substitutions. In the adenine base editor (ABE) system, a Cas9n protein is fused with an engineered Escherichia coli adenosine deaminase (ecTadA), which catalyzes the conversion of adenine (A) to inosine (I). The I is recognized as guanine (G) by DNA polymerase during replication, resulting in A-to-G substitutions. Prime editing (PE) is accomplished by M-MLV-Cas9n-pegRNA complex, in which pegRNA functions as a guide RNA and also provides primer binding site (PBS) and reverse transcriptase (RT) template. The nicked target DNA sequence hybridizes to the PBS, priming reverse transcription of the template into the DNA sequence. The desired mutations within the pegRNA are indicated in purple. PAM, protospacer adjacent motif.
In contrast to CBE, adenine base editor (ABE), composed of a Cas9n and an adenosine deaminase effector, induces A:T>G:C conversions[62] (Fig. 2). Adenosine deaminase deaminates adenosines (A) to inosines (I), which are recognized as guanosines by DNA polymerase during DNA replication[10]. Due to the lack of natural adenosine deaminase, the Escherichia coli tRNA adenosine deaminase (ecTadA) was used for extensive directed evolution, and evolved ecTadA variants (ecTadA*) were used to develop ABEs[62]. Plant-compatible ABEs have been successfully applied in Arabidopsis, rapeseed, rice, wheat, tomato, and soybean[63−65]. Intriguingly, the promoters used for the expression of ABEs in plants were reported to have an impact on editing efficiency[63,65]. For example, the RPS5A promoter is more efficient than CaMV35S or YAO promoter in driving the expression of ABEs in Arabidopsis and rapeseed[63]. Similarly, in tomato, seven different promoters were tested with ABEs and the SlEF1α promoter outperformed the others and resulted in the highest editing efficiency[65].
CBE and ABE could be combined to create dual base editor (DuBE), in which a cytidine deaminase, an adenosine deaminase, a Cas9n (D10A) and an UGI are fused together, to conduct C:G > T:A and A:T > G:C editing simultaneously[66−68]. Plant DuBEs such as saturated targeted endogenous mutagenesis editors (STEMEs) and pDuBE1 have been developed to mediate robust dual editing[67,68], which is very useful for crop trait improvement and plant direct evolution.
Prime editing
-
Although base editors could perform precise genome modifications, only certain base substitutions can be achieved. However, many plant traits are associated with different heterogeneous mutations. To overcome the limitation problem, a 'search-and-replace' genome editing technology named prime editing was recently developed to perform multiple types of modifications as designed[69]. Prime editors (PEs) consist of a Moloney murine leukemia virus (M-MLV) reverse transcriptase (RT) and Cas9n (H840A) (Fig. 2). A prime editing guide RNA (pegRNA) that contains a RT template and primer binding site (PBS) region was used instead of sgRNA (Fig. 2), and the information carried by the RT template could be copied directly and introduced into the target site after prime editing[69]. Compared with previous base editor systems, PEs exhibited higher specificity and the editing efficiency was comparable with that obtained using base editors. As a game-changing technology, PEs have been adapted rapidly for plant genome editing[70−73]. However, low editing efficiency is the major factor that limits the applications of PEs in plants. Lin et al.[72] found that optimizing the melting temperature (Tm) of the PBS could improve prime editing efficiency. By comparing multiple pegRNAs with different PBS Tm, the researchers found that pegRNAs with the PBS Tm approaching 30 °C exhibited higher activities. Moreover, adoption of a dual-pegRNA strategy further improved prime editing efficiency[72]. A user-friendly online tool named PlantPegDesigner was developed accordingly[72], which is expected to facilitate the applications of PEs in plants.
Chromosome engineering
-
Due to the advances in CRISPR technologies, CRISPR/Cas-mediated chromosome engineering has been possible in plants. Chromosomal inversions of up to 18 kb have been accomplished using SaCas9 protein combined with an egg cell-specific promoter in Arabidopsis[74]. The method designed for generating heritable chromosomal translocations was recently established in Arabidopsis, and these CRISPR/Cas9-induced reciprocal translocations were achieved between chromosome 1 and 2, and chromosome 1 and 5, respectively[75]. Both inversions and translocations play an important role in adaptation, speciation and genome evolution[76]. Furthermore, these chromosomal rearrangements generally alter genetic linkages between plant traits. For clonally propagated crops with high genome heterozygosity, deleterious mutations linked together cannot be removed through conventional breeding. In grapevine, for instance, deleterious alleles are usually hidden at the heterozygous state in the genome[77]. Moreover, the color polymorphism of grape berry was reported to be involved in chromosomal rearrangements[78,79]. CRISPR/Cas-induced chromosome engineering provides a way to break the linkage of deleterious genes or to fix genetic linkage of target genes contributing to desirable traits.
-
Grape is an economically important fruit crop, and the CRISPR/Cas9 system has been used for precise genome editing in this species (Table 1). In recent years, several grape characteristics such as quality and disease resistance, have been improved by using CRISPR/Cas9 technology. Moreover, optimization of the currently used CRISPR/Cas9 system was also reported in grapevine. In general, embryogenic callus (or cells) induced from anthers and filaments are used for stable transformation, which is usually mediated by Agrobacterium cells that contain CRISPR/Cas9 constructs (Fig. 3; Table 1). In addition, protoplasts isolated from grape callus or leaves are also suitable for CRISPR experiments (Fig. 3; Table 1).
Table 1. Applications of the CRISPR/Cas9 system in grape.
Target
genessgRNA/Cas9 promoter Explant Delivery method Modification Target traits Editing efficiency Off-target effect Reference IdnDH AtU6/35S Embryogenic cells of 'Chardonnay' Agrobacterium-mediated transformation KO Tartaric acid synthesis 100% No Ren et al.[12] MLO-7 Not mentioned Embryogenic calli of 'Chardonnay' PEG-mediated transformation KO Powdery mildew resistance 0.1% ND Malnoy
et al.[13]VvPDS AtU6/
PcUbi4-2Embryonic calli of
'Neo Muscat'Agrobacterium-mediated transformation KO Albino phenotype 2%−86% No Nakajima et al.[81] VvPDS AtU6/2×35S Embryogenic cells of 'Chardonnay' and
'41B'Agrobacterium-mediated transformation KO Albino phenotype 22.2%−59.9% ('Chardonnay')
30.3%−86.6% ('41B')ND Ren et al.[89] VvWRKY52 AtU3, AtU6/2×35S Proembryonal masses (PEM) of 'Thompson Seedless' Agrobacterium-mediated transformation KO Botrytis cinerea resistance 31% No Wang
et al.[83]CCD8 AtU6/35S Embryogenic cells
of '41B'Agrobacterium-mediated transformation KO Strigolactones biosynthesis and shoot branching 66.7% No Ren et al.[82] VvPR4b AtU6/35S PEM of 'Thompson Seedless' Agrobacterium-mediated transformation KO Powdery mildew resistance 68.8% No Li et al.[85] VvMLO3, VvMLO4 AtU3, AtU6/2×35S PEM and somatic embryos of 'Thompson Seedless' Agrobacterium-mediated transformation KO Powdery mildew resistance 12.8%−38.5% ND Wan
et al.[84]TAS4,
MYBA7MtU6.6/
ZmUbiEmbryogenic callus
of rootstock '101-14'Agrobacterium-mediated transformation KO Anthocyanin accumulation related to Pierce disease (PD) and Grapevine Red Blotch Virus (GRBV) Not mentioned Yes Sunitha & Rock[80] PDS
TMT1,
TMT2AtU6, VvU3, VvU6/35S, VvUbi Embryogenic cells of '41B' Agrobacterium-mediated transformation KO Albino phenotype
Sugar accumulation23.5%−43.2% (PDS)
10.4%−20.9% (TMTs)ND Ren et al.[90] Abbreviations: AtU6, Arabidopsis thaliana U6 promoter; MtU6.6, Medicago truncatula U6.6 promoter; VvU6, Vitis vinifera U6 promoter; PcUbi4-2, Petroselinum crispum ubiquitin4-2 promoter; ZmUbi, Zea mays ubiquitin promoter; VvUbi, V. vinifera ubiquitin promoter; 35S, CaMV 35S promoter; KO, Knock out; ND, Not determined. 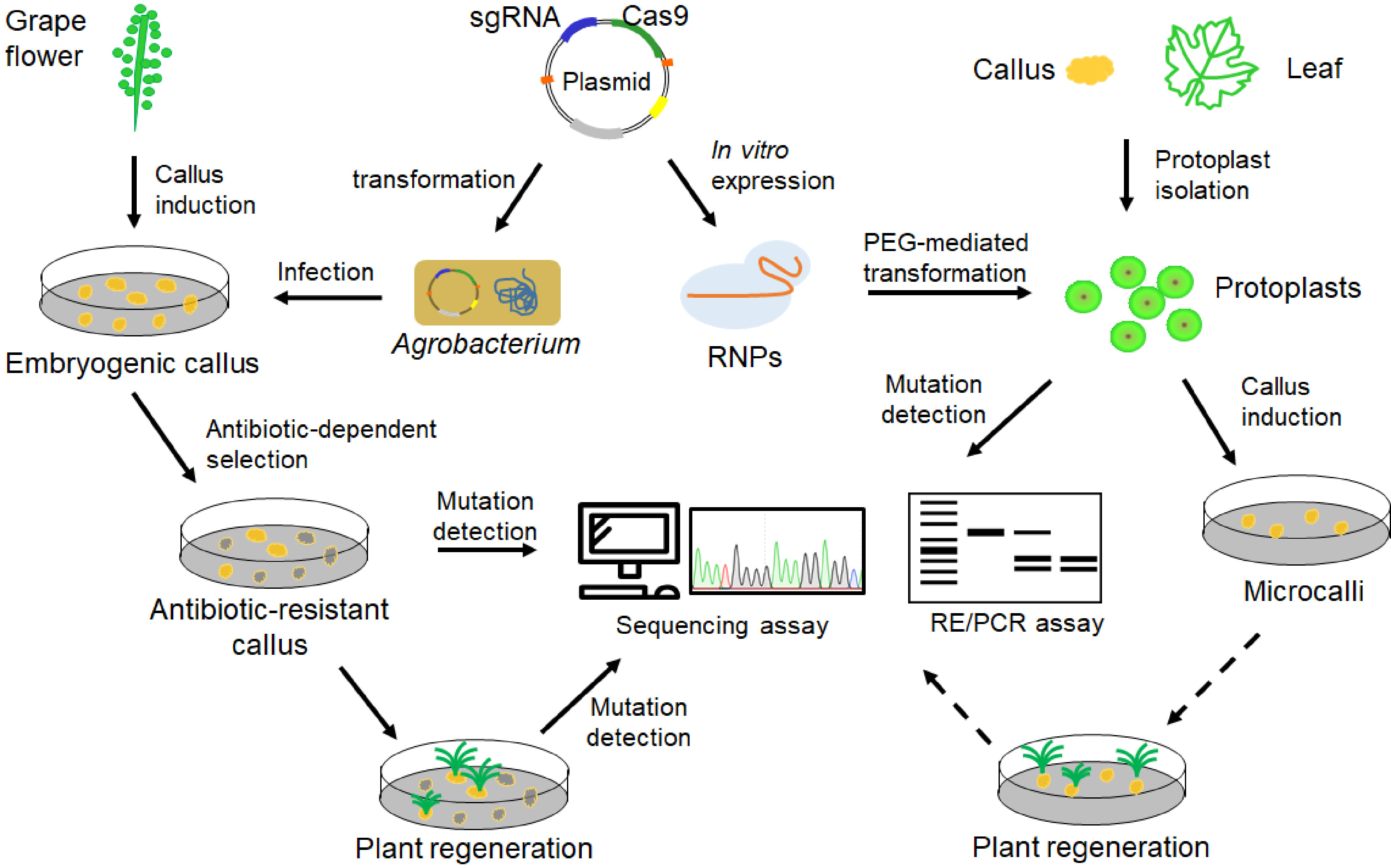
Figure 3. Pipeline of CRISPR/Cas9-mediated genome editing in grape. Both embryogenic callus and protoplasts could be used as materials for grape genome editing. CRISPR/Cas9 reagents such as plasmid constructs and CRISPR/Cas9 ribonucleoproteins (RNPs) can be delivered into the embryogenic callus and protoplasts, respectively. Edited grapevine plants could regenerate from embryogenic callus. Microcalli could be induced from grape protoplasts, but plant regeneration from the protoplasts-induced calli (indicated by broken lines) has not been achieved.
Targeted editing of genes involved in quality and growth
-
In 2016, CRISPR/Cas9-mediated genome editing was first reported in 'Chardonnay' by targeting the L-idonate dehydrogenase (IdnDH) gene, which encodes the enzyme that functions in biosynthesis of tartaric acid (TA) in grape[12]. Disruption of the IdnDH gene resulted in the decrease of TA accumulation in grape cells[12]. The trans-acting small-interfering locus4 (TAS4) and MYBA7 genes are thought to be associated with pathogen-related anthocyanin accumulation in grape, and targeted mutagenesis of the two genes were conducted using the CRISPR/Cas9 system in grapevine rootstock '101-14' to address this relationship[80]. Two TAS4b edited lines and five bi-allelic lines of MYBA7 were obtained, however no visible anthocyanin accumulated in these transgenic plants, possibly due to the functional redundancy of TAS4c and MYBA5/6 loci[80]. Furthermore, targeted knockout of V. vinifera phytoene desaturase (VvPDS) gene was achieved in table grape 'Neo Muscat', and VvPDS mutation in grapevine plants led to the development of albino leaves, with the editing efficiencies ranging from 2% to 86%[81]. Carotenoid cleavage dioxygenase 7 (CCD7) and CCD8 are two enzymes that are required for biosynthesis of strigolactones, which control axillary bud outgrowth in plants[82]. To investigate the role for CCD7 and CCD8 in the control of shoot branching in grapevine, the VvCCD7 and VvCCD8 genes were mutated in grapevine rootstock '41B' by using CRISPR/Cas9, and the obtained ccd8 mutants showed increased shoot branches[82].
Targeted editing of genes involved in disease resistance
-
Wang et al.[83] used the CRISPR/Cas9 system to edit the transcription factor gene VvWRKY52 in 'Thompson Seedless' and obtained 22 independent mutants, among which 15 plants were identified as bi-allelic mutants. Four out of the 22 edited plants were tested with enhanced resistance to Botrytis cinerea[83]. Similarly, in two recent studies, the grapevine cultivar 'Thompson Seedless' was engineered to improve its resistance to powdery mildew[84,85]. In the first study, Wan et al.[84] employed the CRISPR/Cas9 system to edit the mildew resistance Locus O3 (VvMLO3) and VvMLO4 genes, with the editing efficiency varying from 0 to 38.5%. Four VvMLO3-edited plants showed enhanced powdery mildew resistance[84]. In the other study, pathogenesis-related protein 4b (VvPR4b) gene was successfully edited, and mutation of the VvPR4b gene increased the susceptibility of grapevine mutants to pathogen Plasmopara viticola, which suggested that the VvPR4b gene may contribute to powdery mildew resistance in grapevine[85].
Although the efficacy of the CRISPR/Cas12a system in grape genome editing has not yet been demonstrated, a method known as plasmonic CRISPR Cas12a assay has been recently developed for colorimetric detection of grapevine redblotch virus infection. In this detection system, the presence of viral marker induced the degradation of the single stranded substrate (S) that serves as a linker between two DNA functionalized plasmonic gold nanoparticles, resulting in a visible blue-to-red color transition[86]. This CRISPR/Cas12a-based detection method could be used to detect virus infections in the vineyard.
Optimization of the CRISPR/Cas9 system in grapevine
-
Following the successful applications of the CRISPR/Cas9 system in grape genome editing[12,13], in 2018 the protocols for CRISPR experiments in grape based on plasmid-mediated and CRISPR/Cas9 ribonucleoproteins (RNPs)-mediated procedures were established[87]. In addition, the CRISPR database, designed for easy selection of sgRNAs for grape genome editing, is also available[88]. Successful genome editing is expected to be accomplished using these tools. The editing efficiency, however, could be further improved. To optimize editing efficiency, Ren et al.[89] surveyed the effect of three key parameters, i.e., GC content of sgRNA, expression level of Cas9, and genotypes of grape, on grape genome editing. According to the results, CRISPR/Cas9 editing efficiency increased with sgRNA GC content. In addition, the expression level of Cas9 and grape genotypes also had an influence on editing efficiency[89]. Very recently, grape U3/U6 and ubiquitin (UBQ) promoters were isolated and used to drive the expression of sgRNA and Cas9 instead of the Arabidopsis U6 (AtU6) and CaMV35 promoters, respectively, during genome editing in grape[90]. The use of grape promoters significantly improved the editing efficiency by increasing the expression of sgRNA and Cas9[90].
Multiplex genome editing
-
CRISPR/Cas9 toolkits for multiplex genome editing were first designed and applied in Arabidopsis and crops, such as rice and maize[91,92]. Two strategies are commonly used to generate multiple sgRNAs targeting different gene loci: (i) constructing different sgRNA expression cassettes into all-in-one vector[91,92], and (ii) producing multiple sgRNAs together with Cas9 from a single transcription unit (STU) by using endoribonuclease Csy4, tRNA, or ribozymes (RZs)[93,94]. In the study reported by Wang et al.[83], four sgRNAs designed for targeting the VvWRKY52 gene were assembled into the pYLCRISPR/Cas9P35S-N binary vector for genome editing. However, multiplex genome editing involving different genes was not reported until recently. Simultaneous editing of the grape tonoplastic monosaccharide transporter (TMT) gene TMT1 and TMT2 was conducted by using conventional multiple sgRNA expression cassettes (Multi-sgRNA/Cas9) and polycistronic tRNA-sgRNA cassette (PTG/Cas9), respectively, with the overall editing efficiencies being higher than 10%[90]. Simultaneous mutations of TMT1 and TMT2 resulted in low sugar accumulation in grape cells[90]. The development of Multi-sgRNA/Cas9 and PTG/Cas9 systems paves the way for multiplex genome editing involving different traits in grapevine.
Off-target effect in grape genome editing
-
In previous studies, most of the editing events in grape were specific and no off-target effect was detected[12,81−83,85]. In contrast, during the editing of TAS4 and MYBA7 in rootstock '101-14', an off-target effect was detected in TAS4b[80]. In general, the possible off-target sites are predicted using online tools, such as CRISPR-P (http://crispr.hzau.edu.cn/CRISPR2) and Plant-CRISPR (www.grapeworld.cn/pc/design.html), and then the predicted off-target sequences were amplified by PCR followed by Sanger sequencing assays[12,81−83,85]. Most recently, whole-genome sequencing (WGS) was employed to assess off-target effect in Cas9-edited grapevine plants[95]. A number of three wild-type plants and seven CRISPR/Cas9-edited plants were analyzed in greater depth by using WGS analysis, and among the 3,272 tested off-target sites, only one off-target site was identified with indel mutation[95]. These results suggest that the CRISPR/Cas9 system shows high specificity in grapevine genome editing.
Delivery of CRISPR/Cas9 components
-
CRISPR/Cas9 constructs are commonly delivered into grape callus or cells via Agrobacterium-mediated transformation[81−85]. The transformation method based on Agrobacterium is mostly efficient and low-cost, but some grapevine varieties, however, are found to be refractory to Agrobacterium infection[96]. Biolistic delivery could be used to solve the problem, given that delivery of CRISPR/Cas9 reagents could be accomplished by particle bombardment with physical force. Alternatively, polyethyleneglycol (PEG)-mediated transformation was also employed to deliver CRISPR/Cas9 RNPs into grape protoplasts[13]. Intriguingly, microcalli could be induced from edited grapevine protoplasts[87], which provides a possible way to obtain edited grapevine plants. Novel approaches, such as nanoparticle-assisted transformation for plant genetic engineering, have been recently reviewed[97]. Although nanoparticles could serve as cargo carriers for plant transformation, there are many challenges when using this technology. The biggest problem of nanoparticles-based genetic transformation is the regeneration of transgenic plants (except magnetic transfection of pollen)[97]. Whether this approach could be used for grapevine transformation needs to be further investigated in the future.
Detection of targeted mutagenesis in grape genome
-
A critical step of genome editing is the detection of targeted mutagenesis. In grape, targeted mutagenesis of genes of interest is generally detected using typical approaches. Firstly, to detect mutations in grape callus or cells after transformation, the target sequences were amplified from the grape genome and the resulting PCR products were analyzed by direct sequencing. Targeted mutations could be judged by the presence of overlapping peaks in sequencing chromatograms[82]. This method could be used to quickly confirm the existence of targeted mutagenesis. However, desired mutations might not be detected using this method in the presence of large numbers of unedited cells. Secondly, the PCR fragments could be treated with T7EI or CEL I endonuclease. The wild-type-mutated DNA hybrids could be recognized and digested by T7EI and CEL I endonucleases, resulting in the generation of two smaller DNA fragments[12,89,90]. In addition to the mutations, editing efficiencies can also be calculated according to the intensities of digested bands[89,90]. However, homologous or bi-allelic mutations could not be detected using this method. Additionally, the restriction enzyme (RE)/PCR approach is also adopted for mutagenesis detection[90]. Nevertheless, this approach requires the presence of proper endonuclease recognition sites within the target sequence. Thirdly, to explore mutation types accurately, the amplified target sequences are usually cloned into a cloning vector (like pLB vector) and then analyzed by Sanger sequencing. This detection method has been applied in both grape cells and plants[12,81−85,90]. The editing efficiencies could be obtained by calculating the mutated amplicons relative to the total analyzed[12,81]. Other approaches like high resolution melting (HRM) and high-throughput illumina sequencing are also adopted for detection of mutations in plants[98]. However, compared with the typical methods described above, these approaches are still more expensive and require specific instruments.
-
Applications of CRISPR/Cas9 rely on efficient transformation systems. The lack of efficient transformation methods is a bottleneck that restricts the application of CRISPR technologies in grapevine. The grape flowers, or rather, the anthers and filaments have been demonstrated to be suitable explants for induction of embryonic callus[81,87], which could be used for Agrobacterium-mediated transformation and plant regeneration. However, whether this method could be used in other grape varieties or species is still unknown. In addition, some cultivars are found to be recalcitrant to Agrobacterium-mediated transformation[96], hindering the delivery of CRISPR/Cas9 reagents. Furthermore, it is laborious and inefficient to discriminate transgenic or edited calli from a pool of grape materials using current screening methods, which relies on antibiotic-dependent selection, followed by PCR identification and Sanger sequencing assay[82,83,85]. Considering the low transformation rate of grape calli, more efficient screening methods need to be developed. It also usually takes a long time (> 12 months) to obtain the regenerated plants from embryonic calli[81,83,85], and as a perennial fruit crop, grapevine has a long juvenile stage. All these factors make it difficult to evaluate and optimize CRISPR/Cas systems in grapevine plants, and the effect of targeted editing of candidate genes on grape berry development or quality can be difficult to identify in a short period of time. More importantly, grapevine plants are vegetatively propagated, and it is hard to obtain transgene-free editing plants through next generation segregation.
-
As a robust and versatile technology, applications of CRISPR/Cas9 greatly facilitate the basic and applied research of grapevine. However, most of the up-to-date CRISPR technologies have not yet been applied in grapevine. The current genome editing in grapevine is mainly focused on gene knockout, and it remains a long and arduous task to enrich the CRISPR toolbox for grape genome editing. Precise genome editing technologies, i.e., base editing and prime editing, will facilitate the improvement of desirable traits. Due to the difficulties in grape transformation, alternative transformation systems based on protoplasts and hairy roots could be used to evaluate efficacy and efficiency of new CRISPR platforms in grape. A recent study reported a method of de novo meristem induction to generate transgenic plants, which promises to overcome the bottleneck of plant tissue culture[99]. Although this strategy has been proven to be effective in grapevine[99], the generation of chimeric plants is an unavoidable problem when using this method. In addition, rapid and robust detection methods are also needed to screen edited grape calli or plants more efficiently. In a previous study, recovery of non-functional-enhanced green fluorescence protein (nEGFP) was used as an indicator of successful editing in grape cells[100]. Visible markers that could be detected by the naked eye are preferred and can be used without the need for specific instruments. Considering the public concerns on genetically modified organisms, in the future, genome editing methods should be developed to produce transgene-free edited grapevine plants. Site-specific recombinases and newly developed PEs promise to remove T-DNA sequences from the plant genome. In conclusion, the use of CRISPR/Cas technologies has already revolutionized, and will continue to revolutionize, basic and applied research of grapevine.
- This work is funded by grants from the National Natural Science Foundation of China (32001994) and Agricultural Breeding Project of Ningxia Hui Autonomous Region (NXNYYZ20210104).
- The authors declare that they have no conflict of interest.
- Copyright: © 2022 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Ren C, Lin Y, Liang Z. 2022. CRISPR/Cas genome editing in grapevine: recent advances, challenges and future prospects. Fruit Research 2: 7 doi: 10.48130/FruRes-2022-0007

CRISPR/Cas genome editing in grapevine: recent advances, challenges and future prospects
- Received Date: 24 November 2021
- Accepted Date: 12 April 2022
- Published Online: 16 May 2022
Abstract: The development of clustered regularly interspaced short palindromic repeats/CRISPR associated protein 9 (CRISPR/Cas9) system has revolutionized genome editing and plant breeding. Applications of CRISPR/Cas9 technology in fruit crops, including grapevine, enable precise improvement of agronomically important traits. In this review, we first describe genome editing based on the most widely used CRISPR/Cas9 system and recently developed CRISPR technologies. We then focus on applications of CRISPR/Cas9 in improvement of disease resistance, optimization of CRISPR/Cas9 systems, multiplex genome editing and off-target effect analysis in grapevine. We also discuss the challenges facing genome editing that should be overcome to realize the potential of CRISPR technology in grapevine. Finally, we highlight possible future experimental considerations for more precise and efficient genome editing in grapevine.
-
Key words:
- CRISPR/Cas9 /
- Genome editing /
- Grapevine /
- Advance