








-
Fruit is an organ formed by the pistil of angiosperms through pollination and fertilization, and the development of the ovary or other parts of the flower such as the receptacle and sepals[1]. After fertilization, the ovary is metabolically active and grows rapidly. The ovule develops into the seed, and the wall of the ovary develops into the pericarp that wraps around the seed to form the fruit. From a botanical point of view, the edible parts of some vegetables, such as cucurbitaceous plants, are also fruits. Fruits are closely related to human life. They are rich in glucose, fructose and sucrose, as well as a variety of inorganic salts, vitamins and other nutrients[2]. Fruits are not only good for eating fresh, but can also be processed into dried fruit, jam, preserves, wine, juice, fruit vinegar and other foods. In addition, dates, citrus, hawthorn, apricot, longan and other fruits are also commonly used in traditional Chinese medicine. The versatility and large-scale production of fruits require in-depth research on their growth and development, quality formation and maintenance. At present, fruit research focuses more on cultivation improvement and post-harvest techniques, while the basic research on biochemistry, genetics and molecular biology of most fruits still lags behind that of field crops. Fruits are unique experimental materials for scientific research since many phenomena are not found in field crops. Deeper basic research on fruits will improve our understanding of these diverse and unique phenomena and accelerate technological innovation to better utilize the fruits.
Among the many regulatory factors in the process of fruit growth, development, ripening and stress response, miRNAs, as a major and important class of small RNAs, have attracted increasing attention[3−6]. In the past two decades, numerous miRNAs in the main edible parts of diverse fruits have been identified through computational analysis, high-throughput sequencing and experimental validation[7−11]. In addition, studies on miRNA function in fruits have been conducted, revealing mechanistic complexity and unique regulatory features compared to those studied in model plants. However, current functional characterization of miRNAs has been limited to a few fruit species, including tomato, grape, strawberry, pear, apple and kiwifruit[12−17]. So far, the miRNA functions associated with unique traits in many characteristic fruits remain largely unknown, mainly due to many fruit-bearing species exhibiting genetic complexity and long juvenility. In this review, we summarize miRNA research progress on the identification and functional characterization of miRNAs in fruit development, quality formation, and stress response, and aim to provide comprehensive information on miRNAs for future fruit research and breeding.
-
While most miRNA research focuses on model plants, small RNA sequencing has been applied to fruit-bearing horticultural crops and diverse fruits. Of the plant species registered in the miRBase database that collects miRNA information on all organisms, 13 fruit species are included (http://mirbase.org, release 22.1). A more comprehensive sRNA annotation of 143 plants, including 40 fruit species, has been recently reported (http://www.plantsrnas.org)[18]. Here, we summarize fruit miRNA information based on this public database and we include six more fruit species, including blueberry[19], litchi[20], sponge gourd[21], persimmon[22], pitaya[23] and passion fruit[24], according to the newly available data. In total, 41 MIRNA families, including both conserved and non-conserved ones, are presented to illustrate the miRNA conservation in the 46 fruit species (Fig. 1). Most dicotyledonous fruit species belong to several big orders of angiosperms, including Rosales (apple, peach, pear, strawberry, cherry and almond), Cucurbitales (cucumber, pumpkin, zucchini, squash, gourd and melon), Solanales (tomato, pepper and eggplant) and Rutales (pomelo and orange), with other species distributed in the orders of Sapindales (litchi, longan and mango), Theales (kiwifruit, papaya and passion fruit), and Rhamnales (grape and date). In contrast, only a few fruits are monocotyledonous mainly in the order of Principes (coconut, oil palm, and phoenix date), while banana and pineapple belong to Zingiberales and Bromeliales, respectively (Fig. 1).
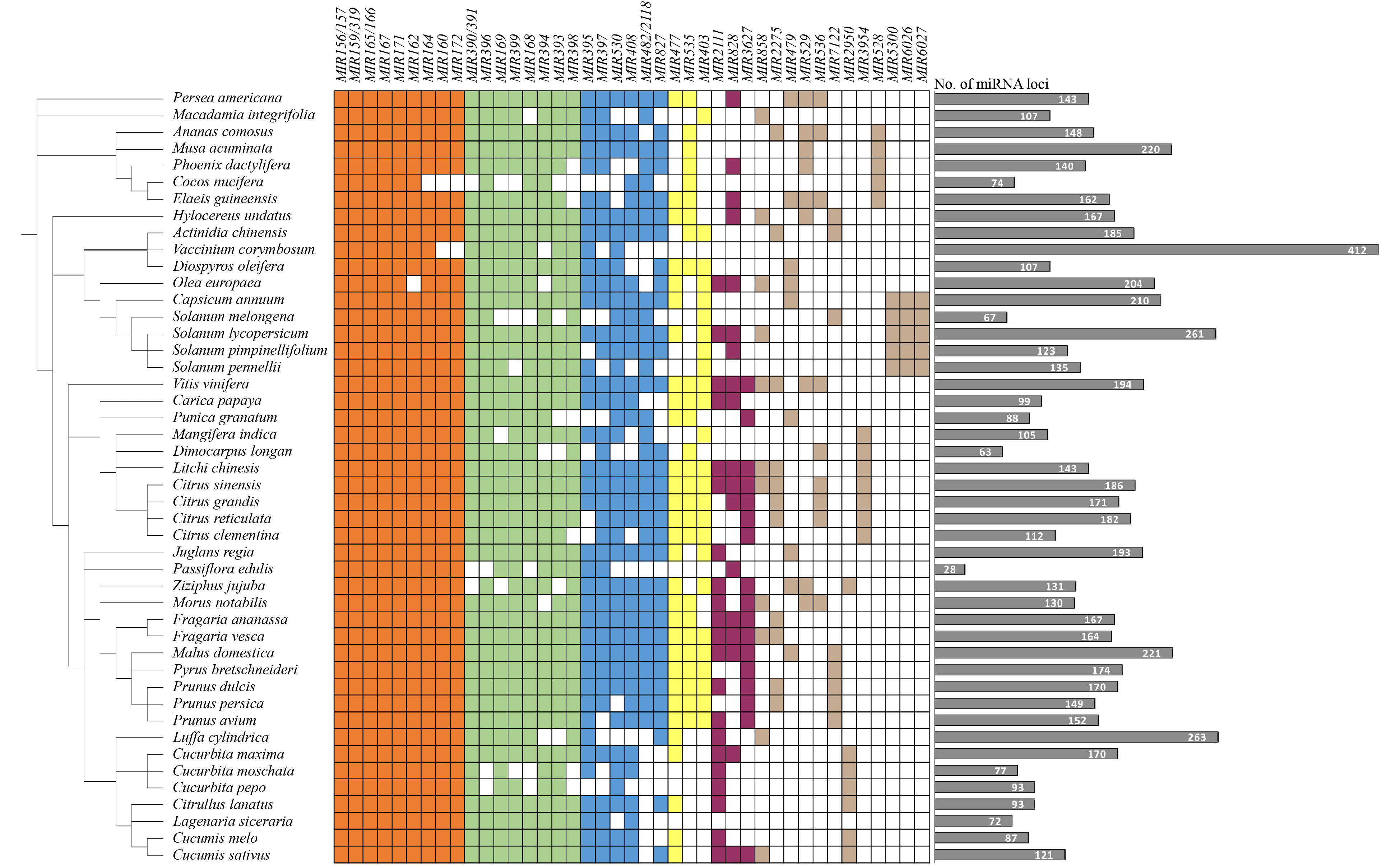
Figure 1. Conservation of miRNAs in fruits. MIRNA families among 46 fruit species, most of which are included in the database repository-sRNAanno[18] with six fruit species appended based on the newly released data[19−24]. The tree diagram on the left shows the general evolutionary relationship of the fruit species. Boxes are highlighted if a MIRNA family has been identified in the fruit species via sequencing or annotated based on its genome sequence. Groups of MIRNA families are highlighted in different colors according to the degree of conservation. Fruit species lacking a MIRNA family or with uncertain identification are left blank. The bar graph on the right shows the number of miRNA loci in each fruit species.
Not quite as observed in the whole plant kingdom, more than half of the 41 MIRNA families in fruits are highly conserved, while a small number of MIRNA families are lineage- or species-specific, suggesting that the evolutionary rate of fruit MIRNA genes is relatively slow. Specifically, 23 MIRNA families are present in over 70% of the fruit species, including MIR156/157, MIR159/319, MIR160, MIR162, MIR164, MIR165/166, MIR167, MIR171 and MIR172 (> 95%); MIR168, MIR169, MIR390/391, MIR393, MIR394, MIR396, MIR398 and MIR399 (85%−95%); MIR395, MIR397, MIR408, MIR482/2118, MIR530 and MIR827 (70%−85%). It is worth noting that the MIR482/2118 family is missing in all cucurbitaceous fruits. In other cases where highly conserved MIRNA families are absent in individual species, it may partly be attributed to the insufficient quality of the genome annotation. Some of the 18 non-conserved MIRNA families appear to be evolved as lineage-specific. For example, the MIR3627 family is restricted in the rosaceous and rutaceous fruits, while the MIR535 family seems to be missing in solanaceous and cucurbitaceous fruits. Three families, i.e. MIR5300, MIR6026 and MIR6027, are specific to solanaceous fruits, and the families of MIR7122, MIR2950 and MIR3954 appear to be mainly restricted in rosaceous, cucurbitaceous and sapindaceous/rutaceous fruits. In the fruits of a few monocotyledonous species, MIR403 and MIR2111 families are lost, while the MIR528 family has been evolved as monocot-specific (Fig. 1).
In general, the genes targeted by conserved miRNAs are also conserved, that is, the target genes of a conserved MIRNA family belong to the same family in different plant species. However, exceptions appear where it has been recently reported that the target genes of miR528 in different monocots may have a large scale of function differentiation[25]. In fruits, highly conserved miRNAs tend to regulate fruit morphogenesis, such as size and shape[16,26,27]. In contrast, pathways regulated by species-specific miRNAs often reflect the quality characteristics of the species, such as the formation of unique color, aroma, taste and resistance of fruits[13−15,28].
-
Fruit is a highly specialized organ useful in understanding the evolution of angiosperm reproductive innovation. In general, fruit development is a unique coordination between embryonic development and seed formation. Following fertilization, fruits typically undergo a dramatic enlargement, which is accompanied by cell division and expansion. The primary fruit provides protection and nourishment for the growing seeds, and also helps seed dispersal at maturity[1]. Genetic studies on the Arabidopsis fruit model have identified several regulatory genes related to fruit differentiation and also outlined the genetic interaction framework for the fruit patterning model[29]. Fruit development is mainly regulated by FRUITFULL (FUL) proteins, which contain conserved MADS-box domain required for normal growth and differentiation of the cells. In addition, plant hormones play an essential role during fruit growth, especially auxin with the AUXIN RESPONSE FACTORs (ARFs), key transcription factors involved in the regulation of auxin signaling. Recently, a study clearly shows that miR172 is critical for proper fruit growth after fertilization, by targeting the transcriptional repressor APETALA2 (AP2). Furthermore, the MADS-domain protein interacts with ARF proteins to directly activate the expression of the MIR172C gene, which delineates the upstream transcriptional architecture regulating miR172 during fruit growth[30]. It has also been proposed that such MADS-ARF associations may occur in diverse fruits, highlighting a miRNA-centered regulatory pathway for fruit development. An early study of sRNA profiling in tomato fruit, covering the period from closed flower bud to ripened fruit at ten time points, reveals differential miRNAs during fruit development. For example, miR172 and miR390 are mainly expressed at the flowering stage but gradually decreased upon fruit set and early development. In contrast, miR159 is most abundant in early fruit development, and miR156 and miR396 are both significantly increased after fruit set and during early fruit development[31]. Later reports on tomato miRNAs further unveil their conserved and novel targets, among which SQUAMOSA PROMOTER-BINDING PROTEIN-LIKE–COLORLESS NON-RIPENING (SPL-CNR) and AP2a are targeted by miR156 and miR172 respectively, and both pathways have been characterized to play key roles in fruit development[32,33]. In addition, MIR159-overexpressing tomato plants exhibit precocious fruit set and parthenocarpy without modifying fruit shape, through the miR159-GIBBERELLIN-SPECIFIC MYB1/2 (GAMYB1/2) module[34].
Parthenocarpy is a unique phenomenon in which the ovary forms the fruit without fertilization, so contains no seeds or abortive seeds. Seedless or soft-seeded fruits are favored by consumers because of their high quality, good taste and convenience to eat. It has been speculated that differentially expressed miR164e and miR172b targeting NAM/ATAF/CUC (NAC), WRKY and MYELOCYTOMATOSIS (MYC) genes may be related to seed hardness in pomegranate[35]. Grapevine miR058 targets the polyphenol oxidase (PPO) gene regulating the synthesis of lignin, whose accumulation can promote berry stone development in grape fruit[36]. A recent study reveals that miR159 is involved in seedless berry development, via regulating its target GAMYB in response to GA treatment[37]. Moreover, miR160a/b/c are also associated with GA-induced grape parthenocarpy through targeting auxin response factors ARF10/16/17. These findings are of great significance for the molecular breeding of seedless grapes[38]. In addition, tomato can develop seedless fruit when miR172 is overexpressed[39]. However, overexpression of a miR160-insensitive version of ARF10 in tomato can also lead to seedless fruit or fruit with abnormal seeds that fail to germinate[26]. Similarly, transgenic tomato plants overexpressing two cucumber TRANSPORT INHIBITOR RESPONSE1/AUXIN SIGNALING F-BOX (TIR1/AFB) genes (CsTIR1 and CsAFB2) also display a parthenocarpic fruit phenotype[40].
-
Ripening is essential for quality acquisition and also signals the beginning of senescence, both of which are integral parts in the life cycle of fruits[41]. Many studies have demonstrated that miRNAs serve as a bridge between transcriptional regulation and hormone response of fruit ripening. In general, miRNAs can be divided into two groups, which play positive and negative regulation on fruit ripening and senescence, respectively. In the group with positive effects, overexpression of miR172 accelerates fruit ripening with enhanced ethylene biosynthesis, by specifically repressing SlAP2a but no other ripening-related genes such as RIPENING INHIBITOR (RIN), NEVER RIPE (NR), TOMATO AGAMOUS-LIKE1 (TAGL1) and HD-ZIP HOMEOBOX PROTEIN-1 (HB-1)[42]. The tomato rin mutant displays no climacteric rise of ethylene and impaired ripening characteristics, and RIN can directly bind to the MIR172a promoter to affect miR172 accumulation[43]. Similarly, miR1917 targeting CONSTITUTIVE TRIPLE RESPONSE4 (CTR4) splice variants mediates ripening and overexpression of miR1917 enhances ethylene response and accelerates fruit ripening in tomato. However, repressing ETHYLENE INSENSITIVE2 (EIN2), the essential positive regulator of ethylene signaling, can reverse this phenotype[44]. More miRNAs fall into the group with negative effects. For example, overexpressing miR157 delays tomato fruit ripening by targeting CNR, a key SPL gene for fruit ripening[45]. A novel strawberry miRNA (fan-miR73) has been reported to prevent fruit ripening by negatively regulating its target ABSCISIC ACID INSENSITIVE5 (ABI5) gene, which is critical for ABA signaling[46]. In kiwifruit, miR164 is involved in fruit ripening by negatively regulating its target NAC6/7, and the miR164-NAC module also exists in various fruit species like citrus, grape and rosaceous fruits[17]. In strawberry, miR164 negatively regulates NAC to play an important role in delaying fruit senescence during low temperature storage[47]. Recently, it has also been demonstrated that pre-harvest UVC-delayed senescence of strawberry fruit may contribute to the enhancement of the antioxidant system mediated by miR159 and miR398, which target lipoxygenase (LOX) and Cu/Zn superoxide dismutases (CSD), respectively[48]. A greater number of miRNAs have been identified to be involved in regulating fruit ripening and senescence. For example, comparative analyses of miRNAs and their targets in banana treated with exogenous ethylene or 1-MCP have revealed several differential miRNA-target pairs, including miR156a-SPL7/9, miR395d-EIN3, miR166e-endo-1,3-β-glucosidase14 (EG14), miR172a-HOMEOBOX-LEUCINE ZIPPER PROTEIN9 (HOX9), miR535a-SODIUM/HYDROGEN EXCHANGER6 (NHE6), all of which may be related to banana fruit ripening[49,50]. In grape, a mutant that matures one month earlier than the wild type displays miR399 downregulation of miR477b upregulation[51]. Furthermore, genes associated with fruit softening and membrane lipid peroxidation, including LOX, β-galactosidase (BGA) and pectinesterase (PE), predicted as targets of miR479, miR397a, miR3627 and novel_miR22, are believed to be involved in fruit senescence[52]. Another study shows that 1 kHz sound waves can delay tomato fruit ripening, partly through differentially expressed miR6022 and miR6024[53]. In addition, miR160, miR164, miR167, miR171, miR172, miR858 are found to be associated with fruit ripening and senescence in high CO2-treated strawberry and sweet cherry[9,54,55]. In litchi, several miRNA-target pathways are actively involved in senescence-induced pericarp browning, including miR2118-PHOSPHOGLYCERALDEHYDE KINASE (PGK)/Ca2+-ATPase, miR397-LACCASE (LAC), miR396-ETHYLENE RESPONSE FACTOR (ERF)/CALLOSE SYNTHASE and miR858-MYB[56,57]. By comparing sRNAome between two varieties of sponge gourd with resistant and sensitive browning features, a group of differentially expressed miRNAs, in particular miR172a and miRn60, have been identified and a miRNA-mediated regulatory model has been proposed for the pulp browning[21]. Surface browning of fresh-cut apples reflects the degree of senescence, which seriously affects the acceptance of consumers. Through miRNA and degradome sequencing, several miRNA-target pathways, such as miR156-SPL, miR164-NAC, miR6478-patatin-like protein 2 (PLP2) and miR319-TB/CYC/PCF4 (TCP4)/GAMYB/acyl-CoA-binding protein 4 (ACBP4) regulating ROS, phenylpropanoid, and lipid metabolism have been found to participate in the browning inhibition of fresh-cut apples by hydrogen sulfide (H2S) treatment[58]. However, it should be noted that most of the above reports, except for tomato, still focus on the identification and expression profiling of miRNAs and their target genes, while reports on their biological functions related to fruit ripening and senescence remain limited.
-
Fruit size has a big impact on consumer acceptance, with large fruits usually more expensive than smaller ones. miR172 makes a significant contribution to fruit growth via its target gene AP2, and recent studies reveal that this and miR172-AP2 module differentially affects fruit size, depending on the tissue type from which the fruit derive[39]. In apple, the fruit derive mainly from sepal tissue that is positively regulated by AP2, and overexpression of miR172 causes the downregulation of AP2 and then a dramatic reduction in fruit size. Moreover, larger fruit displaying a higher AP2 expression coincide with a lower miR172 level under a transposon insertional allele of miRNA172[16]. However in Arabidopsis, the fruit (silique) derive from carpel tissue, which is negatively regulated by AP2, so miR172 overexpression leads to bigger siliques[30]. Interestingly, tomato fruit are similar to Arabidopsis in ovary origin, but also share similarity with apple fruit in pulp development. This may partially explain the seedless and smaller fruit phenotype resulting from the overexpression of miR172 in tomato fruit since hormones released from seeds are thought to promote fruit enlargement after fertilization[39]. It has been also reported that through the short tandem target mimic (STTM) approach, the down-regulation of miR396a/b and up-regulation of their target GROWTH-REGULATING FACTORs (GRFs) in tomato lead to bigger fruit than the wild type[59]. Similarly, miR1917 knockdown mutant displays increased fruit size, with elevated expression of its target gene CTR4 and altered ethylene response in tomato[60]. Based on these findings, it is speculated that miR396 and miR1917 might have complementary functions in regulating fruit size. On the contrary, miR171 may regulate fruit size through its target GAI/RGA/SCR24 (GRAS24), with the evidence that overexpression of GRAS24 causes smaller fruit via regulating gibberellin and auxin homeostasis to inhibit cell division and expansion in tomato[27]. Smaller fruit are also observed in the miR168-resistant ARGONAUTE1s (AGO1s) transgenic tomato plants, indicating the interaction between miR168 and AGO1s in determining fruit expansion[61]. In cucumber, the expression level of miR156a decreases in ovaries during the expanding period compared with that in nonexpanding ovaries, implicating the involvement of miR156a in fruit size[62]. Recently, a novel strawberry miRNA, Fa_novel6, has been reported to target HERCULES1 (HERK1) encoding a receptor-like kinase, which likely contributes to fruit size determination in fruit ripening[6].
Role of miRNAs in fruit shape
-
In addition to regulating fruit size, miRNAs have also been reported to affect fruit shape. For example, overexpression of miR172 in tomato develops into parthenocarpic fruit with ectopic ovaries inside the fruit[39]. Similarly, a fruit-out-fruit phenotype is displayed in the transgenic tomato overexpressing AtMIR156b, probably due to the presence of meristem-like structures inside the ovaries as shown by scanning electron microscopy and histological analyses[12]. Overexpressing a miR166-resistant version of REVOLUTA (REV) also results in ectopic fruit formation on receptacles, and most of the secondary fruits are irregularly spaced without placenta and ovules[63]. Auxin signaling is largely mediated by miR160-targeted ARFs. It has been reported that ectopic expression of a miR160-insensitive ARF10 in tomato gives rise to cone-shaped fruit, accompanied by a sharper angle at the distal fruit edge and a higher ratio of proximal/distal diameters in the transgenic fruit[26]. Further, downregulation of miR160 causes a significantly increased expression of its target ARF10A/10B/17 and leads to elongated, pear-shaped fruit. In fact, the ovaries of the transgenic flowers are narrower and longer, markedly different from those of the wild type, and these phenotypes become evident in early bud development and are maintained until flowering[64]. A recent study shows that intra-species grafting can induce alterations in the fruit shape of Cucurbita pepo cultivars, with altered expression patterns of scion miRNAs. For example, miR159, miR164 and miR171 all show significant downregulation in the hetero-graft that leads to the smaller fruit[65]. These miRNAs have been reported to be involved in plant meristem maintenance and organ boundary by targeting SPL/GAMYB, NAC and GRAS transcription factor genes[3]. Hence, it is likely that changes in the expression of these miRNAs may remodel key genes during fruit growth, resulting in phenotypic changes in fruit size.
Role of miRNAs in fruit color
-
For fruit, the attractive color is often a decisive external quality that promotes consumer purchase. The fruit color depends on the type and level of diverse pigments, including chlorophyll, carotenoids, anthocyanins and flavonoids. Previous studies on miRNA regulation of fruit color have been focused on anthocyanins that are synthesized via the phenylpropanoid metabolic pathway. The composition and amount of anthocyanins are determined by multiple structural and regulatory genes. The regulatory genes include the transcription factors MYB, basic helix-loop-helix (bHLH), and WD40 proteins, which synergistically regulate the anthocyanin biosynthesis and are highly conserved among different fruit species[66]. Also highly conserved is the miRNA regulation of R2R3-type MYBs in the MYB–bHLH–WD40 complex. Based on high-throughput sequencing and functional analyses, the regulation of most R2R3-type MYBs by miR828 and/or miR858 to influence anthocyanin biosynthesis has been unveiled in diverse fruits[19,67−70], suggesting the conservation of miR828/858-MYB pathway in fruit coloration. In tomato, silencing miR858 causes significantly increased expression levels of its target MYB7-like gene, leading to increased anthocyanin accumulation[71]. In grape, miR828 and miR858 jointly target MYB114 that controls anthocyanin accumulation[13], and heterogeneous expression of vvi-miR828 in Arabidopsis shows a phenotype of lighter colored leaves, which is consistent with the phenotype of grape berries in the control and indicates that miR828 acts as a repressor for anthocyanin biosynthesis[72]. It is also reported that in red-colored kiwifruit, miR858 negatively regulates anthocyanin biosynthesis by targeting a MYB family member (named MYBC1) via transient co-transformation, and overexpressing miR858 or silencing MYBC1, both could significantly inhibit fruit coloration[73].
Besides this conserved miR828/858-MYB pathway, miR156-targeted SPLs have been recently characterized to interfere with the interaction between MYB and bHLH in the MYB–bHLH–WD40 complex, thus negatively regulating anthocyanin biosynthesis[74]. Such a regulatory mechanism has also been reported in fruits. For example, a recent study in blueberry reveals that the miR156a-SPL12 module coordinates the accumulation of chlorophylls and anthocyanins during fruit ripening[75]. Similarly in pear, the abundance of miR156 variants increases during anthocyanin accumulation, with multiple target SPLs downregulated. Yeast two-hybrid assays further show that SPL10/13 can interact with MYB10, suggesting the involvement of a miR156-SPL module in anthocyanin biosynthesis by affecting the MYB-bHLH-WD40 complex[76]. In litchi, miR156a regulates anthocyanin in a similar way via targeting SPL1 that interacts with MYB1[70]. However, another study on the role of long noncoding RNAs in apple fruit reveals an opposite function of miR156-targeted SPLs in the light-induced rapid anthocyanin accumulation compared with Arabidopsis[77]. Additionally, miR5072 in apple and a novel miRNA (NEW41) in litchi have been recently identified to target anthocyanidin reductase (ANR) and chalcone isomerase (CHI), respectively, both of which are associated with anthocyanin metabolism[70,78].
In addition to anthocyanins, lycopene, carotenoids and betalain, as major plant pigments in fruits, have also been implicated to be regulated by specific miRNAs. Of the differentially expressed miRNAs identified from wild-type sweet orange and its spontaneous red-flesh mutant, miR1857 can target lycopene β-cyclase (LYCb) gene, which encodes a rate-limiting enzyme in the conversion of lycopene to downstream cyclic carotenes. Therefore, it has been proposed that the differential expression of miR1857 may result in abnormal lycopene accumulation in the mutant[79]. By in silico analysis of the relationship between carotenoid content and miRNA expression in different tissues of tomato, seven miRNAs have been identified as related to the carotenoid biosynthetic pathway[80]. In pitaya, whose color is mainly attributed to betalain accumulation, miR157, miR160, miR6020, miR828 and miR858 are found to be likely involved in betalain biosynthesis[23]. By comparing miRNAs in a russet skin mutant pear with its wild type, the differentially expressed miR396, miR408 and novel miR102, miR274, miR42, and miR442 are speculated to play potential roles in suberin biosynthesis[81].
-
As one of the important edible qualities, most fruits have a characteristic aroma, which directly affects consumer preference. Fruit aroma is determined by different aromatic substances and those are mixtures of various volatiles, including alcohols, aldehydes, ketones, esters, terpenes and sulfur-containing compounds. The main aroma substances of fruits come from fatty acids, amino acids and secondary metabolism. At present, there are few studies on the miRNA regulation of aroma anabolism in fruits. In 'Nanguo' pear during cold storage, differentially expressed miR172, miR159, miR160, miR395, miR399, miR535 and miR7120 negatively regulate their target genes encoding LOXs, hydroperoxide lyase (HPL), and alcohol dehydrogenase (ADH), all of which are key enzymes in the oxidation process of fatty acids[82]. In addition, it has been speculated that both miR156 and miR477 may contribute to fatty acid accumulation during the development of Camellia oleifera fruit, via targeting genes that encode a butanol dehydrogenase and an acetyl-CoA carboxylase carboxyl transferase subunit, respectively[83]. In tomato, a newly identified miRNA, miR1534, is predicted to be involved in the targeting of a terpene synthase[84]. Moreover, a comprehensive study in Arabidopsis and fragrant herb Pogostemon cablin reveals that the miR156-targeted SPL can activate the expression of sesquiterpene synthase (TPS) and patchoulol synthase (PTS) genes, respectively, to moderate the biosynthesis of sesquiterpene[85]. Considering that terpenes are the main characteristic aroma components in fruits such as grape and litchi, it is reasonable to speculate that the miR156-SPL-TPS pathway may also play a role in the synthesis of terpene aroma in fruits.
Besides the characteristic aroma of fruit, its unique flavor is also a key edible quality. Fruit flavor mainly depends on the composition and level of sugars, organic acids and specific flavor substances. For example, it is found that soluble solid content is positively correlated with phosphorus level in ripening strawberry fruit[86]. Also, phosphorous metabolism is partially under miRNA regulation in that miR399 guides the cleavage of the PHOSPHATE OVERACCUMULATOR2 (PHO2) gene, which negatively affects Pi remobilization. A study provides further evidence that overexpression of miR399a in strawberry can significantly enhance the fruit flavor through increased Pi level and higher sugar content[14]. Similarly, a comparative analysis of miRNAome in pear has identified nine miRNAs that putatively participate in sugar and acid metabolism during the formation of fruit flavor[87]. In addition, miR319 has been reported to negatively regulate tomato fruit flavor via targeting LANCEOLATE (LA), which encodes a transcription factor from the TCP family. Overexpression of miR319 in tomato results in a reduction of titratable acid and soluble sugar content, while overexpressing its target LA can obtain opposite results, thus improving fruit flavor[88]. Additional miRNAs have been identified that are involved with sugar and acid metabolism in fruits, including grape, wolfberry and peach[89−91]. It is well established that glutamate is accumulated in tomato fruit during ripening, which contributes to the 'umami' flavor, and a novel tomato miRNA, miR-W*, is validated to be able to target two different genes encoding membrane-bound ATPase and glutamate permease, both of which are associated with glutamate accumulation. Furthermore, the expression of MIR-W gene decreases at the breaker stage of the fruit, suggesting that this novel miRNA is likely to make a contribution to the fresh flavor in tomato[31].
Role of miRNAs in fruit taste
-
miRNAs have been recently reported to regulate fruit taste, a factor greatly affecting the edible quality, which is largely determined by textural characteristics. In climacteric fruits, softening usually marks the beginning of fruit ripening, indicating that the fruit is ready to eat. In tomato, miR396, miR482 and a novel miRZ7 have been revealed to regulate fruit softening by targeting genes that encode endo-1, 4-β-glucanase, pectate lyase, and β-galactosidase, respectively[92]. Furthermore, both miR156 and miR157 have been demonstrated to sequentially regulate the softening process of tomato fruit at different ripening stages[45]. Granulation is one of the key problems affecting the taste of citrus fruit during storage. It has been found that the accumulation of two miRNA families (miR397 and miR828) negatively correlates with juice sac granulation, probably by regulating lignin biosynthesis in citrus[93]. Similarly, an increase in lignified stone cells tends to reduce the taste of pear fruit. Transient overexpression of miR397a in pear fruit can reduce lignin content and stone cell number via silencing of the LAC gene. Furthermore, stable overexpression of miR397a in tobacco plants displays a similar phenotype but does not change the ratio of syringyl- and guaiacyl-lignin monomers[15]. In peach, melting and stony hard flesh present very different tastes. It is found that the expression of MIR171 is induced in melting type flesh but remains at a low level in stony hard type flesh in peach, suggesting the involvement of miR171 in the texture of peach[94]. An unacceptably astringent taste of fruit is usually caused by soluble tannins. In persimmon, miR156j, miR396g, miR395p, miR858b and miR2911a may play important roles in improving the taste of fruit, through targeting tannin biosynthesis pathway genes[95].
Role of miRNAs in fruit nutrition
-
Fruits are rich in phytochemicals, especially antioxidants such as vitamins and flavonoids, many of which are beneficial to human health[96]. Fruits also contain a variety of micronutrients, including Cu, Zn, Fe, Mn, P and S, which are assimilated during growth and development. Moreover, dietary fibers in some fruits have been recognized to be beneficial for preventing chronic diseases. It is well known that miRNAs are vital for maintaining nutrient homeostasis in plants, and many conserved miRNA pathways suggest the involvement of miRNAs in the balance of nutrient levels, by regulating the expression of genes that are involved in the synthesis, transport, conversion and metabolism of various nutrients. So far, miRNAs have been reported to be widely involved in the biosynthesis of plant secondary metabolites, including flavonoids, terpenoids, alkaloids, saponins, and tannins[97]. Conceivably, miRNAs may also exert similar functions in fruit nutrient metabolism, but so far there is no reported research on the explicit function of miRNAs in regulating fruit nutrition. Other studies have reported that plant miRNAs have potential therapeutic functions in the prevention of chronic inflammation-related diseases[98]. A growing number of evidence shows that miRNAs have great benefits in disease treatment[99]. More intriguingly, dietary microRNAs have become one of the hot topics due to their potential health benefits[100]. It becomes increasingly apparent that enhancing fruit quality, in particular, nutritional values, holds high priority for the future of fruit research, where miRNA will definitely play an important and unique role. It will be meaningful to develop fruit varieties rich in specific nutrients through gene editing of specific miRNA pathways, or to directly use dietary miRNAs for targeted disease therapy.
-
Fruit resistance to abiotic or biotic stresses significantly affects their storability and quality. Currently, few studies on the miRNA regulation of fruit stress response have focused on unsuitable temperatures and diseases. Chilling injury often occurs in cold chain logistics used to maintain the freshness of fruits, which usually causes tissue browning and impaired fruit quality. It has been recently reported that miR528 plays a pivotal role through targeting PPO, leading to a ROS surge and subsequent peel browning of banana fruit under cold stress[25,101]. Another study on the effect of miRNAs on the quality of sweet orange under high temperature stress reveals that with the decline of fruit quality, reflected as juice sac granulation, the expression of miR156, miR169, miR171 and miR397 in the samples stored at 30 °C for 6 h all show upregulation compared with the control[102]. In strawberry, a sRNA profiling of red ripening fruit after inoculation with Botrytis cinerea identifies many disease-responsive miRNAs, among which miR5290a can negatively regulate fruit disease resistance by targeting a Plant Ras-associated LRR (PIRL) gene. Functional analysis via transient transformation shows that strawberry fruit overexpressing a miR5290-insensitive PIRL are more resistant to B. cinerea infection than the control[28].
-
A major challenge for the modern fruit industry is to cultivate new fruit varieties with excellent quality to meet growing consumption demand. During the past few decades, with rapid development and reduced cost of next-generation sequencing as well as increasingly powerful bioinformatics tools, many key miRNAs that regulate various fruit traits have been uncovered. Fruits not only share conserved miRNA functions with model plants, but also have evolved unique species-specific miRNA pathways that may confer specific fruit traits. Here we present a table listing representative studies of miRNAs involved in fruit development, quality formation and stress response (Table 1).
Table 1. Representative functional miRNAs characterized in fruits.
Fruit traits Fruit species miRNA Target miRNA regulation Transgenic verification? References Development Fruit set Tomato miR159 GAMYB1/2 Positive yes [34] Seed hardening Pomegranate miR164/172 NAC1/WRKY, MYC Negative no [35] Seed formation Tomato miR393 TIR1/AFB2 Positive yes [40] Seed formation Grape miR159/160 GAMYB/ARFs Negative no [37,38] Ripening and senescence Fruit ripening Tomato miR157 SPL-CNR Negative yes [45] Fruit ripening Tomato miR172 AP2a Positive yes [42] Fruit ripening Tomato miR1917 CTR4 Positive yes [44] Fruit ripening Strawberry Fan-miR73 ABI5 Negative no [46] Fruit ripening Kiwifruit miR164 NAC6/7 Negative yes-transient [17] Fruit senescence Strawberry miR164 NACs Negative no [47] External quality Fruit size Tomato miR156 SPL-CNR Negative yes [32] Fruit size Tomato miR168 AGO1 Positive yes [61] Fruit size Tomato miR171 GRAS24 Positive yes [27] Fruit size Apple miR172 AP2 Negative yes [16] Fruit size Tomato miR396 GRFs Negative yes [59] Fruit size Tomato miR1917 CTR4 Negative yes [60] Fruit shape Tomato miR156 SPLs / yes [12] Fruit shape Tomato miR160 ARF10 / yes [64] Fruit shape Tomato miR166 REV / yes [63] Fruit color Tomato miR858 MYB7-like Negative yes [71] Fruit color Grape miR828/858 MYB114 Positive no [13] Fruit color Apple miR156 SPL2-like/33 Negative yes-transient [77] Fruit color Pear miR156 SPLs Positive no [76] Fruit color Kiwifruit miR858 MYBC1 Negative yes [73] Internal quality Fruit flavor Strawberry miR399 PHO2 Positive yes [14] Fruit flavor Tomato miR-W* glutamate permease Negative no [31] Fruit flavor Camellia oleifera fruit miR477 acetyl-CoA carboxylase carboxyl transferase
subunit alphaNegative no [83] Fruit taste Pear miR397 LAC Positive yes-transient [15] Fruit taste Tomato miR156 SPL-CNR Positive yes [45] Stress response Cold tolerance Banana miR528 PPO Positive no [25] Heat tolerance Orange miR156/169/171/397 not reported Positive no [102] Disease resistance Strawberry miR5290a PIRL Negative yes-transient [28] Given the diverse functions of the fruit miRNAs, it is promising to genetically modify them to obtain better quality. This may be achieved by changing spatiotemporal expression of endogenous miRNAs/target genes, or sequestering specific miRNAs by target mimics, or creating artificial targets of specific miRNAs. But functional studies of miRNAs in non-model fruits are still limited, with most current reports focusing on identification and expression profiling. Certainly, there are many obstacles to overcome. Firstly, our understanding of the miRNA-mediated regulatory network is in its infancy. Up to now, most miRNAs have only been extensively studied in the model fruit tomato, and only a few functional miRNAs have been reported in other fruit species (Table 1). It becomes apparent that the function of conserved miRNAs in a given fruit species cannot be inferred solely on the basis of homology, as neofunctionalization is widespread in the process of speciation. This neofunctionalization may not necessarily result from the miRNA itself, but from the evolution of new functions in miRNA targets and downstream pathways[25]. Obviously, the miRNA neofunctionalization in fruit species with characteristic quality traits will be the focus of future research. Secondly, the genetic redundancy among miRNA family members may prevent their full utilization. Thirdly, conserved miRNAs tend to be pleiotropic, which may lead to the decline of other fruit traits while improving a specific trait. Faced with these obstacles, there are several possible solutions. One possibility is to use tissue- or developmental-specific promoters to precisely control the expression of specific miRNAs for targeting traits that require improvement. Another possibility is to create an artificial crosstalk by introducing a miRNA recognition element in the genetic function of other important pathways. Phylogenetic and functional studies indicate that every existing miRNA may retain a subset of functions from its ancestral MIRNA genes. Therefore, the use of precise genome editing systems can facilitate more sophisticated manipulation of specific miRNA family members and ultimately achieve the goal of accurately regulating a specific fruit trait without causing pleiotropic phenotypes. In fact, the biggest obstacle is that functional analysis in most fruit-bearing species remains challenging due to the lack of fast, efficient and stable transformation systems. However, it is believed that through more investment and effort, there will be breakthroughs in technological innovation in the future such as nano particle-based foliar spray of miRNAs[103]. In conclusion, more extensive bioinformatic interpretation and explicit functional validation will help us better understand the role of fruit miRNAs, which can be used as targets for genetic engineering to improve fruit quality in the future.
- This work is funded by the National Natural Science Foundation of China (31772371, 32071810) and Natural Science Foundation of Guangdong Province (2021A1515011258)
- The authors declare that they have no conflict of interest.
- Copyright: © 2021 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Kong X, He M, Guo Z, Zhou Y, Chen Z, et al. 2021. microRNA regulation of fruit development, quality formation and stress response. Fruit Research 1: 5 doi: 10.48130/FruRes-2021-0005

microRNA regulation of fruit development, quality formation and stress response
- Received Date: 11 May 2021
- Accepted Date: 09 July 2021
- Published Online: 20 July 2021
Abstract: Fruits, as the reproductive organs of many higher plants, are an integral part of a balanced diet, providing rich nutrients and bioactive substances for our health. Over the years, the research on fruit development, quality formation and stress response has deepened, which sheds light on the regulatory mechanism and application of fruit quality improvement. Among the many regulatory factors, microRNAs (miRNAs) are a class of short non-coding RNAs, ranging from 20 to 24-nt, which post-transcriptionally regulate target gene expression. miRNAs and their functions have been extensively examined in plants especially in model species, and they play critical roles in the regulation of diverse biological processes in response to endogenous developmental signals and external environmental cues, respectively. In fruit crops, the function of miRNAs and their regulation have also been under intensive study. In particular, the novel roles of miRNAs that have not been revealed in the model annual species have been unraveling, which reflect the genetic, physiological, and developmental complexity of gene regulation in fruit crops. Here we review the current research progress achieved, specifically in fruit crops, with a focus on the function of miRNAs in the control of fruit development and quality as well as response to various stresses. The future prospects of miRNAs for quality-targeted fruit breeding are also discussed.
-
Key words:
- fruits /
- miRNA /
- morphogenesis /
- quality /
- stress response