

Abstract
Objective. Hydroxychloroquine (HCQ) is a commonly used medicine for the treatment of systemic lupus erythematosus (SLE), and Th17 cells are closely related to the pathogenesis of SLE. However, the role and mechanism of HCQ on Th17 cell differentiation in SLE is not clearly understood. Here, we investigate the effect of HCQ on Th17 cell differentiation both in vitro and in patients with SLE.
Methods. Twenty-five patients with SLE were divided into 2 treatment groups: prednisone alone and HCQ plus prednisone. Interleukin 17 (IL-17) expression was analyzed by ELISA and real-time (RT)-PCR. Th17 were measured in patients with SLE by flow cytometry before and after HCQ treatment. In vitro, naive T cells were cultured in Th17-inducing conditions with or without HCQ. Cell differentiation and IL-17 expression were analyzed. Finally, transcriptome sequencing identified differential gene expression between naive T cells and induced Th17 cells.
Results. In patients, HCQ plus prednisone treatment inhibited IL-17 production, gene expression, and Th17 cell differentiation. In vitro, HCQ inhibited Th17 cell proliferation and differentiation, as well as IL-17 production. Five microRNA were significantly different in Th17 cells compared with naive T cells, and HCQ treatment reversed this effect. In vivo, microRNA-590 (miR-590) was verified and was significantly decreased in Th17 cells, compared with naive T cells from lupus-prone mice. Moreover, miR-590 was increased in patients treated with HCQ plus prednisone.
Conclusion. HCQ inhibited Th17 cell differentiation and IL-17 production both in vitro and in patients with SLE. Our study provides additional evidence for HCQ as a treatment for SLE.
Systemic lupus erythematosus (SLE) is a complex autoimmune disease whose pathogenesis is not yet clearly understood. Immune imbalance is closely related to both SLE occurrence and the degenerative characteristic of the disease1. Moreover, it is generally accepted that activation and expansion of effector T cells or B cells occur as part of SLE, making these cells effective therapeutic targets for SLE.
The Th17 lineage, a subset of effector CD4+ T cells characterized by the production of interleukin (IL)-17, exhibits developmental and functional features that are distinct from classic Th1 and Th2 lineages2,3. Th17 cells are associated with the development and pathogenesis of a growing list of chronic inflammatory diseases, including rheumatoid arthritis (RA), systemic sclerosis, and psoriatic arthritis4,5,6. Our previous studies, as well as those of others, have proved that Th17 cells are increased in the peripheral blood of patients with SLE, and that Th17 cells are closely related to nephritis, vasculitis, central nervous system infection, and the production of autoantibodies in patients with SLE7,8,9,10. Further, it has gradually been recognized that Th17 cells are a promising therapeutic target for chronic inflammatory injury11,12, and neutralized antibodies against IL-17 and the IL-17 receptor have been developed to treat diseases such as psoriasis, arthritis, and ankylosing spondylitis6,13,14.
Hydroxychloroquine (HCQ) is a medicine commonly used to treat or prevent malaria; however, it is also an antirheumatic medicine that can alleviate symptoms associated with RA, discoid lupus, or SLE15,16. Accordingly, HCQ is widely used in clinical settings and has become an essential drug for the treatment of SLE. In addition, previous studies have shown that HCQ can interfere with antigen processing and presentation, reduce cutaneous reactions to UV light, alleviate oxidative stress, and inhibit the production of proinflammatory cytokines such as IL-6, IL-1β, and tumor necrosis factor (TNF)-α15,17. However, neither the effect of HCQ treatment on Th17 cells and regulatory T (Treg) cells in patients with SLE, nor the effect of HCQ on the differentiation of Th17 and Treg cells in vitro, are currently known.
In our study, we aimed to investigate the effect of HCQ on Th17 cells and cytokine production, both alone and in combination with the corticosteroid drug prednisone. We demonstrate that treatment with HCQ, combined with prednisone, can inhibit Th17 cell differentiation and production of IL-17 (a key cytokine produced by Th17 cells) in vivo, when compared with prednisone treatment alone. In addition, HCQ treatment prevented Th17 cell differentiation and proliferation, as well as IL-17 production in vitro. The results of transcriptome sequencing revealed that microRNA (miR)-590 is sensitive to HCQ treatment, and closely related to Th17 cell differentiation. Further, we investigated the role of miR-590 in Th17 cells by using an autoimmune, lupus-prone mouse model (MRL/lpr mice). The data showed that decreased expression of miR-590 was verified in induced Th17 cells, compared with naive T cells, from lupus-prone MRL/lpr mice. The expression of miR-590 could be upregulated in patients with SLE following treatment with HCQ. Our data indicate that HCQ treatment can inhibit Th17 cell differentiation both in vitro and in vivo, and that miR-590 may be an additional therapeutic target for the treatment of SLE.
MATERIALS AND METHODS
Patient selection and healthy controls
Our study was approved by the Ethics Committee of Zhongshan Hospital, and Huashan Hospital, Fudan University (2015-9-02). All patients enrolled in the study after giving informed and written consent. The animal study was approved by the institutional animal care and use committee of Zhongshan Hospital, Fudan University (ZS17018). Twenty-five adult patients (22 women; 3 men; mean age 28.3 ± 7.5 yrs) with a diagnosis of SLE, based on the American College of Rheumatology 1997 revised criteria18, were included in the study. All patients were referred to either the Department of Dermatology in Zhongshan Hospital or the Division of Rheumatology in Huashan Hospital at Fudan University (Shanghai, China) and were enrolled in the study after giving their informed and written consent. Patients with SLE were divided into 2 groups: 1 group received prednisone treatment (n = 10; 8 women, 2 men; mean age 30.7 ± 9.6 yrs), while the second group received prednisone plus HCQ treatment (n = 15; 14 women, 1 man; mean age 26.7 ± 5.4 yrs). A control group was also included, consisting of 10 age- and sex-matched healthy individuals, who were enrolled after giving their informed, written consent.
T cell isolation, culture conditions, and Th17 differentiation
Human naive T cells were purified from peripheral blood mononuclear cells (PBMC) of healthy donors, according to the manufacturer’s instructions (Miltenyi Biotec). To stimulate differentiation into Th17 cells, purified T cells (2 × 106 cells/ml) were cultured for 6 days in RPMI 1640 (Gibco) containing anti-CD3/CD28 beads (5 μl/ml; Life Dynabeads), IL-1β (50 ng/ml), transforming growth factor (TGF)-β (1 ng/ml), IL-6 (30 ng/ml), IL-23 (5 ng/ml), anti-IL-4 antibodies (5 ng/ml), anti-IL-12 antibodies (5 ng/ml), and anti-IL-2 antibodies (20 U/ml). All cytokines and antibodies were from Peprotech. For some experiments, 20 μM of HCQ (Abcam) was added to the culture media according to previous reports19.
Flow cytometry
To detect Th17 cells in the PBMC of patients with SLE, PBMC were isolated from patients and incubated for 5 h with 50 ng/ml phorbol 12-myristate 13-acetate and 750 ng/ml ionomycin in the presence of 20 μg/ml monensin (Sigma-Aldrich). Surface staining was performed with PE-Cy5-conjugated anti-CD3 and FITC-conjugated anti-CD8 (BD Pharmingen) for 15 min. Then, cells were re-suspended in fixation/permeabilization solution, according to the manufacturer’s instructions (Invitrogen Caltag), and intracellular staining with PE-conjugated anti-IL-17 was performed, also according to the manufacturer’s protocol (eBioscience). Finally, IL-17+ cells were analyzed with flow cytometry by applying a CD3+CD8− gate.
For some experiments, naive T cells were cultured in Th17 cell culture conditions (described above) and treated with 20 μM HCQ for 6 days. Then, staining was performed with FITC-conjugated anti-CD4 and PE-conjugated anti-IL-17, prepared according to the manufacturer’s protocol (eBioscience). IL-17+ cells were sorted from CD4+ T cells by flow cytometry. For some experiments, naive T cells were labeled with carboxyfluorescein diacetate succinimyl ester (CFSE), and cultured in Th17 cell culture conditions with or without 20 μM HCQ for 6 days, and the percentage of CFSE+ cells was analyzed by flow cytometry.
Transcriptome sequencing
Human naive T cells were purified from the PBMC of healthy donors, according to the manufacturer’s instructions (Miltenyi Biotec). To promote differentiation into Th17 cells, purified T cells (2 × 106 cells/ml) were cultured in Th17 culture conditions (as described above) in the presence or absence of 20 μM HCQ for 6 days.
The study consisted of 3 groups (n = 3 replicates/group): naive CD4+ T cells, Th17 cells without HCQ treatment, and Th17 cells with HCQ treatment. Total RNA was extracted from each group and mRNA was enriched using oligo (dT) magnetic beads, cDNA synthesis, and gene amplification by PCR, and then detected by sequencing (Illumina). Screening of differentially expressed genes was performed using cluster and pathway enrichment analyses. These procedures were performed by the Beijing Genomics Institute (Shenzhen, China).
ELISA
Sera were collected from patients with SLE and healthy controls, and IL-17 concentrations were determined by ELISA (eBioscience). In addition, isolated T cells were cultured in Th17 cell culture media with or without HCQ (20 μM) for 6 days, and IL-17 was similarly detected in the supernatants by ELISA.
Mice
Female MRL/lpr mice (12 weeks of age) were purchased from the Shanghai SLAC laboratory animal company. The animal study was approved by the institutional animal care and use committee of Zhongshan Hospital, Fudan University (ZS17018). All mice were maintained under pathogen-free conditions. For some experiments, naive CD4+ T cells were sorted from the spleen of mice (Miltenyi Biotec). To stimulate differentiation into Th17 cells, purified T cells (2 × 106 cells/ml) were cultured in media containing anti-CD3/CD28 beads (5 μl/ml; Life Dynabeads), TGF-β (1 ng/ml, Peprotech), IL-6 (30 ng/ml, Peprotech), and IL-1β (50 ng/ml, Peprotech) for 5 days, and the miRNA expression was analyzed by quantitative real-time PCR (qRT-PCR).
Analysis of gene expression using PCR
Total RNA was purified from the PBMC of patients and healthy controls using Trizol reagent (Invitrogen). Synthesis of cDNA was performed using Primescript RT Master Mix Perfect Real-time Kit (TaKaRa), and mRNA expression was determined with a Bio-Rad iCycler 7500 Optical System (Bio-Rad) using a SYBR Premix EX Taq Real-time PCR Master Mix (TaKaRa). The 2-ΔΔCt method was used to normalize transcription to β-actin and to calculate the fold change relative to controls. The following primer pairs were used: Hum β-actin (forward: ATC ATG TTT GAG ACC TTC AAC A; reverse: CAT CTC TTG CTC GAA GTC CA), Hum IL-17A (forward: AAA GTG GCC CGG ATG TGA GA; reverse: GAC ATT GTG CCC TGC CCT TCT), retinoic acid receptor–related orphan receptor gamma t (ROR-gamma-t; forward: TGA GAA GGA CAG GGA GCC AA; reverse: CCA CAG ATT TTG CAA GGG ATC A), and forkhead box P3 (FOXP3; forward: GAA ACA GCA CAT TCC CAG AGT TC; reverse: ATG GCC CAG CGG ATG AG). To detect miR-590 expression in the PBMC of patients with SLE and in the spleens of mice, the 2-ΔΔCt method was used to normalize the expression level of miR-590 to U6 RNA. Primers were provided by Ruibo.
Statistical analyses
Results are expressed as the mean ± SD. Statistical significance was determined using ANOVA for comparisons of multiple means, followed by the Student t test, and p < 0.05 was considered significant.
RESULTS
HCQ inhibits Th17 cell differentiation and IL-17 production in patients with SLE
We first investigated the effect of either prednisone treatment alone or prednisone combined with HCQ on the production of IL-17 in the serum of patients with SLE. The results of ELISA showed that HCQ plus prednisone inhibits IL-17 production (Figure 1A) compared with prednisone treatment alone. Similarly, RT-PCR results showed that IL-17 mRNA expression in the PBMC of patients with SLE was also decreased by the combined treatment, when compared with prednisone treatment alone (Figure 1B). In addition, we assessed the expression of FOXP3, because it is a key transcription factor in the differentiation of Treg cells20. We found that HCQ plus prednisone treatment did not effectively increase FOXP3 gene expression, when compared with prednisone treatment only (Figure 1C).

HCQ inhibits IL-17 production in patients with SLE. A. Concentration of IL-17 in the serum of patients with SLE was assessed by ELISA following either prednisone treatment alone (n = 10) or HCQ plus prednisone treatment (n = 15), and compared with healthy controls (n = 10). B. The expression of IL-17 mRNA in the PBMC of patients with SLE was measured by RT-PCR following either prednisone treatment (n = 10) or HCQ plus prednisone treatment (n = 15), and compared with healthy controls (n = 10). C. The expression of FOXP3 was assessed by RT-PCR in the PBMC of patients with SLE following treatment with either prednisone alone (n = 10) or HCQ plus prednisone treatment (n = 15), and compared with healthy controls (n = 10). HCQ: hydroxychloroquine; SLE: systemic lupus erythematosus; IL: interleukin; PBMC: peripheral blood mononuclear cells; RT-PCR: real-time PCR; Pred: prednisone; FOXP3: forkhead box P3.
To provide additional evidence, 6 patients with SLE were dynamically followed before and after HCQ treatment. An analysis by flow cytometry showed that after 4 weeks of treatment with HCQ plus prednisone, the percentage of Th17 cells was inhibited, compared with initial values (Figure 2A). Interestingly, the percentage of Treg cells was increased following treatment with HCQ plus prednisone, compared with the starting values (Figure 2B). We next performed ELISA and found that the production of IL-17, in the serum of patients with SLE, was decreased following 4 weeks of the combined treatment (Figure 2C). Finally, the expression of FOXP3 mRNA was increased following 4 weeks of treatment with HCQ plus prednisone, compared with the starting values (Figure 2D). These data together indicate that HCQ treatment plus prednisone can inhibit Th17 cell expansion and IL-17 production in patients with SLE.
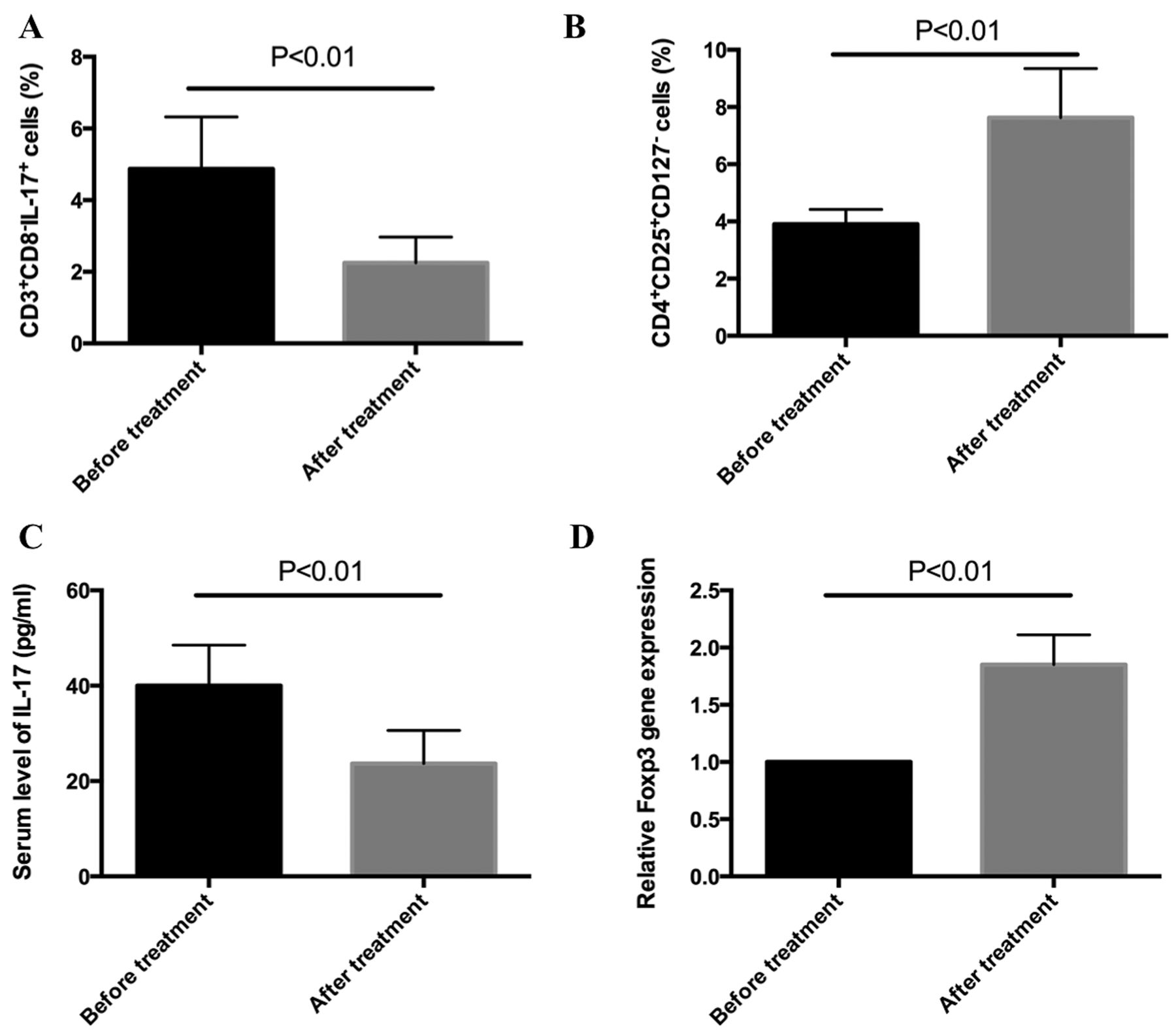
HCQ inhibits Th17 cells in patients with SLE. A. The percentage of CD3+CD8–IL-17+ Th17 cells was analyzed by flow cytometry in patients with SLE both before and after 4 weeks of treatment with HCQ plus prednisone treatment (n = 6 for each group). B. The percentage of CD4+CD25+CD127–Treg cells was analyzed by flow cytometry in patients with SLE both before and after 4 weeks of treatment with HCQ plus prednisone (n = 6 for each group). C. The concentration of IL-17 in the serum of patients with SLE was analyzed by ELISA both before and after 4 weeks of treatment with HCQ plus prednisone (n = 6 for each group). D. The expression of FOXP3 mRNA in PBMC of patients with SLE was analyzed by RT-PCR both before and after 4 weeks of treatment with HCQ plus prednisone (n = 6 for each group). HCQ: hydroxychloroquine; SLE: systemic lupus erythematosus; IL: interleukin; RT-PCR: real-time PCR; FOXP3: forkhead box P3; PBMC: peripheral blood mononuclear cells.
HCQ inhibits Th17 cell differentiation and IL-17 production in vitro
We next investigated whether HCQ inhibits Th17 cell differentiation in vitro. For this, naive T cells were sorted from the PBMC of healthy controls and cultured in Th17-inducing cell culture conditions with or without HCQ. The percentage of CD4+IL-17+ Th17 cells was analyzed by flow cytometry, which revealed that 20 μM HCQ inhibited CD4+IL-17+ Th17 cell differentiation (Figure 3A). In addition, we found that HCQ inhibited IL-17 production and gene expression (Figure 3B and 3C). ROR-gamma-t, a key transcript factor for differentiation of Th17 cells, was also inhibited by HCQ treatment (Figure 3D). HCQ could time-dependently prevent CD4+IL-17+ Th17 cell expansion (Figure 3E), and the combination of HCQ and prednisone enhanced the inhibitory effect on CD4+IL-17+ Th17 cell differentiation (Supplementary Figure 1A, available from the authors on request), and induced the apoptosis of Th17 cells (Supplementary Figure 1B, available from the authors on request). In addition, HCQ could inhibit CFSE-labeled Th17 cell proliferation (Figure 3F and 3G). However, HCQ did not promote FOXP3 gene expression and Treg cell differentiation in vitro (data not shown). These data indicate that HCQ treatment could inhibit Th17 cell differentiation and IL-17 production. Our further data showed that HCQ treatment could also inhibit Th1/Th2-related cytokines interferon (IFN)-γ and IL-4 production (Supplementary Figure 2, available from the authors on request).

HCQ inhibits Th17 cell differentiation and IL-17 production in vitro. Naive CD4+ T cells were sorted from PBMC of healthy controls and cultured for 6 days in conditions designed to induce Th17 differentiation, either with or without 20 μM HCQ treatment. A. The percentage of CD4+IL-17+ Th17 cells was analyzed by flow cytometry (n = 3 for each group). B. The concentration of IL-17 in supernatants was analyzed by ELISA (n = 3 for each group). C. Expression of IL-17 mRNA was analyzed by RT-PCR (n = 3 for each group). D. Naive CD4+ T cells were cultured for 12 days in conditions designed to induce Th17 differentiation, with or without 20 μM HCQ; the gene expression of retinoic acid receptor–related orphan receptor gamma t (RORγt) was analyzed by RT-PCR. E. The number of CD4+IL-17+ cells could be induced in a time-dependent manner, and 20 μM HCQ could time-dependently inhibit CD4+IL-17+ cell proliferation (n = 3 for each group). These experiments were performed 3 times with similar results. F. Sorted naive CD4+ T cells were labeled with CFSE and cultured for 6 days in conditions designed to induce Th17 cell differentiation, with or without 20 μM HCQ. CFSE+ Th17 cells were analyzed by flow cytometry. G. The percentage of CFSE+ among CD4+ cells. Results shown are representative of at least 3 independent experiments. HCQ: hydroxychloroquine; IL: interleukin; RT-PCR: real-time PCR; PBMC: peripheral blood mononuclear cells; CFSE: carboxyfluorescein diacetate succinimyl ester.
Differential gene expression in Th17 cells with and without HCQ treatment
To comprehensively and systematically investigate gene expression in Th17 cells, transcriptome sequencing was performed and differential gene expression profiles were compared between naive T cells, Th17 cells with HCQ treatment, and Th17 cells without HCQ treatment. Our data revealed that 1151 genes were upregulated and 661 genes were downregulated in Th17 cells, when compared with naive CD4+ T cells (Figure 4A). A pathway analysis revealed that pathways for processes, such as cell cycle, human T cell lymphotrophic virus type 1 infection, and Epstein-Barr virus infection, were all involved in the differentiation of Th17 cells, when compared with naive CD4+ T cells (Figure 4B). Finally, a clustering analysis showed that HCQ treatment affects the expression of a large number of genes in Th17 cells (Figure 4C).
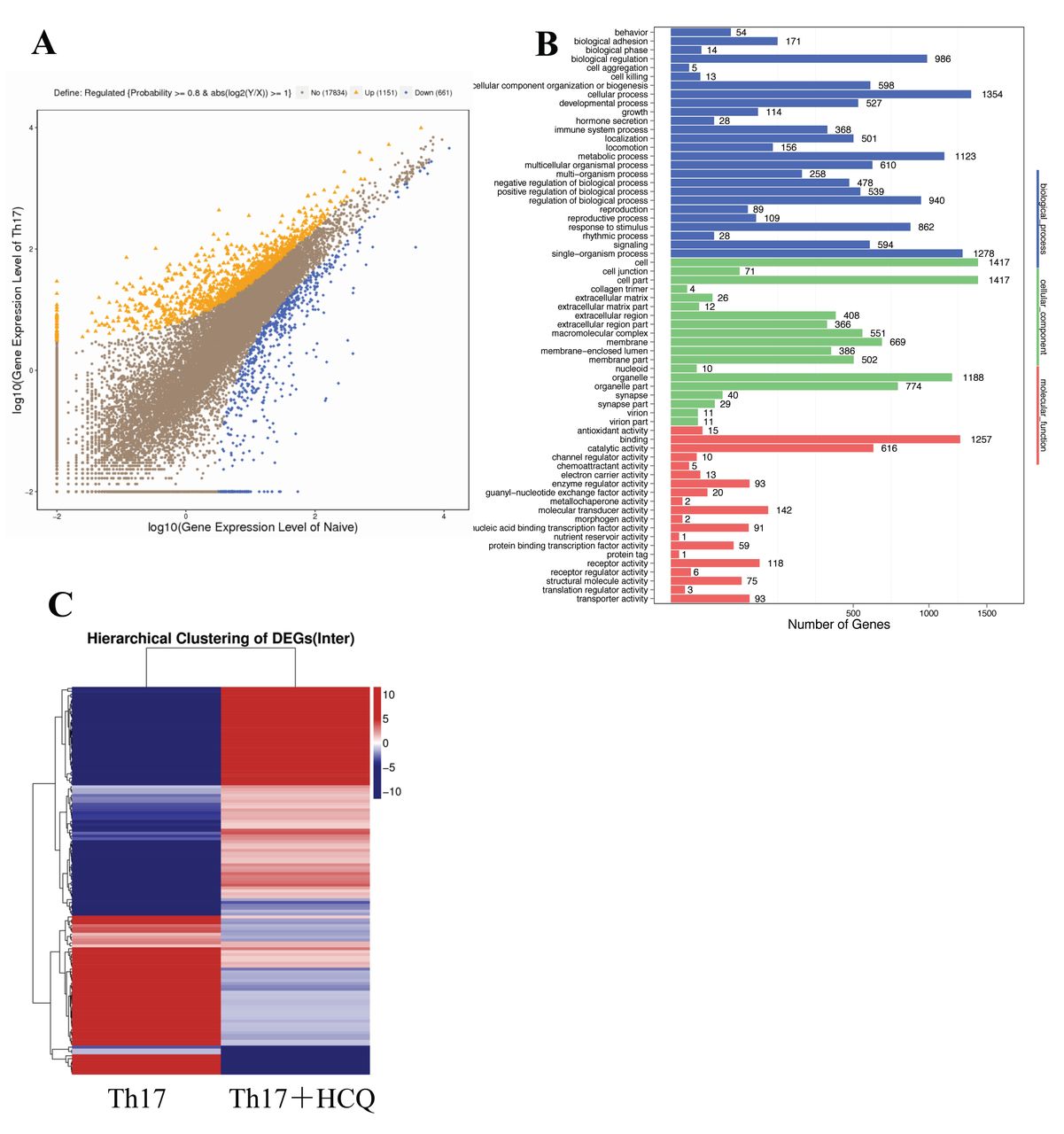
Gene expression profiles of Th17 cells with or without HCQ treatment. Naive CD4+ T cells were sorted from PBMC of healthy controls and cultured for 6 days in conditions designed to induce Th17 differentiation, either with or without HCQ treatment. The expression of the entire genome was then analyzed by transcriptome sequencing. A. Differential gene expression between induced Th17 cells and naive CD4+ T cells. B. The main pathways activated in Th17 cells are displayed, when compared with naive CD4+ T cells. C. A clustering analysis of gene expression between Th17 cells with and without 20 μM HCQ treatment. Blue represents downregulation and red represents upregulation. All data in this figure are the results of 3 independent experiments. HCQ: hydroxychloroquine; PBMC: peripheral blood mononuclear cells; DEG: differentially expressed gene.
The relationship between miR-590 expression and Th17 cell differentiation in the pathogenesis of SLE
Using transcriptome sequencing, we identified 5 miRNA whose expressions were significantly changed in Th17 cells when compared with naive CD4+ T cells: miR-4426, miR-3615, miR-106B, miR-590, and miR-573. Most notably, we found that HCQ treatment reversed the expression of these miRNA, suggesting that they might be related to Th17 cell differentiation and are sensitive to HCQ treatment (Figure 5A).

Expression of miR-590 mRNA both in vitro and in vivo. A. Naive CD4+ T cells were sorted from PBMC of healthy controls and cultured for 6 days in conditions designed to induce Th17 differentiation, either with or without HCQ treatment. The expression of 5 microRNA was analyzed by transcriptome sequencing (n = 3 for each group). B. Naive CD4+ T cells were isolated from the spleens of an SLE mouse model (MRL/lpr mice) and cultured in the presence of vehicle or TGF-β, IL-6, and IL-1β for 5 days. Then, miR-590-5p and (C) miR-590-3p expressions were analyzed by qPCR (n = 3 for each group). D. PBMC were isolated from patients and treated with prednisone or prednisone plus HCQ. The expression of miR-590-3p and (E) miR-590-5p were analyzed by qPCR (n = 10 for each group). miR-590: microR-590; PBMC: peripheral blood mononuclear cells; HCQ: hydroxychloroquine; SLE: systemic lupus erythematosus; TGF: transforming growth factor; IL: interleukin; qPCR: quantitative PCR.
IL-6 is one of the most important cytokines for the differentiation of Th17 cells. Previous reports have indicated that miR-590 is involved in IL-6–mediated inflammatory injuries, and that IL-6 can inhibit miR-590 expression21,22. Here, we sorted naive CD4+ T cells from the spleens of MRL/lpr mice, and cultured them in the presence of vehicle or TGF-β, IL-6, and IL-1β for 5 days. Then, miRNA expression was analyzed by qRT-PCR. We found that the expressions of miR-590-5p and miR-590-3p were significantly decreased in Th17 cells, when compared with naive CD4+ T cells (Figure 5B and 5C). In addition, we verified the expression of miR-590 in the PBMC of patients with SLE with and without HCQ treatment. The data showed that the expression of both miR-590-5p and miR-590-3p was upregulated in patients with SLE treated with HCQ plus prednisone, compared with prednisone treatment alone (Figure 5D and 5E). Taken together, our data indicate a relationship between miR-590 and Th17 cell differentiation, as well as sensitivity to HCQ treatment.
DISCUSSION
Th17 cells are a proinflammatory subset of Th cells that play key roles in the pathogenesis of autoimmune diseases23. Both the numbers of Th17 cells and IL-17 production have been found to be increased in the peripheral blood of patients with SLE, and expanded Th17 cells have been detected in the skin lesions, lungs, and kidneys of these patients8,10. In addition, Th17 cells are increased in SLE-prone mouse models, including BXD2, SNF1, NZB×NZW F1, Ro52 knockout mice, and MRL/lpr mice8,24,25,26. The potent proinflammatory activities of Th17 cells could contribute to numerous pathological features of SLE, such as the induction of vascular inflammation, recruitment of leukocytes, activation of B cells, and autoantibody production27,28. Thus, it has been speculated that inhibition of Th17 cells or antagonism of IL-17 might be promising therapeutic steps for the treatment of SLE28. Here, we showed that HCQ plus prednisone treatment inhibited Th17 cell differentiation and IL-17 production in patients with SLE, compared with prednisone treatment alone. Moreover, we found that Th17 cells and IL-17 production are inhibited by 4 weeks of HCQ treatment. Finally, our in vitro study further confirmed that HCQ treatment inhibits Th17 cell differentiation, cell number proliferation, and IL-17 production. Interestingly, the combined use of HCQ and prednisone enlarged the inhibitory function on the differentiation of Th17 cells, and could induce the apoptosis of Th17 cells. Therefore, we have provided additional evidence for HCQ as an efficacious treatment for SLE, and suggest that it may act to reduce Th17 cells and alleviate subsequent inflammatory reactions. These data were consistent with findings that HCQ could reduce Th17-related cytokines such as IL-6, IL-17, and IL-22 production from PBMC in SLE29. Further, chloroquine almost inhibited activation of all Th cell subsets: secretion of IFN-γ by Th1 cells and IL-17 by Th17 cells was strongly decreased by chloroquine. IL-4 and IL-13 secretion by Th2 cells was markedly decreased by chloroquine30. Here, our data further proved that HCQ could also inhibit the production of Th1/Th2 cytokines from SLE patient–derived PBMC.
Treg cells can modulate the functioning of effector T cells, thereby maintaining immunologic homeostasis and preventing autoimmunity31. Circulating Treg cell numbers are known to decrease during flares in patients with SLE, and the immunosuppressive functions of Treg cells are impaired in SLE32,33. Our study showed that both the percentage of Treg cells and the expression of the FOXP3 gene were upregulated in vivo after combined treatment with prednisone and HCQ, suggesting that HCQ might promote the differentiation of Treg cells. However, parallel-controlled study between prednisone and prednisone plus HCQ treatment groups did not find significant FOXP3 differential expression, and our unpublished in vitro data showed that HCQ did not promote Treg cell differentiation and FOXP3 gene expression, but rather inhibits them. Thus, we speculate that the upregulation of Treg cells and FOXP3 gene expression in vivo might be due to the effect of prednisone, and could contribute to recovery and remission of disease activity.
MicroRNA are endogenous, small noncoding RNA that act as negative regulators of gene expression by degrading mRNA or repressing translation34. Moreover, microRNA have been implicated in the pathogenesis of many diseases through their regulation of processes such as cell proliferation, differentiation, and morphogenesis34. It has been reported that microRNA, such as miR-155, miR-21, miR-301, miR-326, miR-10a, and miR-20b, contribute to positive or negative Th17 cell differentiation35,36. A number of studies have confirmed that the inhibition or knockdown of miR-155 can alleviate alveolar hemorrhage in SLE mice, reduce autoantibodies, and alleviate lupus nephritis37,38. Based on the important role of microRNA in regulating Th17 cell differentiation, microRNA could also represent a therapeutic target for the treatment of Th17 cell–related SLE.
To discover new mechanisms for the regulation of Th17 cell differentiation, we analyzed whole genomes to identify differentially expressed genes in Th17 cells, using high throughput transcriptome sequencing. A total of 1151 genes were upregulated and 661 genes were downregulated in the induced Th17 cells. Among them, we found 5 novel microRNA that are closely related to Th17 cell differentiation: miR-4426, miR-3615, and micro miR-106B were downregulated, whereas miR-590 and miR-573 were upregulated in Th17 cells. Importantly, HCQ treatment promoted the expression miR-4426, miR-3615, and miR-106B, and inhibited the expression of miR-590 and miR-573. We decided to focus on miR-590, because it has been reported that miR-590 negatively regulates IL-6–mediated inflammatory cytokine production, and IL-6 can inhibit miR-590 expression. The data indicated that IL-6 can inhibit miR-590 expression, and that upregulation of miR-590 can block inflammatory cytokines production21,22. This partly explains the decreased expression of miR-590 in IL-6–induced Th17 cells in our study. Recent studies have proved that miR-590-3p mimic could diminish proinflammatory IL-18 production and reduce angiogenesis in the autoimmune disease of RA39. Upregulation of miR-590 could inhibit IL-6 and TNF-α production in vivo of experimental autoimmune myocarditis22, and miR-590 could attenuate proinflammatory IL-6, TNF-α, and IL-1β production in macrophages21. One study reports that miR-590-3p mimics could also alleviate the intracellular oxidative stress caused by angiotensin II, and function as antiinflammatory effects for endothelial cell damage40. These data suggested that miR-590 exerts anti-inflammatory function in the inflammatory disease. No studies, to our knowledge, have reported a link between miR-590 and Th17 cell differentiation and IL-17 production. Previous studies showed that IL-6 could inhibit miR-590 expression, and upregulation of miR-590 reversely inhibited IL-6 production21,22, which indicates that miR-590 might be negative related to IL-6 production. IL-6 is one of the key cytokines for promoting Th17 cell differentiation; thus, upregulation of miR-590 might inhibit IL-6 production, and subsequently block Th17 cell differentiation and IL-17 production. Therefore, miR-590 might be a promising therapeutic target for the treatment of Th17-related autoimmune diseases.
Here, we further verified the expression of miR-590 in naive T cells from MRL/lpr mice and induced Th17 cells, and found that the expression of miR-590 was decreased after differentiation into Th17 cells. Interestingly, our in vivo study confirmed that miR-590 expression is increased after treatment with HCQ in our SLE patient group. Taken together, our data indicate that miR-590 may play a negative regulatory role in Th17 cell differentiation, and that it is sensitive to HCQ treatment. Further studies are needed to verify both the role and underlying mechanism of miR-590 in the differentiation of Th17 cells and in the pathogenesis of SLE.
Our study indicates that HCQ can inhibit Th17 cell differentiation and IL-17 production both in vitro and in patients with SLE. We report that miR-590 is sensitive to HCQ treatment and that it might negatively regulate Th17 cell differentiation, which could be a therapeutic target for the treatment of Th17-related autoimmune diseases.
Footnotes
Supported by grants from the National Natural Science Foundation of China (81472874; 81401346), Medical Guide Project from Shanghai Municipal Science and Technology (134119a8400), Cultivation Plan of Young Doctor of Shanghai (2014), and Outstanding Talent Plan of Zhongshan Hospital, Fudan University (2015ZSYXGG13). This work was also supported by grants from Shanghai Public Health Construction Projects Foundation (Grant No. 15GWZK0501), and the Foundation of Shanghai Municipal Commission of Health and Family Planning (Grant No. 201640096).
- Accepted for publication December 11, 2017.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}