

Abstract
Objective. The dysregulation of the NLRP3 (NLR containing a pyrin domain) inflammasome is involved in autoinflammatory diseases. Adult-onset Still disease (AOSD) is regarded as an autoinflammatory disease. However, the pathogenic involvement of NLRP3 inflammasome in AOSD remains unclear and NLRP3 activators in AOSD are currently unknown.
Methods. The mRNA expression of NLRP3 inflammasome signaling in peripheral blood mononuclear cells (PBMC) from 34 patients with AOSD and 14 healthy subjects was determined using quantitative-PCR (qPCR). The changes in mRNA and protein levels of NLRP3 inflammasome signaling in PBMC treated with the potential activator [imiquimod (IMQ)] or inhibitor of NLRP3 were evaluated using qPCR and immunoblotting, respectively. The supernatant levels of interleukin (IL)-1β and IL-18 were determined by ELISA.
Results. Significantly higher mRNA levels of NLRP3 inflammasome signaling were observed in patients with AOSD compared with healthy controls. NLRP3 expressions were positively correlated with disease activity in patients with AOSD. IMQ (an effective Toll-like receptor 7 ligand; 10 µg/ml and 25 µg/ml) stimulation of PBMC from patients with AOSD induced dose-dependent increases of mRNA expression of NLRP3 (mean ± standard error of the mean, 2.06 ± 0.46 and 6.05 ± 1.84, respectively), caspase-1 (1.81 ± 0.23 and 4.25 ± 0.48), IL-1β (5.68 ± 1.51 and 12.13 ± 3.71), and IL-18 (2.32 ± 0.37 and 4.81 ± 0.51) compared with controls (all p < 0.005). IMQ stimulation of PBMC from patients similarly induced greater increases in protein expressions of NLRP3 inflammasome compared with controls. The protein expressions of NLRP3, IL-1β, and IL-18 on PBMC significantly decreased after treatment with NLRP3 inhibitor in patients with AOSD.
Conclusion. Increased expression of NLRP3 inflammasome and its positive correlation with disease activity in AOSD suggest its involvement in disease pathogenesis. IMQ upregulated expressions of NLRP3 inflammasome signaling, and IMQ might be an activator of NLRP3 inflammasome in AOSD.
The innate immune system encompasses the germ-line-encoded pattern recognition receptors, including Toll-like receptors (TLR) and cytosol-expressed NLR1,2. Various levels of crosstalk between these pathways have been identified3,4.
NLR containing a pyrin domain (NLRP) is the major component of inflammasome, which has a pathogenic involvement in innate immunity and inflammation5,6,7. Activation of NLRP by stimuli such as ATP recruits caspase-1, which cleaves pro-interleukin (IL)-1β and pro-IL-18 into their active biologic forms5,6,7,8. Increasing evidence has indicated that the dysregulation of NLRP3 inflammasome is involved in autoinflammatory diseases9,10.
Adult-onset Still disease (AOSD), a systemic inflammatory disorder, is characterized by spiking fever, rash, arthritis, hepatosplenomegaly, lymphadenopathy, variable multisystemic involvement, and increases in acute-phase reactants11,12,13. AOSD has been considered an autoinflammatory disease because of its typical phenotypes without significant increase of autoantibodies levels, similar to other autoinflammatory diseases14,15. In addition, rapid response to IL-1 receptor antagonist anakinra observed in patients with refractory AOSD16,17 suggested an important involvement of IL-1–related inflammasome in the pathogenesis of this disease. However, the pathogenic involvement of NLRP3 inflammasome in AOSD remains unclear, and the precise activators of NLRP3 in AOSD are still currently unknown.
Several activators of NLRP3 inflammasome, such as crystalline particulates, have been identified18. We have shown that TLR7 activation, which leads to enhanced production of proinflammatory cytokines through myeloid differentiation primary-response protein 88 (MyD88)–dependent signaling, may be involved in AOSD pathogenesis19. Moreover, Kanneganti, et al demonstrated that TLR7 ligand was an activator of the NLRP3 inflammasome in a murine model20. In addition, an antagonist of TLR7/8/9 could suppress lupus-associated inflammation and IL-1β levels in NZBW/F1 mice21. Given that imiquimod (IMQ) has been shown to be an effective ligand for TLR7, capable of activating proinflammatory responses22,23, we hypothesized that IMQ, a TLR7 ligand, might activate NLRP3 inflammasome and enhance the production of downstream cytokines in AOSD. However, there was no related study that analyzed the involvement of IMQ in the inflammasome-mediated inflammation in AOSD.
In our study, we investigated (1) whether the expressions of NLRP3 inflammasome components and downstream cytokines differed significantly between patients with AOSD and healthy subjects; (2) the association of NLRP3 inflammasome expressions with disease activity in patients with AOSD; and (3) whether TLR7 ligand IMQ affects the expressions of NLRP3 inflammasome signaling in patients with AOSD.
MATERIALS AND METHODS
Subjects
Thirty-four patients with active AOSD were enrolled, each fulfilling the Yamaguchi criteria24. Patients with infections, malignancies, or other rheumatic diseases were excluded. The disease activity of each patient with AOSD was assessed using a modified Pouchot score described by Rau, et al25. After initial investigation, all patients were treated with corticosteroids, and 30 (88.2%) of them also received at least 1 synthetic disease-modifying antirheumatic drug (sDMARD; Table 1). Because there is no IL-1 inhibitor available in Taiwan, 5 patients with refractory AOSD were then treated with IL-6 receptor inhibitor [tocilizumab (TCZ)], which has shown effectiveness in the treatment of this disease26,27. Fourteen healthy volunteers who had no rheumatic disease were enrolled as control subjects. The Institutional Review Board of our hospital approved this study (CF11309), and each participant’s written consent was obtained according to the Declaration of Helsinki.
Demographic data and mRNA expression levels of NLRP3 inflammasome components as well as downstream cytokines in active patients with AOSD and HC. Values are n (%) or median (interquartile range) unless otherwise specified.
Determination of mRNA expressions of NLRP3 inflammasome signaling using quantitative PCR (qPCR)
Because NLRP were reported to be expressed in a wide range of cell types, including macrophages, lymphocytes, and peripheral blood mononuclear cells (PBMC)28,29, we examined the expression of NLRP inflammasome signaling on PBMC, as described in previous reports30.
PBMC were immediately isolated from venous blood using the Ficoll-Paque PLUS (GE Healthcare Biosciences) density gradient centrifugation. Total RNA was obtained from PBMC by the guanidinium isothiocyanate method31. A 2.5 μg RNA aliquot was then reverse transcribed using 200U of Moloney murine leukemia virus reverse transcriptase (Fermentas, Thermo Fisher Scientific Inc.). The mRNA expression levels of NLRP3 inflammasome including NLRP3, caspase-1, and downstream cytokines including IL-1β and IL-18 were determined by TaqMan qPCR assay (Applied Biosystems). To standardize mRNA expression levels of NLRP3 inflammasome signaling, the mRNA levels of the housekeeping gene GAPDH were also determined in parallel for each sample. The expression level of NLRP3 inflammasome signaling was calculated using the comparative threshold cycle (Ct) method and evaluated by 2−ΔΔCt, where

Ex vivo assay to investigate the effect of IMQ on mRNA expressions of NLRP3 inflammasome signaling molecules
PBMC from 14 patients with AOSD and 4 healthy volunteers were resuspended in RPMI 1640 medium (Thermo Fisher Scientific Inc.) supplemented with 100 units/ml penicillin, 100 μg/ml streptomycin, and 10% fetal blood serum at a final concentration of 5 × 106 cells/well. We examined the change in the mRNA expressions of NLRP3 inflammasome and downstream cytokines in PBMC treated with IMQ (10 μg/ml and 25 μg/ml; InvivoGen), an effective ligand for TLR723 and an activator of NLRP3 inflammasome20. The mRNA expression levels of NLRP3, caspase-1, and downstream IL-1β and IL-18 were determined by TaqMan qPCR assay (Applied Biosystems). The fold change represents the ratio of mRNA levels comparing TLR7 ligand–treated PBMC with medium alone.
The supernatant levels of IL-1β and IL-18 were determined using ELISA kits (RayBiotech Inc. and Medical and Biology Laboratories Co. Ltd., respectively) according to each of the manufacturer’s instructions.
Ex vivo assay to investigate the effect of IMQ on protein expressions of NLRP3 inflammasome signaling
Total proteins were extracted from PBMC lysates treated with IMQ (25 μg/ml). The proteins were separated with 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and then transferred to PVDF membranes (BIO-RAD). Immunoblots were performed using primary antibodies against NLRP3 (Abcam), caspase-1 (Abcam), IL-1β (NOVUS), IL-18 (Santa Cruz Biotechnology), and GAPDH (Santa Cruz Biotechnology), and secondary horseradish peroxidase–conjugated anti-mouse or rabbit IgG antibodies (Santa Cruz Biotechnology). The reaction mixtures, requiring an equal amount of cell extracts from each set of experiments, were fractionated on 10%∼12% SDS-PAGE in running buffer (25 mM Tris, 192 mM glycine, 0.1% SDS). The membranes were blocked with 5% skimmed milk in Tris buffered saline with tween [150 mM NaCl, 20 mM Tris-HCl (pH 7.4), 0.1% Tween-20] at room temperature for 1 h, and subsequently probed with anti-NLRP3 (1:1000; Abcam), anti-caspase-1 (1:1000; Abcam), anti-IL-1β (1:1000; NOVUS), anti-IL-18 (1:1000; Santa Cruz Biotechnology), and anti-GAPDH (1:5000; Santa Cruz Biotechnology) at 4°C overnight. The membranes were incubated with peroxidase-conjugated secondary antibodies (1:10,000) at room temperature for 1 h. Immunoreactive bands were detected by radiographic film using the ECL detection system (Advansta), and band intensity was determined by Image J software. The protein levels of NLRP3 inflammasome and cytokines were normalized to GAPDH, and values are expressed relative to control.
Ex vivo assay to investigate the effect of NLRP3 inhibitor on protein expressions of NLRP3 inflammasome signaling
PBMC from patients with AOSD and controls were primed with IMQ (25 μg/ml; InvivoGen) for 2 h at 37°C in 5% CO2, and then treated with ATP (2 mM, Sigma-Aldrich) for 30 min. PBMC were subsequently treated with NLRP3 inhibitor glybenclamide (IMGENEX) for 4 h at 37°C in 5% CO2. The protein levels of NLRP3 inflammasome and downstream cytokines (IL-1β and IL-18) were determined by immunoblotting.
Statistical analysis
Prior to the start of our study, the total number of enrolled subjects sufficient for the statistical power of 80% and α error of 0.05 for NLRP3 expression investigation was calculated to be 45 (32 in the study group and 13 in the control group). Therefore, we enrolled 48 subjects including 34 patients with AOSD and 14 healthy volunteers. The results were presented as the mean ± SD or standard error of mean (SEM), or median (interquartile range). The nonparametric Mann-Whitney U test was used for between-group comparison of expression levels of NLRP3 inflammasome and downstream cytokines. The correlation coefficient was calculated using the nonparametric Spearman rank correlation test. The Wilcoxon signed-rank test was used to compare mRNA levels of NLRP3 inflammasome and downstream cytokines before and after treatment. A probability of < 0.05 was considered significant.
RESULTS
Clinical characteristics of patients with AOSD
Among the 34 patients with active AOSD, spiking fevers (≥ 39°C), evanescent rash, arthralgia or arthritis, sore throat, and lymphadenopathy were noted in 33 (97.1%), 30 (88.2%), 25 (73.5%), 24 (70.6%), and 13 patients (38.2%), respectively. At the 6-month followup, 14 (48.3%) of 29 patients who received sDMARD therapy alone and 4 (80%) of 5 TCZ-treated patients achieved complete remission defined as total absence of clinical symptoms. One TCZ-treated patient had a partial remission with arthritis as a main persisting symptom. There were no significant differences in the age at entry or in the proportion of women between the patients with AOSD and healthy controls.
The comparisons of mRNA expression levels of NLRP3 inflammasome signaling between patients with AOSD and healthy controls
As shown in Figure 1 and Table 1, significantly higher mRNA expression levels of inflammasome components including NLRP3, caspase-1, and downstream cytokines IL-1β and IL-18 were observed in patients with AOSD than in healthy subjects. Considering the variation of cytokine levels in different age groups, we further examined the differences in mRNA levels of NLRP3, caspase-1, IL-1β, and IL-18 between patients with AOSD and healthy subjects using propensity score–matching analysis for age. The results showed that their differences were still statistically significant (Table 1).

The comparisons of relative mRNA levels of NLRP3 inflammasome signaling, including (A) NLRP3, (B) caspase-1, (C) IL-1β, and (D) IL-18 between 34 patients with AOSD and 14 HC. Horizontal bar indicates median value. NLRP: NLR containing a pyrin domain; IL: interleukin; AOSD: adult-onset Still disease; HC: healthy controls.
The correlation between NLRP3 mRNA levels and AOSD disease activity
As illustrated in Figure 2A, NLRP3 mRNA levels were positively correlated with disease activity scores in patients with AOSD. Moreover, mRNA expression levels of NLRP3, caspase-1, IL-1β, and IL-18 significantly declined (mean ± SEM, 7.2 ± 4.7 vs 1.8 ± 1.0; 3.4 ± 0.9 vs 1.3 ± 0.3; 57.4 ± 43.8 vs 8.5 ± 5.5; and 14.3 ± 10.5 vs 2.8 ± 2.0; respectively, all p < 0.05) in patients with AOSD after 6-month therapy with sDMARD alone, paralleling clinical remission (Figure 2B). Similarly, mRNA expression levels of NLRP3, caspase-1, IL-1β, and IL-18 significantly declined (11.6 ± 3.3 vs 2.0 ± 0.9; 7.9 ± 3.0 vs 3.4 ± 2.0; 34.1 ± 19.4 vs 6.5 ± 3.6; and 47.3 ± 32.4 vs 13.0 ± 8.0; respectively, all p < 0.05) in patients with AOSD after 6-month TCZ therapy, paralleling clinical remission (Figure 2C).

The correlations between NLRP3 mRNA levels and disease activity of AOSD. (A) A positive correlation was present between NLRP3 mRNA expression levels and disease activity scores in patients with AOSD. Spearman rank test was used in the correlation analysis. The change in mRNA expression levels of (B) NLRP3 inflammasome components and (C) downstream cytokines was observed in patients with AOSD after 6-month therapy with sDMARD alone or IL-6Ri, respectively. Bars and error bars indicate mean and standard error of mean, respectively. *p < 0.005, versus before treatment, was determined by the Wilcoxon signed-rank test. NLRP: NLR containing a pyrin domain; AOSD: adult-onset Still disease; sDMARD: synthetic disease-modifying antirheumatic drugs; IL-6Ri: interleukin 6 receptor inhibitor.
The change in expressions of NLRP3 inflammasome and cytokines on PBMC treated with IMQ, a potential activator of NLRP3
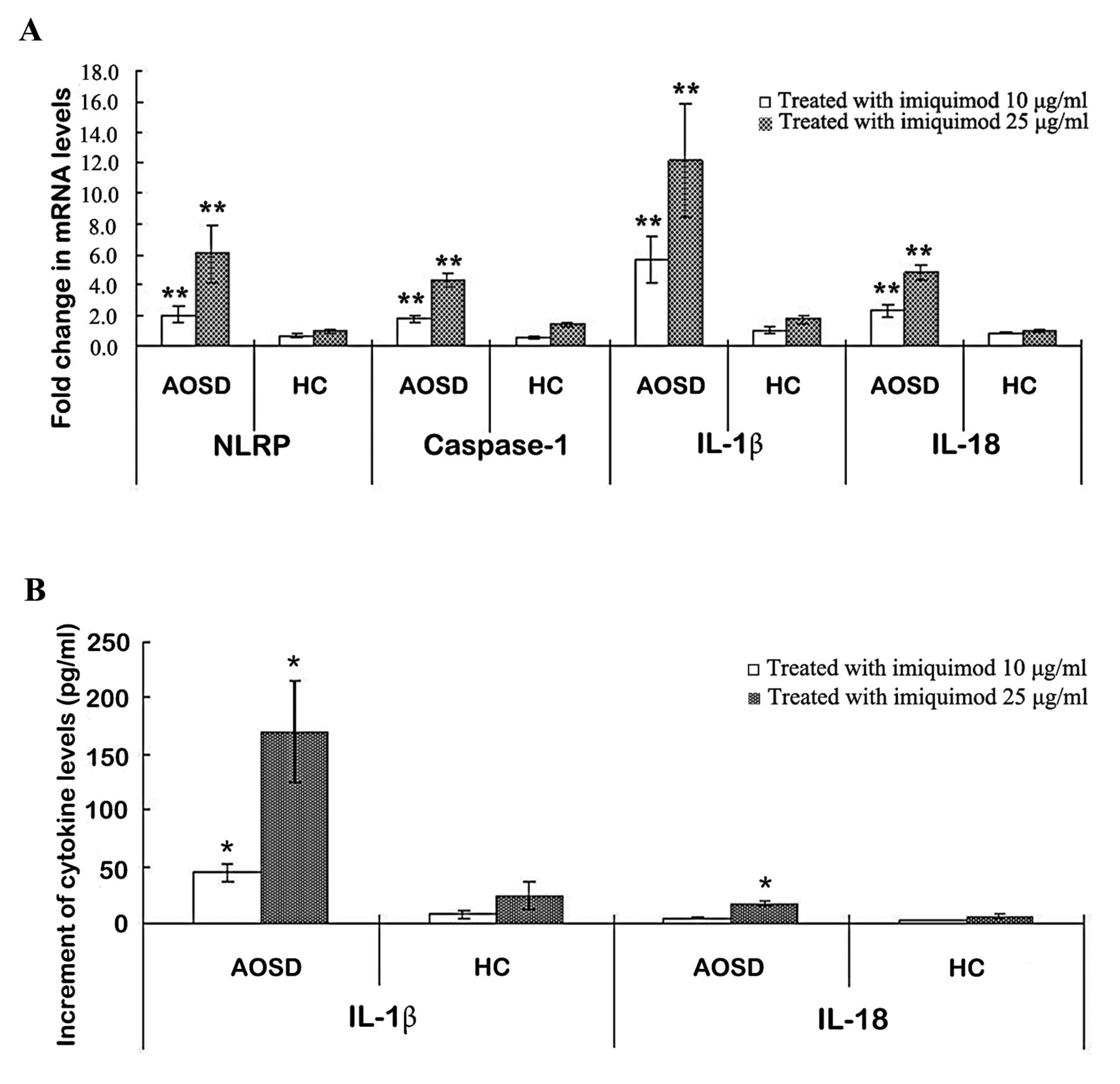
To study the effects of IMQ (a TLR7 ligand) on NLRP3 inflammasome activation, we examined the changes of mRNA levels of NLRP3 inflammasome signaling on PBMC after stimulation with this potential activator. IMQ (10 µg/ml and 25 µg/ml) stimulation of PBMC induced much greater and dose-dependent increases of mRNA levels of NLRP3 (2.06 ± 0.46 and 6.05 ± 1.84, respectively), caspase-1 (1.81 ± 0.23 and 4.25 ± 0.48, respectively), IL-1β (5.68 ± 1.51 and 12.13 ± 3.71, respectively), and IL-18 (2.32 ± 0.37 and 4.81 ± 0.51, respectively) in patients with AOSD compared with healthy controls (0.69 ± 0.12 and 0.97 ± 0.15 for NLRP3; 0.59 ± 0.09 and 1.43 ± 0.18 for caspase-1; 1.00 ± 0.23 and 1.76 ± 0.29 for IL-1β; 0.84 ± 0.04 and 1.05 ± 0.11 for IL-18; respectively, all p < 0.005; Figure 3A).
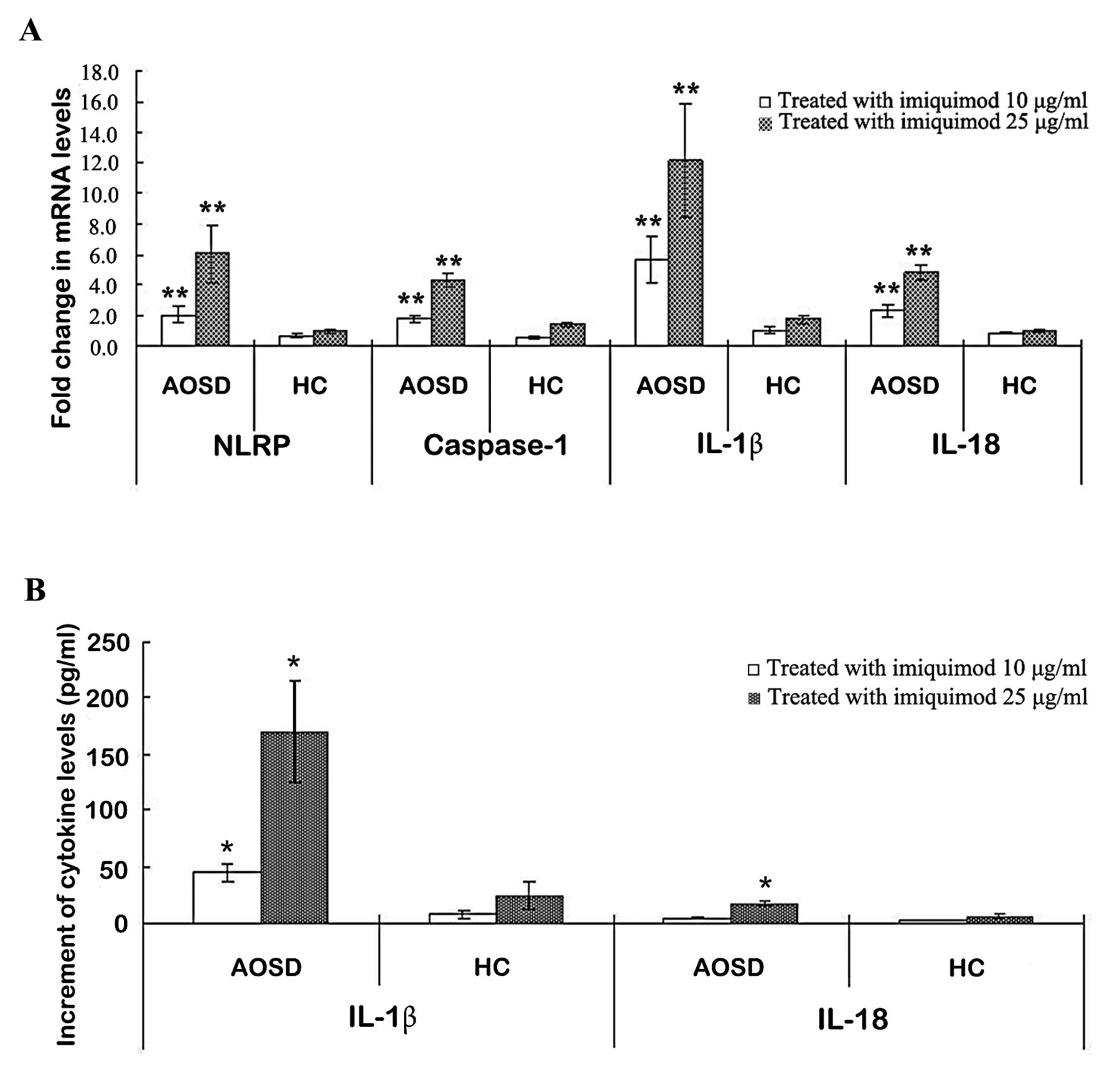
The change in expressions of NLRP3 inflammasome and cytokines on PBMC treated with IMQ, a potential activator of NLRP3. (A) The fold-changes in mRNA levels of NLRP3 inflammasome and downstream cytokines in PBMC stimulated with different concentrations of IMQ (10 µg/ml and 25 µg/ml) in both patients with AOSD and HC. (B) The change in the supernatant levels of downstream cytokines IL-1β and IL-18 on PBMC treated with different concentrations of imiquimod. Bars and error bars indicate mean and standard error of mean, respectively. * p < 0.05, ** p < 0.005, versus no treatment group. NLRP: NLR containing a pyrin domain; PBMC: peripheral blood mononuclear cells; IMQ: imiquimod; AOSD: adult-onset Still disease; HC: healthy controls; IL: interleukin.
As for the change in supernatant levels of downstream cytokines, IMQ (10 µg/ml and 25 µg/ml) stimulation of PBMC also resulted in greater and dose-dependent increment of IL-1β (44.75 ± 8.77 pg/ml and 170.22 ± 45.17 pg/ml, respectively) and IL-18 (4.15 ± 1.07 pg/ml and 17.24 ± 2.78 pg/ml, respectively) in patients with AOSD compared with controls (7.16 ± 3.05 pg/ml and 24.21 ± 12.09 pg/ml; 1.35 ± 0.28 pg/ml and 5.75 ± 2.38 pg/ml; respectively, all p < 0.05; Figure 3B).
In the immunoblotting examination, IMQ-stimulated PBMC from patients with AOSD induced more increases in protein expressions of NLRP3 (1.56 ± 0.22), caspase-1 (1.43 ± 0.12), IL-1β (2.38 ± 0.15), and IL-18 (1.49 ± 0.24) compared with healthy controls (0.91 ± 0.04, p < 0.005; 0.98 ± 0.04, p < 0.005; 1.36 ± 0.13, p < 0.005; and 1.03 ± 0.09, p < 0.05; respectively; Figures 4A–B).
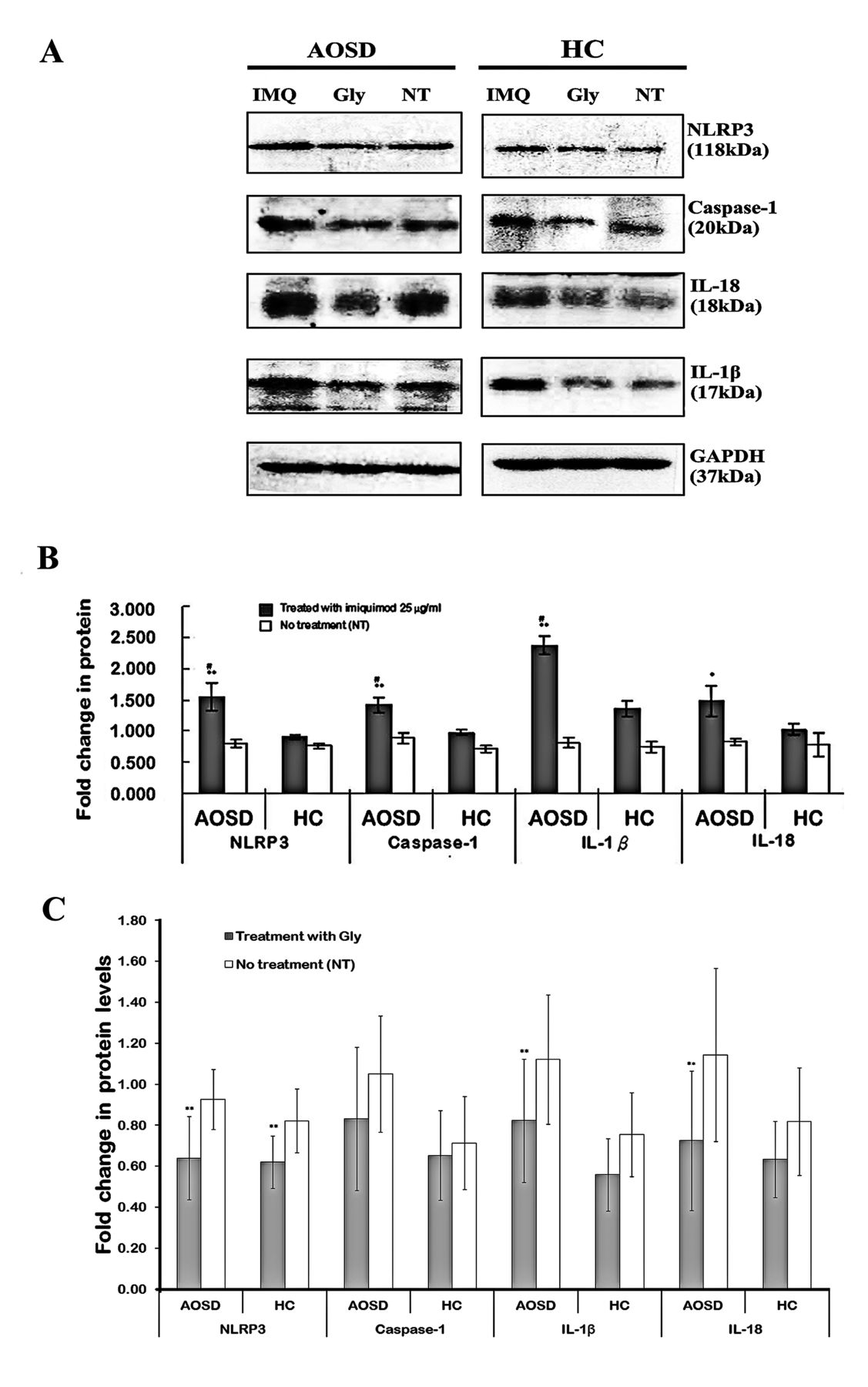
The change in protein levels of NLRP3 inflammasome and downstream cytokines in PBMC treated with NLRP3 activator or inhibitor. (A) A representative example of immunoblotting analyses in protein expression levels of NLRP3 inflammasome and cytokines in IMQ-treated or Gly-treated PBMC lysates obtained from 1 patient with AOSD and 1 HC. The fold changes in protein expressions of NLRP3, caspase-1, IL-1β, and IL-18 in PBMC stimulated with (B) IMQ (25 µg/ml) or (C) NLRP3 inhibitor (Gly) in both patients with AOSD and HC. Bars and error bars indicate mean and standard error of mean, respectively. * p < 0.05, ** p < 0.005, versus NT group; # p < 0.05, versus HC. NLRP: NLR containing a pyrin domain; PBMC: peripheral blood mononuclear cells; IMQ: imiquimod; Gly: glybenclamide; AOSD: adult-onset Still disease; HC: healthy controls; IL: interleukin; NT: no treatment.
The change in protein levels of NLRP3 inflammasome and downstream cytokines in PBMC treated with NLRP3 inhibitor
The inflammatory response to NLRP3 inhibitor was evaluated to analyze the involvement of NLRP3 in the activation of its inflammasome signaling in patients with AOSD. Our results showed that treatment with NLRP3 inhibitor (glybenclamide) could suppress the protein expression levels of NLRP3 (from 0.93 ± 0.15 to 0.64 ± 0.20, p = 0.004), caspase-1 (from 1.05 ± 0.28 to 0.83 ± 0.35, p = 0.065), IL-1β (from 1.12 ± 0.32 to 0.82 ± 0.30, p < 0.001), and IL-18 (from 1.14 ± 0.42 to 0.72 ± 0.34, p = 0.011) on PBMC from patients with AOSD (Figure 4C).
DISCUSSION
To our knowledge, ours is the first study to demonstrate significantly higher expression levels of NLRP3 inflammasome signaling including NLRP3, caspase-1, IL-1β, and IL-18 in patients with AOSD compared with healthy subjects. The mRNA expression levels of NLRP3 inflammasome components and downstream cytokines were also positively correlated with disease activity in patients with AOSD. Moreover, the decrease in expression levels of NLRP3 inflammasome signaling paralleled disease remission. These findings suggest the involvement of NLRP3 inflammasome in AOSD pathogenesis. In addition, IMQ stimulation of PBMC from patients with AOSD induced significantly greater and dose-dependent increases in the expression levels of NLRP3 inflammasome signaling compared with controls, indicating that IMQ may be a potential activator of NLRP3 inflammasome in AOSD.
Inflammasomes are cytoplasmic multiprotein complexes, and the dysregulation of NLRP3 inflammasome is involved in autoinflammatory diseases9,10. Although AOSD has been considered an autoinflammatory disease14,15, the pathogenic involvement of NLRP3 inflammasome in AOSD remains unclear. Our results showed that NLRP3 inflammasome components were significantly upregulated in AOSD, and NLRP3 expressions were positively correlated with AOSD activity scores. Moreover, the mRNA expressions of NLRP3 inflammasome components significantly declined after 6-month therapy, paralleling clinical remission. Our results were similar to the findings of 1 case report of AOSD with a marked activation of NLRP3 inflammasome32. These observations suggested an involvement of NLRP3 inflammasome in AOSD pathogenesis.
Although several NLRP3 activators such as crystalline particulates have been identified2,18, the precise ligands of NLRP3 in AOSD are currently unknown. Given the pathogenic involvement of TLR7 in AOSD19 and IMQ being a TLR7 ligand and activator of NLRP320,22,23, we then examined the involvement of IMQ in the activation of NLRP3 inflammasome signaling in patients with AOSD. Our ex vivo study revealed that IMQ could upregulate both the mRNA levels and protein levels of NLRP3 inflammasome signaling in a dose-dependent manner, a finding similar to previous studies that IMQ could activate NLRP3 inflammasome signaling20. Our findings were also supported by a previous study using mouse J774 cells in which NLRP3 inflammasome mRNA expressions were induced by the exposure of cells to TLR7 agonist33. Hence, IMQ (a TLR7 agonist) is a proven activator of NLRP3 inflammasome34, and its downstream cytokines are the mediators of the innate immune response in AOSD. Although the studied diseases were different, our data support previous findings that TLR7 antagonists could inhibit NLRP3 inflammasome activation in a murine model of psoriasis33 and a NZBW/F1 murine model of systemic lupus erythematosus21.
Considering that IMQ stimulation could upregulate the expressions of NLRP3 inflammasome components including caspase-1 and downstream cytokines in patients with AOSD, the NLRP3 activation should be functional. To further prove this novel finding, the inflammatory response to NLRP3 inhibitor was evaluated on PBMC. Our ex vivo study revealed that the protein expressions of NLRP3 and its downstream cytokines including IL-1β and IL-18 could be significantly suppressed by NLRP3 inhibitor, supporting the findings of a previous report35 and reinforcing the involvement of NLRP3 inflammasome–mediated inflammation in patients with AOSD.
There were, however, some limitations in our study. The lack of a significant change in the protein expression levels of NLRP3 inflammasome components such as caspase-1 on PBMC treated with NLRP3 inhibitor could be due to the small sample size. In addition, we did not elucidate the biological function of IMQ using a disease (AOSD)-based animal model because this is not currently available. Therefore, future studies dissecting the functional involvement and pathogenic mechanisms of IMQ in NLRP3 inflammasome signaling are certainly needed. In addition, further investigation into the association of other ligands of TLR7 with the activation of NLRP3 inflammasome in AOSD will provide more validation.
The upregulation of NLRP3 inflammasome signaling correlated with disease activity suggests its involvement in AOSD pathogenesis. With the precise activators of NLRP3 still unknown in AOSD, our results identified IMQ (a TLR7 agonist) as a potential activator of NLRP3 inflammasome, capable of enhancing IL-1β and IL-18 production. This elucidation of the potential role of NLRP3 inflammasome–mediated inflammation in AOSD will allow for future development of effective therapeutic agents targeting NLRP3 inflammasome36,37, or its downstream cytokines such as IL-1β16,17,37,38.
Acknowledgment
The authors thank the Biostatistics Task Force of Taichung Veterans General Hospital, Taichung, Taiwan.
Footnotes
Supported by a grant (NSC-101-2314-B-010-030-MY3) from the National Science Council, Taiwan.
- Accepted for publication April 7, 2017.








{kind=link}
{kind=link}
{kind=link}
{kind=link}