
Abstract
Agency has been shown to facilitate episodic memory. However, most paradigms use simple list-learning tasks, which preclude the ability to characterize more associative forms of memory, such as binding of items into spatial and temporal context. Across two studies, we characterize the role of agency on associative memory formation. We delineate agentive from passive memory encoding by allowing agency participants to play an online text-based game meant to simulate free exploration of objects in different rooms and yoking each passive participant’s trajectory to an agentive participant, thus equating their exposure to the stimuli. To assess memory differences, we gathered three measures: item descriptions, spatial location, and temporal order. While memory for spatial location and item features did not differ between groups, there was a marked enhancement of temporal order memory in the agentive group across two independent samples. These findings support a model of self-directed learning, in which agency facilitates the binding of items into a temporal context, which allows for the sequential binding of information into continuous narratives.
Similar content being viewed by others


Individuals are intrinsically motivated to explore novel environments, exerting agency to make decisions and observe the causal outcomes (Cho et al., 2016; Leotti & Delgado, 2011; Ly et al., 2019). These types of agentive decisions influence memory, such that when individuals are given the opportunity to actively manipulate their environment, they have better memory for the outcomes (Bridge et al., 2017; Chrastil & Warren, 2012; DuBrow et al., 2019; Hon & Yeo, 2021; Markant, 2020; Markant et al., 2014; Murty et al., 2015, 2018; Voss et al., 2011a, b). However, prior work has focused mostly on simple measures of item learning that precludes assays of associative memory, such as item-location binding and temporal order. The opportunity to actively manipulate one’s environment confers a sense of control over one’s environment (Ding et al., 2021; Leotti & Delgado, 2011). To test this hypothesis, we characterize the influence of agency on item-description memory, item–spatial context binding, and temporal order memory.
Agency has been conceptualized as a system that creates its own activities and experiences (di Paolo, 2005; Froese & di Paolo, 2011; Maturana & Varela, 1987; Varela et al., 1974, 1991; Weber & Varela, 2002). As systems that create their own activities and experiences, agentive systems acquire a sense of control over their environment and sensory experiences (Frith, 2014; Haggard, 2017). Moreover, a sense of agency can be studied by providing participants opportunities to make their own decisions, a manipulation that has revealed choice-induced performance enhancements for the outcomes of individual’s choices (Ding et al., 2021; Murty et al., 2015). Furthermore, many studies have implicated the mesolimbic dopamine system in choice-induced memory gains (Murty et al., 2015; Leotti & Delgado, 2011; Tanaka et al., 2008; see Murty & Adcock, 2017, for review), which in other contexts has been shown to facilitate associative memory for discrete sensory features of memoranda encountered in the context of reward.
The fact that choice the memory gains of agency being underpinned my neural mechanisms similar to reward motivation (i.e., mesolimbic engagement), suggests that memory mechanisms may be similar across the two learning contexts. In fact, recent research from our laboratory has shown that individual differences in agency-related memory enhancements are associated with value-related signals (DuBrow et al., 2019). We recently developed a theory suggesting that reward motivation facilitates a state of interrogative motivation in which not only the targets of goal pursuit are enhanced but also surrounding contextual features, suggesting that all forms of associative memory will be enhanced (Murty & Adcock, 2017). Here, we predict that this theory is not limited only to reward motivation but also exploration-based motivation as well, such that making choices improves memory for what was chosen. Therefore, Murty and Adcock’s (2017) theory of nonspecific associative memory enhancements from interrogative motivation Leotti and Delgado’s (2011) hypothesis that agency is a form of implicit reward, leads to the prediction that perceived control broadly facilitates associative memory for the learning environment.
Critically, hypotheses about interrogative motivational states broadly facilitating associative memory (i.e., across multiple contextual domains such as time and space) have mainly been grounded in engagement of underlying neural systems that support these processes (i.e., mesolimbic systems, hippocampus). However, few behavioral studies have studied multiple forms of associative memory in motivationally relevant contexts like agency. Thus, an alternative possibility is that memory enhancements may only extend to features of the environment directly related to a participant’s agency (i.e., contextual features they have control over). That is, rather than boosting memory for the learning environment more generally, exploration-based motivated behavior will facilitate associative memory for any information that occurs within proximity to motivationally relevant features directly related to the active exploration. In line with this interpretation, prior literature has demonstrated that agency enhances memory for the spatial location of items on a spatial grid during self-guided exploration (Voss et al., 2011a, b). However, prior work confounded manipulations of agency with meta-memory processes, such that participants could revisit items based on their confidence in their memory. Thus, the boundary conditions on which agency in isolation enhances associative learning remains an open question. Theories founded in meta-memory predicting that memory enhancements emerge from the ability to strategically revisit poorly learned or difficult information, whereas more exploration-based theories predicting more general enhancements in associative memory due to an engendered motivational state.
To study the influence of agency on associative memory for the binding of items to written descriptions, spatial location (i.e., binding items into their spatial context), and temporal sequence (i.e., binding items into their temporal context), the current study had participants complete an interactive text-based game in which participants could either agentively or passively explore multifeatural environments, and then memory was subsequently tested. Moreover, we explored potential interactions between agency and consolidation by manipulating the delay of test. Learning occurred during a free exploration task that included self-directed decisions to explore in the agency group. Participants were placed in multifeatural rooms and were instructed to actively explore different items in the room to find a hidden key to enter the next room. Upon exploring each item, participants were given a rich text-based description of the item, allowing us to dissociate memory for details of the item, the spatial location of the items, and the trajectory a participant made while searching for the key. An agentive participant controlled their trajectory through the encoding phase of the study, while a passive (i.e., yoked) participant was forced to follow the trajectory created by an agency group participant. To further isolate choice, passive participants were still able to click through the trials during the encoding phase (but they could not choose anything that deviated from the agency participant’s trajectory). This experimental approach enables isolation of causal mechanisms underlying agency effects on memory, while controlling for the exposure to different memoranda, study durations, and motor movements. Memory analyses focused on characterizing different forms of associative memory including item–spatial context, temporal order, and item description recall, as well as exploratory analyses characterizing free recall. Using this approach across two independent samples, we characterized associative memory benefits across groups to arbitrate between competing hypotheses of generalized associative memory enhancements or specific memory enhancements directed at binding of items to temporal context, as the order of search events was the feature of learning participants had agency over.
Study 1
Material
The stimuli used in Study 1 were all made by the authors and used the Inquisit 6 software (Millisecond Software, 2020). Specifically, we designed stimuli to represent random objects found in rooms of a fantasy building (a Wizard’s castle, Victorian mansion). Each object, when clicked on, yielded an object description that consisted of approximately two to three sensory descriptors (e.g., “The spellbook is bound with a cover made of tree bark”) and one action descriptor (e.g., “As you near it, its pages turn rapidly by themselves”). Object descriptions were normed prior to Study 1 by gathering ratings from online participants along two dimensions: valence (Was the description negative or positive?) and pleasantness (How pleasant was reading this description on a scale of 1–10?). We made sure the averaged ratings between houses did not differ significantly (data not shown here). Within rooms, objects and object descriptions were the same color to facilitate greater cohesion amongst within-room stimuli.
Method
Participants
One hundred and twenty-six participants were recruited online in eight separate batches for a 2-day study. The “agency” group data was collected first in each batch and later replayed to yoked partners in the “passive” group. If we were not able to successfully recruit a passive participant (because of failure to return on the second day) yoked to an agency participant after four attempts, we excluded the participant in order to expedite data collection, which resulted in the exclusion of 12 participants from the agency group, making the overall sample size n = 114. Informed consent was obtained from each participant, complying with Temple University’s Institutional Review Board. All participants were between the ages of 18 and 45, reported normal or corrected-to-normal vision, and spoke English as their native language. They were monetarily compensated at a rate of 7.25 per hour. Because the experiment was divided into two sessions, they were awarded an additional monetary bonus of 3 dollars for completing Day 2 of the study to promote study retention.
General procedure
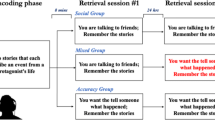
Our task was implemented online using the Prolific Online Testing Platform (https://www.prolific.co). Participants completed the task on desktop computers (we did not allow mobile devices), with stimuli making up an invariant percentage of given pixels. All the task scripts were made in Inquisit 6 (Millisecond Software, 2020). Participants first completed informed consent. Following consent, participants were first given instructions for the encoding task, then performed two runs of the encoding task (detailed below), separated by a 24-hour delay (Fig. 1). In this study, we tested both immediate and 24-hour memory as previous work shows enhanced item memory after a period of consolidation (Murty et al., 2018). The second encoding task was followed by a short distractor task consisting of 2.5 minutes of math problems, a memory test of item recall, cued recall of spatial locations of items, and temporal order memory.

Experimental design. a Schematic of overall experimental design for Study 1. On Day 1, participants completed an encoding task that consisted of two room simulations. Following a 24-hour delay; on Day 2, participants completed a second version of the same encoding task. After the second encoding task, participants completed a math distractor task, and then completed a series of retrieval tasks. b Encoding. During encoding, participants see a room label (e.g., “Room 1”) and item. When an item-word is clicked on, the screen is replaced with the item description. If the last click met the contingency requirements for that room (e.g.. “click on same item twice”), then the screen reveals a brief description about finding a key that takes the participant to the next room. If contingency requirements are not met, the screen is replaced with the previous room. c Retrieval. For retrieval, participants first completed a free recall task for items. Next was an item–spatial context associative memory test, in which participants were cued with the item words and they had to click on the grid tile in which the item word appeared during encoding. Next was a temporal order memory test, where participants were shown the item-words that appeared in a given room and they had to list them in the order in which they clicked on them during encoding. Finally, there was an item-description memory test, where participants were cued with an item and asked to recall as much of the item’s description as they can. d Sequence lengths for each contingency. Sequence lengths are the number of clicks that participants made for a given room. Here, we averaged the number of clicks across contingencies to show that sequence lengths were not considerably long. Sequence length did not influence any associative memory results
Memory encoding task
Next, we describe the encoding task. We first describe the task for the participants in the agency condition, and then detail how we modified this task for the participants in the passive condition. In the agency condition, participants were instructed to explore a series of rooms (2 rooms) to find a key to move to the next room. Each room consisted of 3 to 4 items. When they clicked on the item, they could read about the description of that item. To move from the current room to the next room, they must find a key, hidden amidst a certain item. The key is set to reveal itself when a preestablished contingency is met. These contingencies were not told to the participants beforehand, and thus they had to discover the sequence of items would unlock the key. There were three total contingencies, and thus each participant was exposed to one of the contingencies twice. The contingencies include “6 clicks,” “click the same item twice,” and “click all the items in the given room” (Fig. 1d). We counterbalanced the assignment of contingencies to rooms according to batches, so that sequence lengths were controlled for. There were no significant differences in trial durations or memory between contingencies (data not shown). We equipped rooms with contingencies to ensure that participants will not be able to find a pattern to reach the keys more quickly and thus engender more exploratory behavior. In each room, participants first viewed a screen labeled with the title of a room and a rectangle, in which text descriptions of the items were placed (Fig. 1b). The title tells the participant which “room” they are currently in and the rectangle was meant to simulate the room boundaries. Participants had the freedom to click on any item they want, in any order, with the ability to revisit an item. Once an item is clicked, the screen was replaced with the item’s description. If the item clicked on met the contingency requirements to move to the next room, then the participant was informed that they had found the key that will take them to the next room (Fig. 1b), and then the screen was replaced with the next room (i.e., a new room title and new items). However, if the contingency is not met, then the participant is taken back to the previous room, where they are once again free to click on any item. Both encoding tasks, on Day 1 and Day 2, consisted of two rooms (i.e., a total of four rooms), and so participants had to find two keys in each task to finish.
In the passive condition, participants played the same game as the agency group participants. Thus, the rooms, items, contingencies, and time spent on each screen from one agency participant was exactly the same for one passive participant. One of our main goals, however, was to assess the role that choice plays in driving interrogative goal states. Ergo, the only difference in the passive group game was that participants were forced to explore specific items (which were highlighted in a different color). These trajectories were determined by yoking them to the experimental manipulation of a participant in the agency condition. To illustrate: If an agency participant clicked on the spellbook in Trial 1 (where trials are operationalized as every time a word was clicked by the participant), then the spellbook in Trial 1 would be colored differently for the yoked partner in the passive group. Notably, we controlled for timing by making the minimum duration prior to clicking on items in the yoked group equal to the duration before item clicks in the agency group. Thus, exposure to memoranda, learning durations, and motor actions were controlled across groups.
Memory tests
Memory tests were administered in the same order for all participants (Fig. 1c). That is, first participants did a free recall of items followed by our three associative memory tests. First, they were presented with an item–spatial context test, followed by a temporal order test, and finally an item-description memory test. For all of the memory tests, except for item-descriptions, participants were first prompted to recall items from Day 2 and then prompted to recall items from Day 1. Participants were told prior to encoding that they would be tested on how much they learned about each room, in an attempt to facilitate exploratory behavior, but the specific types of associative memory tests were unknown to the participants as to not in traduce any biases. For this reason, spatial location-related associations were not necessarily irrelevant to participants.
Free recall
Following the second encoding task and distractor on Day 2, participants were instructed to freely recall as many items as possible, in any order. Following being prompted with the name of each room, participants typed responses into a text box at their own pace (i.e., self-paced). They were prompted with the most recent room names first (i.e., the rooms they explored on Day 2), followed by the room names of the rooms they explored on Day 1.
Item–spatial context associative memory
During learning, items could only appear in one of seven locations within the rectangle that was presented on the screen. The seven locations formed a grid-like tiling across the rectangle area. For the spatial location memory task, item words were presented, above a schematic of the room which indicated the potential locations. Participants were instructed to click the tile in which the items were presented.
Temporal order memory
For the temporal order memory task, participants were shown all the items that had appeared in a room and instructed to list them in the order in which they explored the items in a text box. Thus, items could be used more than once or not at all.
Item-description cued recall
For the item-description cued recall task, participants were cued with a specific item and instructed to recall as many descriptors about that item as possible. All items, across rooms from both days, were shown in randomized order.
Analysis
Free recall for items was scored by summing the total number of items that each subject recalled. Spatial location memory was scored by averaging the total number of correct responses.
Temporal order memory was scored using the Levenshtein distance metric (Levenshtein, 1966; Schulz & Mihov, 2003; Yujian & Bo, 2007). Levenshtein distance, also known as edit distance, measures the number of operations required to transform one string into another string. To determine chance performance using this measure, we permuted each recalled sequence 10,000 times and computed the Levenshtein distance between each permutation and the veridical sequence. Then, we computed the average and standard deviation of the permuted Levenshtein distances, used those values to transform the true recalled sequences into z-scores: (true distance − avg permuted distance) / std of permuted distance. We then compared the z-transformed scores against zero. Notably, greater Levenshtein distances scores indicate better memory, whereas lower values indicate worse memory.
Cued item-description recall was scored using an automated machine learning approach which compares the accuracy of free responses with the presented memoranda, which has recently been developed and validated in our laboratory (Shen et al., 2021). In brief, this approach converts the text from encoding as well as the subjects free recall response into a 512-demensional vector using the Universal Sentence Encoder (USE; Cer et al., 2018). Accuracy can then be determined by converting the cosine similarity of sentence embeddings between the memoranda and the free recall response into angular distances (Cer et al., 2018; Vaswani et al., 2017), such that high similarity in this multidimensional space reflects greater accuracy. Thus, we computed semantic similarities between cued item-description recalls and each sensory/action detail in the veridical description participants viewed during encoding, iteratively. To illustrate, if the spellbook has two sensory descriptors, a subject’s recall for the spellbook would be compared against both Descriptor 1 and Descriptor 2, one at a time. For each comparison, scores were binarized according to whether the semantic similarity computed with USE exceeded a threshold. The threshold was optimized by computing the intraclass correlation coefficient (Koo & Li, 2016) between a subset of hand-scored recalls and USE-scored recalls, binarized via a range of arbitrary thresholds. The optimal threshold computed here was ICC = 0.57.
All statistical analyses were run in R. Mixed-effects analysis of variance (ANOVA) models along with effect sizes were calculated using the Plot2WayANOVA function, and the t.test function for post hoc Student’s t tests. A comparison was deemed significant if the p value was less than .05. Additionally, we used the glmer function to run model comparisons.
Results
On average, the duration of encoding sessions for participants in the agency and passive conditions did not significantly differ from each other (agency group: M = 13.1 min, SD = 7.01 min; passive group: M = 14.1 min, SD = 8.91 min, p = .46), confirming our ability to match study duration across groups.
We next present the results from each of our associative memory tests, following the order in which they were presented (Fig. 2). For item–spatial location memory, both the passive group and agency group performed significantly greater than chance levels of performance, agency group: t(56) = 7.95, p < .001; passive group: t(56) = 4.9, p < .001. A mixed-effects ANOVA revealed a main effect of delay, F(1, 224) = 9.93, p = .002, η2 = .038, such that participants had worse memory after a delay (immediate recall: M = .364, SD = 0.218; delayed recall: M = .266, SD = .246), but no effect of agency group, F(1, 224) = 0.001, p = .97, η2 = 0, or Group × Time interaction, F(1, 224) = 0.097, p = .76, η2 = 0. These findings suggest that individuals in both groups had intact associative memory for items and their spatial locations, which decayed over time. However, agency did not have any influence on item–spatial context binding.

Study 1 results. a Item–spatial context associative memory for Study 1 shows an effect of delay but no significant differences between groups. b Temporal order memory shows significant effects of delay and between groups but no group by time interaction, as measured via z-transformed Levenshtein distances between veridical and recalled temporal sequences. Notably greater scores indicate better memory, whereas lower scores indicate worse memory. The agency group scored significantly better on temporal order memory than the passive group. c Item description memory for Study 1 shows an effect of delay but no significant differences between groups. a–c Black dots represent the mean. Error bars represent the standard error of the mean. Asterisks represent p < .05
For temporal order memory, both groups scored above chance, agency group: t(56) = 8.132, p < .001; passive group: t(56) = 4.981, p = .001. A mixed-effects ANOVA revealed a main effect of delay, F(1, 452) = 36.95, p < .001, η2 = .073, such that participants had lower memory after a delay (immediate recall: M = .827, SD = .912; delayed recall: M = .225, SD = .677) and a main effect of group, F(1, 452) = 14.01, p = .001, η2 = .028, with agentive participants recalling sequences with greater accuracy, agency group: M = .71, SD = .934; passive group: M = .34, SD = .728. There was no Group × Time interaction, F(1, 452) = 1.21, p = .267, η2 = .002. Altogether, these findings entail that participants’ memory for their sequences of item clicks was significantly better than chance and that the agency group performed reliably better than the passive group across both delays.
For cued recall of item descriptions, there was a significant effect of delay, t(113) = −5.67 p < .001; F(1, 224) = 26.14, η2 = .111, such that memory was worse when tested at a 24-hour delay versus immediately. There was no main effect of group, F(1, 224) = 1.3, p = .26, η2 = .006, or a Group × Delay interaction, F(1, 224) = 0.003; p = .96, η2 = 0.
Finally, we present the exploratory analyses on free recall of encountered items. Item recall percentages, collapsed across both days, were very low for both groups, agency group: M = .12, SD = .14; passive group: M = .16, SD = .16. There were main effects of group, F(1, 224) = 4.68, p = .032, η2 = .02, and time, F(1, 224) = 76.1, p < .001, η2 = .254, but no Group × Time interaction, F(1, 224) = 3.09, p = .08, η2 = .014. Thus, participants had lower item recall in the agency condition as well as worse item recall after a 24-hour delay. The trending interaction suggests that these differences were more prominent after the 24-hours delay. Notably, these findings contrast with prior work that has found agency-induced item memory gains; however, the effect size for this effect is quite small. The nature of our free recall task (only prompting participants with the room title), however, may have increased the difficulty. That is, it may be the case that agency participants were focused on recalling previously visited items which were goal relevant; while passive participants, who do not have this task-related burden, were able to broaden their attention to other items and the room names.
Discussion: Study 1
Study 1 showed that participants were successful in encoding multiple features from our text-based game, such as descriptions of encountered items, item–spatial context associations, and the temporal sequence of items visited, validating this novel paradigm. Critically, our results showed that agency uniquely enhanced temporal order memory, as opposed to item-description memory and item–spatial context binding. Thus, from the current results we can surmise that memory for the temporal order of experienced events is enhanced under conditions that promote agentive-manipulation of temporal sequences. This supports the idea that reward systems in the brain are targeting information associated with control because the temporal order of the items was all that participants in the agency group could manipulate. We further found surprising results that free recall memory was greater in the control versus agency condition, which differs from our theoretical predictions of greater memory in associative memory. However, prior work has never investigated the influence of agency on free recall, which should be addressed in future research.
Study 2
Motivation
There was a potential confound in Study 1, in that memory tests were always presented in the same order (i.e., spatial location test, followed by temporal order test, followed by item description test). In Study 2, we aimed to replicate our findings and also remove the confound of order effects by intermixing trials of our associative memory tests. Further, given the lack of compelling differences in agency effects across immediate and delayed recall, we limited our design to only testing immediate recall.
Material
All materials used in Study 2 were the same as those used in Study 1.
Method
One hundred and sixty-six participants were recruited online in eight separate batches. Like the first study, agency group data were collected first and later replayed to yoked partners in the passive group. After four attempts of passive participants to complete the game, we excluded that participant, resulting in six excluded participants, making the overall sample size of Study 2 n = 160.
The recruitment constraints, materials, and encoding task of Study 2 were the same as that of Study 1 with the following exceptions. First, we only tested immediate recall because Study 1 results showed no Group × Time interaction in any associative memory tests. Second, we randomized the order in which the item–spatial context binding and temporal order memory tests were presented. Third, given that memory tests were intermixed on a trial-by-trial basis and that temporal order, spatial location, and item-description memory were all cued, we eliminated free recall of items as it may have been influenced by the presentation of the items in the other tests. That is, the intermixing of memory tests means cued recalls can precede free recall which would clearly bias free recall results.
Results
The duration of encoding sessions between groups were statistically similar (agency group: M = 13.86 min, SD = 7.54 min; passive group: M = 14.44 min, SD = 7.57 min; p = .611), again confirming our ability to match study duration.
For spatial location memory, both agency and passive participants performed above chance levels, agency: t(79) = −479.18, p < .001; passive: t(79) = −536.06, p < .001. A t test revealed no significant differences between groups (agency group: M = .35, SD = .261, passive group: M = .32, SD = .234; p = .35) (Fig. 3). These results replicated what we found in the first study—mainly, that associative memory was intact in both groups and that agency did not influence item-description cued recall or item–spatial context binding.

Memory test results from Study 2. a The spatial accuracy of item-spatial context associative memory does not differ significantly between groups. b Accuracy for recalled temporal sequences differs significantly between groups, as measured by z-transformed Levenshtein distances. The agency group demonstrated significantly better temporal order memory. c Item description memory does not differ significantly in the number of item descriptors recalled between groups. a–c Black dots represent the mean. Error bars represent the standard error of the mean. Asterisk represents p < .05
For temporal order memory, both groups again scored above chance when comparing z-transformed Levenshtein distances against zero, agency group: t(79) = 11.485, p < .001, passive group: t(79) = 7.798, p < .001. Further, we replicated the first study’s finding of significantly better temporal order memory in the agency group (agency group: M = .94, SD = .738; passive group: M = .6, SD = .697; p = .002) (Fig. 3). Altogether, the findings from the second study reaffirm a strong agency group effect on temporal order memory.
For item-description cued recall, there was no effect of group on the number of item descriptors recalled (agency group: M = .13, SD = 5.14; passive group: M = .13, SD = 5.05; p = .76) (Fig. 3).
Discussion: Study 2
The results from Study 2 replicated the effects observed from Study 1, while controlling for order effects. That is, both groups had statistically similar spatial location and item-description memory, but participants in the agency group had significantly better temporal order memory than participants in the passive group.
General discussion
In the present experiment, we characterized the effect of agency on different forms of associative memory by having participants explore a multifeatural text-based environment. We conducted two studies, both of which suggest that agency enhances temporal order memory, without significantly influencing item-description or item–spatial context memory. The results from this experiment provide support for a motivational system, perhaps akin to reward motivation, that targets information under one’s control to be encoded in memory, as opposed to more generalized enhancements in all forms of associative memory. Notably, our paradigm allowed us to determine that these effects were driven by active decision-making during agentive choice, as we controlled for general motor responses, information content, and encoding time.
The enhancement of temporal order memory in agentive participants relative to passive participants dovetails with theories of motivated encoding and extend them into the domain of intrinsic motivation centered on exploration. While we and others have emphasized a more general enhancement of associative memory due to intrinsically motivated exploratory behavior, the current study suggests that such benefits are particular to associations with temporal context. Indeed, in the current study, we also tested item–spatial context associative memory, which was sensitive to forgetting but did not differ across agency conditions. Interestingly, this result differs from that of previous studies investigating active learning of item-spatial context associations. For example, Voss et al. (2011a) had participants explore a screen via a sliding window, where they could find objects one at a time. In this paradigm, the agency group demonstrated better memory for the spatial location of objects in a later memory test.
One possible reason why the current study failed to replicate this finding is because, in Voss et al. (2011a), participants discovered objects by actively sliding the window. The sliding of the window thus adds an agency component to spatial memory which was absent in our study. In the current study, participants are shown all the items on a screen at once (i.e., they are passively exposed to spatial locations, whether they are in the agency or passive group). Additionally, the two studies may have induced different motivational contexts as participants in Voss et al. (2011a) were explicitly instructed to memorize the object and object location, whereas, in the current study, participants were told to find the “key.” This interpretation is consistent with theories suggesting that active learning benefits are task-dependent (Chrastil & Warren, 2012). Altogether, these findings open the possibility that an individual’s goal state in the moment can interact with motivational state to broaden the scope of agency on associative memory. This interpretation is further supported by the findings from the prior work, in which goal-driven metamemory processes allowed individuals to revisit spatial locations to support item-spatial context binding (Voss et al., 2011a, b). However, in our task, the salient feature of encoding may have been temporal sequences as individuals were driven to determine the processes that lead to findings keys.
Thus, our findings conflict with our originally conceived hypotheses about more generalized associative memory enhancements and suggests a modification of a prior theory developed in our lab that characterizes interrogative motivation. Specifically, within this model, motivational states that promote active exploration, such as agency, are predicted to enhance all forms of associative memory such that relationships between individual items and their surrounding contexts are maintained (Murty & Adcock, 2017). However, our results show that these benefits to item–context binding may be specified by the goal state of individuals during agency, such that goal-oriented exploration can target different types of associative memory depending on the motivational context. Thus, interrogative motivation consists of testing novel action–outcome relationships with respect to the current motivational context, which highlights what features of an environment are controllable and relevant. Under our revised theory of interrogative motivation, we propose that motivational context elucidates relevant features of the environment that an agent can act on to achieve a goal.
In line with this interpretation, our current findings show that there are no enhancements in memory for the description of items. At first glance, this seems in contrast to past work showing that perceived agency can influence item memory when subjects were able to make arbitrary choices about which information to learn (Coverdale & Nairne, 2019; DuBrow et al., 2019; Hon & Yeo, 2021; Markant et al., 2014; Murty et al., 2015). However, in this prior work, the focus of exploration was the outcome of individual’s decisions. One important difference between the current study is that the content of individual items was not especially relevant to participant’s interrogative goals to find keys. Thus, our negative findings on item-description associative memory further corroborate our interpretation that interrogative motivational states may be bounded by the relevance of the memoranda to the individual’s exploration related goals. An interesting future study that could further dissociate goal-related memory in agency versus passive participants would be to include additional agency conditions, in which spatial information rather than finding a key was prioritized in instructions (i.e., a hidden door is placed underneath an item). This way the two agency conditions are identical in all respects besides their goals. This could be further bolstered by randomizing the spatial presentation of items during search to further highlight that spatial information is stabilized, with the prediction that agency would enhance spatial memory in this learning context.
Thus, the current findings specify our prior models of motivated memory, at least in the context of agency, such that only features of the environment under agentic control receive associative memory benefits. Moreover, these patterns of findings may relate to another well-known theory of agency: intentional binding theory (Haggard, 2017; Moore & Obhi, 2012), which focuses on more perception-related cognition. Within this theoretical framework, intentional binding effects are intimately related to agency effects and a sense of control. Specifically, intentional binding theory (Frith, 2014; Haggard, 2017), says that sensory matches between intentions and action effects increase, while sensory mismatches (i.e., prediction errors) decrease, sense of agency (Blakemore & Decety, 2001; Blakemore et al., 2000, 2002; Frith et al., 2000). Ergo, the comparator model says that ascribing ownership to action-induced sensory effects generates bridges between items and outcomes when the sensorimotor feedback matches the efference copies of the intended action signal (Wen et al., 2019), which can be construed as a mechanism of control (Beck et al., 2017; Frith, 2014; Haggard, 2017, 2019). Within this framework, actions and outcomes become bound together thus providing an architecture that binds items and outcomes with each other, functionally bridging these features of learning over time by embedding them in temporal context. An alternative explanation of our results is that mechanisms associated with intentional binding lead to a chaining of events connected by action, thus allowing individuals to resolve the temporal order of their search history. Future work will need to get more explicit characterizations of intentional binding during agency guided memory encoding to see how well this theoretical framework intersects or dissects from our motivated memory framework.
Together, our findings shed light into the nature of memory representations that are enhanced under states of agentic learning, in that temporal sequence memory is enhanced as individuals actively explore a space to find a goal. These findings not only inform agency’s role in memory, but also broader concepts of curiosity drive (Loewenstein, 1994), free exploration (Gruber & Ranganath, 2019), and information-seeking (Gottlieb et al., 2013). Across these frameworks, interrogative motivational states enhance memory for goal-relevant features of learning providing rich maps of our environment that are tailored to suit our goals in the moment.
Data availability
The data and materials for all experiments are available by contacting the last author (V.P.M.), and none of the experiments was preregistered .
References
Beck, B., di Costa, S., & Haggard, P. (2017). Having control over the external world increases the implicit sense of agency. Cognition, 162. https://doi.org/10.1016/j.cognition.2017.02.002
Blakemore, S. J., & Decety, J. (2001). From the perception of action to the understanding of intention. Nature Reviews Neuroscience, 2(8). https://doi.org/10.1038/35086023
Blakemore, S. J., Smith, J., Steel, R., Johnstone, E. C., & Frith, C. D. (2000). The perception of self-produced sensory stimuli in patients with auditory hallucinations and passivity experiences: Evidence for a breakdown in self-monitoring. Psychological Medicine, 30(5). https://doi.org/10.1017/S0033291799002676
Blakemore, S. J., Wolpert, D. M., & Frith, C. D. (2002). Abnormalities in the awareness of action. Trends in Cognitive Sciences, 6(6). https://doi.org/10.1016/S1364-6613(02)01907-1
Bridge, D. J., Cohen, N. J., & Voss, J. L. (2017). Distinct hippocampal versus frontoparietal network contributions to retrieval and memory-guided exploration. Journal of Cognitive Neuroscience, 29(8). https://doi.org/10.1162/jocn_a_01143
Cer, D., Yang, Y., Kong, S. Y., Hua, N., Limtiaco, N., John, R. S., Constant, N., Guajardo-Céspedes, M., Yuan, S., Tar, C., Sung, Y. H., Strope, B., & Kurzweil, R. (2018). Universal sentence encoder. Proceedings of the 2018 Conference on Empirical Methods in Natural Language Processing: System Demonstrations. https://doi.org/10.18653/v1/D18-2029
Cho, C., Smith, D. V., & Delgado, M. R. (2016). Reward sensitivity enhances ventrolateral prefrontal cortex activation during free choice. Frontiers in Neuroscience, 10(NOV). https://doi.org/10.3389/fnins.2016.00529
Chrastil, E. R., & Warren, W. H. (2012). Active and passive contributions to spatial learning. Psychonomic Bulletin and Review, 19(1). https://doi.org/10.3758/s13423-011-0182-x
Coverdale, M. E., & Nairne, J. S. (2019). The mnemonic effect of choice. Psychonomic Bulletin & Review, 26(4). https://doi.org/10.3758/s13423-019-01575-z
di Paolo, E. A. (2005). Autopoiesis, adaptivity, teleology, agency. Phenomenology and the Cognitive Sciences, 4(4). https://doi.org/10.1007/s11097-005-9002-y
Ding, Z., Jiang, T., Chen, C., Murty, V. P., Xue, J., & Zhang, M. (2021). The effect of choice on intentional and incidental memory. Learning & Memory (Cold Spring Harbor, N.Y.), 28(12). https://doi.org/10.1101/lm.053433.121
DuBrow, S., Eberts, E. A., & Murty, V. P. (2019). A common mechanism underlying choice’s influence on preference and memory. Psychonomic Bulletin & Review, 26(6). https://doi.org/10.3758/s13423-019-01650-5
Frith, C. D. (2014). Action, agency and responsibility. Neuropsychologia, 55(1). https://doi.org/10.1016/j.neuropsychologia.2013.09.007
Frith, C. D., Blakemore, S. J., & Wolpert, D. M. (2000). Abnormalities in the awareness and control of action. Philosophical Transactions of the Royal Society B: Biological Sciences, 355(1404). https://doi.org/10.1098/rstb.2000.0734
Froese, T., & di Paolo, E. A. (2011). The enactive approach Theoretical sketches from cell to society. Pragmatics & Cognition, 19(1). https://doi.org/10.1075/pc.19.1.01fro
Gottlieb, J., Oudeyer, P. Y., Lopes, M., & Baranes, A. (2013). Information-seeking, curiosity, and attention: Computational and neural mechanisms. Trends in Cognitive Sciences, 17(11). https://doi.org/10.1016/j.tics.2013.09.001
Gruber, M. J., & Ranganath, C. (2019). How curiosity enhances hippocampus-dependent memory: The Prediction, Appraisal, Curiosity, and Exploration (PACE) framework. Trends in Cognitive Sciences, 23(12). https://doi.org/10.1016/j.tics.2019.10.003
Haggard, P. (2017). Sense of agency in the human brain. Nature Reviews Neuroscience, 18(4). https://doi.org/10.1038/nrn.2017.14
Haggard, P. (2019). The neurocognitive bases of human volition. Annual Review of Psychology, 70. https://doi.org/10.1146/annurev-psych-010418-103348
Hon, N., & Yeo, N. (2021). Having a sense of agency can improve memory. Psychonomic Bulletin and Review, 28(3):946–952. https://doi.org/10.3758/s13423-020-01849-x
Koo, T. K., & Li, M. Y. (2016). A guideline of selecting and reporting intraclass correlation coefficients for reliability research. Journal of Chiropractic Medicine, 15(2). https://doi.org/10.1016/j.jcm.2016.02.012
Leotti, L. A., & Delgado, M. R. (2011). The inherent reward of choice. Psychological Science, 22(10):1310–1318. https://doi.org/10.1177/0956797611417005
Levenshtein, V. (1966). Binary codes capable of correcting deletions, insertions, and reversals. Soviet Physics Doklady, 10(8).
Loewenstein, G. (1994). The psychology of curiosity: A review and reinterpretation. Psychological Bulletin, 116(1). https://doi.org/10.1037//0033-2909.116.1.75
Ly, V., Wang, K. S., Bhanji, J., & Delgado, M. R. (2019). A reward-based framework of perceived control. Frontiers in Neuroscience, 13(FEB). https://doi.org/10.3389/fnins.2019.00065
Markant, D. B. (2020). Active transitive inference: When learner control facilitates integrative encoding. Cognition, 200. https://doi.org/10.1016/j.cognition.2020.104188
Markant, D., DuBrow, S., Davachi, L., & Gureckis, T. M. (2014). Deconstructing the effect of self-directed study on episodic memory. Memory & Cognition, 42(8). https://doi.org/10.3758/s13421-014-0435-9
Maturana, H. R., & Varela, F. J. (1987). The tree of knowledge: The biological roots of human understanding. New Science Library/Shambhala Publications.
Moore, J. W., & Obhi, S. S. (2012). Intentional binding and the sense of agency: A review. Consciousness and Cognition, 21(1). https://doi.org/10.1016/j.concog.2011.12.002
Murty, V. P., & Adcock, R. A. (2017). Distinct medial temporal lobe network states as neural contexts for motivated memory formation. In D. Hannula & M. Duff (Eds.), The Hippocampus from cells to systems: Structure, connectivity, and functional contributions to memory and flexible cognition (pp. 467–501). https://doi.org/10.1007/978-3-319-50406-3_15
Murty, V. P., DuBrow, S., & Davachi, L. (2015). The simple act of choosing influences declarative memory. Journal of Neuroscience, 35(16). https://doi.org/10.1523/JNEUROSCI.4181-14.2015
Murty, V. P., Dubrow, S., & Davachi, L. (2018). Decision-making increases episodic memory via postencoding consolidation. Journal of Cognitive Neuroscience, 31(9). https://doi.org/10.1162/jocn_a_01321
Schulz, K. U., & Mihov, S. (2003). Fast string correction with Levenshtein automata. International Journal on Document Analysis and Recognition, 5(1).
Shen, X., Houser, T., Smith, D. V., & Murty, V. P. (2021). Machine-learning as a validated tool to characterize individual differences in free recall of naturalistic events. https://doi.org/10.31234/osf.io/uygzv
Tanaka, S. C., Balleine, B. W., & O’Doherty, J. P. (2008). Calculating consequences: Brain systems that encode the causal effects of actions. Journal of Neuroscience, 28(26). https://doi.org/10.1523/JNEUROSCI.1808-08.2008
Varela, F. J. (1991). Organism: A meshwork of selfless selves. In organism and the origins of self (pp. 79–107). Springer, Dordrecht.
Varela, F. G., Maturana, H. R., & Uribe, R. (1974). Autopoiesis: The organization of living systems, its characterization and a model. BioSystems, 5(4). https://doi.org/10.1016/0303-2647(74)90031-8
Vaswani, A., Shazeer, N., Parmar, N., Uszkoreit, J., Jones, L., Gomez, A. N., Kaiser, Ł., & Polosukhin, I. (2017). Attention is all you need. Advances in Neural Information Processing Systems, 2017(December).
Voss, J. L., Gonsalves, B. D., Federmeier, K. D., Tranel, D., & Cohen, N. J. (2011a). Hippocampal brain network coordination during volitional exploratory behavior enhances learning. Nature Neuroscience, 14(1). https://doi.org/10.1038/nn.2693
Voss, J. L., Warren, D. E., Gonsalvesa, B. D., Federmeier, K. D., Tranel, D., & Cohen, N. J. (2011b). Spontaneous revisitation during visual exploration as a link among strategic behavior, learning, and the hippocampus. Proceedings of the National Academy of Sciences of the United States of America, 108(31). https://doi.org/10.1073/pnas.1100225108
Weber, A., & Varela, F. (2002). Life after Kant: Natural purposes and the autopoietic foundations of biological individuality. Phenomenology and the Cognitive Sciences, 1(2). https://doi.org/10.1023/A:1020368120174
Wen, W., Kuroki, Y., & Asama, H. (2019). The sense of agency in driving automation. Frontiers in Psychology, 10. https://doi.org/10.3389/fpsyg.2019.02691
Yujian, L., & Bo, L. (2007). A normalized Levenshtein distance metric. IEEE Transactions on Pattern Analysis and Machine Intelligence, 29(6). https://doi.org/10.1109/TPAMI.2007.1078
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Houser, T.M., Tompary, A. & Murty, V.P. Agency enhances temporal order memory in an interactive exploration game. Psychon Bull Rev 29, 2219–2228 (2022). https://doi.org/10.3758/s13423-022-02152-7
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13423-022-02152-7