
Abstract
We report two experiments designed to investigate how the implied motion of tactile stimuli influences perceived location. Predicting the location of sensory input is especially important as far as the perception of, and interaction with, the external world is concerned. Using two different experimental approaches, an overall pattern of localization shifts analogous to what has been described previously in the visual and auditory modalities is reported. That is, participants perceive the last location of a dynamic stimulus further along its trajectory than is objectively the case. In Experiment 1, participants judged whether the last vibration in a sequence of three was located closer to the wrist or to the elbow. In Experiment 2, they indicated the last location on a ruler attached to their forearm. We further pinpoint the effects of implied motion on tactile localization by investigating the independent influences of motion direction and perceptual uncertainty. Taken together, these findings underline the importance of dynamic information in localizing tactile stimuli on the skin.
Similar content being viewed by others


Introduction
Children sometimes play a game in which one child extends his or her arm, palm upwards, and closes his/her eyes. The other child then runs his/her finger slowly along the skin surface from wrist to elbow. The first child has to indicate when the finger of the other has reached the elbow joint. Typically, the first child will indicate the arrival of the other child’s finger at their elbow too early (that is, before the finger had actually reached the elbow joint). In other words, the finger is perceived as occupying a location further along in its trajectory than is actually the case. While this example is nothing more than a children’s game, knowing where people localize dynamic tactile stimuli turns out to be of both theoretical and practical importance. Yet, until now, only a pure replication (Brugger & Meier, 2015) and a first experiment looking at such endpoint-localization of dynamic tactile stimuli (Macauda, Lenggenhager, Meier, Essick, & Brugger, 2018) have been published. The present study was designed to systematically investigate the influence of implied motion on tactile localization.
Event localization is an important task for our senses and, in particular, for multisensory integration. For instance, if we feel an insect on our skin, we need to know where it is in order to be able to engage with or act on it. Interestingly, even the localization of static, tactile events (e.g., a tap on the skin) is not always error-free. It has, for example, often been reported that a static tactile stimulus may be systematically misperceived at a different spatial location (Harrar & Harris, 2009; Ho & Spence, 2007; Longo, 2017; Mancini, Longo, Iannetti, & Haggard, 2011; Margolis & Longo, 2015; Steenbergen, Buitenweg, Trojan, & Veltink, 2014; Trojan et al., 2006; see Medina & Coslett, 2016, for a discussion). Even on the hand and fingers, locations that are very sensitive to two-point discrimination and localization (see Gallace & Spence, 2014; Stevens & Choo, 1996; Weinstein, 1968), systematic mislocalizations have been documented (e.g., Mancini et al., 2011; Margolis & Longo, 2015). Tactile localization can also be biased due to the changing orientation of the head (Ho & Spence, 2007) as well as the position of the eyes (Harrar & Harris, 2009). Moreover, the perceived location of a tactile stimulus can be shifted toward the location of a preceding or succeeding tactile stimulus (this is known as tactile saltation; Geldard & Sherrick, 1972). When two tactile stimuli are presented simultaneously or else in rapid succession, only one stimulus somewhere between the two stimuli may be perceived (this is known as funneling; Chen, Friedman, & Roe, 2003; Gardner & Spencer, 1972). Those examples show that the precision in tactile localization is variable and can be poor.
In the visual and auditory modalities, it is well known that the localization of stimuli is influenced by their (implied) motion. Freyd and Finke (1984) were the first to report that the location of a stimulus is displaced in the direction of (implied) motion if the stimulus seems to rotate in a single consistent direction (Experiment 1) as compared to a condition in which the stimulus does not (Experiment 2). This forward shift, when the last location of a moving stimulus is perceived further along its trajectory, is a robust and oft-replicated finding in the literature on representational momentum (see Hubbard, 2005, 2014, 2018, for reviews). Many theories have tried to explain the forward shift, ranging from internalization accounts (e.g., Freyd, 1987; Freyd & Finke, 1984) to network-models (e.g., the bow-wave model; Müsseler, Stork, & Kerzel, 2002), as classified by Hubbard (2010) in his overview of theories of representational momentum. On a neuronal level, the forward shift might also be explained with the help of the theory of dynamic predictive coding (e.g., see Clark, 2013; Huang & Rao, 2011; Rao & Ballard, 1999). Following on from this idea, we constantly predict upcoming sensory inputs. Hence, if a stimulus moves, it would be reasonable for the sensory system to predict the next sensory input in the direction of anticipated motion. In line with such a suggestion, Berry and colleagues (1999) found an anticipation effect for visual stimuli moving across the retina (see De sá Teixeira, 2016; Hubbard, 2005, 2014; Kerzel, 2000, for discussion concerning the influence of eye movements on representational momentum). Interestingly, most theories of representational momentum consider this effect to be modality-independent (see Hubbard, 2010).Footnote 1
Localization of dynamic tactile stimuli: Influence of direction and (perceptual) uncertainty
One of the main moderators of the forward shift is the direction of (implied) motion of the moving target. Horizontal moving stimuli elicit a stronger forward shift as compared to stimuli moving vertically (Hubbard, 1990; Hubbard & Bharucha, 1988). In the horizontal plane, most studies have not reported any difference between left- and rightward movement (Cooper & Munger, 1993; Hubbard, 1990, 1995a; Hubbard & Bharucha, 1988; but see Halpern & Kelly, 1993). In the vertical plane, downward movement/descending stimuli tend to elicit stronger forward shifts than upward movement/ascending stimuli, known as representational gravity (e.g., De sá Teixeira, 2016; De sá Teixeira, Hecht, & Oliveira, 2013; Hubbard, 1990, 1995b, 2005). For stimuli that move in the depth plane, a forward shift has been evidenced, yet consistent differences between approaching and receding events have not been reported. Hubbard (1996) documented larger forward shifts for receding targets in his Experiment 1, but was unable to replicate this finding in a subsequent experiment (Experiment 2). Meanwhile, Nagai, Kazai, and Yagi (2002) also failed to find any differences between approaching and receding targets (Experiment 2 – upright posture condition). Overall, these results can be taken to show that the direction of (implied) motion is an important moderator of the forward shift. In this present study, we present dynamic stimuli along the forearm, heading either toward the elbow or the wrist.
Another influential factor on the magnitude of the forward shift is uncertainty. In fact, spatial uncertainty due to an increased blurring of the target (Fu, Shen, & Dan, 2001) or due to decreased spatial acuity (Schmiedchen, Freigang, Rübsamen, & Richter, 2013; see Kanai, Sheth, & Shimojo, 2004, for similar results on the flash-lag effect) has been shown to increase the magnitude of the forward shift. As for the tactile modality, it is well known that different body sites differ in terms of their spatial acuity and discrimination abilities (Cholewiak, 1999; Cholewiak & Collins, 2000; Cholewiak & Craig, 1984; Cody, Garside, Lloyd, & Poliakoff, 2008; Craig & Lyle, 2002; Gallace & Spence, 2014; Green, 1982; Stevens & Choo, 1996; Weinstein, 1968). But even across one specific region of the body surface such as the forearm, for example, spatial acuity can vary. Spatial acuity is highest at the wrist and the elbow and decreases toward the middle of the forearm (Cholewiak & Collins, 2003). Assessing the forward shift at different locations therefore might indicate the influence of spatial acuity on the forward shift. On the other hand, it is important to have at least one location that can be used to assess the forward shift for both motion direction conditions. Only then will spatial acuity be matched for both direction conditions, and hence any difference in the forward shift be attributed to the different directions of motion.
As a central aspect of this study, the tactor (a mechanical device used to present vibrotactile stimulation) location estimate of the implied motion condition is compared to that of the control condition, not to the actual tactor location. The comparison with a control condition is necessary in order to get an estimation of the effect of the implied motion on tactile localization, independent of any general mislocalization at that body site. Following the same reasoning, Getzmann and Lewald (2007, 2009) investigated the influence of motion in auditory space with the help of a control stimulus. In fact, the lack of any control estimate constitutes our main criticism of a previous study by Macauda et al. (2018). For the tactile modality, the authors found a backward shift, but the estimated location was never compared to the perceived location of a control stimulus at that location, only to the actual tactor location. This shortcoming makes the interpretation of their results particularly problematic. At the forearm, the perceived location is not necessarily identical to the actual location, and the perceived location is further influenced by the intensity of the tactile stimulus (Steenbergen et al., 2014). Therefore, the difference between the actual and the perceived location in the study by Macauda and colleagues might, in fact, constitute a general localization error, independent of any influence of a coherent motion in a single direction.
Overview
The goal of the present study was to investigate the influence of implied motion on tactile localization against the background of possible moderators such as motion direction and uncertainty. Five tactors were used to present vibrotactile stimuli to the participant’s left forearm. The tactors were attached in a straight line, one next to the other. Three 250-ms vibrations, separated by 250-ms empty intervals, were presented on each trial. These are the same timing parameters as documented in visual and auditory representational momentum experiments (e.g., Freyd & Finke, 1984; Hubbard & Courtney, 2010). With those timing parameters, three distinct vibrotactile stimuli were perceived. Further, these stimuli have the characteristics of a motion stimulus and imply a motion in a specific direction, but do not elicit tactile mislocalizations attributable to illusions such as the tactile saltation or funneling illusion. The latter are known to influence the perceived location of tactile sensations.Footnote 2 By being presented next to each other in a single consistent direction, the three vibrations implied motion in one direction (implied motion condition). The locations of the three vibrations were chosen at random for the control condition (for examples, see Fig. 1). Using two different dependent variables, participants judged the location of the third vibration as closer to the elbow or wrist (Experiment 1) or else indicated the location on a ruler attached to the forearm (Experiment 2). These two dependent measures were derived and adapted from two of the most oft-used dependent measures, the probe comparison (e.g., Freyd & Finke, 1984; Freyd & Johnson, 1987; Kerzel, 2003) and the absolute estimation task (e.g., Hubbard & Bharucha, 1988; Kerzel & Gegenfurtner, 2003). Typically, probe comparisons involve a comparison of the moving targets’ final location with another target stimuli, presented either at the same or at a slightly different location (e.g., Kerzel, 2003). Due to the experimental set-up (tactors of 3 cm, tactor spacing of 3.5 cm), we could not conduct such a probe comparison. Instead, we used comparisons of each individual target stimulus with a predefined location (i.e., wrist and elbow). Please note that the pointing response that is evaluated in Experiment 2 is a conventional dependent variable used in representational momentum studies (e.g., Kerzel & Gegenfurtner, 2003).

Mapping of actual tactor location and target location on the participant’s arm. The outermost, outer, or central target locations for a distal direction are tactors E, D, and C, respectively; for the proximal direction, tactors A, B, and C, respectively. Schematic display of the three different trials types used for the central target location, namely implied motion trials with proximal and distal motion as well as control trials (one example presented, for detailed description see the main text). Filled black circles indicate a vibration at that location; filled gray circles indicate those tactors that were attached to the arm bandage but never used during the experiments
To foreshadow the results, systematic forward shifts for dynamic stimuli were observed and the direction of implied motion was identified as a moderating factor. The results of Experiment 2 further indicated that uncertainty (here the spatial acuity) independently of the direction of implied motion influenced forward shifts.
Pilot study: Questionnaire on the perception of the stimuli
A pilot study on the perception of the stimuli set-up used in the experiments reported here was conducted. Therefore, a short questionnaire, which focused on motion characteristics inherited by the presented stimuli, was devised.
Methods
Participants
Thirteen participants (nine female, 21–57 years old, none left-handed) from the University of Trier took part in this study. All of the participants reported normal or corrected-to-normal vision and no sensory impairment on the forearm.
Design, apparatus, and stimuli
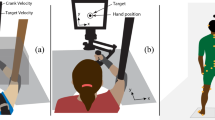
Each participant was tested individually in a dark, sound-attenuated room. All of the furniture was painted black, and all sources of illumination (e.g., from electrical equipment) were eliminated. All of the stimuli were coded in E-Prime (Version 2.0). Visual stimuli were presented on a 24-in. TFT screen controlled by a standard PC. The participant’s forearm was oriented away from his or her body (see Fig. 2), with the dorsal side facing upwards (see Fig. 3). During the experiment, the participants wore a custom-made arm bandage with seven tactors (Model C-2, Engineering Acoustic, Inc.; controlled via the serial interface) on the interior surface.

Bird’s-eye view of the experimental set-up used during Experiment 1 with an additional inset panel showing the placement of the tactors on the skin surface. In the pilot study, and in Experiment 2, the occluder box was omitted; see main text for details

The two postures used for the two experiments. (a) In Experiment 1, the volar side of the forearm faced upward. (b) In Experiment 2 and the pilot study, the dorsal side of the forearm faced upward. For Experiment 2, a ruler was attached on top of the self-made arm bandage
For this study and due to technical restrictions, only the inner five tactors were used. The tactors (3 cm in diameter; 0.79 cm thick; centrally located skin contactor of 0.76 cm) were ordered in a straight line with a center-to-center distance of 3.5 cm. The tactile stimuli (~250 Hz, about 200 μm peak-to-peak amplitude) were applied to the volar side of the left forearm. In order to rule out any impact of the sounds caused by the operation of the tactors, the participants wore ear plugs (Noise reduction: 29 dB) on top of which white noise was presented over headphones (Over-ear headphones: about 95 dB). For this pilot study, a short paper-and-pencil questionnaire with four items was devised that had to be completed after every trial (see Fig. 4). Participants were asked to continue the tactile pattern and to indicate if the next vibration would be located closer to the wrist, the elbow, or in no specific direction (Item 1). On a ten-point scale, participants rated if the sequence felt like one common, continuous event or like individual, separated events (Item 2), and to what extent did the sequence feel like a motion (Item 3) or a random pattern (Item 4).

The four items and results of the questionnaire on the perception of the stimuli. The response frequency (Item 1) and mean rating (Items 2–4) are presented; error bars depict the standard error of the means. The questionnaire was conducted in German, translation by the authors (see Appendix for the original wording)
Procedure
Each trial started with the visual presentation of a plus sign for 400 ms. Thereafter, three vibrotactile stimuli were presented successively for 250 ms each with an interstimulus-interval of 250 ms. Following the offset of the third vibration, the participants were asked to answer the four items of the questionnaire without time constrains. Then the experimenter started a new trial.
Three different conditions were presented, either one of two implied motion trials (in the distal or proximal direction), or a control trial (see Fig. 1). For the implied motion trials, the vibrations were presented adjacent to each other in one single consistent direction. Therefore, these stimuli implied a motion from one tactor to the next along the participant’s forearm. For the control trials, the locations of the vibrations’ locations were selected randomly without replacement with the restriction that implied motion condition trials never occurred. Overall, participants worked through nine trials (three per condition), for all trials, the target vibration was presented at tactor location C (central tactor location; see Fig. 1).
Results and discussion
When asked in which direction participants would continue the presented pattern, participants indicated to expect the pattern to continue in the implied direction (for both implied motion conditions, on average, 2.73 out of three times the participants would continue in the implied direction; see Item 1, Fig. 4).Footnote 3 When asked to what extent the sequence resembled a motion pattern (Item 3), the three different conditions were perceived differently. Overall, the implied motion conditions had a high resemblance to a motion pattern (7.19 on a scale from 0 to 9, Item 3). A one-factorial (implied proximal motion vs. implied distal motion vs. control condition) multivariate analysis of variances (MANOVA)Footnote 4 with Pillai’s trace as criterion revealed a significant main effect, F(2, 11) = 19.73, p < .001, ɳp² = .78. Helmert contrasts revealed that the average of both implied motion sequences resembled more a motion pattern compared to the control pattern, F(1, 12) = 15.60, p = .002, ɳp² = .57. Interestingly, the second contrast revealed that the implied motion sequence for the proximal direction resembled more a motion pattern than the implied motion stimulus in the distal direction condition, F(1, 12) = 5.45, p = .038, ɳp² = .31. Similar results were obtained from Item 4. The MANOVA revealed a significant main effect, F(2, 11) = 32.75, p < .001, ɳp² = .86, when asked to what extent the sequence resembled a random pattern. Once again, Helmert contrasts were conducted and revealed that the control sequence resembled more a random pattern in comparison with the average of both implied motion sequences, F(1, 12) = 23.63, p < .001, ɳp² = .66. The second contrast between the two implied motion sequences was not significant, F(1, 12) = 2.65, p = .130, ɳp² = .18. These results clearly indicate that the presented implied motion sequence is perceived to have more consistent motion characteristics than the control condition.
Experiment 1
In Experiment 1, participants were presented with a sequence of three vibrations at different locations on the forearm. The three vibrations either implied motion along the forearm in a consistent direction, or else they suggested a random pattern. A two-alternative forced choice task (2 AFC) was used to assess the perceived location. The participants’ task was to indicate if they perceived the last vibration, the target vibration, as being located closer to the wrist or to the elbow. A forward shift in the proximal direction was measured as a higher percentage of elbow responses for the proximal direction condition as compared to the control condition. A forward shift for the distal direction condition was measured as a lower percentage of elbow responses for the distal implied motion condition as compared to the control condition. Based on the literature reviewed above, forward shifts for both directions were expected.
Methods
Participants
Based on the effect sizes of studies investigating the influence of (implied) motion on visual and auditory localization (Getzmann & Lewald, 2007, 2009; Cohen’s ds ranging between 0.68 and 1.89), an effect of dz = 0.7 was used for sample size calculations. Those studies used an absolute judgment of the perceived last location (comparable to Experiment 2 of this study). Absolute judgment scores are known to elicit stronger forward shifts as compared to relative probe judgments (Kerzel, 2003; Kerzel & Gegenfurtner, 2003). In Experiment 1, a relative judgment score was used, the target vibration was judged in relation to a landmark (elbow/wrist). Therefore, the expected effect size was reduced to dz = 0.5 for Experiment 1. With α = .05 and a power (1-β) = .9, we calculated a sample size of N = 36 using G*Power Version 3.1.9.2 (Faul, Erdfelder, Lang, & Buchner, 2007). The sample sizes were slightly increased (N = 40) to be able to exclude participants if needed.
The final sample consisted of 38 (22 female; 18–35 years old; three left-handed) students from the University of Trier who took part in this study in return for course credit or a payment of 5€. All of the participants reported normal or corrected-to-normal vision and no sensory impairment on the forearm. The data from one participant was excluded due to a failure to comply with the task instructions. The data from another participant was excluded because he was aware of the purpose of the study.
Design
The experiments were designed with the three within-participants conditions of direction (proximal vs. distal), experimental condition (implied motion vs. control), and target location (central: 0 cm vs. outer: ±3.5 cm vs. outermost: ±7 cm). All of the analyses were computed with the frequency of elbow responses as the dependent variable.
Apparatus, stimuli, and procedure
The apparatus, stimuli, and procedure were identical to the pilot study with the following exceptions. The volar side of the left forearm faced upwards (see Fig. 3) and a black occluder covered the left forearm of the participants during the experiment. The trial procedure was identical to that of the pilot study with the difference that participants didn’t answer a paper-and-pencil questionnaire. The participants had to indicate whether the last vibration in the sequence was perceived closer to the wrist or to the elbow with the help of the computer mouse. After a response had been recorded (or after 2,000 ms had elapsed), the next trial started after an interval of 1,000 ms.
The experiment consisted of a total of 384 trials organized in two fully counterbalanced blocks. All of the five tactor locations were used as the endpoint of the sequence. In half of all trials, the sequence ended at the central tactor location (192 trials). Half of those trials were control trials (96 trials), the other half were implied motion trials, in either the proximal or the distal direction (48 trials each). The remaining trials did not end at the central tactor location, with each of those four target end locations (two outer end locations, locations B and D; two outermost end location, locations A and E) being estimated equally often (24 trials in the implied motion and control conditions, respectively). The outer and outermost end locations were presented at a distance of 3.5 cm and 7 cm from the central location. Importantly, only for the central tactor location, did both motion direction conditions end at the same location, namely tactor location C (see Fig. 1). Besides ending on the central tactor location, proximal direction trials also ended on the outer location B (stimulus sequence: D, C, B) or outermost location A (stimulus sequence: C, B, A). Similarly, distal direction trials ended on the outer location D (stimulus sequence: B, C, D) or outermost location E (stimulus sequence: C, D, E).
Data analysis
Those trials in which the participants did not respond in the 2,000-ms response window (1.01% of the trials) were excluded. We analyzed the frequency of elbow responses with generalized linear mixed effect models (GLMMs) in R (packages “lme4”, Bates, Mächler, Bolker, & Walker, 2015; R Core Team, 2018), accounting for the two-alternative forced choice task by using the binomial-logit function. For each direction of implied motion, we fitted four models to the data (see Table 1). The simplest model explains the data only with an intercept (Model 1, intercept only). For the next three models, we successively added an additional fixed effect. For Model 2, the continuous predictor location was added to Model 1 (ranging from 0 to +7 in the distal direction condition, ranging from 0 to -7 in the proximal direction condition). For Model 3, the previous Model 2 was extended to include the categorical predictor experimental condition (implied motion vs. control). For Model 4, the previous Model 3 was extended to include the interaction between the two predictors (full model; Model 4). We compared each model with its previous model in order to probe if the addition of the fixed effect resulted in a significantly better model fit. The best-fitting model was determined by the last model, which explained the data significantly better. All models treated the intercept of individual participants as random effects.Footnote 5 The raw data, diagrams depicting single participant data with model fitting curves for each participant as well as the R-script are available for open access (Merz, Meyerhoff, Spence, & Frings, 2018).
Results
The visual impression of a forward shift in the proximal direction condition was confirmed by the GLMM analysis. The model with the predictors of location as well as experimental condition was the most appropriate (Model 3; Table 1, I). In other words, a location closer to the elbow (more negative values) resulted in a higher frequency of elbow responses in the proximal direction condition. Importantly, a forward shift was found, the implied motion condition resulted in a higher frequency of elbow responses as compared to the control condition. In contrast, for the distal direction condition, no forward shift was found. The most appropriate model to explain the distal data was Model 2 (Table 1, II), which only incorporated the predictor location. Models with more predictors did not lead to a significantly better fit of the data (see Table 1, sections I and II).
In order to compare the two direction conditions directly, we analyzed the central target location separately. A forward shift, that is, a significant difference between the control and implied motion condition, was found for the proximal, t(37) = 2.27, p = .029, d = 0.37, but not for the distal direction condition, t(37) = -0.23, p = .819. This result fits with the GLMM analysis.
Discussion
In Experiment 1, the participants had to judge whether the last of three sequentially-presented vibrations was perceived closer to the wrist or to the elbow. Our results show a significant forward shift for an implied motion sequence in the proximal direction was obtained. That is, participants perceived the target location as being misplaced further along its trajectory as compared to the control sequence. Notice that this is the first time that a purely tactile forward shift resulting from implied motion has been evidenced.
In contrast to our expectations, no forward shift in implied motion direction was found for the distal sequence. One possibility – which we confirm in Experiment 2 - is that the effect in the distal direction is smaller and thus not appropriately captured by the binominal dependent variable. Therefore, we conceptually replicated and reinvestigated the pattern of results of Experiment 1 with a continuous dependent variable in Experiment 2.
Experiment 2
In Experiment 2, the participants were once again presented with a sequence of three vibrations at different locations on the forearm, which either implied a motion in a single consistent direction or did not. Participants indicated the target location on a ruler attached to their forearm. A 250-mm ruler was attached on top of the arm bandage, but on the dorsal side of the forearm (see Fig. 3). On this ruler, starting with 0 mm at the wrist and ending with 250 mm at the elbow, the participants had to indicate the location of the target vibration. Note that this changes our dependent variable from a relative to an absolute score (each answer indicates the absolute location estimation of the perceived target location).
Methods
Participants
As stated above, based on the effect sizes of studies investigating the influence of (implied) motion on visual and auditory localization (Getzmann & Lewald, 2007, 2009; Cohen’s ds ranging between 0.68 and 1.89), an effect of dz = 0.7 was used for sample size calculations. With α = .05 and a power of (1-β) = .9, we calculated a sample size of N = 19 using G*Power Version 3.1.9.2 (Faul et al., 2007). The sample size was slightly increased (N = 21) to be able to exclude participants if needed. Twenty-one new students (14 female, 20–35 years old, one left-handed) from the University of Trier took part in this study in return for course credit or a payment of 8€. All of the participants reported normal or corrected-to-normal vision and no sensory impairment on the forearm.
Design, apparatus, stimuli, and procedure
The design, apparatus, stimuli, and procedure were identical to Experiment 1 with the following exceptions. The occluder was omitted so that participants were able to see the outline of their forearm. Instead, a 250-mm ruler with 0 mm at the wrist and 250 mm at the elbow was attached to the top of the arm bandage. The ruler was attached to the dorsal side of the forearm, which faced-upward in this experiment (see Fig. 3). The participants were instructed to indicate the location of the last vibration by pointing with their right index finger at the corresponding location on the ruler. The experimenter (who was unaware of the presented sequence) sat next to the participant in order to note the indicated location. The experimenter initiated a new trial when the indicated location was noted and the participant as well as the experimenter were ready.
All of the participants completed 192 experimental trials preceded by eight practice trials. Every location was targeted equally often (16 trials in the implied motion and control conditions, respectively), except for the central location, which served as a target in the proximal as well as the distal direction condition (16 trials each and corresponding 32 control trials). The participants were allowed to pause after every 24 trials.
Data analysis
We analyzed the absolute location estimation with linear mixed effect models (LMMs) in R. As in Experiment 1, we fitted four models to the data of each direction condition (see Table 1). All models treated the intercept of individual participants as random effects.Footnote 6 The raw data, diagrams depicting single participant data with model fitting curves for each participant as well as the R-script are available for open access (Merz, Meyerhoff, Spence, & Frings, 2018).
Results
For both motion direction conditions, a forward shift at the central location, which decreases toward the outer locations, is shown in Fig. 5. The LMM analysis supports this description (see Table 1, sections III and IV). For both motion directions, the data are best explained with the full model (Model 4), which incorporated the fixed effects of location, experimental condition as well as their interaction. As expected, the location of the target vibration influenced the location estimation, a target vibration closer to the elbow was perceived as closer to the elbow. Importantly, for both motion directions, the difference between the implied motion and control condition, the forward shift, was strongest at the central location and decreased toward the outermost location (see Fig. 5, bar diagram). In the proximal direction condition, the implied motion trials were perceived closer to the elbow than the control trials. In the distal direction condition, this was reversed, the implied motion trials were perceived closer to the wrist than the control trials, as expected by the forward shift. For both motion conditions, this difference decreased toward the outermost locations.

Plot of the data from Experiment 1 (upper panel) and Experiment 2 (lower panel) as a function of location (x-axis). Line diagram: Mean scores of the frequency of elbow response (Experiment 1) and absolute location estimation (Experiment 2) for the control as well as both implied motion conditions. Bar diagram: The forward shift, the difference between the implied motion and control score, as a function of direction. Positive values represent an overestimation in implied motion direction. Error bars represent standard errors
In order to compare the magnitude of the forward shift in the two direction conditions, we analyzed the central target location separately. A significant difference between the implied motion and control condition, a forward shift, was observed for the proximal, t(20) = 5.36, p < .001, d = 1.17, as well as the distal direction condition, t(20) = 2.51, p = .021, d = 0.55. For both direction conditions, the central target location was misperceived further along its trajectory as compared to the control condition. The magnitude difference between the forward shifts for the two direction conditions (7.1 mm) was also significant, t(20) = 2.89; p = .009; d = 0.63, indicating a stronger forward shift for the proximal direction compared to the distal direction.
Discussion
In Experiment 2, the participants indicated the target location on a ruler attached to their forearm. A strong forward shift at the central target location for approaching stimuli moving in the proximal direction was documented. This replicates the main finding of Experiment 1 and the influence of implied motion on tactile localization. Furthermore, a forward shift for the distal direction was also found, underlining the importance of implied motion in tactile localization in general. The effect of implied motion on the localization of tactile stimuli was the same for the two directions, with stronger forward shifts for the proximal direction. For both direction conditions, the strength of the forward shift was strongest at the central tactor location and decreased to the outer and outermost tactor locations. This result fits with the change of spatial uncertainty along the forearm (Cholewiak & Collins, 2003), which is discussed in more detail in the General discussion section.
General discussion
In two experiments, we investigated the influence of implied motion on tactile localization. To investigate the effect of implied motion on tactile localization, a control condition was conducted. Systematic differences between the control and the implied motion conditions can only be attributable to the dynamic property of the motion in a single consistent direction. Using two different experimental approaches, a systematic forward shift was observed in the tactile modality for the first time. Participants perceive the last location of a dynamic stimulus to be further along its trajectory. These results are in line with studies using visual or auditory targets (see Hubbard, 2005, 2014).
As one important influencing factor, the direction of implied motion was systematically investigated throughout our study. A stronger forward shift for targets heading in the proximal direction was found. An implied motion stimulus in the proximal direction approaches the elbow but further heads toward the participant’s egocentre at the upper body and the head. We perceive the self at this area of our body (Alsmith & Longo, 2014; Limanowski & Hecht, 2011; Starmans & Bloom, 2012). Furthermore, in their review of the graphesthesia task, Arnold, Spence, and Auvray (2017) identified an overall predominance of the head-centered perspective in the interpretation of ambiguous tactile stimuli presented to the skin surface. Therefore, we argue that the stimuli in our study approach to and recede from our self (i.e., the head/torso). Looking at the forward shift for approaching and receding stimuli in the visual modality, different results have been obtained (Hubbard, 1996; Nagai et al., 2002). As already mentioned in the Introduction, no differences or even greater forward shifts for receding targets (Hubbard, 1996, Experiment 1) were found. This stands in contrast to stronger forward biases for approaching stimuli in the tactile modality, indicating modality-specific differences in the perception of (implied) motion in depth.
Besides the representational momentum literature, there is plenty of evidence for differences between approaching and receding stimuli. For example, looming/approaching stimuli have been shown to elicit stronger psychophysiological (Bach, Neuhoff, Perrig, & Seifritz, 2009; Bach et al., 2007) as well as emotional (Bach et al., 2009; Schiff, Caviness, & Gibson, 1962) responses. Approaching (in comparison to receding) stimuli are perceived as lasting longer (Grassi & Pavan, 2012; Kline & Reed, 2013) and as changing more in terms of their loudness (Neuhoff, 1998, 2001). Such differences are often explained by the differing importance of approaching and receding stimuli for survival. According to the argument that is often made in the literature, approaching stimuli need to be attended to so that an appropriate action can be initiated (such as stepping out of the way if a car is rapidly approaching; Spence, Lee, & Van der Stoep, 2017). Therefore, according to researchers, participants err on the “side of safety” (Neuhoff, 2001, p. 88). This is evidenced, for example, by the underestimation of time of arrival judgments for approaching auditory stimuli (Rosenblum, Wuestefeld, & Saldana, 1993; Schiff & Oblak, 1990). Further, approaching stimuli tend to be mislocalized and perceived as being closer to the observer’s body than are receding stimuli of the same objective distance from the observer (Neuhoff, 2001; Neuhoff, Planisek, & Seifritz, 2009). The error on the side of safety might help to generate responses faster and might increase the chance to be able to respond (if necessary) to the approaching stimulus (see Neuhoff, 2018, for a recent review of the looming literature). Interestingly, both Guski (1992) and Popper and Fay (1997) have attributed a warning function to the auditory system. The auditory system might be helpful in directing an observer’s visual resources toward the event or to initiate a fast response (turn around; jump out of the way) if necessary. Similar functions might underlie the tactile system, thus providing “quick and dirty” information erring on the side of safety.
With the reported experiments here, it is not possible to differentiate between approaching/receding stimuli in a spatiotopic and a somatotopic frame of reference. Participants extended their arms and placed them on the desk (see Fig. 2), but never changed the position of their arms. Therefore, an experiment in which different arm positions were used (e.g., bending the arm at the elbow so that the hand is directed toward the upper body/head, e.g., Gallace, Soto-Faraco, Dalton, Kreukniet, & Spence, 2008; Pritchett, Carnevale, & Harris, 2012) can distinguish between the two spaces of references. Alternatively, the self, located at the upper body/torso, might be interpreted as a landmark. Previous research (Hubbard & Ruppel, 1999) indicates that objects moving toward a landmark undergo landmark attraction, which adds to the representational momentum effect and therefore increases the forward shift.Footnote 7
In this study, an influence of spatial uncertainty on the forward shift was documented. The strength of the forward shift was strongest at the central tactor location and decreased to the outer and outermost tactor locations (Experiment 1 – proximal direction condition; Experiment 2 – proximal and distal direction condition). This is in line with differences in the localization performance along the forearm (Cholewiak & Collins, 2003). The overall localization performance increases from the central to the outermost target location along the forearm. An increase in spatial acuity leads to a decrease in the magnitude of the forward shift. These results are in line with previous findings reported in the visual and auditory modality (Fu et al., 2001; Schmiedchen et al., 2013) and findings with the flash-lag effect (Kanai et al., 2004). Spatial acuity increases from the central to the outermost location, but the stimuli also approach the boundary of the experimental set-up at these extreme locations, which was also found to decrease the forward shift (Hubbard & Motes, 2005).
To date, a few studies already exist investigating the tactile localization of moving stimuli. Due to different foci, and therefore crucial differences in their experimental set-up, it is difficult to draw any conclusion from these studies to the current study. The studies by Seizova-Cajic, Taylor, and colleagues (Nguyen, Taylor, Brooks, & Seizova-Cajic, 2016; Seizova-Cajic & Taylor, 2014) used moving tactile brushing stimuli along the forearm and participants judged the last location where they perceived the brush. The authors focused on the effects of a gap in the tactile stimulation (10 out of 20 cm of skin was not contacted by the brush due to an occluder) and if this cap could be overcome by manipulating the space-time continuum. For this, the tactile stimulation was irregular and not continuous, therefore it is difficult to compare any results with the current study. Trojan et al. (2010) investigated the perceived location of all three tactile stimuli in a reduced rabbit/saltation illusion design. Hereby, the first two stimuli were presented to the same location (stimulus onset asynchrony: 1,020 ms), the third using different stimulus onset asynchronies (20–1,020 ms) 10.5 cm apart. Overall, the third (last) tactile stimuli was misperceived toward the location of the first two stimuli (indicating a backward shift; see Figs. 3 and 4 in their paper). Interestingly, the participants were asked to indicate the location of a single tactile stimulus (see Fig. 2, location 8 in their paper), and this showed a similar mislocalization. Since these two mislocalizations were never compared, it is difficult to draw a conclusion on whether in fact a forward, backward, or no shift would be found with the cutaneous rabbit/saltation illusion.
Whitsel et al. (1986) were interested in the perceived length of a motion stimulus for differing velocities and showed that the perceived length decreases with an increase in velocity. In one of the experiments (Experiment 1), participants judged the perceived start- and endpoint of a motion stimulus in a single consistent direction (experimental stimulus) in comparison to the start- and endpoint of another motion stimulus in a single consistent direction (control stimulus). The control stimulus was also a motion stimulus in one single direction, so no control condition without a motion in a single consistent direction exists. Therefore, no conclusion about the existence of a forward/backward-shift in the tactile modality can be drawn from this study. In contrast, in this study, we used a control condition without motion in a consistent direction by presenting the first two vibrations at random locations. Therefore, the control as well as the implied motion stimuli inherited motion characteristics (as evidenced in the Pilot study). Crucially, the implied motion trials indicated a motion in a single consistent direction with a consistent speed while the control trials did not. This difference resulted in the occurrence of the forward shift. Nevertheless, location estimation of a single, stationary vibration could be used in prospective research. We would not expect any difference between the control condition of the present study and a stationary control.
The notion of where we localize dynamic tactile sensations applied to the skin surface is also relevant in applied contexts. To unload and support the visual sensory system during difficult and attention-demanding tasks (e.g., driving a car in rush hour traffic), the focus was turned to the auditory and especially the tactile modality to present supporting information (e.g., warning signals, Ho, Gray, & Spence, 2014; Meng, Gray, Ho, Ahtamad, & Spence, 2015a; Meng, Ho, Gray, & Spence, 2015b). Interestingly, recent studies found that auditory as well as tactile looming/approaching warning signals decrease braking response times (Gray, 2011; Ho, Spence, & Gray, 2013; Meng, Gray, et al., 2015a; Meng, Ho, et al., 2015b). This can be explained in terms of people erring on the side of safety to be able to initiate fast responses. Combining the existing knowledge about dynamic stimuli and their perception will lead to further developments and improvement in tactile interface design and hopefully to a safer environment for all people.
Conclusions
The present study provides the first demonstration of the influence of implied motion on the localization of sequences of spatially-distributed vibrotactile stimuli. Comparable to the localization of dynamic events in the visual and auditory modalities, a dynamic tactile event is mislocated further along its trajectory. The present data suggest that motion patterns approaching people’s egocenter are perceived further along their trajectory as compared to receding motion patterns. The results would appear to stand in contrast to the localization of approaching and receding stimuli in the visual modality. We speculate that this may be an adaptive mechanism to be able to attend and respond to approaching stimuli quickly. Further, the spatial uncertainty influences the forward shift. With increasing spatial acuity to localize the target stimulus, the influence of implied motion on tactile localization decreases.
Notes
For an exhaustive discussion of the tentative evidence regarding the interplay between localization and motion or motion-like sensations in the tactile modality, see the General discussion.
For the present series of experiments, it was our intention to avoid tactile mislocalization due to illusions like, for example, the funneling illusion (Chen et al., 2003; Gardner & Spencer, 1972). The basic funneling illusion occurs when two simultaneously-presented tactile stimuli are perceived as one tactile stimulus localized in-between the two stimulated locations. Successive tactile stimuli with an interstimulus interval of 250 ms were used in the present study to prevent the occurrence of this illusion. Another illusion that might have influenced the location perception is the tactile saltation or ‘cutaneous rabbit’ illusion (Cholewiak & Collins, 2000; Flach & Haggard, 2006; Geldard, 1982; Geldard & Sherrick, 1972). In this illusion, a tactile stimulus is mislocated toward the location of a preceding or succeeding tactile stimulus. Once again, the occurrence of this illusion in our experiments is very unlikely. A long interstimulus interval combined with a long vibrotactile stimulation duration (250 ms) was used. In contrast, those studies that have investigated the cutaneous rabbit illusion have typically used shorter interstimulus intervals (about 200 ms or less) as well as durations (about 100 ms and less; e.g., Blankenburg, Ruff, Deichmann, Rees, & Driver, 2006; Flach & Haggard, 2006; Geldard & Sherrick, 1972).
For Item 1, two participants had to be excluded since they reported afterwards that they had not read the question properly and therefore answered wrongly.
Note that all repeated-measures designs are inherently multivariate and the MANOVA has the advantage that sphericity cannot influence the results (see e.g., Tabachnick & Fidell, 2007).
Reanalyzing the models with the intercept as well as the slopes of the two predictor variables of location and condition of individual participants as well as their interaction treated as random effects identified the same best-fitting models.
Reanalyzing the models with the intercept as well as the slopes of the two predictor variables of location and condition of individual participants as well as their interaction treated as random effects identified the same best-fitting models.
We thank an anonymous reviewer for bringing this potential explanation to our attention.
References
Alsmith, A. J., & Longo, M. R. (2014). Where exactly am I? Self-location judgements distribute between head and torso. Consciousness and Cognition, 24, 70-74.
Arnold, G., Spence, C., & Auvray, M. (2017). A unity of the self or a multiplicity of locations? How the graphesthesia task sheds light on the role of spatial perspectives in bodily self-consciousness. Consciousness and Cognition, 56, 100-114.
Bach, D. R., Neuhoff, J. G., Perrig, W., & Seifritz, E. (2009). Looming sounds as warning signals: The function of motion cues. International Journal of Psychophysiology, 74(1), 28-33.
Bach, D. R., Schächinger, H., Neuhoff, J. G., Esposito, F., Salle, F. D., Lehmann, C., ... Seifritz, E. (2007). Rising sound intensity: An intrinsic warning cue activating the amygdala. Cerebral Cortex, 18(1), 145-150.
Bates, D., Mächler, M., Bolker, B., & Walker, S. (2015). Fitting linear mixed-effects models using lme4. Journal of Statistical Software, 67(1), 1-48.
Berry, M. J., Brivanlou, I. H., Jordan, T. A., & Meister, M. (1999). Anticipation of moving stimuli by the retina. Nature, 398(6725), 334-338.
Blankenburg, F., Ruff, C. C., Deichmann, R., Rees, G., & Driver, J. (2006). The cutaneous rabbit illusion affects human primary sensory cortex somatotopically. PLoS Biology, 4(3):e69.
Brugger, P., & Meier, R. (2015). A new illusion at your elbow. Perception, 44(2), 219-221.
Chen, L. M., Friedman, R. M., & Roe, A. W. (2003). Optical imaging of a tactile illusion in Area 3b of the primary somatosensory cortex. Science, 302(5646), 881-885.
Cholewiak, R. W. (1999). The perception of tactile distance: Influences of body site, space, and time, Perception, 28(7), 851-875.
Cholewiak, R. W., & Collins, A. A. (2000). The generation of vibrotactile patterns on a linear array: Influences of body site, time, and presentation mode, Attention, Perception, & Psychophysics, 62(6), 1220-1235.
Cholewiak, R. W., & Collins, A. A. (2003). Vibrotactile localization on the arm: Effects of place, space, and age, Attention, Perception, & Psychophysics, 65(7), 1058-1077.
Cholewiak, R. W., & Craig, J. C. (1984). Vibrotactile pattern recognition and discrimination at several body sites. Attention, Perception, & Psychophysics, 35(6), 503-514.
Clark, A. (2013). Whatever next? Predictive brains, situated agents, and the future of cognitive science. Behavioral and Brain Sciences, 36(3), 181-204.
Cody, F. W., Garside, R. A., Lloyd, D., & Poliakoff, E. (2008). Tactile spatial acuity varies with site and axis in the human upper limb. Neuroscience Letters, 433(2), 103-108.
Cooper, L. A., & Munger, M. P. (1993). Extrapolating and remembering positions along cognitive trajectories: Use and limitations of analogies to physical motion. In N. Eilan, R. A. McCarthy, & B. Brewer (Eds.), Spatial representation: Problems in philosophy and psychology (pp. 112-131). Cambridge: Blackwell Publishing.
Craig, J. C., & Lyle, K. B. (2002). A correction and a comment on Craig and Lyle (2001). Attention, Perception, & Psychophysics, 64(3), 504-506.
De Sá Teixeira, N. A. (2016). The visual representations of motion and of gravity are functionally independent: Evidence of a differential effect of smooth pursuit eye movements. Experimental Brain Research, 234(9), 2491-2504.
De Sá Teixeira, N. A., Hecht, H., & Oliveira, A. M. (2013). The representational dynamics of remembered projectile locations. Journal of Experimental Psychology: Human Perception and Performance, 39(6), 1690-1699.
Faul, F., Erdfelder, E., Lang, A. G., & Buchner, A. (2007). G* Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behavior Research Methods, 39(2), 175-191.
Flach, R., & Haggard, P. (2006). The cutaneous rabbit revisited. Journal of Experimental Psychology: Human Perception and Performance, 32(3), 717-732.
Freyd, J. J. (1987). Dynamic mental representations. Psychological Review, 94(4), 427-438.
Freyd, J. J., & Finke, R. A. (1984). Representational momentum. Journal of Experimental Psychology: Learning, Memory, and Cognition, 10(1), 126-132.
Freyd, J. J., & Johnson, J. Q. (1987). Probing the time course of representational momentum. Journal of Experimental Psychology: Learning, Memory, and Cognition, 13(2), 259-269.
Fu, Y. X., Shen, Y., & Dan, Y. (2001). Motion-induced perceptual extrapolation of blurred visual targets. Journal of Neuroscience, 21(20):RC172:1-5.
Gallace, A., Soto-Faraco, S., Dalton, P., Kreukniet, B., & Spence, C. (2008). Response requirements modulate tactile spatial congruency effects. Experimental Brain Research, 191(2), 171-186.
Gallace, A., & Spence, C. (2014). In touch with the future: The sense of touch from cognitive neuroscience to virtual reality. Oxford: Oxford University Press.
Gardner, E. P., & Spencer, W. A. (1972). Sensory funneling. I. Psychophysical observations of human subjects and responses of cutaneous mechanoreceptive afferents in the cat to patterned skin stimuli. Journal of Neurophysiology, 35(6), 925-953.
Geldard, F. A. (1982). Saltation in somesthesis. Psychological Bulletin, 92(1), 136-175.
Geldard, F. A., & Sherrick, C. E. (1972). The cutaneous" rabbit": A perceptual illusion. Science, 178(4057), 178-179.
Getzmann, S., & Lewald, J. (2007). Localization of moving sound. Perception & Psychophysics, 69(6), 1022-1034.
Getzmann, S., & Lewald, J. (2009). Constancy of target velocity as a critical factor in the emergence of auditory and visual representational momentum. Experimental Brain Research, 193(3), 437-443.
Grassi, M., & Pavan, A. (2012). The subjective duration of audiovisual looming and receding stimuli. Attention, Perception, & Psychophysics, 74(6), 1321-1333.
Gray, R. (2011). Looming auditory collision warnings for driving. Human Factors, 53(1), 63-74.
Green, B. G. (1982). The perception of distance and location for dual tactile pressures. Attention, Perception, & Psychophysics, 31(4), 315-323.
Guski, R. (1992). Acoustic tau: An easy analogue to visual tau? Ecological Psychology, 4(3), 189-197.
Halpern, A. R., & Kelly, M. H. (1993). Memory biases in left versus right implied motion. Journal of Experimental Psychology: Learning, Memory, and Cognition, 19(2), 471-484.
Harrar, V., & Harris, L. R. (2009). Eye position affects the perceived location of touch. Experimental Brain Research, 198(2-3), 403-410.
Ho, C., Gray, R., & Spence, C. (2014). Reorienting driver attention with dynamic tactile cues. IEEE Transactions on Haptics, 7(1), 86-94.
Ho, C., & Spence, C. (2007). Head orientation biases tactile localization. Brain Research, 1144, 136-141.
Ho, C., Spence, C., & Gray, R. (2013). Looming auditory and vibrotactile collision warnings for safe driving. In Proceedings of the 7th International Driving Symposium on Human Factors in Driver Assessment, Training, and Vehicle Design (No. 83, pp. 551-557).
Huang, Y., & Rao, R. P. (2011). Predictive coding. Wiley Interdisciplinary Reviews: Cognitive Science, 2(5), 580-593.
Hubbard, T. L. (1990). Cognitive representation of linear motion: Possible direction and gravity effects in judged displacement. Memory & Cognition, 18(3), 299-309.
Hubbard, T. L. (1995a). Cognitive representation of motion: Evidence for friction and gravity analogues. Journal of Experimental Psychology: Learning, Memory, and Cognition, 21(1), 241-254.
Hubbard, T. L. (1995b). Environmental invariants in the representation of motion: Implied dynamics and representational momentum, gravity, friction, and centripetal force. Psychonomic Bulletin & Review, 2(3), 322-338.
Hubbard, T. L. (1996). Displacement in depth: Representational momentum and boundary extension. Psychological Research, 59(1), 33-47.
Hubbard, T. L. (2005). Representational momentum and related displacements in spatial memory: A review of the findings. Psychonomic Bulletin & Review, 12(5), 822-851.
Hubbard, T. L. (2010). Approaches to representational momentum: Theories and models. In R. Nijhawan & B. Khurana (Eds.), Space and time in perception and action (pp. 338-365). Cambridge: Cambridge University Press.
Hubbard, T. L. (2014). Forms of momentum across space: Representational, operational, and attentional, Psychonomic Bulletin & Review, 21(6), 1371-1403.
Hubbard, T. L. (2018). Influences on representational momentum. In T. L. Hubbard (Ed.), Spatial biases in perception and cognition (pp. 180-190). Cambridge, Cambridge University Press.
Hubbard, T. L., & Bharucha, J. J. (1988). Judged displacement in apparent vertical and horizontal motion. Perception & Psychophysics, 44(3), 211-221.
Hubbard, T. L., & Courtney, J. R. (2010). Cross-modal influences on representational momentum and representational gravity. Perception, 39(6), 851-862.
Hubbard, T. L., & Motes, M. A. (2005). An effect of context on whether memory for initial position exhibits a Fröhlich effect or an onset repulsion effect. The Quarterly Journal of Experimental Psychology Section A, 58(6), 961-979.
Hubbard, T. L., & Ruppel, S. E. (1999). Representational momentum and the landmark attraction effect. Canadian Journal of Experimental Psychology, 53(3), 242-256.
Kanai, R., Sheth, B. R., & Shimojo, S. (2004). Stopping the motion and sleuthing the flash-lag effect: Spatial uncertainty is the key to perceptual mislocalization. Vision Research, 44(22), 2605-2619.
Kerzel, D. (2000). Eye movements and visible persistence explain the mislocalization of the final position of a moving target. Vision Research, 40(27), 3703-3715.
Kerzel, D. (2003). Mental extrapolation of target position is strongest with weak motion signals and motor responses. Vision Research, 43(25), 2623-2635.
Kerzel, D., & Gegenfurtner, K. R. (2003). Neuronal processing delays are compensated in the sensorimotor branch of the visual system. Current Biology, 13(22), 1975-1978.
Kline, S. R., & Reed, C. L. (2013). Contextual influences of dimension, speed, and direction of motion on subjective time perception. Attention, Perception, & Psychophysics, 75(1), 161-167.
Limanowski, J., & Hecht, H. (2011). Where do we stand on locating the self? Psychology, 2(04), 312-317.
Longo, M. R. (2017). Expansion of perceptual body maps near–but not across–the wrist. Frontiers in Human Neuroscience, 11:111.
Macauda, G., Lenggenhager, B., Meier, R., Essick, G., & Brugger, P. (2018). Tactile motion lacks momentum. Psychological Research, 82, 889-895.
Mancini, F., Longo, M. R., Iannetti, G. D., & Haggard, P. (2011). A supramodal representation of the body surface. Neuropsychologia, 49(5), 1194-1201.
Margolis, A. N., & Longo, M. R. (2015). Visual detail about the body modulates tactile localisation biases. Experimental Brain Research, 233(2), 351-358.
Medina, J., & Coslett, H. B. (2016). What can errors tell us about body representations?. Cognitive Neuropsychology, 33(1-2), 5-25.
Meng, F., Gray, R., Ho, C., Ahtamad, M., & Spence, C. (2015a). Dynamic vibrotactile signals for forward collision avoidance warning systems. Human Factors, 57(2), 329-346.
Meng, F., Ho, C., Gray, R., & Spence, C. (2015b). Dynamic vibrotactile warning signals for frontal collision avoidance: towards the torso versus towards the head. Ergonomics, 58(3), 411-425.
Merz, S., Meyerhoff, H.S., Spence, C., & Frings, C. (2018). Data for ‘Implied tactile motion: localizing dynamic stimulations on the skin’ [Data files, single subject diagrams and R-scripts]. Available from doi:https://doi.org/10.23668/psycharchives.1848
Müsseler, J., Stork, S., & Kerzel, D. (2002). Comparing mislocalizations with moving stimuli: The Fröhlich effect, the flash-lag, and representational momentum, Visual Cognition, 9(1-2), 120-138.
Nagai, M., Kazai, K., & Yagi, A. (2002). Larger forward memory displacement in the direction of gravity. Visual Cognition, 9(1-2), 28-40.
Neuhoff, J. G. (1998). Perceptual bias for rising tones. Nature, 395(6698), 123-124.
Neuhoff, J. G. (2001). An adaptive bias in the perception of looming auditory motion. Ecological Psychology, 13(2), 87-110.
Neuhoff, J. G. (2018). Adaptive biases in visual and auditory looming perception. In T. L. Hubbard (Ed.), Spatial biases in perception and cognition (pp. 180-190). Cambridge: Cambridge University Press.
Neuhoff, J. G., Planisek, R., & Seifritz, E. (2009). Adaptive sex differences in auditory motion perception: Looming sounds are special. Journal of Experimental Psychology: Human Perception and Performance, 35(1), 225-234.
Nguyen, E. H., Taylor, J. L., Brooks, J., & Seizova-Cajic, T. (2016). Velocity of motion across the skin influences perception of tactile location. Journal of Neurophysiology, 115(2), 674-684.
Popper, N., & Fay, R. (1997). Evolution of the ear and hearing: Issues and questions. Brain, Behavior and Evolution, 50(4), 213-221.
Pritchett, L. M., Carnevale, M. J., & Harris, L. R. (2012). Reference frames for coding touch location depend on the task. Experimental Brain Research, 222(4), 437-445.
Rao, R. P., & Ballard, D. H. (1999). Predictive coding in the visual cortex: A functional interpretation of some extra-classical receptive-field effects. Nature Neuroscience, 2(1), 79-87.
Rosenblum, L. D., Wuestefeld, A. P., & Saldaña, H. M. (1993). Auditory looming perception: Influences on anticipatory judgments. Perception, 22(12), 1467-1482.
Schiff, W., Caviness, J. A., & Gibson, J. J. (1962). Persistent fear responses in rhesus monkeys to the optical stimulus of “looming”. Science, 136(3520), 982-983.
Schiff, W., & Oldak, R. (1990). Accuracy of judging time to arrival: Effects of modality, trajectory, and gender, Journal of Experimental Psychology: Human Perception and Performance, 16(2), 303-316.
Schmiedchen, K., Freigang, C., Rübsamen, R., & Richter, N. (2013). A comparison of visual and auditory representational momentum in spatial tasks. Attention, Perception, & Psychophysics, 75(7), 1507-1519.
Seizova-Cajic, T., & Taylor, J. L. (2014). Somatosensory space abridged: Rapid change in tactile localization using a motion stimulus. PLoS One, 9(3), e90892.
Spence, C., Lee, J., & van der Stoep, N. (2017). Responding to sounds from unseen locations: Crossmodal attentional orienting in response to sounds presented from the rear. European Journal of Neuroscience, 1-17. doi:https://doi.org/10.1111/ejn.13733
Starmans, C., & Bloom, P. (2012). Windows to the soul: Children and adults see the eyes as the location of the self. Cognition, 123(2), 313-318.
Steenbergen, P., Buitenweg, J. R., Trojan, J., & Veltink, P. H. (2014). Tactile localization depends on stimulus intensity. Experimental Brain Research, 232(2), 597-607.
Stevens, J. C., & Choo, K. K. (1996). Spatial acuity of the body surface over the life span. Somatosensory & Motor Research, 13(2), 153-166.
Tabachnick, B. G., & Fidell, L. S. (2007). Using multivariate statistics (5th). New York: Allyn & Bacon.
Team, R. C. (2018). R: A language and environment for statistical computing. R Foundation of Statistical Computing, Vienna. Available from https://www.R-project.org/ .
Trojan, J., Kleinböhl, D., Stolle, A. M., Andersen, O. K., Hölzl, R., & Arendt-Nielsen, L. (2006). Psychophysical ‘perceptual maps’ of heat and pain sensations by direct localization of CO2 laser stimuli on the skin. Brain Research, 1120(1), 106-113.
Trojan, J., Stolle, A. M., Mršić Carl, A., Kleinböhl, D., Tan, H. Z., & Hölzl, R. (2010). Spatiotemporal integration in somatosensory perception: Effects of sensory saltation on pointing at perceived positions on the body surface. Frontiers in Psychology, 1:206.
Weinstein, S. (1968). Intensive and extensive aspects of tactile sensitivity as a function of body part, sex, and laterality. In D. R. Kenshal (Ed.), The skin senses (pp. 195-222). Springfield: Thomas.
Whitsel, B. L., Franzen, O., Dreyer, D. A., Hollins, M., Young, M., Essick, G. K., & Wong, C. (1986). Dependence of subjective traverse length on velocity of moving tactile stimuli. Somatosensory Research, 3(3), 185-196.
Acknowledgements
The research reported here was supported by a grant from the Deutsche Forschungsgemeinschaft to Christian Frings and Charles Spence (FR2133/5-3). We would like to thank Stephanie Blasl for the drawings incorporated in Figs. 1, 2, and 3 and Moritz Breit and Johannes Stricker for their help with data collection.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Appendix
Appendix
Pilot Study – Original German wording
-
Item 1:
Setze das Muster fort: Im Vergleich zur letzten / dritten Vibration, wäre die nächste Vibration … [mark one of the three possibilities] (a) … näher am Handgelenk, (b) … näher am Ellenbogen, (c)...in keiner bestimmten Richtung/weiß nicht.
-
Item 2:
Haben sich die drei Vibrationen wie ein gemeinsames, durchgängiges Event oder wie einzelne, separate und nicht zusammengehörige Events angefühlt? [Ten-point scale, ranging from] gemeinsames, durchgängiges Event [to] einzelne, separate Events.
-
Item 3:
In wie weit hat sich die letzte Vibrationsabfolge als ein Bewegungsmuster angefühlt? [Ten-point scale, ranging from] Überhaupt nicht wie ein Bewegungsmuster [to] wie ein Bewegungsmuster.
-
Item 4:
In wie weit hat sich die letzte Vibrationsabfolge als ein Zufallsmuster angefühlt? [Ten-point scale, ranging from] Überhaupt nicht wie ein Zufallsmuster [to] wie ein Zufallsmuster.
Rights and permissions
About this article

Cite this article
Merz, S., Meyerhoff, H.S., Spence, C. et al. Implied tactile motion: Localizing dynamic stimulations on the skin. Atten Percept Psychophys 81, 794–808 (2019). https://doi.org/10.3758/s13414-018-01645-9
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-018-01645-9