
Abstract
Consciousness and working memory are two widely studied cognitive phenomena. Although they have been closely tied on a theoretical and neural level, empirical work that investigates their relation is largely lacking. In this study, the relationship between visual consciousness and different working memory components is investigated by using a dual-task paradigm. More specifically, while participants were performing a visual detection task to measure their visual awareness threshold, they had to concurrently perform either an executive or visuospatial working memory task. We hypothesized that visual consciousness would be hindered depending on the type and the size of the load in working memory. Results showed that maintaining visuospatial content in working memory hinders visual awareness, irrespective of the amount of information maintained. By contrast, the detection threshold was progressively affected under increasing executive load. Interestingly, increasing executive load had a generic effect on detection speed, calling into question whether its obstructing effect is specific to the visual awareness threshold. Together, these results indicate that visual consciousness depends differently on executive and visuospatial working memory.
Similar content being viewed by others
The role of consciousness has intrigued memory researchers since Ebbinghaus (1885). A classical view is that working memory (WM) processes are strictly related to conscious input and operations. This view is echoed in theories of consciousness that similarly propose an intricate relation between WM and consciousness based on their remarkable resemblance (Baars, 1993). First, both consciousness and WM are believed to be highly limited in capacity, temporarily accommodating one unitary concept in consciousness (Crick & Koch, 1990) or maximally four items in WM (Cowan, 2001). Second, both provide a platform for integrating information in a multimodal way (Baddeley, 2000; Zeki & Bartels, 1999). Third, consciousness and WM have both been related to temporally maintaining information in similar reverberant neural circuits (D’Esposito, 2007; Sergent, Baillet, & Dehaene, 2005). This point was even made more clearly in an fMRI meta-analysis, showing a largely overlapping frontoparietal network for both consciousness and WM (Naghavi & Nyberg, 2005).
Although consciousness and WM clearly share important functional properties, how these properties are related has only recently become a topic of investigation. In one line of research, the influence of WM content on consciousness is investigated. This line of research focuses on the maintenance of visuospatial representations and typically probes consciousness by measuring the threshold for visual awareness with a technique called “breaking continuous flash suppression” (b-CFS; Jiang, Costello, & He, 2007). In b-CFS, a dynamical pattern that is flashed to one eye suppresses the visibility of a static target stimulus that is simultaneously presented to the other eye (Tsuchiya & Koch, 2005). Participants are asked to respond as soon as the target stimulus breaks through the binocular rivalry and they become aware of it, which typically takes a few seconds. In one of these studies, participants remembered a visual patch (i.e., a color) while performing a circle detection task (Gayet, Paffen, & Van der Stigchel, 2013). Results showed that visuospatial WM content influenced the awareness threshold for the circles; it was lower when the circle’s color matched the color maintained in WM compared with when the circle's color differed from that in WM. Similar results were obtained in a series of experiments by Pan, Lin, Zhao, and Soto (2014) in which the awareness threshold in a face detection task was lower when the target face on the screen was identical to the face that participants stored in memory. Together, these studies suggest that the content in visuospatial WM can influence the threshold for visual awareness, with WM possibly biasing the competition for conscious access.
In another line of research, consciousness has been investigated in relation to WM processes typically involving prefrontal functions. In one study, participants memorized a string of letters while they were performing a masking task in which they had to detect a target number (De Loof, Verguts, Fias, & Van Opstal, 2013). Results showed that detection performance decreased with the number of letters that needed to be remembered, indicating that the awareness threshold increases with a frontal WM load (Lavie, 2005). This effect was recently replicated in a task where participants had to detect a briefly presented grating while remembering and manipulating 1 or 4 letters (Maniscalco & Lau, 2015), thus showing that a higher load on executive WM resulted in an increased threshold for visual awareness.
These previous findings suggest that WM directly influences consciousness. However, questions remain about the exact nature of their relation. It remains, for example, unknown whether an increase in the quantity of the remembered items in visuospatial WM will have the same effect on the awareness threshold as loading executive WM. Opposite effects of increasing executive versus visuospatial WM load on visual perception have already been demonstrated in a visual search. Konstantinou and Lavie (2013), for example, demonstrated that detection sensitivity for a peripheral stimulus enhanced with increasing executive WM load while it reduced with increasing visuospatial WM load. To investigate how executive and visuospatial WM load affect the threshold for visual awareness, in this study we used a dual-task approach in which one task, a detection task, was used to measure visual consciousness and the other task to load WM. In the detection task, the threshold for visual awareness was measured with b-CFS. In addition, non-rivalry control trials were added to verify whether detection times truly reflected the awareness threshold or more general decision processes (Jiang et al., 2007). WM was loaded either by a secondary visuospatial WM task or a secondary executive WM task. To be able to study the effect of load within each WM component, secondary tasks were given with low and high loads.
Method
Participants
Nineteen participants (9 males, mean age: 23.4 years) gave informed consent and were paid €10 for participation. All participants were naive to the purpose of the experiment.
Stimuli and material
The stimulus set consisted of twenty pictures of Caucasian faces from the Face Database of the Park Aging Mind Laboratory (10 males, aged 19-79 years; Minear & Park, 2004). All pictures (3.6° × 3.6° visual angle) were luminance scaled to avoid additional luminance-based variation in reaction times (RTs). Twenty Mondrians were created (15° × 15° visual angle) consisting of squares with a width and height ranging from 0.7° to 3.2° visual angle in all possible RGB palette colors.
The experiment was run on a DELL Latitude E6430 laptop running Windows 7 Professional and an external DELL E2213 screen (1680 × 1050; 100 Hz). Stimuli were generated using Psychtoolbox (Brainard, 1997; Pelli, 1997) in MATLAB 2013a (Mathworks). Responses were collected through a Cedrus RB-730 response box.
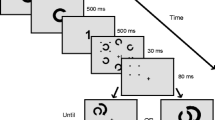
Participants were seated approximately 90 cm from the screen and wore stereoscopic goggles to merge the visual input from both sides of the screen into one visual percept through binocular fusion. The screen was divided into a left and right half (see Fig. 1 for an illustration of the screen layout and task design). On each half of the screen the stimulus display extended over a 15°×15° visual angle square, surrounded by a checkerboard frame to facilitate binocular fusion (checkers measuring 1°×1° visual angle).

The screen layout and task design are illustrated for both types of WM tasks (a: visuospatial and executive WM in the low and high load version) and detection tasks (b: b-CFS trials; c: control trials). The visuospatial WM task starts with the presentation of the 16 rectangles with one (low load) or six (high load) rectangles in the target color for 2 seconds. The executive WM task starts with the instruction to count back in steps of one (low load) or three (high load) followed by a digit number for 2 seconds. In the b-CFS version of the detection task (b), the Mondrians are presented to the right eye and the face to the left eye. In the control version of the detection task (c), the face is blended into the Mondrians and presented to both eyes
In the b-CFS version of the detection task, the Mondrians were presented to the right eye and the face pictures to the left eye. The Mondrians changed at a rate of 10 Hz and covered the entire right side of the stimulus display. The face picture was presented on the top or bottom half of the left display. The visibility of the faces gradually increased at 50 Hz from none to full visibility over a span of 4.5 seconds to avoid an immediate breakthrough (Jiang et al., 2007).
The non-rivalry control version of the detection task was identical to the b-CFS version, but now both the Mondrians and the face picture were presented at both sides of the display.
Procedure and design
Participants started with a practice phase of 20 b-CFS trials (without WM load) to become accustomed to the binocular presentation. Participants were instructed to respond as fast as possible when a picture became visible. The left and right index fingers were used to indicate whether the picture appeared at the bottom or top position and were positioned in corresponding locations on the response box. The participants were reminded that they had to press the button as soon as they saw any (part of the) picture and were not supposed to postpone their answer until they were sure of the picture’s identity.
Next, participants were instructed about the WM tasks and performed two b-CFS trials of each WM task in both the low and high load version. If participants had no further questions the experiment was started. The 2 (Masking: b-CFS vs. control) × 2 (WM task: visuospatial vs. executive) × 2 (Load: low vs. high) blocked design resulted in a total of 8 blocks. Each block consisted of 35 trials and was followed by a break. The position of the picture on the screen (bottom versus top) was balanced across conditions.
In the visuospatial WM task (based on that from Sheremata, Bettencourt, & Somers, 2010), an array of 16 blue and red rectangles was presented at the start of each trial. Each of the rectangles had a horizontal or vertical orientation. For half of the participants, the blue rectangles were the targets and the red rectangles the distracters (instruction: “Remember the position of the blue rectangles”). The reversed mapping was applied for the other half of the participants. In the low-load version, only one rectangle was presented in the target color while in the high load version there were six targets (Fig. 1). The array of rectangles remained on screen for 2 seconds and participants were instructed to remember the location and orientation of the targets. Next, participants performed the detection task. At the end of the trial the array of rectangles reappeared. In half of the trials this array was identical to the one presented at the start, whereas in the other half one target rectangles had changed orientation. Participants responded with their left or right index finger to indicate whether or not they thought one of the target rectangles had changed orientation.
For the executive WM task, participants were instructed at the start of each trial to either count back in steps of one (low load) or in steps of three (high load). Next, a random number between 70 and 100 appeared and the participant started counting back out loud. After 2 seconds, the number disappeared from the screen and the presentation of the Mondrians and face picture started. When the face picture had been detected and a response was given, the participant could stop counting backwards. The end of the trial was announced and participants could start the following trial by pushing a button. Although performance was not being recorded, participants were told that an audio recording was made for scoring afterwards.
Results
All participants performed well throughout the entire task, amounting to an average of 97 % correctly located pictures (accuracy range: 92.7 % to 99.6 %). All further analyses are therefore performed on the data of all 19 subjects, excluding the 20 practice trials for the detection task and the 8 practice trials for the WM tasks. Together with error trials (i.e., trials where the picture was not located correctly; 2.5 % of the data), trials with RTs below 350 ms were labeled as invalid (0.44 % of the data) and excluded from further analysis.
WM task performance showed a clear difference in the error rate between the low and high load version of the visuospatial WM task. In the low-load version, the error rate ranged from 1.47 % to 40.6 % (mean: 15.39 %), whereas in the high-load version the error rate ranged from 28.8 % to 49.3 % (mean: 41.6 %). All participants made more errors in the high compared with the low-load version. Furthermore, RTs were significantly higher in the high-load (1858 ms) compared with the low-load condition (983 ms), χ2(1, N = 19) = 1044, p < 0.001, indicating that participants continued to perform the task despite the high error rate in the high-load condition. No performance was recorded during the executive task.
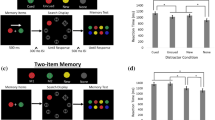
To investigate the influence of the WM task manipulations on the detection task, the RTs on the detection task were entered in a linear mixed model with a random intercept per participant, a random intercept per target picture and the interaction between Masking (b-CFS vs. control), WM task (visuospatial vs. executive) and Load (low vs. high) as fixed effect predictors. There was a significant main effect of Masking, χ2(1, N = 19) = 1123, p < 0.001, with a higher detection threshold on b-CFS trials compared with control trials. There was no main effect of WM task, χ2(1, N = 19) = 2.11, p = 0.15, but there was a very strong and significant interaction between Masking and WM task, χ2(1, N = 19) = 26.2, p < 0.001. As Fig. 2a indicates, RTs in the control version of the detection task were higher under executive load compared with visuospatial load, but this pattern reversed in the b-CFS version where RTs were higher under visuospatial load compared to executive load.

The analysis of the detection task RTs revealed a significant interaction between WM task and Masking (a: b-CFS and control trials on the horizontal axis) and a significant interaction between WM task and Load (b: low and high load on the horizontal axis). The average RTs and their 95 % confidence intervals (vertical axis, in seconds) are presented for the executive (blue) and visuospatial WM task (red). (a) There is a main effect of Masking (p < 0.001) with faster detection in control trials compared with b-CFS trials. However, a significant interaction between WM task and Masking reveals that the difference in RTs between b-CFS trials and control trials is larger in the visuospatial WM task compared to the executive WM task. (b) Follow-up analysis on the significant interaction between WM task and Load (p < 0.001) show that RTs during the executive WM task are higher under high load compared with low load (p < 0.001), but there is no effect of Load during the visuospatial WM task (p = 0.81)
Second, there was a main effect of Load, χ2(1, N = 19) = 25.8, p < 0.001, with the high-load version of the tasks resulting in higher detection thresholds. However, this main effect of Load needs to be interpreted in the light of a significant interaction between Load and WM task, χ2(1, N = 19) = 22.6, p < 0.001. As illustrated in Fig. 2b, follow-up tests revealed that high load trials resulted in higher detection threshold in the executive WM task, χ2(1, N = 19) = 60, p < 0.001, but not in the visuospatial WM task, χ2(1, N = 19) = 0.070, p = 0.79. There was no significant interaction between Masking and Load, χ2(1, N = 19) = 2.27, p = 0.13, and no significant three-way interaction, χ2(1, N = 19) = 1.49, p = 0.22.
Discussion
The results of this study revealed two main findings. First, the visuospatial WM task delayed detection more in the b-CFS trials compared with the non-rivalry control trials. Second, an increase in executive WM load resulted in slower detection times while an increase in visuospatial WM did not. Taken together, these results give us an unprecedented overview of how different aspects of WM interact with visual awareness, as we will discuss below.
First, maintenance of visuospatial information in WM hinders the breakthrough from rivalry in the b-CFS trials more than it hinders the detection process in the control trials. Crucially, the continuous rivalry process underlying CFS is thought to tax visual processing much heavier compared to the decision process of the control trials (Clark, 2013; Yuval-Greenberg & Heeger, 2013). Because the sheer maintenance of visual information—irrespective of the load—involves similar visual processing areas (Ruchkin, Grafman, Cameron, & Berndt, 2003), it likely interfered with b-CFS more than with the control trials. To our knowledge, no other studies have compared the effects of visuospatial WM maintenance on different detection paradigms and further investigation is needed to verify this claim.
Second, in line with early theories and previous empirical work it was expected that visual awareness is directly related to executive functions. Indeed, an increase in prefrontal executive load increased the awareness threshold in a masking study (De Loof et al., 2013), and in visual detection (Maniscalco & Lau, 2015). In the current study, these results are replicated with a different detection task (b-CFS), and with a secondary task that is better suited to load executive functions as the backward counting task crucially involves manipulation of WM content and not only maintenance (D’Esposito, Postle, Ballard, & Lease, 1999). Interestingly, our results reveal that detection was not only progressively hindered by increasing executive WM load in the b-CFS trials but also in the non-rivalry control trials. This calls into question whether executive WM load specifically influences the threshold for visual awareness or rather affects more general decision processes. Indeed, the same prefrontal areas assumed to play a pivotal role in consciousness (Dehaene & Naccache, 2001) are also critically involved in manipulating information in WM (Cohen et al., 1997) and in decisional processes (Philiastides, Auksztulewicz, Heekeren, & Blankenburg, 2011). In their detection study, Maniscalco & Lau (2015) measured the metacognitive certainty participants had about their detection responses. Trials that strongly involved manipulation of WM content were characterized by a decrease in metacognition. Not surprisingly, metacognition also is associated with activation in prefrontal areas (Fleming, Huijgen, & Dolan, 2012). Taken together, it cannot be determined whether increased demands on executive WM truly affect the threshold for visual awareness or merely influence subsequent stages such as deciding on a response or assessing the metacognitive quality of the visual awareness. Ultimately, the answer will remain uncertain until a control experiment can be developed that only differs from the masking technique in its relation to conscious awareness and not to any other decisional process (Sterzer, Stein, Ludwig, Rothkirch, & Hesselmann, 2014).
Third, an increase in visuospatial WM load did not influence detection times in the b-CFS trials or the control trials. Analysis of the performance on the secondary visuospatial WM task (higher error rates and RTs in the high load condition) indicate that the absence of an effect on detection times is not caused by a failure of the visuospatial load manipulation or a floor effect of the difficulty in this WM task. Interestingly, this difference between visuospatial load and executive load on detection time is reminiscent to that found in visual search. When comparing a single task and a dual task that loaded WM, applying an executive WM task severely impairs visual search performance (Han & Kim, 2004). By contrast, while visual search efficiency was hindered when loading the spatial subsystem of visuospatial WM, it remained nearly identical when loading the object subsystem of visuospatial WM (Woodman & Luck, 2004; Woodman, Vogel, & Luck, 2001). These results may mirror the current findings as our visuospatial WM test mainly queried the orientation of the target rectangles (object subsystem) and not their location, causing the spatial subsystem to be only minimally involved.
The lack of interference between visual awareness and increasing visuospatial WM load could appear to be inconsistent with the observation that visual awareness and maintaining content in visuospatial WM both involve a reverberant loop in a frontoparietal network (D’Esposito, 2007; Dehaene et al., 2006; Naghavi & Nyberg, 2005). A possible answer may come from a study by Rissman, Gazzaley, and D’Esposito (2008) in which the communication between the prefrontal cortex and posterior visual association areas was probed under changing visual WM load. While low load conditions mainly tapped into a frontoparietal network, high load conditions received support from the hippocampus during the retention period. Accordingly, high demands on the maintenance of visuospatial information might strategically call on hippocampal areas and free up resources for other tasks involving frontoparietal communication. It remains to be investigated whether a similar process is at play here.
References
Baars, B. (1993). A cognitive theory of consciousness. Cambridge: Cambridge University Press.
Baddeley, A. (2000). The episodic buffer: A new component of working memory? Trends in Cognitive Sciences, 4(11), 417–423. doi:10.1016/S1364-6613(00)01538-2
Brainard, D. H. (1997). The psychophysics toolbox. Spatial Vision, 10(4), 433–436. doi:10.1163/156856897X00357
Clark, A. (2013). Whatever next? Predictive brains, situated agents, and the future of cognitive science. The Behavioral and Brain Sciences, 36(3), 181–204. doi:10.1017/S0140525X12000477
Cohen, J. D., Perlstein, W. M., Braver, T. S., Nystrom, L. E., Noll, D. C., Jonides, J., & Smith, E. E. (1997). Temporal dynamics of brain activation during a working memory task. Nature, 386(6625), 604–608. doi:10.1038/386604a0
Cowan, N. (2001). The magical number 4 in short-term memory: A reconsideration of mental storage capacity. Behavioral and Brain Sciences, 24(1), 87–114. doi:10.1017/S0140525X01003922
Crick, F., & Koch, C. (1990). Towards a neurobiological theory of consciousness. In Seminars in the neurosciences (pp. 263–275). Saunders Scientific Publications.
D’Esposito, M. (2007). From cognitive to neural models of working memory. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 362(1481), 761–772. doi:10.1098/rstb.2007.2086
D’Esposito, M., Postle, B. R., Ballard, D., & Lease, J. (1999). Maintenance versus manipulation of information held in working memory: An event-related fMRI study. Brain and Cognition, 41(1), 66–86. doi:10.1006/brcg.1999.1096
De Loof, E., Verguts, T., Fias, W., & Van Opstal, F. (2013). Opposite effects of working memory on subjective visibility and priming. Journal of Experimental Psychology. Learning, Memory, and Cognition, 39(6), 1959–1965. doi:10.1037/a0033093
Dehaene, S., Changeux, J.-P., Naccache, L., Sackur, J., & Sergent, C. (2006). Conscious, preconscious, and subliminal processing: A testable taxonomy. Trends in Cognitive Sciences, 10(5), 204–211. doi:10.1016/j.tics.2006.03.007
Dehaene, S., & Naccache, L. (2001). Towards a cognitive neuroscience of consciousness: Basic evidence and a workspace framework. Cognition, 79(1-2), 1–37. doi:10.1016/S0010-0277(00)00123-2
Ebbinghaus, H. (1885). Memory: A contribution to experimental psychology. New York: Dover.
Fleming, S. M., Huijgen, J., & Dolan, R. J. (2012). Prefrontal contributions to metacognition in perceptual decision making. The Journal of Neuroscience : The Official Journal of the Society for Neuroscience, 32(18), 6117–6125. doi:10.1523/JNEUROSCI.6489-11.2012
Gayet, S., Paffen, C. L. E., & Van der Stigchel, S. (2013). Information matching the content of visual working memory is prioritized for conscious access. Psychological Science, 24(12), 2472–2480. doi:10.1177/0956797613495882
Han, S.-H., & Kim, M.-S. (2004). Visual search does not remain efficient when executive working memory is working. Psychological Science, 15(9), 623–628. doi:10.1111/j.0956-7976.2004.00730.x
Jiang, Y., Costello, P., & He, S. (2007). Processing of invisible stimuli: Advantage of upright faces and recognizable words in overcoming interocular suppression. Psychological Science, 18(4), 349–355. doi:10.1111/j.1467-9280.2007.01902.x
Konstantinou, N., & Lavie, N. (2013). Dissociable roles of different types of working memory load in visual detection. Journal of Experimental Psychology. Human Perception and Performance, 39(4), 919–924. doi:10.1037/a0033037
Lavie, N. (2005). Distracted and confused?: Selective attention under load. Trends in Cognitive Sciences, 9(2), 75–82. doi:10.1016/j.tics.2004.12.004
Maniscalco, B., & Lau, H. (2015). Manipulation of working memory contents selectively impairs metacognitive sensitivity in a concurrent visual discrimination task. Neuroscience of Consciousness, 1–13. doi:10.1093/nc/niv002
Minear, M., & Park, D. C. (2004). A lifespan database of adult facial stimuli. Behavior Research Methods, Instruments, & Computers, 36(4), 630–633. doi:10.3758/BF03206543
Naghavi, H. R., & Nyberg, L. (2005). Common fronto-parietal activity in attention, memory, and consciousness: Shared demands on integration? Consciousness and Cognition, 14(2), 390–425. doi:10.1016/j.concog.2004.10.003
Pan, Y., Lin, B., Zhao, Y., & Soto, D. (2014). Working memory biasing of visual perception without awareness. Attention, Perception & Psychophysics, 76(7), 2051–2062. doi:10.3758/s13414-013-0566-2
Pelli, D. G. (1997). The videotoolbox software for visual psychophysics: Transforming numbers into movies. Spatial Vision, 10(4), 437–442. doi:10.1163/156856897X00366
Philiastides, M. G., Auksztulewicz, R., Heekeren, H. R., & Blankenburg, F. (2011). Causal role of dorsolateral prefrontal cortex in human perceptual decision making. Current Biology, 21(11), 980–983. doi:10.1016/j.cub.2011.04.034
Rissman, J., Gazzaley, A., & D’Esposito, M. (2008). Dynamic adjustments in prefrontal, hippocampal, and inferior temporal interactions with increasing visual working memory load. Cerebral Cortex (New York, N.Y, 18(7), 1618–1629. doi:10.1093/cercor/bhm195
Ruchkin, D. S., Grafman, J., Cameron, K., & Berndt, R. S. (2003). Working memory retention systems: A state of activated long-term memory. Behavioral and Brain Sciences, 26(06), 709–728. doi:10.1017/S0140525X03000165
Sergent, C., Baillet, S., & Dehaene, S. (2005). Timing of the brain events underlying access to consciousness during the attentional blink. Nature Neuroscience, 8(10), 1391–1400. doi:10.1038/nn1549
Sheremata, S. L., Bettencourt, K. C., & Somers, D. C. (2010). Hemispheric asymmetry in visuotopic posterior parietal cortex emerges with visual short-term memory load. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience, 30(38), 12581–12588. doi:10.1523/JNEUROSCI.2689-10.2010
Sterzer, P., Stein, T., Ludwig, K., Rothkirch, M., & Hesselmann, G. (2014). Neural processing of visual information under interocular suppression: A critical review. Frontiers in Psychology, 5, 1–12. doi:10.3389/fpsyg.2014.00453
Tsuchiya, N., & Koch, C. (2005). Continuous flash suppression reduces negative afterimages. Nature Neuroscience, 8(8), 1096–1101. doi:10.1038/nn1500
Woodman, G. F., & Luck, S. J. (2004). Visual search is slowed when visuospatial working memory is occupied. Psychonomic Bulletin & Review, 11(2), 269–274. doi:10.3758/BF03196569
Woodman, G. F., Vogel, E. K., & Luck, S. J. (2001). Visual search remains efficient when visual working memory is full. Psychological Science, 12(3), 219–224. doi:10.1111/1467-9280.00339
Yuval-Greenberg, S., & Heeger, D. J. (2013). Continuous flash suppression modulates cortical activity in early visual cortex. The Journal of Neuroscience : The Official Journal of the Society for Neuroscience, 33(23), 9635–9643. doi:10.1523/JNEUROSCI.4612-12.2013
Zeki, S., & Bartels, A. (1999). Toward a theory of visual consciousness. Consciousness and Cognition, 8(2), 225–259. doi:10.1006/ccog.1999.0390
Acknowledgments
This work was partially funded by ERC Advanced Grant RADICAL to Axel Cleeremans. Axel Cleeremans is a Research Director of the F.R.S.-FNRS (Belgium).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
De Loof, E., Poppe, L., Cleeremans, A. et al. Different effects of executive and visuospatial working memory on visual consciousness. Atten Percept Psychophys 77, 2523–2528 (2015). https://doi.org/10.3758/s13414-015-1000-8
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-015-1000-8