
Abstract
Perceptual learning produces an improvement in visual functions such as an increase in visual acuity (VA) and contrast sensitivity in participants with both amblyopia and refractive defects. This improvement has been observed in the presence of lateral masking, which is known to bring about lateral interactions between detectors in early cortical pathways. Improvement has also been revealed in the absence of flankers in healthy individuals and those with amblyopia. This study seeks to understand whether a perceptual training regime really needs to be based on lateral interactions in cases where poor vision is not due to cortical dysfunction, such as myopia. Ten participants with mild myopia (max –2D) were recruited. A battery of tests measuring visual function was administered prior to (pre-test) and following (post-test) the training. The participants carried out an 8-week behavioural training using a single Gabor perceptual learning paradigm, completing a total of 24 sessions. Results indicate that training using a single Gabor protocol results in a VA improvement of 0.16 logMAR. The present study supports the idea that, in the absence of cortical deficits, as is the case in myopia, some sort of compensatory mechanism can take place at the cortical level by means of perceptual learning, resulting in more effective processing of the received blurred input. However, regarding training based on lateral masking, here we found that improvement of visual functions was smaller and limited to VA. This might suggest that training based on lateral masking, which is able to modify the strength of facilitatory and inhibitory lateral interactions, could be more effective for optimal recovery of blurred vision.
Similar content being viewed by others

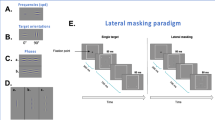
Introduction
Since studies in the early 1980s (e.g. Fiorentini & Berardi 1980), perceptual learning was observed in many visual tasks and was found to be highly specific to the trained stimulus characteristics or even to the trained eye (Ahissar & Hochstein 1996; Karni & Sagi 1991; Fiorentini & Berardi 1980; Campana & Casco 2003; Fahle & Poggio 2002), pointing to neural plasticity at early cortical stages, thus suggesting that sensory plasticity extends much beyond the critical period into adulthood (Sagi 2011). In fact, although some authors support the idea that learning occurs at the high-level decision stage (see, for example Yu et al. 2004), neuroimaging and electrophysiological results support the view that the striate cortical area V1 is often involved in perceptual learning and associated plastic changes (Casco et al. 2004; Gilbert et al. 2001; Pourtois et al. 2008; Schwartz et al. 2002). The mechanisms underlying perceptual learning could be a fine tuning (or selective weighting) of independent early detectors or channels (gain control) (Saarinen & Levi 1995; Schwabe & Obermayer 2005), a modification of interactions between detectors, either via horizontal (Polat & Sagi 1993, 1994a, 1994b, 2006; Tanaka & Sagi 1998) or feedback (Ahissar & Hochstein 1993, 1996, 1997) connections, or a reduction of external or internal noise (Huang et al. 2009; Lu & Dosher 2004) that could occur at either the sensory level (Bejjanki et al. 2011), or the decision stage (Yu et al., 2004).
Some perceptual tasks, however, have been reported to produce no improvement with practice, except in individuals showing high initial thresholds (Fahle & Henke-Fahle 1996) or when stimulus uncertainty is eliminated (Swift & Smith 1983), suggesting that some type of processing, probably occurring at low-sensory level, could be hard-wired and not modifiable, already showing the best possible performance (Sagi 2011). In the case of contrast detection or discrimination, Adini and colleagues (Adini et al. 2002, 2004) found that contrast discrimination of a Gabor stimulus can improve with practice only if it is flanked by pairs of similar Gabor stimuli. The mechanism underlying improvement of contrast detection with flankers has been ascribed to an increase of the range of facilitation between collinear elements resulting from a cascade of local connections between detectors based on Hebbian synaptic mechanisms (Polat 1999; Polat & Sagi 1994b). However, other authors found that improvement in contrast detection or discrimination can occur even in absence of flankers, provided that the range of employed contrasts is narrow (Yu et al. 2004).
Perceptual learning has been shown to be useful for improving visual functions such as visual acuity (VA) and contrast sensitivity (CS) in individuals with amblyopia (Chung et al. 2006; Polat et al. 2004; Huang et al. 2008) and also in those with refractive defects (Durrie & McMinn 2007; Tan & Fong 2008). It is also a promising technique for improving peripheral visual functions in patients with central visual loss (Maniglia et al. 2011).
In their 1994 studies, Polat and colleagues demonstrated that perceptual learning, using a lateral masking paradigm, results in an increase in the cortical spatial range of lateral interactions by a factor of six (Polat & Sagi, 1994a; 1994b). In a later study, Polat and colleagues (2004) trained adults with amblyopia using the same lateral masking technique and found that learning was generalised to higher level tasks such as letter recognition, and VA. Thus, by the end of the training the participants learned to see better. This and other studies using lateral-masking paradigms point to plasticity of spatial interactions in adults following repetitive training on a target-flanker task.
Different perceptual learning paradigms, however, have been implemented in adults with amblyopia, which resulted in effective improvement of CS or VA. For example, training on a contrast detection task using single Gabor patches (in the absence of flankers), either with (Huang et al. 2009) or without (Zhou et al. 2006) external noise. Zhou and his colleagues indicated that training improved VA and CS functions in amblyopic eyes that were retained for up to 1 year. Other studies have trained amblyopic participants on Vernier tasks, showing that repetitive practice not only leads to significant improvement in measured Vernier acuity but also a substantial improvement in (standard) VA and CS measurements, which, in some cases, reached up to normal vision. Despite the fact that these studies have shown that perceptual learning training on single targets (Gabor or Vernier stimuli) improved performance in the amblyopic eye, results obtained with the lateral masking paradigm seemed to be more efficient: the improvement in CS was 9.5 dB with the lateral masking paradigm, 4.9 dB with single Gabor training, and 3.5 dB with the Vernier task (Zhou et al. 2006).
Our ability to see contours and distinguish objects is also dependent upon the integrity of our eyes in effectively processing light, resulting in clear images. In myopia, the neuronal connectivity has developed normally in childhood and is capable of processing images efficiently; however, visual input is limited by an optical de-focus. In individuals with myopia, visibility of high spatial frequencies (SFs) is perceived as low contrast even when their physical contrast is high, thus degrading VA (Tan & Fong 2008). Despite the fact that perceptual or sensory training cannot modify the structure of the eye and the aforementioned ocular defects, positive results using perceptual learning paradigms have also been found using similar lateral masking techniques on refractive defects and thus individuals with myopia and presbyopia (Durrie & McMinn 2007; Tan & Fong 2008). How perceptual learning can improve vision in refractive disorders is still a matter of debate. The current explanation is that an increase in cortical processing efficiency can overcome the poor resolution of the image formed on the retina. The aim of this study is to investigate if an efficacious perceptual training, able to improve visual functions such as VA, really needs to be based on lateral interactions between detectors. It is still possible that cortical dysfunctions such as amblyopia, where connectivity between neurons is impaired, might obtain the most beneficial effects from a training based on lateral masking. Indeed, the amount of VA improvement found by Zhou and colleagues (2006), with no lateral masking, was nearly half that obtained by Polat and colleagues (2004) where lateral masking has been used.
However, in cases of poor vision not due to cortical dysfunctions, such as mild myopia, does a perceptual training regime really need to be based on lateral interactions? A contrast detection training with single Gabor patches was used on individuals with mild myopia to assess if it can have equally positive effects on CS and VA, similar to those reported using a lateral interaction paradigm. A battery of tests, including VA measured with Landolt C, Vernier acuity, CS and a test of lateral interactions using the lateral masking procedure, were administered in order to assess changes in visual functions before and after the training.
Methods
Participants
Ten participants with mild myopia were recruited from the University of Padova (mean age 24.22 years, range 22–27), all of which fitting the following inclusion criteria: refractive error up to –2 Diopter (D) in each eye (minimum was –0.75 D), with astigmatism not exceeding –0.5 D in either eye. The participants had a stable refractive index for the 6 months prior to training. Exclusion criteria included any other ocular condition or cause for reduced VA other than simple myopia and/or mild astigmatism; these included diabetes mellitus, pregnancy, presence of myopia-related ocular complications and any previous ocular surgery. To ensure the inclusion and exclusion criteria, prior to training every participant carried out a detailed assessment by a local optometrist.
This study was approved by the Local Ethics Committee. Informed consent was obtained from each participant prior to enrolment in the study highlighting that, at any point during the experiment, they were allowed to withdraw from the study. Two participants withdrew from the study following the complete training and post-training tests, thus they did not carry out the follow up 2 months post-training.
Experimental procedure
Each participant carried out a series of baseline visual functioning tests, which served as the pre-test measurements. These tests measured VA, Vernier hyperacuity, CS and lateral interactions. Following the pre-test, participants carried out an 8-week behavioural training using a single Gabor contrast detection task, completing a total of 24 sessions each lasting approximately 45 min. The same battery of tests was re-administered at the end of the treatment (post-test) and 2 months follow-up from the end of the treatment. All the pre- and post-tests as well as the training were carried out binocularly and without the use of corrective lenses. Furthermore, in order to investigate any possible effects of the intervention on optical eye characteristics, each participant also carried out an eye examination before and after the training at a local optometrist, who assessed pupil size, near point of accommodation and refractive error (Polat et al. 2012).
Apparatus
Stimuli were displayed on a 22-inch (55.88 cm) Philips Brilliance 202P4 monitor with a refresh rate of 60 Hz. Both the stimuli used in the training and in the lateral interaction test were generated with the Matlab Psychtoolbox (Brainard 1997; Pelli 1997), whereas stimuli for measuring VA, CS and Vernier acuity were generated using the Freiburg acuity and contrast test (FrACT 3.8) (Bach 1996). All stimuli were presented in foveal vision. The screen resolution was 1,280 x 1,024 pixels; each pixel subtended 0.33 arcmin at a viewing distance of 3 m, and 0.67 arcmin at a viewing distance of 1.5 m. Viewing distance was equal to 3 m for all tests except for the lateral interaction test and the training, which was administered from 1.5 m. Display linearisation was performed before the beginning of this study by means of a dedicated screen calibrator. Both the tests and training were carried out in a dark, silent room. Background screen luminance was 31.5 cd/m2 for all stimuli (FrACT contrast sensitivity test, training and lateral interaction stimuli) measured using a Gossen Mavo-Monitor luminance meter.
Freiburg acuity and contrast test
Stimuli used for measuring CS at the pre- and post-test measurements were sinusoidal gratings presented in a circular window with a narrow Gaussian taper. The size of the gratings used was 3° and grating orientations were 0, 45, 90 and 135°. The task of the participant entailed discriminating the orientation of the grating at different SFs, ranging from 1 cpd to 15 cpd, in separate blocks.
Landolt C optotypes were used to assess VA. The task of the participants was to indicate, in every trial, the orientation of the gap of the Landolt C out of eight possible orientations.
Vernier acuity was also assessed using two vertical lines, each 0.25° long, with no vertical separation between them, and with a variable horizontal offset. The task of the participants was to indicate, in every trial, the direction of the offset (left vs right) of the upper line with respect to the lower line.
For all FrACT stimuli, the Best-Pest adaptive procedure was used to calculate the absolute threshold for each of these tests. The thresholds were calculated by setting the discrimination rate in the middle between 100 % and the guessing rate (so if the guessing rate was equal to 25 % in the case of four choices, the threshold was set at 62.5 %) (Bach 1996). Note that these low-thresholds could overestimate the performance of participants (when the number of choices is larger than 2) respect to the more conservative threshold of 75 %.
Stimulus duration lasted until the participants’ response. An auditory cue was presented upon stimulus presentation and a different auditory cue was implemented as feedback for error responses.
Gabor stimuli (training and lateral interactions in pre/post-tests)
Stimuli used in the lateral interaction test and in the training comprised of Gabor patches consisting of a co-sinusoidal carrier enveloped by a stationary Gaussian. Standard deviation of the luminance Gaussian envelope (σ) was equal to the co-sinusoidal wavelength (λ); that is, the size of the Gabor patch covaried with its SF. Additionally, the spatial phase of the co-sinusoidal carrier equalled to zero (evenly symmetric Gabor patch). Stimulus duration lasted 200 ms.
In the lateral interaction test, two high-contrast Gabor patches (0.6 Michelson contrast), collinear to, and with the same SF of, a centrally presented low-contrast target Gabor, were located at various distances from the target (i.e. 2λ, 3λ, 4λ, and 8λ). Single SF, vertically oriented and collinear Gabor flankers (besides a condition with no flankers) were used. The SF used in the lateral masking task was the middle SF amongst the three SFs used in the training (see “training” section). The results of this test allowed us to investigate whether individuals with mild myopia who train on a single Gabor patch altered the strength of facilitatory (4λ) or inhibitory (2λ) collinear lateral interactions (Polat & Sagi 1993).
Training procedure
Following the baseline measurements, the participants undertook a series of training sessions using a single Gabor patch in a contrast detection task with a two interval forced choice (2IFC) procedure.
In a typical training task, the participant was presented with two consecutive displays where only one of the displays contained the target Gabor stimulus presented in the centre of the screen. Note that the same procedure was implemented in the lateral interaction test, with the exception that one interval contained both target and flankers, and one contained only the two flankers. Participants responded by pressing a key according to which interval he or she perceived the target; e.g. ‘press ‘z’ if the target was perceived in the first temporal interval and press ‘m’ if it was seen in the second temporal interval. The threshold corresponding to 79 % of correct discrimination was determined separately for each block by using a 1up/3down staircase procedure (Levitt 1971). In order to eliminate spatial or temporal uncertainty, and to avoid the possibility that practice improved performance by reducing uncertainty, both an auditory and a spatial cue were implemented. At the start of each block the participant was reminded to remain focused on the centre of the screen which was facilitated by providing a central fixation point (positional cue) preceding the presentation of each interval, as well as an auditory cue, indicating when the stimulus (if present) appeared. Performance feedback was also provided to the participants in the form of an auditory beep (500 Hz, 40 ms) following an incorrect response.
Each participant carried out three sessions per week for 8 weeks, comprising a total of 24 sessions. Within the same week and across sessions, the SF was varied; thus participants trained on only one SF per session and three SFs per week. The orientation of the Gabor Patches on the other hand was varied weekly (four orientations in 4 weeks). This protocol was then repeated for the 2nd month of training. In the first session of each week, participants trained with the lower SF (3 cpd) and were progressively presented with the higher ones (5 or 7 cpd for day 2, and 7 or 11 cpd for day 3). Therefore, each participant trained on three different SF, which changed daily, and four different orientations, which varied weekly (0°, 45°, 90° and 135°), thereby covering all stimulus orientations and three levels of SF. The three trained SFs were chosen individually for each participant on the basis of individual performance on the pre- training CS (“Grating”) task, choosing a lower SF range (i.e. 3, 5, 7 cpd) for participants with a CS value below 5 in the 15 cpd condition or below 10 in 11 cpd, or a higher SF range (i.e. 3, 7, 11 cpd) for participants with a CS above those values for the two highest tested SFs.
Each training session was made up of eight blocks, each block contained 60 trials, amounting to a total training time of 40 min per session. The battery of baseline tests were re-administered to each participant following 24 training sessions (8 weeks) in order to obtain post-training measurements of visual performance. Additionally, follow-up sessions were carried out two months following the end of the training in order to determine the long-term effectiveness of single Gabor techniques in improving visual functions.
Re-training
In case the training with single Gabor patches resulted in a VA improvement of less than 0.05 LogMAR, the participants were offered the chance to participate in a second training, which employed the lateral masking paradigm (Polat et al. 2004; Tan & Fong 2008; Durrie & McMinn 2007). This also allowed us to compare the results of training with single Gabors to lateral interaction training. Participants were trained with the same SF and orientation as in the first training, but for only 1 month (12 sessions). The same battery of tests was administered after this second training, in order to assess the presence of any further improvement.
Results
Training data
Since different participants were trained on different SFs, only those two SFs that were used by all participants (i.e., 3 cpd and 7 cpd) were analysed. Given the high variability of performance in each block within a session, and in order to reduce the weight of outliers or extreme data, the median contrast threshold of each session for each participant was calculated. A three-way ANOVA on these medians was carried out with SF (3 cpd vs 7 cpd), repetition (1st vs 2nd month), and orientation (0°, 45°, 90°, and 135°) as factors. Note that the only way to assess the presence of any learning effect on the present data is to compare the same conditions in different sessions. However, given the high number of conditions used in the training, the same conditions were repeated only twice: during the 1st month the combination of four orientations by three SF was administered across 12 sessions (3 sessions per week), and during the 2nd month the same conditions (in the same order as in the 1st month) were re-administered. This is why only the first vs. the second repetition of each condition can be compared. It is important also to point out that two participants missed at least one session, and because of these missing data the ANOVA considered only eight participants.
The main effect of SF was significant (F1,7 = 34.16, P < 0.001, η2 p = 0.83), indicating higher contrast thresholds with 7 cpd (0.287) with respect to 3 cpd (0.061). The main effect of orientation was marginally significant (F3,21 = 2.51, P = 0.086, η2 p = 0.26), but neither contrasts nor Bonferroni-corrected post-hoc t-tests revealed any significant difference between contrast thresholds with different orientations (all P > 0.05). The main effect of repetition (learning) was not significant (F1,7 = 3.13, P > 0.05, η2 p = 0.31), indicating that overall there was no learning improvement of performance between the 1st and the 2nd month. The interaction SF by repetition was marginally significant (F1,7 = 5.3, P = 0.055, η2 p = 0.43). Post-hoc t-tests showed no effect of repetition with SF set at 3 cpd (t7 = 0.87, P > 0.05), but a marginal effect of repetition with SF set at 7 cpd (t7 = 2, P = 0.086). Indeed, mean contrast thresholds at 7 cpd decreased from 0.3 in the first month, to 0.27 in the second month. Altogether, these data suggest that some learning occurred only at higher SFs, probably due to a floor effect at low SFs.
Pre- and post-tests
Each pre/post-test was analysed independently in order to identify whether any improvement has been made following the training.
VA tests results (Landolt-C) were assessed by using a one-way ANOVA with time (pre-, post-test and follow-up measurements) as main factor, followed by simple contrasts and Bonferroni-corrected post-hoc t-tests. The results revealed a significant main effect of time (F2,14 = 4.72, P < 0.05, η2 p = 0.4). Simple contrast showed a significant VA improvement (see Fig. 1) from pre- to post-test values (F1,7 = 7.95, P < 0.05, η2 p = 0.53) but no significant differences from pre- to follow-up test values (F1,7 = 3.64, P > 0.05, η2 p = 0.34). Bonferroni-corrected post-hoc t-tests confirmed the same picture: a marginally significant VA improvement from pre- to post-test values (t9 = 2.6, P = 0.057) and no significant difference from pre- to follow-up (t7 = 1.91, P = 0.19). The improvement of 0.16 LogMAR from pre- (0.427 LogMAR) to post-tests (0.267 LogMAR) decreased by only 10 % at follow-up tests (0.283 LogMAR) despite yielding a statistically non-significant result. This may be due to a reduced sample size in the analysis since two participants dropped out following the post-tests and did not carry out the follow up assessment.

Mean visual acuity (VA) measured with Landolt C before the training (pre-test), after the training (post-test) and at 2 months from the end of the training (follow-up). Error bars Standard error of the mean (SE)
Concerning Vernier hyperacuity, despite average pre-training hyperacuity was reported as 92.78 arcsecs and post-training hyperacuity at 61.85 arcsecs, a paired-samples t-test did not reveal any significant difference (t9 = 1.85, P > 0.05).
Log-transformed CS results (Fig. 2) were assessed using a two-way ANOVA with time (pre- post- measurements) and SF (1, 3, 5, 7, 9, 11, 15 cpd) as main factors. No significant main effect of time (F1,8 = 0.42, P > 0.05, η2 p = 0.05), nor time x SF interaction (F6,48 = 1.1, P > 0.05, η2 p = 0.12) was found.

Contrast sensitivity (CS) function (geometric mean) measured before (pre-test) and after the training (post-test). Error bars SE
Lateral interactions tests (Fig. 3), assessed using a two-way ANOVA with time (pre-, post-test measurements) and target-to-flankers distance (2, 3, 4 and 8λ) as main factors, revealed no statistically significant results of time (F1,9 = 0.09, P > 0.05, η2 p = 0.01), target-to-flankers distance (F3,27 = 2.56, P > 0.05, η2 p = 0.22), or interaction (F3,27 = 2.5, P > 0.05, η2 p = 0.22). In order to analyse the effects of single Gabor training on the facilitatory and inhibitory components of lateral interactions, the contrast thresholds obtained with each target-to-flanker distance were normalised by subtracting the contrast thresholds of a single-Gabor block (with the same SF and same orientation) obtained before and after training (Fig. 3). A two-way ANOVA (pre- vs post-test, target-to-flanker distance) revealed no significant differences between pre- and post-test results (F1,9 = 0.22, P > 0.05, η2 p = 0.023), a marginally significant difference between the various target-to-flanker distances (F3,27 = 2.56, P = 0.076, η2 p = 0.22), and a marginally significant interaction (F3,27 = 2.5, P = 0.081, η2 p = 0.22), probably due to the different mean thresholds at 2 and 8λ (see Fig. 3). However, post-hoc t-tests revealed no significant differences between these normalised pre- and post-measures at any target-to-flanker distance (all P > 0.05), nor any significant differences between normalised pre- or post- measures and zero (all P > 0.05), further supporting the idea of altered lateral interactions in myopia.

Mean normalised contrast thresholds (threshold differences respect to baseline thresholds without flankers) in the lateral masking paradigm as a function of target-flankers distance (λ), before (pre-test) and after the training (post-test). Positive values indicate inhibitory effects of the flankers, negative values facilitatory effects. Error bars SE
In order to assess whether training modified the optical characteristics of the eye, measurement of pupil size (in scotopic conditions), refractive error and near point of accommodation (in mesopic conditions) were taken for each participant. Mean pupil size before and after training was, respectively, 4.33 mm and 4.35 mm for the right eye, and 4.32 mm and 4.37 mm for the left eye. A two-way ANOVA with time (pre- vs post) and eye as factors showed no significant differences between pupil size before and after training (F1,9 = 0.17, P > 0.05, η2 p = 0.019). Mean near point of accommodation before and after training was, respectively, 10.6 mm and 10.7 mm for the right eye, and 11.1 mm and 11.3 mm for the left eye. A two-way ANOVA with time (pre- vs post) and eye as factors showed no significant differences between the near point of accommodation before and after training (F1,9 = 3.8, P > 0.05, η2 p = 0.3). Mean refractive error before and after training was, respectively, 1.30 D and 1.35 D for the right eye, and 1.35 D and 1.35 D for the left eye. A two-way ANOVA with time (pre- vs post) and eye as factors showed no significant differences between the refractive error before and after training (F1,9 = 0.12, P > 0.05, η2 p = 0.01).
Control group
In order to exclude the possibility that the improvement in VA was only due to the repetition of the same VA (and/or CS) tests at pre- and post-tests, we collected a small sample of six participants with the same criteria as the experimental group. These participants carried out the same VA and CS pre- and post-tests (Landolt C and Gratings of the FrACT, respectively), with a 1-week interval between them. The results showed no significant difference in VA between the pre- (0.31 LogMAR) and post-test (0.34 LogMAR) (t5 = –0.64, P > 0.05), and no significant difference between pre- and post-test in Log-transformed CS (F1,5 = 1.09, P > 0.05, η2 p = 0.17) for any of the tested SF (F6,30 = 1.25, P > 0.05, η2 p = 0.2). No participant improved VA by more than 0.05 LogMAR between the first and the second test.
Re-training
Three participants who obtained a VA improvement of less than 0.05 LogMAR agreed to participate in the 1-month re-training with lateral masking stimuli.
After retraining, participants showed an additional improvement in their VA ranging from 0.1 to 0.28 LogMAR (Fig. 4), and a consistent improvement in their CS at most of the tested SFs (improvement ranging from 10 % to above 100 %, Fig. 5). No further improvement in Vernier acuity was found in any participant.

Individual VA of three participants before and after re-training with lateral masking

Individual CS of three participants before (black symbols) and after (grey symbols) re-training with lateral masking. Symbols connected by lines represent the geometric means of the three subjects
Discussion
The present study investigated the effects on VA and CS outcomes of single Gabor training in the absence of lateral masking in a group of individuals with mild myopia (maximum –2D). The lack of significant differences between the pre- and post-tests of the control group suggest that any improvements seen in the experimental group following the training were not due to any learning effects of the baseline visual tasks.
The results indicate that training using a single Gabor protocol (training on three SF and four orientations) results in an improvement on VA of 0.16 LogMAR (on average) following 24 training sessions, with one participant out of ten exhibiting a slightly worsened VA. This degree of improvement, although both statistically significant and clinically relevant, is not as strong in magnitude as that found in training protocols using lateral masking, which is reported as being 0.22 LogMAR in various groups of visual difficulties, namely, myopia, presbyopia and amblyopia (Durrie & McMinn 2007; Polat et al. 2004, 2012; Tan & Fong 2008). Nevertheless, and despite high inter-participant variability in VA improvement, the single Gabor training did improve VA up to 0.44 LogMAR. Although in principle, such improvement in VA could also be due to a more efficacious attentional focusing, the presence of an auditory cue both in the training task and in the VA task (besides the fact that in the VA task the stimulus was shown with no time limits) should have already produced an optimal attentional focusing, with not much room for further enhancement. Additionally, the specific improvement of VA performance (whereas CS and vernier hyperacuity did not improve) does not support the idea of high-level learning of rules for performing a visual discrimination task (Zhang et al. 2014).
Surprisingly, our findings relating to CS and Vernier hyperacuity are not as suggestive as those found for VA improvement, pointing to a limited effect on visual cortical processing of single Gabor training using the present protocol. No statistically significant effects of single Gabor training were found on improvement of CS. One reason for this may be the difference in size of the gratings used in the training and in the pre/post-tests measuring CS. Since training was carried out using small gratings while pre- and post- CS tests used larger gratings, it is possible that no transfer of the training on CS took place. However, a number of studies involving training on contrast detection tasks have found transfer of CS improvement using a variety of different CSF measurement tools and sizes of stimuli. For example, Maniglia and colleagues found an improvement limited to high spatial frequencies using computer-generated stimuli of different sizes with respect to the trained ones. Durrie & McMinn (2007), Tan and Fong (2008) and Polat et al. (2009), found CS improvement used wall-mounted sinewave contrast sensitivity charts with stimuli with constant size, whereas the size of trained stimuli was dependent on spatial frequency, and Yalcin and Balci (2013) found an improvement using a similar procedure but implemented in an automatic slide package tool (“Optec functional vision analyser”).
Other explanations for the absence of improvement found in contrast sensitivity could reside in the different experimental protocols used in perceptual learning research. For example, studies showing greatest improvement in both VA and CS, trained participants using a range of SFs under lateral masking conditions. In each training session, the SF was varied; shifting progressively higher, and four orientations were presented at each SF (Polat et al. 2004, 2012; Polat 2009). Polat and his colleagues attribute this improvement to enhanced neuronal lateral interactions, as can also be witnessed through increased facilitation from nearby flankers. Interestingly, however, Zhou and colleagues in 2006 also show improvement on the trained task as well as transfer onto VA and CS, after practicing contrast detection without the use of flankers. This group of researchers trained two groups of participants, one of which were trained using one SF known as the cut-off SF (defined as the SF at which the estimated contrast threshold from pre-training CS measurements was 0.50), and the other group trained on a variety of SFs. The results indicated that the greatest improvement on VA and CS (in both the trained and fellow eye) was seen in the group that trained on only one SF at the cut-off performance of each participant. Whereas in the group training on a variety of SFs no significant effect was found on the improvement on CS and VA in the fellow eyes and the overall improvement in the trained eye was much weaker compared to the first group training on cut-off (Zhou et al. 2006). Thus, in cases where cortical processing is non-optimal (as in the case of amblyopia), it is unclear whether improvement in VA and CS is based primarily on the improvement of lateral interactions or rather to over-learning of specific stimulus attributes.
Thus, another explanation for the disparity of results in contrast sensitivity found with single Gabor versus lateral masking training could reside in the variations of training protocols and stimulus characteristics, not just the absence vs presence of flankers. Several studies have shown that neural noise in the early visual system can be modulated through the appropriate choice of stimulus parameters and training conditions (e.g. Durrie & McMinn 2007; Polat et al. 2012). In all psychophysical studies, the importance of finding the right stimulus parameters for signal detection is well reported (Huang et al. 2008; Zhou et al. 2006). Improving the signal-to-noise ratio of cortical neurons in the primary visual cortex is a mechanism frequently proposed to account for the improvement in contrast sensitivity following perceptual learning (Polat et al. 2012; Polat 2009; Webster et al. 2002). Another explanation is that the implemented stimulus parameters (e.g. choice of SF) were not suitable or sensitive enough to bring about this neuronal plasticity and, consequently, improvements in contrast sensitivity. In his recent review paper, Sagi (2011) mentions that perceptual learning is seen on almost every occasion where the observer’s vision is challenged. Huang and colleagues also point out that lower SFs are not as effective in producing a positive effect on VA and CS when compared to higher or cut-off SFs (Huang et al. 2008; Zhou et al. 2006). The present study supports these findings in a group of participants with non-cortical visual difficulties. The choice of three SFs, of which likely just one was at cut-off, may have resulted in a limited effect of the training on cortical processing. Many perceptual learning paradigms showing an improvement in both VA and CS make use of the NeuroVision Technology system (Durrie & McMinn 2007; Tan & Fong 2008; Polat et al. 2012), which provides on-going subject-specific algorithms adapted to each participant. This results in an individualised treatment, implementing subject-specific stimuli thar change each training session according to the participants’ on-going performance during the training (Polat et al. 2012; Tan & Fong 2008). Although the stimuli chosen in this study were selected carefully according to each individual’s performance on pre-training contrast sensitivity, the SFs chosen were not altered according to their on-going training performance.
Another difference between the present study and lateral masking studies concerns the duration of training. Finding the gold standard when choosing the number of training sessions required to bring about a functional improvement in vision is also necessary. Whereas in the present study the duration was fixed and equal to 24 sessions, in other studies the training duration was tailored on the performance of participants and was, on average, slightly longer. For example, in the study of Tan and Fong, an improvement of 0.21 logMAR was found over a training period between 20 and 30 sessions over 3 months. Another study using lateral masking in myopia also found an improvement of 0.22 logMAR following 30 training sessions (Durrie & McMinn 2007). An improvement of 0.2 logMAR in VA and in CS of 0.26 log units (average of 95 %, where the peak improvement was 164 % at 12 cpd), was also achieved in a study on presbyopia using lateral masking over 37.4 (±10.7) training sessions (Polat et al. 2012). In the present study, with a group of mild myopia participants, no significant improvement on contrast sensitivity was found after 24 training sessions and no strong learning trend was observed over the course of 24 sessions (8 weeks). A possible explanation for the absence of learning (or only slight learning) could be the presence vs absence of high-order optical aberrations: Zhou and colleagues (Zhou et al. 2012) found that perceptual learning was much larger when correcting for high-order optical aberrations that might strongly limit the quality of the retinal image. In fact, since in our study training was performed without optical correction, both low-order and high-order optical aberrations could have contributed to decrease the optical quality and thus interfere with perceptual learning and its transfer to VA and CS. Note, however, that this very small improvement in the trained task may also be due to the lack of comparable conditions (since on each day a different condition was trained). This variability may also account for the lack of improvement seen in CS, raising the importance of selecting the right stimulus parameters for contrast detection training. Nonetheless, in their mini review study comparing different perceptual learning paradigms as a potential treatment for amblyopia, Levi and Li (2009) point out that not all contrast detection training experiments lead to large changes in the trained task. Studies showed that neither identification of near threshold contrast defined letters nor tasks of luminance defined letters resulted in substantial learning of the trained task (Chung et al. 2006, 2008; Levi 2005).
Although explanations such as choice of stimulus parameters and duration of training cannot be ignored, the data obtained on re-training with the lateral masking technique on participants that did not improve with the single Gabor training still suggests stronger reliability of lateral masking as a training for improving visual functions, not just in participants with cortical dysfunctions such amblyopia, but also in participants with refractive defects. Indeed, the participants who achieved only small improvements on VA with 24 sessions of training with single Gabors, seemed to have improved both in VA and CS with an additional 12 sessions of training using a lateral masking paradigm (using the same stimulus conditions). The VA and CS improvement of these three participants upon re-training with the lateral masking paradigm might suggest that optimal tuning of visual cortical processing able to overcome blurred images due to mild refractive defects, possibly requires the strengthening of facilitatory and inhibitory lateral interactions between collinear detectors brought about through lateral masking. The absence of significant differences between contrast thresholds at various target-to-flanker distances, as well as their normalised contrast thresholds not significantly different from zero, suggest that these (uncorrected) myopic participants might have altered lateral interactions between collinear detectors.
The distinct organic differences of different visual conditions calls into question whether rehabilitation of visual functioning on an organic and functional level follows the same processes and thus requires the same conditions of learning. In amblyopia, studies reveal improved visual functioning following perceptual learning both in the presence and absence of flankers, although larger improvements are often found with the lateral masking paradigm (Zhou et al. 2006; Polat et al. 2004; Huang et al. 2009; Zhang et al. 2014). The underlying mechanisms proposed are said to involve strengthening connections through Hebbian learning, resulting in recovery of function implicating lateral, feed-forward and feedback mechanisms (e.g. Rosa et al. 2013; Li & Levi 2004; Polat et al. 2004). Myopia, on the other hand, is a condition presenting with intact cortical functioning and difficulties at the ocular level. The uncorrected myopic neural system operates under sub-optimal conditions as it receives a blurred input resulting in lower contrast and resolution and reduced visual acuity. In myopia, thus far, no study has investigated whether training in the absence of flankers can improve visual acuity and contrast sensitivity. The present study suggests that when there is no cortical deficit, such as in refractive defects, some sort of compensatory mechanism can take place at the cortical level by means of perceptual learning, even in the absence of lateral masking, which results in more effective processing of the received blurred input, although contrast sensitivity does not seem to be affected. Training to detect low-contrast, small Gabor stimuli could have increased the ability of the visual system to detect small signals in noise (blurred image), thus transferring this ability to VA, with only small modifications of lateral interactions between detectors responding to oriented, collinear stimuli. In fact, when lateral masking paradigms have been used (Durrie & McMinn 2007; Tan & Fong 2008), a larger improvement on VA and CS was found, indicating that, not just in amblyopia, but also in refractive defects, a modification of the strength of both facilitatory and inhibitory lateral interactions via trainings based on lateral masking could be more effective for an optimal recovery of blurred vision.
Conclusion
This study investigated the effect of perceptual learning using single Gabor training on a group of participants with mild myopia. The results reveal a positive effect of the training on VA and less so on CS.
When applying psychophysical techniques to training of visual functions (i.e. clinical application), the main question arising is which training protocol and which stimulus parameters are therefore best suited to yield effective positive results for each individual participant, i.e. improved VA and CS. Additionally, considering the variation of visual defects (such as amblyopia, myopia, presbyopia), training protocols also need to be tailored to each condition.
The present research raises two main queries: (1) is there a gold standard training protocol for perceptual learning to be most efficient and effective to daily visual functioning? (2) If so, by which neuronal mechanism is this transfer made possible. The latter raises the importance of introducing electrophysiological and other brain imaging techniques in combination with a perceptual learning paradigm for visual defects in order to better understand the underlying neural mechanisms by which perceptual learning takes place under different conditions. Understanding these two questions is key in order to put into practice the current literature on psychophysics and perceptual learning as a rehabilitation tool for improving visual functions in a variety of visual defects.
References
Adini, Y., Sagi, D., & Tsodyks, M. (2002). Context-enabled learning in the human visual system. Nature, 415(6873), 790–793.
Adini, Y., Wilkonsky, A., Haspel, R., Tsodyks, M., & Sagi, D. (2004). Perceptual learning in contrast discrimination: The effect of contrast uncertainty. Journal of Vision, 4(12), 993–1005.
Ahissar, M., & Hochstein, S. (1993). Attentional control of early perceptual learning. Proceedings of the National Academy of Sciences of the United States of America, 90(12), 5718–5722.
Ahissar, M., & Hochstein, S. (1996). Learning pop-out detection: Specificities to stimulus characteristics. Vision Research, 36(21), 3487–3500.
Ahissar, M., & Hochstein, S. (1997). Task difficulty and the specificity of perceptual learning. Nature, 387(6631), 401–406.
Bach, M. (1996). The “Freiburg Visual Acuity Test”—automatic measurement of visual acuity. Optometry and Vision Science, 73, 49–53.
Bejjanki, V. R., Beck, J. M., Lu, Z. L., & Pouget, A. (2011). Perceptual learning as improved probabilistic inference in early sensory areas. Nature Neuroscience, 14(5), 642–648.
Brainard, D. H. (1997). The psychophysics toolbox. Spatial Vision, 10, 443–446.
Campana, G., & Casco, C. (2003). Learning in combined-feature search: Specificity to orientation. Perception & Psychophysics, 65(8), 1197–1207.
Casco, C., Campana, G., Grieco, A., & Fuggetta, G. (2004). Perceptual learning modulates electrophysiological and psychophysical response to visual texture segmentation in humans. Neuroscience Letters, 371(1), 18–23.
Chung, S. T., Li, R. W., & Levi, D. M. (2006). Identification of contrast-defined letters benefits from perceptual learning in adults with amblyopia. Vision Research, 46(22), 3853–3861.
Chung, S. T., Li, R. W., & Levi, D. M. (2008). Learning to identify near-threshold luminance-defined and contrast-defined letters in observers with Amblyopia. Vision Research, 48, 2739–2750.
Durrie, D., & McMinn, S. P. (2007). Computer-based primary visual cortex training for treatment of low myopia and early presbyopia. Transactions of the American Ophthalmological Society, 105, 132–140.
Fahle, M., & Henke-Fahle, S. (1996). Interobserver variance in perceptual performance and learning. Investigative Ophthalmology and Visual Science, 37(5), 869–877.
Fahle, M., & Poggio, T. (2002). Perceptual learning. Cambridge, Masssachusetts: MIT Press.
Fiorentini, A., & Berardi, N. (1980). Perceptual learning specific for orientation and spatial frequency. Nature, 287(5777), 43–44.
Gilbert, C. D., Sigman, M., & Crist, R. E. (2001). The neural basis of perceptual learning. Neuron, 31(5), 681–697.
Huang, C. B., Lu, Z. L., & Zhou, Y. (2009). Mechanisms underlying perceptual learning of contrast detection in adults with anisometropic amblyopia. Journal of Vision, 9(11), 1–14.
Huang, C. B., Zhou, Y., & Lu, Z. L. (2008). Broad bandwidth of perceptual learning in the visual system of adults with anisometropic amblyopia. Proceedings of the National Academy of Sciences of the United States of America, 105(10), 4068–4073.
Karni, A., & Sagi, D. (1991). Where practice makes perfect n texture discrimination: Evidence for primary visual cortex plasticity. Proceedings of the National Academy of Sciences of the United States of America, 88, 4966–4970.
Levi, D. M. (2005). Perceptual learning in adults with amblyopia: A re-evaluation of critical periods in human vision. Developmental Psychobiology, 46(3), 222–232.
Levi, D. M., & Li, R. W. (2009). Perceptual learning as a potential treatment for amblyopia. Vision Research, 21, 2535–2549.
Levitt, H. (1971). Transformed up-down methods in psychoacoustics. Journal of the Acoustical Society of America, 49(2), 467–477.
Li, R. W., & Levi, D. M. (2004). Characterizing the mechanisms of improvement for position discrimination in adult amblyopia. Journal of Vision, 4, 476–487.
Lu, Z. L., & Dosher, B. A. (2004). Spatial attention excludes external noise without changing the spatial frequency tuning of the perceptual template. Journal of Vision, 4(10), 955–966.
Maniglia, M., Pavan, A., Cuturi, L. F., Campana, G., Sato, G., & Casco, C. (2011). Reducing crowding by weakening inhibitory lateral interactions in the periphery with perceptual learning. PLoS One, 6(10), e25568.
Pelli, D. G. (1997). The VideoToolbox software for visual psychophysics: Transforming numbers into movies. Spatial Vision, 10, 437–442.
Polat, U. (1999). Functional architecture of long-range perceptual interactions. Spatial Vision, 12, 143–162.
Polat, U. (2009). Making perceptual learning practical to improve visual functions. Vision Research, 49, 2566–2573.
Polat, U., Ma-Naim, T., Belkin, M., & Sagi, D. (2004). Improving vision in adult amblyopia by perceptual learning. Proceedings of the National Academy of Sciences of the United States of America, 101(17), 6692–6697.
Polat, U., Ma-Naim, T., & Spierer, A. (2009). Treatment of children with amblyopia by perceptual learning. Vision Research, 49(21), 2599–2603.
Polat, U., & Sagi, D. (1993). Lateral interactions between spatial channels: Suppression and facilitation revealed by lateral masking experiments. Vision Research, 33(7), 993–999.
Polat, U., & Sagi, D. (1994a). The architecture of perceptual spatial interactions. Vision Research, 34(1), 73–78.
Polat, U., & Sagi, D. (1994b). Spatial interactions in human vision: From near to far via experience-dependant cascades of connections. Proceedings of the National Academy of Sciences of the United States of America, 91(4), 1206–1209.
Polat, U., & Sagi, D. (2006). Temporal asymmetry of collinear interactions. Vision Research, 46, 953–960.
Polat, U., Schor, C., Tong, J. L., Zomet, A., Lev, M., Yehezkel, O., ... Levi, D. M. (2012). Training the brain to overcome the effect of aging on the human eye. Scientific Reports, 2, 278.
Pourtois, G., Rauss, K. S., Vuilleumier, P., & Schwartz, S. (2008). Effects of perceptual learning on primary visual cortex activity in humans. Vision Research, 48(1), 55–62.
Rosa, A. M., Silva, M. F., Ferreira, S., Murta, J., & Castelo-Branco, M. (2013). Plasticity in the human visual cortex: An ophthalmology-based perspective. BioMedical Research International, ID 568354, 13.
Saarinen, J., & Levi, D. M. (1995). Perceptual learning in Vernier acuity: What is learned? Vision Research, 35(4), 519–527.
Sagi, D. (2011). Perceptual learning in Vision Research. Vision Research, 51, 1552–1566.
Schwabe, L., & Obermayer, K. (2005). Learning in top-down gain control of feature selectivity in a recurrent network model of a visual cortical area. Vision Research, 45(25–26), 3202–3209.
Schwartz, S., Maquet, P., & Frith, C. (2002). Neural correlates of perceptual learning: A functional MRI study of visual texture discrimination. Proceedings of the National Academy of Sciences of the United States of America, 99(26), 17137–17142.
Swift, D. J., & Smith, R. A. (1983). Spatial frequency masking and Weber’s law. Vision Research, 23(5), 495–505.
Tan, D. T., & Fong, A. (2008). Efficacy of neural vision therapy to enhance CS and visual acuity in low myopia. Journal of Cataract and Refractive Surgery, 34(4), 570–577.
Tanaka, Y., & Sagi, D. (1998). Long-lasting, long-range detection facilitation. Vision Research, 38, 2591–2599.
Webster, M. A., Georgeson, M. A., & Webster, S. M. (2002). Neural adjustments to image blur. Nature Neuroscience, 5, 839–840.
Yalcin, E., & Balci, O. (2013). Efficacy of perceptual vision therapy in enhancing visual acuity and contrast sensitivity function in adult hypermetropic anisometropic amblyopia. Clinical Ophthalmology, 8, 49–53.
Yu, C., Klein, S. A., & Levi, D. M. (2004). Perceptual learning in contrast discrimination and the (minimal) role of context. Journal of Vision, 4(3), 169–182.
Zhang, J. Y., Cong, L. J., Levi, D. M., Klein, S. A., & Yu, C. (2014). Perceptual learning improves adult amblyopic vision through rule-based cognitive compensation. Investigative Ophthalmology and Visual Science, 55(4), 2020–2030.
Zhou, Y., Huang, C., Xu, P., Tao, L., Qiu, Z., Li, X., & Lu, Z. L. (2006). Perceptual learning improves CS and VA in adults with anisometropic amblyopia. Vision Research, 46(5), 739–750.
Zhou, J., Zhang, Y., Dai, Y., Zhao, H., Li, R., Hou, F., ... Zhou, Z. (2012). The eye limits the brain's learning potential. Nature Scientific Reports, 364, 1–6.
Acknowledgements
We would like to thank Stefano Cappello (optician/optometrist, Padova, Italy) for the optometric assessment of the participants as well as Silvia Rigoni for helping out with data collection.
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
About this article

Cite this article
Camilleri, R., Pavan, A., Ghin, F. et al. Improving myopia via perceptual learning: is training with lateral masking the only (or the most) efficacious technique?. Atten Percept Psychophys 76, 2485–2494 (2014). https://doi.org/10.3758/s13414-014-0738-8
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-014-0738-8