
Tidal Flushing Rather Than Non-Point Source Nitrogen Pollution Drives Nutrient Dynamics in A Putatively Eutrophic Estuary
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
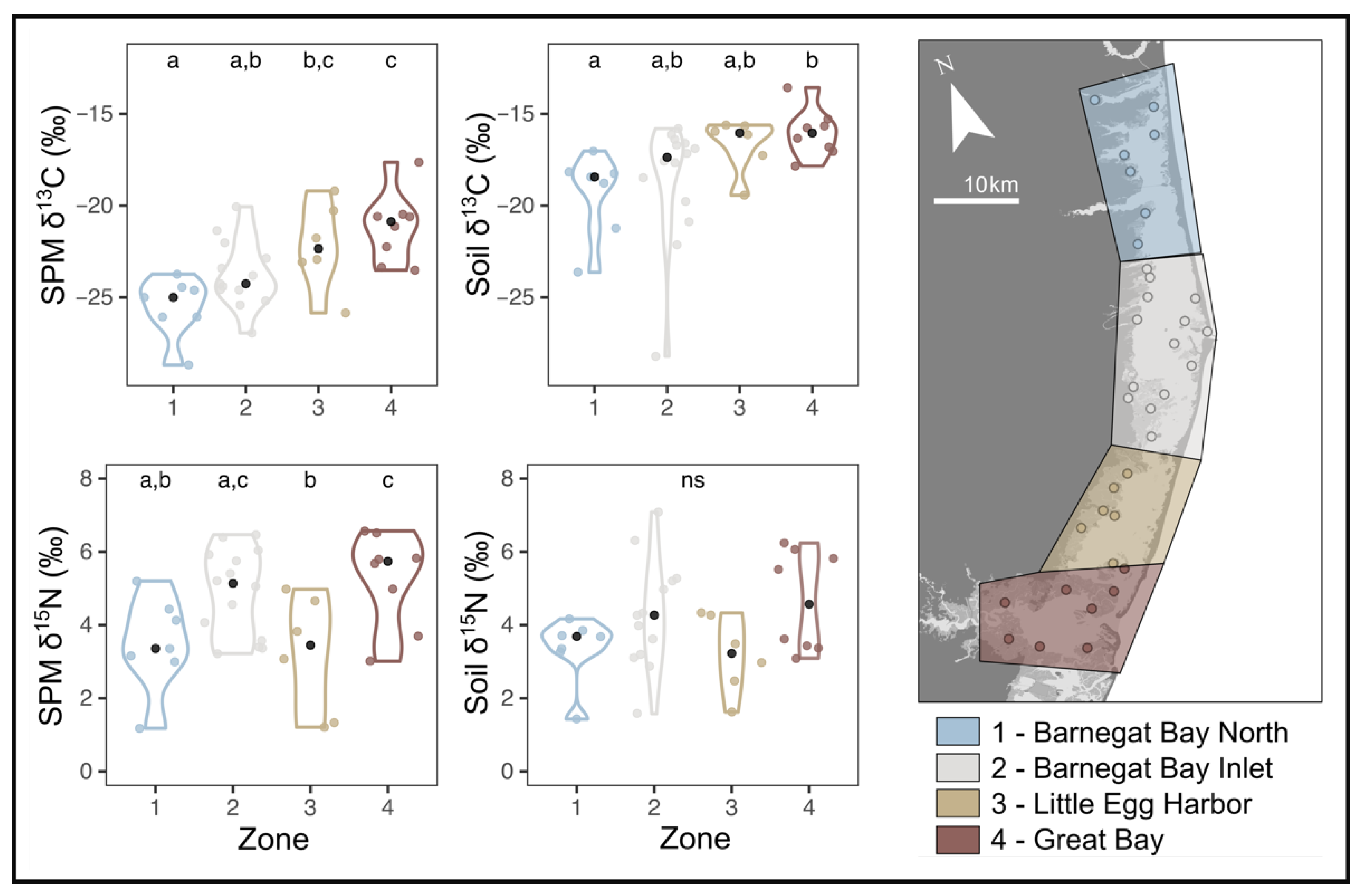
2.1. Study System
2.2. Sample Collection
2.3. Laboratory Analysis
2.3.1. Stable Isotopes
2.3.2. Dissolved Inorganic Nutrients
2.3.3. Salt Marsh Cores
2.4. Data Analysis
3. Results
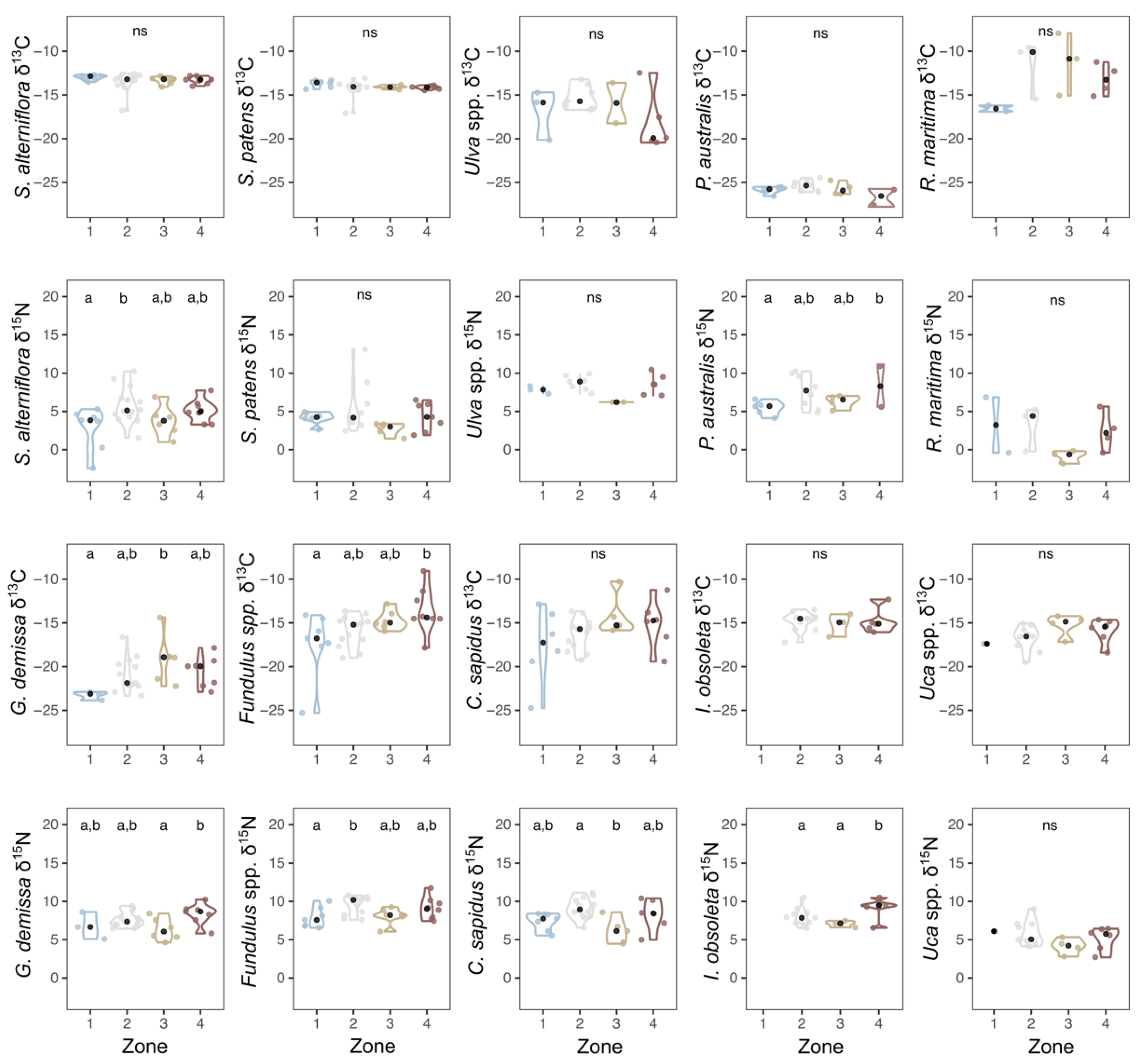
3.1. Modern Stable Isotopes
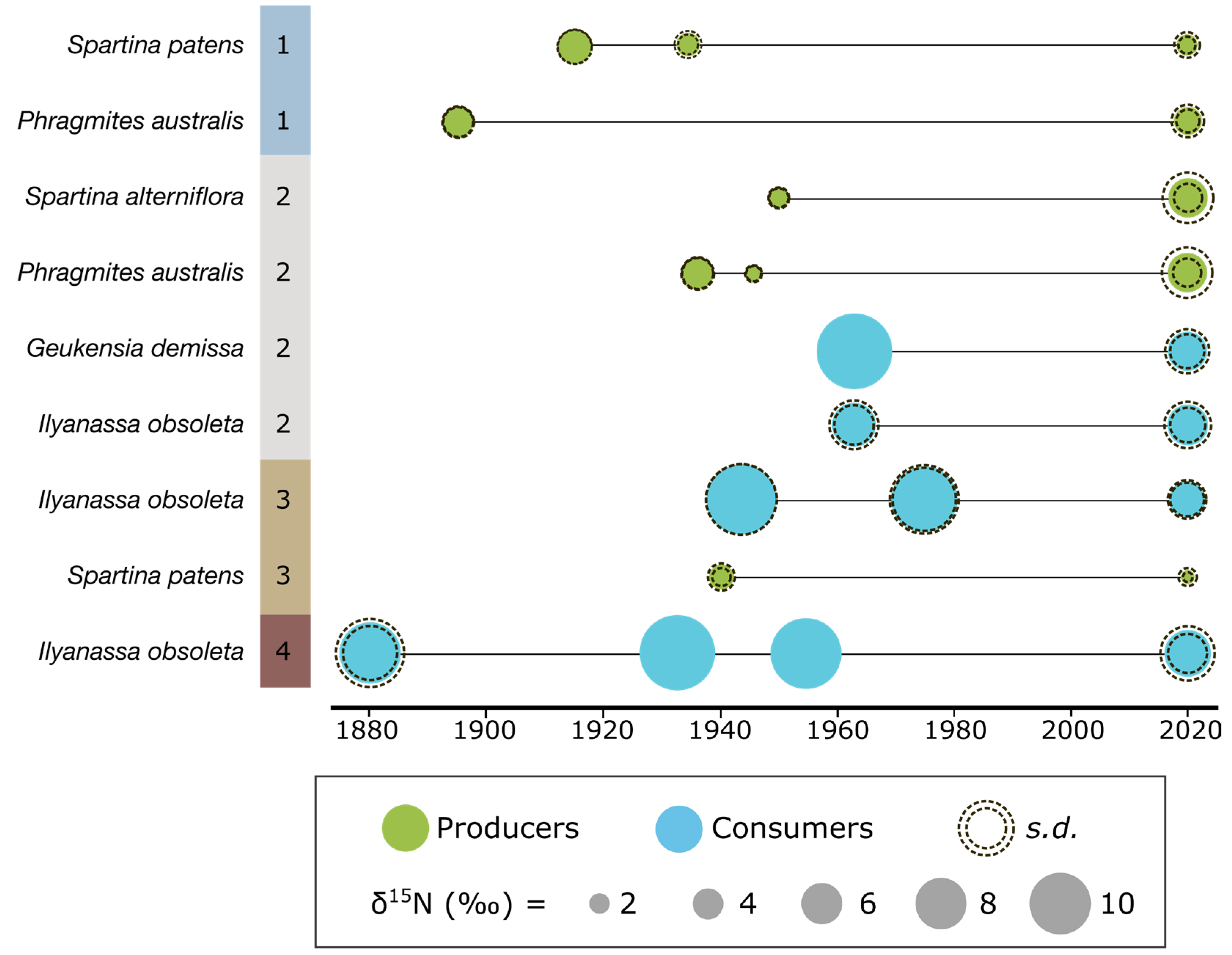
3.2. Archival and Modern Analogue Material
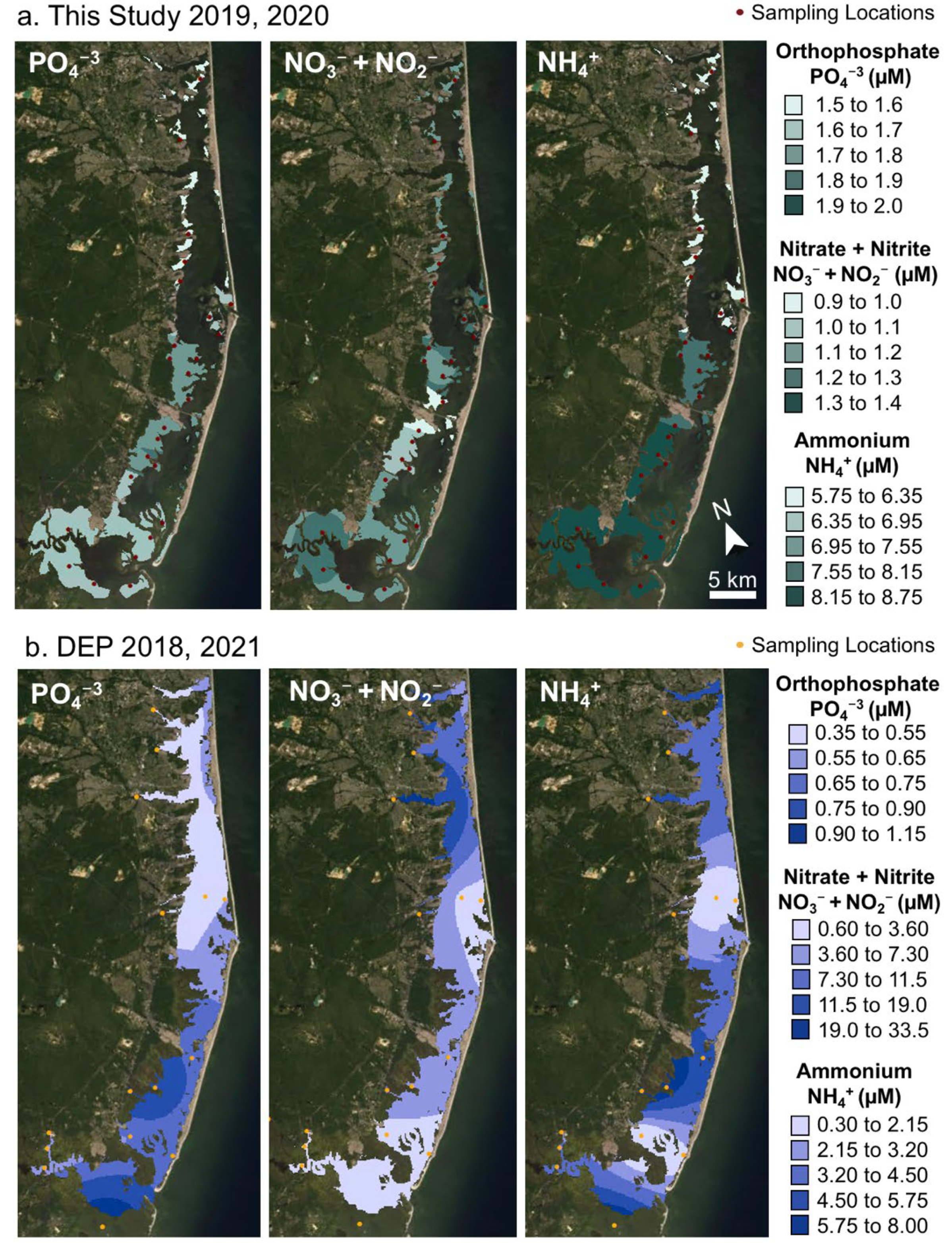
3.3. Water Quality and Salt Marsh Cores
4. Discussion
4.1. Spatial Patterns of Macronutrient Availability
4.2. Estuarine Isoscapes in the Region
4.3. Spatial Pattern in δ13C
4.4. Spatial Pattern in δ15N
4.5. Temporal Trends in δ15N
4.6. Current Status of BB-LEH-GB in Relation to Nutrient Pollution
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cloern, J. Our evolving conceptual model of the coastal eutrophication problem. MEPS 2001, 210, 223–253. [Google Scholar] [CrossRef] [Green Version]
- Howarth, R.; Chan, F.; Conley, D.J.; Garnier, J.; Doney, S.C.; Marino, R.; Billen, G. Coupled Biogeochemical Cycles: Eutrophication and Hypoxia in Temperate Estuaries and Coastal Marine Ecosystems. Front. Ecol. Environ. 2011, 9, 18–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, D.M.; Gilbert, P.M.; Burkholder, J.M. Harmful algal blooms and eutrophication: Nutrient sources, composition, and consequences. Estuaries 2002, 25, 704–726. [Google Scholar] [CrossRef]
- Cosper, E.M.; Dennison, W.; Milligan, A.; Carpenter, E.J.; Lee, C.; Holzapfel, J.; Milanese, L. An examination of the environmental factors important to initiating and sustaining “brown tide” blooms. In Novel Phytoplankton Blooms; Cosper, E.M., Bricelj, V.M., Carpenter, E.J., Eds.; Springer: Berlin, Heidelberg, 1989; pp. 317–340. [Google Scholar] [CrossRef]
- Deegan, L.A.; Johnson, D.S.; Warren, R.S.; Peterson, B.J.; Fleeger, J.W.; Fagherazzi, S.; Wollheim, W.M. Coastal eutrophication as a driver of salt marsh loss. Nature 2012, 490, 388–392. [Google Scholar] [CrossRef]
- Duarte, C.M.; Borja, A.; Carstensen, J.; Elliott, M.; Krause-Jensen, D.; Marbà, N. Paradigms in the recovery of estuarine and coastal ecosystems. Estuaries Coast 2015, 38, 1202–1212. [Google Scholar] [CrossRef]
- Lefcheck, J.S.; Orth, R.J.; Dennison, W.C.; Wilcox, D.J.; Murphy, R.R.; Keisman, J.; Gurbisz, C.; Hannam, M.; Landry, J.B.; Moore, K.A.; et al. Long-term nutrient reductions lead to unprecedented recovery of a temperate coastal region. Proc. Natl. Acad. Sci. USA 2018, 115, 3658–3662. [Google Scholar] [CrossRef] [Green Version]
- Staehr, P.A.; Testa, J.; Carstensen, J. Decadal changes in water quality and net productivity of a shallow Danish estuary following significant nutrient reductions. Estuaries Coast 2017, 40, 63–79. [Google Scholar] [CrossRef]
- Hunchak-Kariouk, K.; Nicholson, R.S. Watershed contributions of nutrients and other nonpoint source contaminants to the Barnegat Bay-Little Egg Harbor estuary. J. Coast Res. 2001, 32, 28–81. [Google Scholar]
- Taylor, D.I.; Oviatt, C.A.; Giblin, A.E.; Tucker, J.; Diaz, R.J.; Keay, K. Watewater input reductions reverse historic hypereutrophication ofBoston Harbor, USA. Ambio 2020, 49, 187–196. [Google Scholar] [CrossRef]
- Oczkowski, A.J.; Schmidt, C.; Santos, E.; Miller, K.; Hanson, A.; Cobb, D.; Krumholz, J.; Pimenta, A.; Heffner, L.; Robinson, S.; et al. How the distribution of anthropogenic nitrogen has changed in Narragansett Bay (RI, USA) following major reductions in nutrient loads. Estuaries Coast 2018, 41, 2260–2276. [Google Scholar] [CrossRef]
- Bowen, J.L.; Ramstack, J.M.; Mazzilli, S.; Valiela, I. NLOAD: An interactive, web-based modeling tool for nitrogen management in estuaries. Ecol. Appl. 2007, 17, S17–S30. [Google Scholar] [CrossRef]
- Santos, I.R.; Chen, X.; Lecher, A.L.; Sawyer, H.H.; Moosdorf, N.; Rodellas, V.; Tamborski, J.; Cho, H.-M.; Dimova, N.; Sugimoto, R.; et al. Submarine groundwater discharge impacts on coastal nutrient biogeochemistry. Nat. Rev. Earth Environ. 2021, 2, 307–323. [Google Scholar] [CrossRef]
- Green, L.; Magel, C.; Brown, C. Management pathways for the successful reduction of nonpoint source nutrients in coastal ecosystems. Reg. Stud. Mar. Sci. 2021, 45, 101851. [Google Scholar] [CrossRef]
- Burnett, W.C.; Aggarwal, P.K.; Aureli, A.; Bokuniewicz, H.; Cable, J.E.; Charette, M.A.; Kontar, E.; Krupa, S.; Kulkarni, K.M.; Loveless, A.; et al. Quantifying Submarine Groundwater Discharge in the Coastal Zone via Multiple Methods. Sci. Total Environ. 2006, 367, 498–543. [Google Scholar] [CrossRef] [PubMed]
- Sabo, R.D.; Sullivan, B.; Wu, C.; Trentacoste, E.; Zhang, Q.; Shenk, G.W.; Bhatt, G.; Linker, L.C. Major Point and Nonpoint Sources of Nutrient Pollution to Surface Water Have Declined throughout the Chesapeake Bay Watershed. Environ. Res. Commun. 2022, 4, 045012. [Google Scholar] [CrossRef]
- Taniguchi, M.; Burnett, W.C.; Cable, J.E.; Turner, J.V. Investigation of Submarine Groundwater Discharge. Hydrol. Process. 2002, 16, 2115–2129. [Google Scholar] [CrossRef]
- Greening, H.; Janicki, A.; Sherwood, E.T.; Pribble, R.; Johansson, J.O.R. Ecosystem Responses to Long-Term Nutrient Management in an Urban Estuary: Tampa Bay, Florida, USA. Estuar. Coast Shelf Sci. 2014, 151, A1–A16. [Google Scholar] [CrossRef]
- Kennish, M.J.; Fertig, B. Application and Assessment of a Nutrient Pollution Indicator Using Eelgrass (Zostera marina L.) in Barnegat Bay–Little Egg Harbor Estuary, New Jersey. Aquat. Bot. 2012, 96, 23–30. [Google Scholar] [CrossRef]
- Fry, B. Stable Isotope Ecology; Springer: New York, NY, USA, 2006; ISBN 978-0-387-30513-4. [Google Scholar]
- McClelland, J.W.; Valiela, I.; Michener, R.H. Nitrogen-Stable Isotope Signatures in Estuarine Food Webs: A Record of Increasing Urbanization in Coastal Watersheds. Limnol. Oceanogr. 1997, 42, 930–937. [Google Scholar] [CrossRef]
- Oczkowski, A.; Taplin, B.; Pruell, R.; Pimenta, A.; Johnson, R.; Grear, J. Carbon Stable Isotope Values in Plankton and Mussels Reflect Changes in Carbonate Chemistry Associated with Nutrient Enhanced Net Production. Front. Mar. Sci. 2018, 5, 43. [Google Scholar] [CrossRef]
- Watson, E.B.; Powell, E.; Maher, N.P.; Oczkowski, A.J.; Paudel, B.; Starke, A.; Szura, K.; Wigand, C. Indicators of Nutrient Pollution in Long Island, New York, Estuarine Environments. Mar. Environ. Res. 2018, 134, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Oczkowski, A.; Nixon, S.; Granger, S.; El-Sayed, A.-F.M.; Altabet, M.; McKinney, R. A Preliminary Survey of the Nitrogen and Carbon Isotope Characteristics of Fish from the Lagoons of Egypt’s Nile Delta. Estuaries Coast 2008, 31, 1130–1142. [Google Scholar] [CrossRef]
- Weinstein, M.P.; Litvin, S.Y.; Bosley, K.L.; Fuller, C.M.; Wainright, S.C. The Role of Tidal Salt Marsh as an Energy Source for Marine Transient and Resident Finfishes: A Stable Isotope Approach. Trans. Am. Fish Soc. 2000, 129, 797–810. [Google Scholar] [CrossRef]
- Martinetto, P.; Teichberg, M.; Valiela, I. Coupling of Estuarine Benthic and Pelagic Food Webs to Land-Derived Nitrogen Sources in Waquoit Bay, Massachusetts, USA. Mar. Ecol. Prog. Ser. 2006, 307, 37–48. [Google Scholar] [CrossRef]
- Litvin, S.Y.; Weinstein, M.P. Life History Strategies of Estuarine Nekton: The Role of Marsh Macrophytes, Benthic Microalgae, and Phytoplankton in the Trophic Spectrum. Estuaries 2003, 26, 552–562. [Google Scholar] [CrossRef]
- Black, H.D.; Andrus, C.F.T.; Lambert, W.J.; Rick, T.C.; Gillikin, D.P. Δ15N Values in Crassostrea Virginica Shells Provides Early Direct Evidence for Nitrogen Loading to Chesapeake Bay. Sci. Rep. 2017, 7, 44241. [Google Scholar] [CrossRef] [Green Version]
- Carmichael, R.; Hattenrath, T.; Valiela, I.; Michener, R. Nitrogen Stable Isotopes in the Shell of Mercenaria Mercenaria Trace Wastewater Inputs from Watersheds to Estuarine Ecosystems. Aquat. Biol. 2008, 4, 99–111. [Google Scholar] [CrossRef]
- Gillikin, D.P.; Lorrain, A.; Jolivet, A.; Kelemen, Z.; Chauvaud, L.; Bouillon, S. High-Resolution Nitrogen Stable Isotope Sclerochronology of Bivalve Shell Carbonate-Bound Organics. Geochim. Cosmochim. Acta 2017, 200, 55–66. [Google Scholar] [CrossRef] [Green Version]
- Darrow, E.S.; Carmichael, R.H.; Andrus, C.F.T.; Jackson, H.E. From Middens to Modern Estuaries, Oyster Shells Sequester Source-Specific Nitrogen. Geochim. Cosmochim. Acta 2017, 202, 39–56. [Google Scholar] [CrossRef]
- Graniero, L.E.; Grossman, E.L.; O’Dea, A. Stable Isotopes in Bivalves as Indicators of Nutrient Source in Coastal Waters in the Bocas Del Toro Archipelago, Panama. PeerJ 2016, 4, e2278. [Google Scholar] [CrossRef]
- Watson, E.B.; Szura, K.; Powell, E.; Maher, N.; Wigand, C. Cultural Eutrophication Is Reflected in the Stable Isotopic Composition of the Eastern Mudsnail, Nassarius Obsoletus. J. Environ. Qual. 2018, 47, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Teece, M.; Fogel, M.; Tuross, N.; McCourt, R.; Spamer, E. The Lewis and Clark Herbarium of The Academy of Natural Sciences Part 3. Modern Environmental Applications of a Historic Nineteenth Century Botanical Collection. Not. Nat. 2002, 477, 1–20. [Google Scholar]
- Hinz, H.; Moranta, J.; Balestrini, S.; Sciberras, M.; Pantin, J.R.; Monnington, J.; Zalewski, A.; Kaiser, M.J.; Sköld, M.; Jonsson, P.; et al. Stable Isotopes Reveal the Effect of Trawl Fisheries on the Diet of Commercially Exploited Species. Sci. Rep. 2017, 7, 6334. [Google Scholar] [CrossRef]
- Defne, Z.; Ganju, N.K. Quantifying the Residence Time and Flushing Characteristics of a Shallow, Back-Barrier Estuary: Application of Hydrodynamic and Particle Tracking Models. Estuaries Coast 2015, 38, 1719–1734. [Google Scholar] [CrossRef]
- Kennish, M.J.; Bricker, S.B.; Dennison, W.C.; Glibert, P.M.; Livingston, R.J.; Moore, K.A.; Noble, R.T.; Paerl, H.W.; Ramstack, J.M.; Seitzinger, S.; et al. Barnegat Bay-Little Egg Harbor Estuary: Case Study of a Highly Eutrophic Coastal Bay System. Ecol. Appl. 2007, 17, S3–S16. [Google Scholar] [CrossRef] [Green Version]
- Kennish, M.J.; Fertig, B.M.; Lathrop, R.G. Assessment of Nutrient Loading and Eutrophication in Barnegat Bay-Little Egg Harbor, New Jersey in Support of Nutrient Management Planning; Rutgers University: New Brunswick, NJ, USA, 2012. [Google Scholar] [CrossRef]
- Pang, H.; Ingelido, P.; Hirst, B.; Pflaumer, J.; Witt, A.; Zaman, A.; Aiello, J. Water Quality Condition and Assessment within the Barnegat Bay Watershed between 2011 and 2015. J. Coast Res. 2017, 78, 7–21. [Google Scholar] [CrossRef]
- U.S. Army Corps of Engineers New Jersey Back Bays Coastal Storm Risk Management Draft Integrated Feasibility Report and Tier1 Environmental Impact Statement 2021. Available online: https://www.nan.usace.army.mil/Portals/37/NYNJHATS%20Draft%20Integrated%20Feasibility%20Report%20Tier%201%20EIS.pdf (accessed on 7 September 2022).
- Oczkowski, A.; Gumbley, T.; Carter, B.; Carmichael, R.; Humphries, A. Establishing an Anthropogenic Nitrogen Baseline Using Native American Shell Middens. Front. Mar. Sci. 2016, 3, 79. [Google Scholar] [CrossRef] [Green Version]
- Kennish, M.J. Physical Description of the Barnegat Bay—Little Egg Harbor Estuarine System. J. Coast Res. 2001, 32, 13–27. [Google Scholar]
- Tetra Tech. New Jersey State Hazard Mitigation Plan, Appendix P. Watesheds of New Jersey; Office of Emergency Management: West Trenton, NJ, USA, 2014.
- NJ DEP. Land Use/Land Cover of New Jersey 2015. 2019. Available online: https://gisdata-njdep.opendata.arcgis.com/documents/6f76b90deda34cc98aec255e2defdb45/about (accessed on 12 August 2022).
- Bricker, S.B.; Clement, C.G.; Pirhalla, D.E.; Orlando, S.P.; Farrow, D.R.G. National Estuarine Eutrophication Assessment: Effects of Nutrient Enrichment in the Nation’s Estuaries; NOAA, National Ocean Service, Special Projects Office and the National Centers for Coastal Ocean Science: Silver Spring, MD, USA, 1999; Volume 2.
- Kennish, M.J.; Sakowicz, G.P.; Fertig, B. Recent Trends of Zostera marina (Eelgrass) in a Highly Eutrophic Coastal Lagoon in the Mid-Atlantic Region (USA). OJE 2016, 06, 243–253. [Google Scholar] [CrossRef] [Green Version]
- U.S. Department of Agriculture, Natural Resources Conservation Service. National Soil Survey Handbook, Title 430-VI. Available online: http://www.nrcs.usda.gov/wps/portal/nrcs/detail/soils/ref/?cid=nrcs142p2_054242 (accessed on 1 June 2022).
- Walthert, L.; Graf, U.; Kammer, A.; Luster, J.; Pezzotta, D.; Zimmermann, S.; Hagedorn, F. Determination of Organic and Inorganic Carbon, δ13C, and Nitrogen in Soils Containing Carbonates after Acid Fumigation with HCl. Z. Pflanz. Bodenk 2010, 173, 207–216. [Google Scholar] [CrossRef]
- Soil Survey Staff. Kellogg Soil Survey Laboratory Methods Manual; Soil Survey Investigations Report; U.S. Department of Agriculture, Natural Resources Conservation Service: Washington, DC, USA, 2022.
- NJ DEP. Coastal Water Quality Network. Available online: https://www.nj.gov/dep/bmw/coastalwaterquality.htm (accessed on 28 July 2022).
- NOAA. NOAA C-CAP Regional Landcover Coastal Change Analysis Program (C-CAP) Regional Land Cover; NOAA Office for Coastal Management: Charleston, SC, USA, 2010.
- Paudel, B.; Velinsky, D.; Belton, T.; Pang, H. Spatial Variability of Estuarine Environmental Drivers and Response by Phytoplankton: A Multivariate Modeling Approach. Ecol. Inform. 2016, 34, 1–12. [Google Scholar] [CrossRef]
- Gastrich, M.D.; Lathrop, R.; Haag, S.; Weinstein, M.P.; Danko, M.; Caron, D.A.; Schaffner, R. Assessment of Brown Tide Blooms, Caused by Aureococcus Anophagefferens, and Contributing Factors in New Jersey Coastal Bays: 2000–2002. Harmful Algae 2004, 3, 305–320. [Google Scholar] [CrossRef]
- Baker, R.J.; Wieben, C.M.; Lathrop, R.G.; Nicholson, R.S. Concentrations, Loads, and Yields of Total Nitrogen and Total Phosphorus in the Barnegat Bay-Little Egg Harbor Watershed, New Jersey, 1989–2011, at Multiple Spatial Scales; Scientific Investigations Report; 2014-5072; U.S. Geological Survey: Reston, VA, USA, 2014; 64p. [CrossRef]
- Deegan, L.A.; Garritt, R.H. Evidence for Spatial Variability in Estuarine Food Webs. Mar. Ecol. Prog. Ser. 1997, 147, 31–47. [Google Scholar] [CrossRef] [Green Version]
- Zimmerman, A.R.; Canuel, E.A. Bulk Organic Matter and Lipid Biomarker Composition of Chesapeake Bay Surficial Sediments as Indicators of Environmental Processes. Estuar. Coast Shelf Sci. 2001, 53, 319–341. [Google Scholar] [CrossRef] [Green Version]
- Wainright, S.C.; Fuller, C.M.; Mcguinness, L.R. Stable Isotopic Composition of Suspended Organic Matter in the Mullica River/Great Bay Estuary (Southern New Jersey, USA). Bull. N. J. Acad. Sci. 1999, 44, 8. [Google Scholar]
- Oczkowski, A.; Kreakie, B.; McKinney, R.A.; Prezioso, J. Patterns in Stable Isotope Values of Nitrogen and Carbon in Particulate Matter from the Northwest Atlantic Continental Shelf, from the Gulf of Maine to Cape Hatteras. Front. Mar. Sci. 2016, 3, 252. [Google Scholar] [CrossRef]
- Peterson, B.J.; Howarth, R.W.; Garritt, R.H. Multiple Stable Isotopes Used to Trace the Flow of Organic Matter in Estuarine Food Webs. Science 1985, 227, 1361–1363. [Google Scholar] [CrossRef]
- Pruell, R.J.; Taplin, B.K.; Lake, J.L.; Jayaraman, S. Nitrogen Isotope Ratios in Estuarine Biota Collected along a Nutrient Gradient in Narragansett Bay, Rhode Island, USA. Mar. Pollut. Bull. 2006, 52, 612–620. [Google Scholar] [CrossRef]
- Raven, J.A.; Walker, D.I.; Johnston, A.M.; Handley, L.L.; Kübler, J.E. Implications of 13 C Natural Abundance Measurements for Photosynthetic Performance by Marine Macrophytes in Their Natural Environment. Mar. Ecol. Prog. Ser. 1995, 123, 193–205. [Google Scholar] [CrossRef] [Green Version]
- Peterson, B.J.; Howarth, R.W. Sulfur, Carbon, and Nitrogen Isotopes Used to Trace Organic Matter Flow in the Salt-Marsh Estuaries of Sapelo Island, Georgia1: Stable Isotopes in a Salt Marsh. Limnol. Oceanogr. 1987, 32, 1195–1213. [Google Scholar] [CrossRef]
- Krause, J.; Oczkowski, A.; Watson, E.B. Improved Mapping of Coastal Salt Marsh Habitat Change at Barnegat Bay (NJ, USA) Using Object-Based Image Analysis of High-Resolution Aerial Imagery. Remote. Sens. Appl. Soc. Environ. 2023, 29, 100910. [Google Scholar] [CrossRef]
- Pruell, R.J.; Taplin, B.K.; Oczkowski, A.J.; Grear, J.S.; Mendoza, W.G.; Pimenta, A.R.; Hanson, A.R.; Miller, K.M. Nitrogen Isotope Fractionation in a Continuous Culture System Containing Phytoplankton and Blue Mussels. Mar. Pollut. Bull. 2020, 150, 110745. [Google Scholar] [CrossRef] [PubMed]
- Marcarelli, A.M.; Fulweiler, R.W.; Scott, J.T. Nitrogen Fixation: A Poorly Understood Process along the Freshwater-marine Continuum. Limnol. Oceanogr. Lett. 2022, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Romero, I.C.; Jacobson, M.; Fuhrman, J.A.; Fogel, M.; Capone, D.G. Long-Term Nitrogen and Phosphorus Fertilization Effects on N2 Fixation Rates and NifH Gene Community Patterns in Mangrove Sediments: Long-Term Nitrogen and Phosphorus Fertilization Effects. Mar. Ecol. 2012, 33, 117–127. [Google Scholar] [CrossRef]
- Wigand, C.; Roman, C.T.; Davey, E.W.; Stolt, M.; Johnson, R.L.; Hanson, A.R.; Watson, E.B.; Moran, S.B.; Cahoon, D.R.; Lynch, J.C.; et al. Below the Disappearing Marshes of an Urban Estuary: Historic Nitrogen Trends and Soil Structure. Ecol. Appl. 2014, 24, 633–649. [Google Scholar] [CrossRef] [Green Version]
- Velinsky, D.J.; Paudel, B.; Belton, T.J.; Sommerfield, C.K. Tidal Marsh Record of Nutrient Loadings in Barnegat Bay, New Jersey. J. Coast Res. 2017, 78, 79–88. [Google Scholar] [CrossRef]
- Tyler, A.C.; Mastronicola, T.A.; McGlathery, K.J. Nitrogen Fixation and Nitrogen Limitation of Primary Production along a Natural Marsh Chronosequence. Oecologia 2003, 136, 431–438. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krause, J.R.; Gannon, M.E.; Oczkowski, A.J.; Schwartz, M.J.; Champlin, L.K.; Steinmann, D.; Maxwell-Doyle, M.; Pirl, E.; Allen, V.; Burke Watson, E. Tidal Flushing Rather Than Non-Point Source Nitrogen Pollution Drives Nutrient Dynamics in A Putatively Eutrophic Estuary. Water 2023, 15, 15. https://doi.org/10.3390/w15010015
Krause JR, Gannon ME, Oczkowski AJ, Schwartz MJ, Champlin LK, Steinmann D, Maxwell-Doyle M, Pirl E, Allen V, Burke Watson E. Tidal Flushing Rather Than Non-Point Source Nitrogen Pollution Drives Nutrient Dynamics in A Putatively Eutrophic Estuary. Water. 2023; 15(1):15. https://doi.org/10.3390/w15010015
Chicago/Turabian StyleKrause, Johannes R., Michelle E. Gannon, Autumn J. Oczkowski, Morgan J. Schwartz, Lena K. Champlin, David Steinmann, Martha Maxwell-Doyle, Emily Pirl, Victoria Allen, and Elizabeth Burke Watson. 2023. "Tidal Flushing Rather Than Non-Point Source Nitrogen Pollution Drives Nutrient Dynamics in A Putatively Eutrophic Estuary" Water 15, no. 1: 15. https://doi.org/10.3390/w15010015