
Construction and Characterization of an Aeromonas hydrophila Multi-Gene Deletion Strain and Evaluation of Its Potential as a Live-Attenuated Vaccine in Grass Carp

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Declaration of Ethical Approval
2.2. Bacterial Strains and Growth Conditions
2.3. Construction of the A. hydrophila Five-Gene Deletion Mutant and Phenotype Characterization
2.4. Blood and Skin Mucus Killing Assay
2.5. Median Lethal Dose (LD50) Assays
2.6. Quantitative Real-Time Reverse Transcriptase-PCR (qRT-PCR)
2.7. Vaccination and Challenge Assay
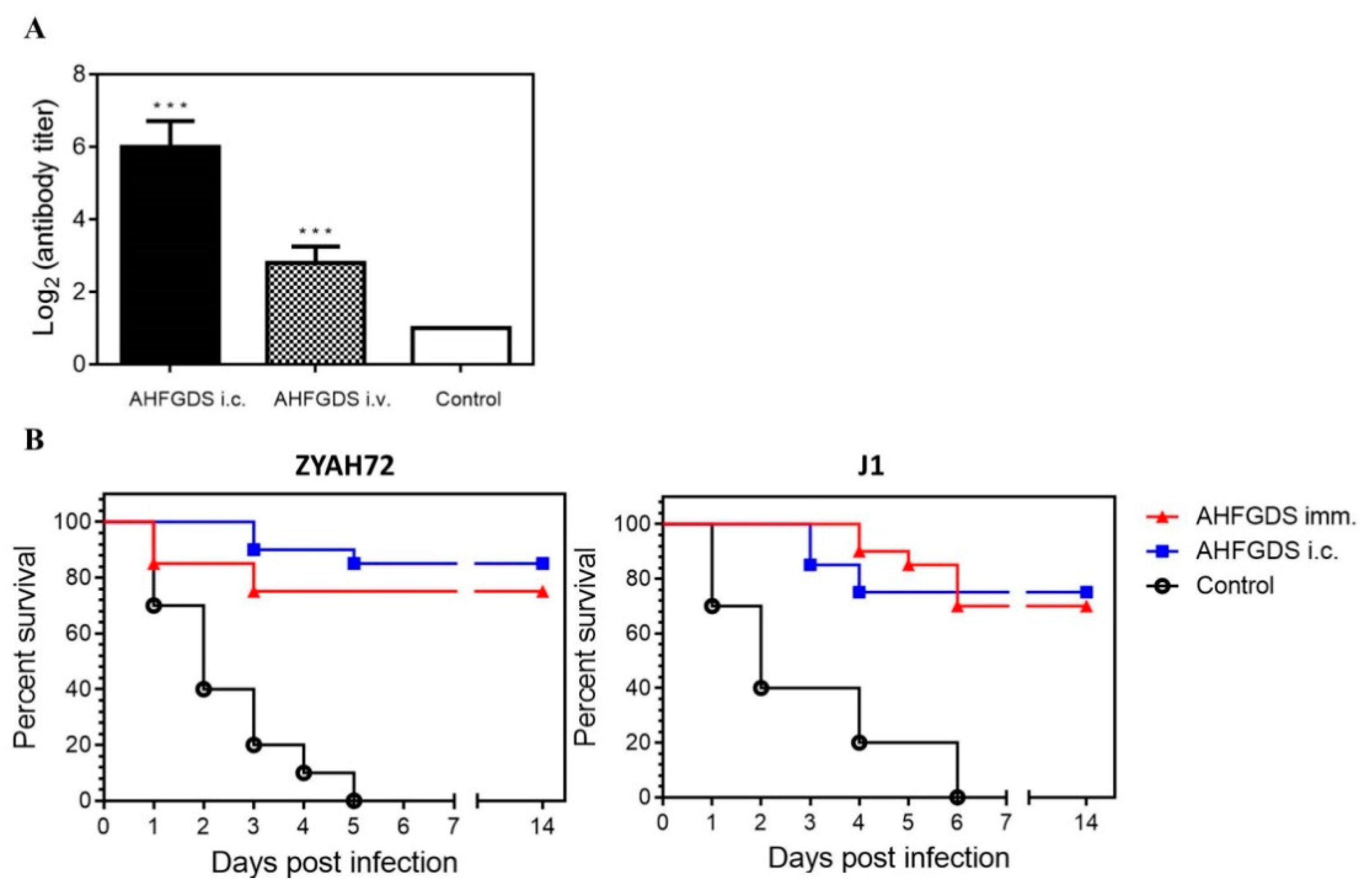
2.8. Agglutinating Antibody Titer
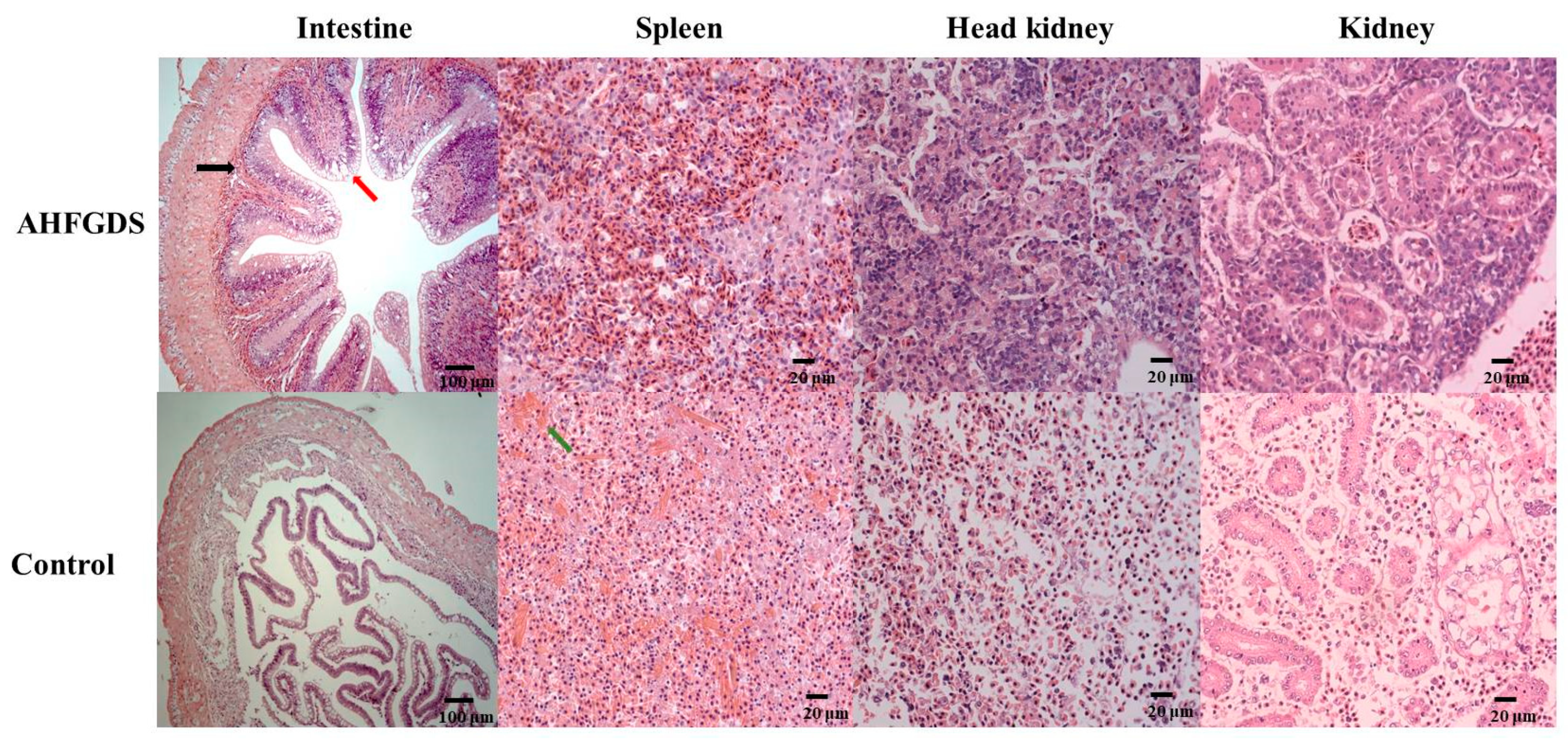
2.9. Histopathological Studies
2.10. Statistical Analysis
3. Results
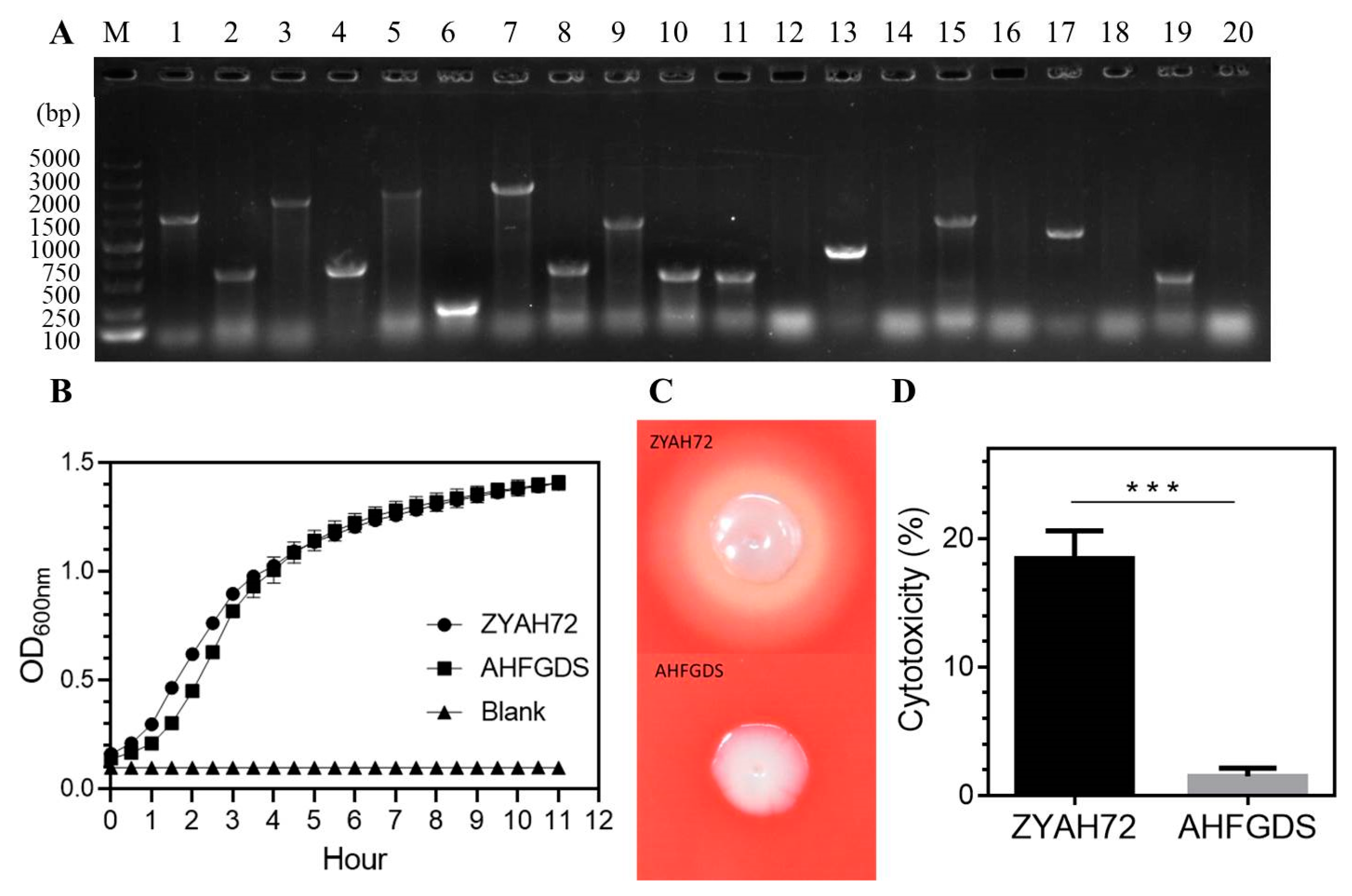
3.1. Construction and Characterization of the A. hydrophila Mutant AHFGDS
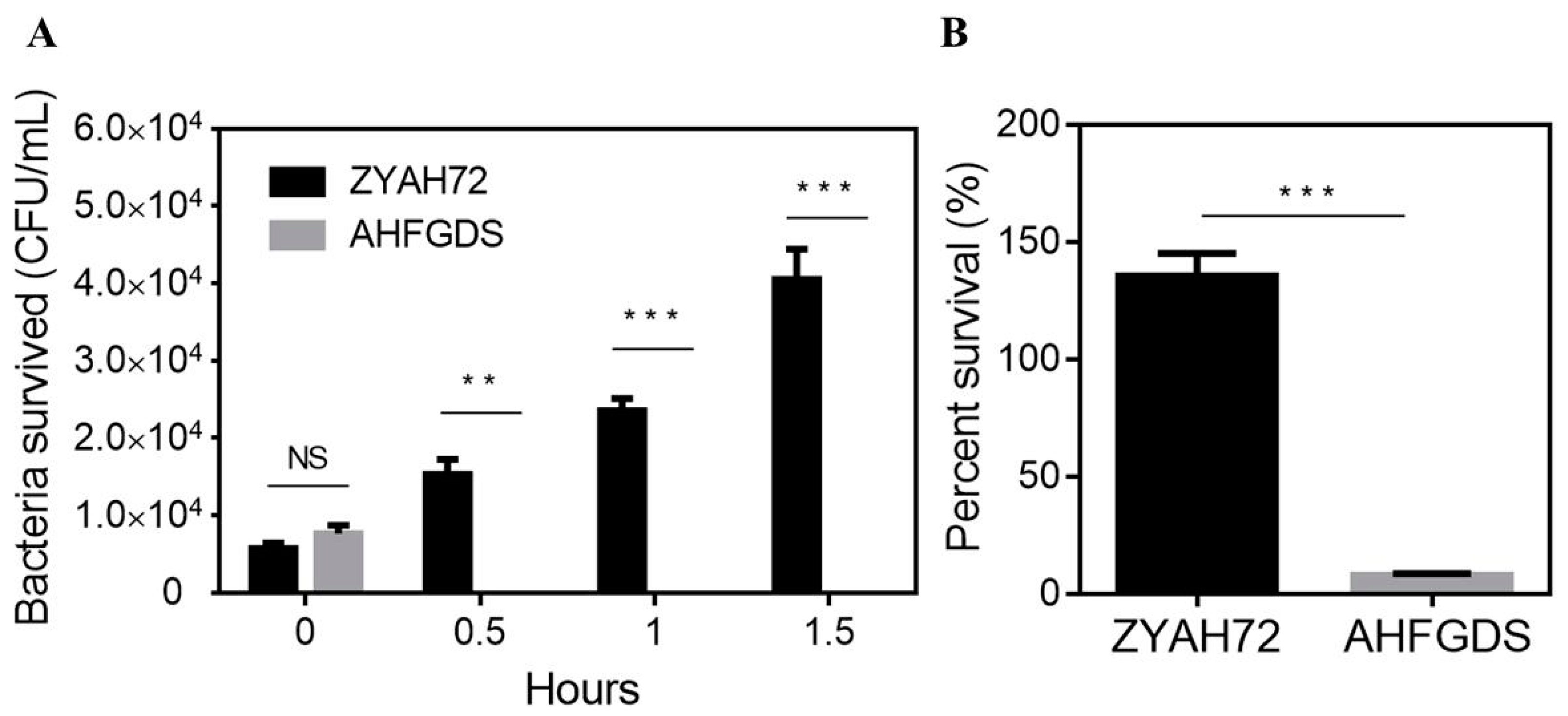
3.2. AHFGDS Was Sensitive against Host Clearance
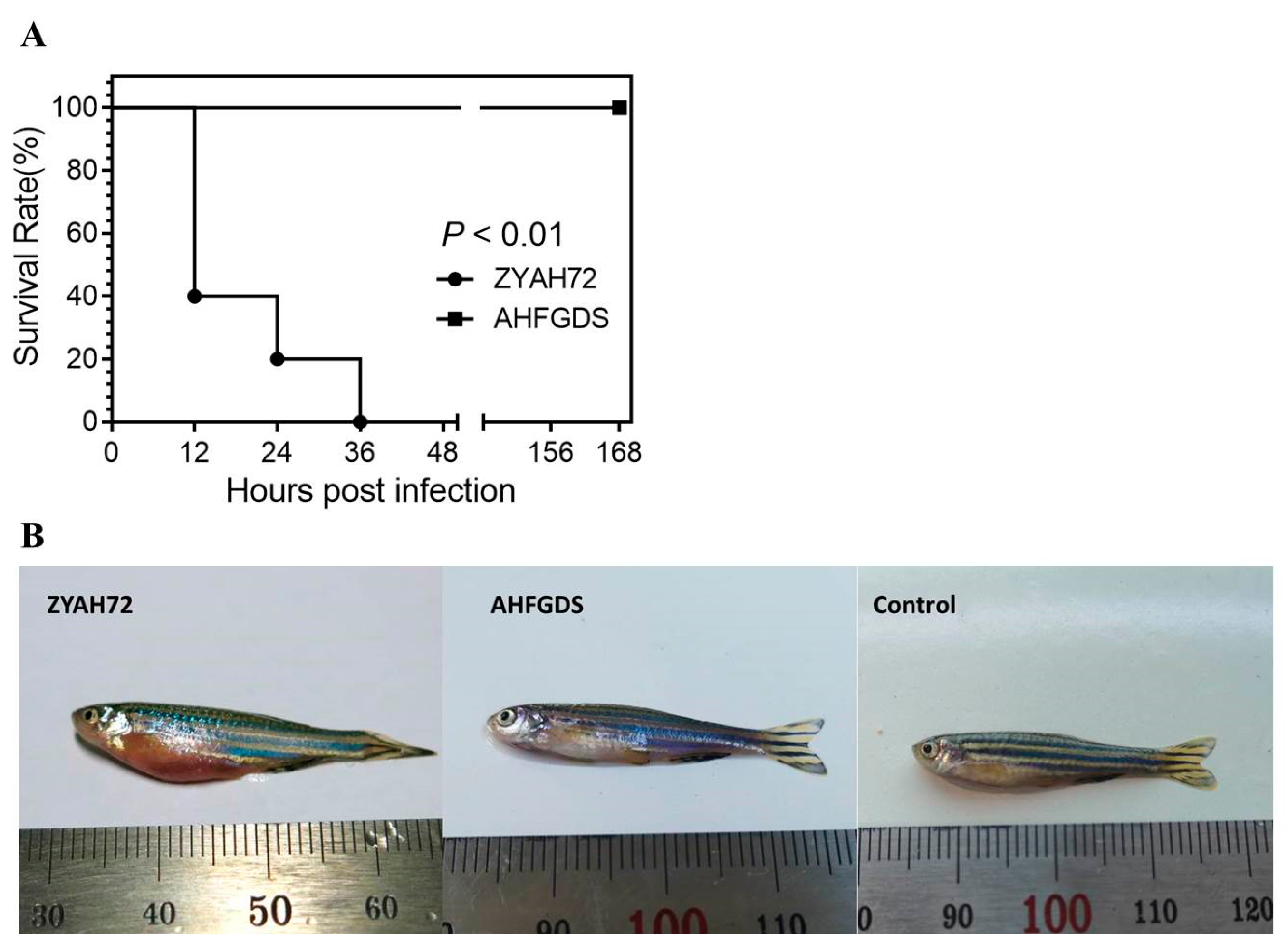
3.3. AHFGDS Was Highly Attenuated in Zebrafish
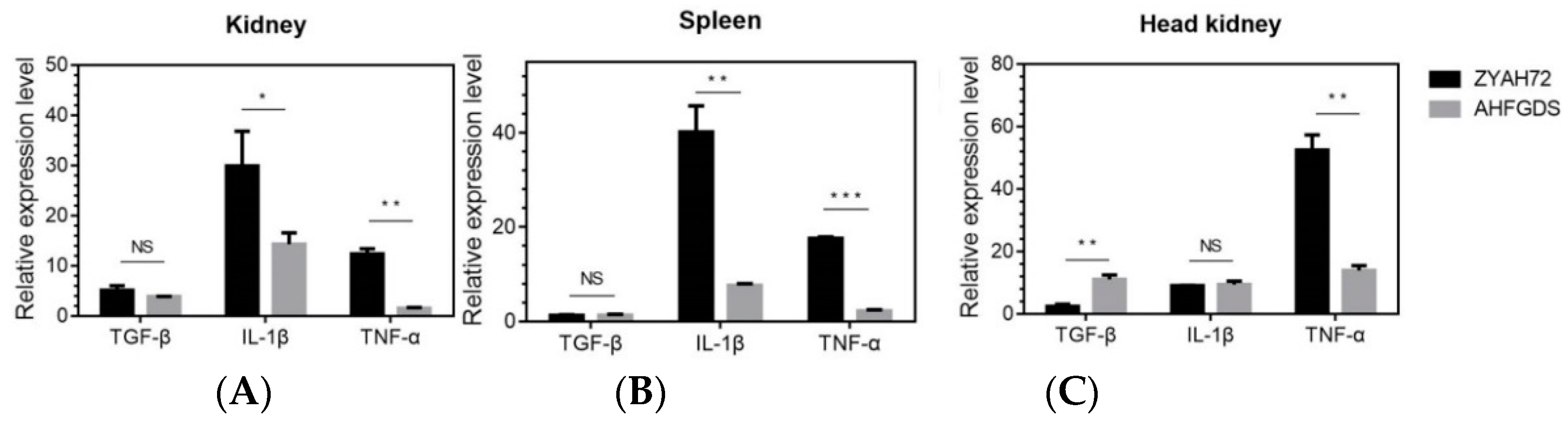
3.4. AHFGDS Elicited Weakened Inflammatory Responses
3.5. AHFGDS Offered Grass Carp Effective Protection against A. hydrophila Infection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Fernández-Bravo, A.; Figueras, M.J. An Update on the Genus Aeromonas: Taxonomy, Epidemiology, and Pathogenicity. Microorganisms 2020, 8, 129. [Google Scholar] [CrossRef] [Green Version]
- Pang, M.; Xie, X.; Wu, Y.; Dong, Y.; Zhang, W.; Yao, H.; Lu, C.; Liu, Y.; Jiang, J.; Kwok, A.H.Y.; et al. Novel insights into the pathogenicity of epidemic Aeromonas hydrophila ST251 clones from comparative genomics. Sci. Rep. 2015, 5, 9833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Y.; Li, S.; Han, S.; Wang, D.; Zhao, J.; Xu, L.; Liu, H.; Lu, T. Characterization and application of a novel Aeromonas bacteriophage as treatment for pathogenic Aeromonas hydrophila infection in rainbow trout. Aquaculture 2020, 523. [Google Scholar] [CrossRef]
- Pridgeon, J.W.; Yildirim-Aksoy, M.; Klesius, P.H.; Srivastava, K.K.; Reddy, P.G. Attenuation of a virulent A eromonas hydrophila with novobiocin and pathogenic characterization of the novobiocin-resistant strain. J. Appl. Microbiol. 2012, 113, 1319–1328. [Google Scholar] [CrossRef]
- Rasmussen-Ivey, C.R.; Hossain, M.J.; Odom, S.E.; Terhune, J.S.; Hemstreet, W.G.; Shoemaker, C.A.; Zhang, D.; Xu, D.-H.; Griffin, M.J.; Liu, Y.-J.; et al. Classification of a Hypervirulent Aeromonas hydrophila Pathotype Responsible for Epidemic Outbreaks in Warm-Water Fishes. Front. Microbiol. 2016, 7, 1615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patil, H.J.; Benet-Perelberg, A.; Naor, A.; Smirnov, M.; Ofek, T.; Nasser, A.; Minz, D.; Cytryn, E. Evidence of Increased Antibiotic Resistance in Phylogenetically-Diverse Aeromonas Isolates from Semi-Intensive Fish Ponds Treated with Antibiotics. Front. Microbiol. 2016, 7, 1875. [Google Scholar] [CrossRef] [Green Version]
- Kaskhedikar, M.; Chhabra, D. Multiple drug resistance in Aeromonas hydrophila isolates of fish. Food Microbiol. 2010, 28, 157–168. [Google Scholar] [CrossRef] [Green Version]
- Marshall, B.M.; Levy, S.B. Food animals and antimicrobials: Impacts on human health. Clin. Microbiol. Rev. 2011, 24, 718–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Ji, W.; Xu, Z. Current use and development of fish vaccines in China. Fish Shellfish Immunol. 2020, 96, 223–234. [Google Scholar] [CrossRef]
- Rao, Y.V.; Das, B.; Jyotyrmayee, P.; Chakrabarti, R. Effect of Achyranthes aspera on the immunity and survival of Labeo rohita infected with Aeromonas hydrophila. Fish Shellfish Immunol. 2006, 20, 263–273. [Google Scholar] [CrossRef]
- Magnadottir, B. Immunological Control of Fish Diseases. Mar. Biotechnol. 2010, 12, 361–379. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; Xu, K.; Liu, Z.; Ge, W.; Shao, S.; Li, P.; Yan, N.; Li, X.; Zhang, Z. Oral yeast-based DNA vaccine confers effective protection from Aeromonas hydrophila infection on Carassius auratus. Fish Shellfish Immunol. 2019, 84, 948–954. [Google Scholar] [CrossRef]
- Liu, L.; Gong, Y.-X.; Liu, G.-L.; Zhu, B.; Wang, G.-X. Protective immunity of grass carp immunized with DNA vaccine against Aeromonas hydrophila by using carbon nanotubes as a carrier molecule. Fish Shellfish Immunol. 2016, 55, 516–522. [Google Scholar] [CrossRef]
- Mzula, A.; Wambura, P.N.; Mdegela, R.H.; Shirima, G.M. Current State of Modern Biotechnological-BasedAeromonas hydrophilaVaccines for Aquaculture: A Systematic Review. BioMed Res. Int. 2019, 2019, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Detmer, A.; Glenting, J. Live bacterial vaccines—A review and identification of potential hazards. Microb. Cell Fact. 2006, 5, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Santos, Y.; Toranzo, A.E.; Barja, J.L.; Nieto, T.P.; Villa, T.G. Virulence properties and enterotoxin production of Aeromonas strains isolated from fish. Infect. Immun. 1988, 56, 3285–3293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dooley, J.S.; Trust, T.J. Surface protein composition of Aeromonas hydrophila strains virulent for fish: Identification of a surface array protein. J. Bacteriol. 1988, 170, 499–506. [Google Scholar] [CrossRef] [Green Version]
- El-Bahar, H.M.; Ali, N.G.; Aboyadak, I.M.; Khalil, S.A.E.S.; Ibrahim, M.S. Virulence genes contributing to Aeromonas hydrophila pathogenicity in Oreochromis niloticus. Int. Microbiol. 2019, 22, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ni, X.; Liu, Y.; Lu, C. Detection of three virulence genes alt, ahp and aerA in Aeromonas hydrophila and their relationship with actual virulence to zebrafish. J. Appl. Microbiol. 2011, 110, 823–830. [Google Scholar] [CrossRef]
- Wong, C.Y.F.; Heuzenroeder, M.W.; Flower, R.L.P. Inactivation of two haemolytic toxin genes in Aeromonas hydrophila attenuates virulence in a suckling mouse model. Microbiology 1998, 144, 291–298. [Google Scholar] [CrossRef] [Green Version]
- Klemm, D.J.; Stober, Q.J.; Lazorchak, J.M. Guidance for AssessingChemical ContaminantData for Use in FishAdvisories; US Environmental Protection Agency: Cincinnati, OH, USA, 1993.
- Kennedy, M.J.; Yancey, R.J.; Sanchez, M.S.; Rzepkowski, R.A.; Kelly, S.M.; Curtiss, R. Attenuation and immunogenicity of Δcya Δcrp derivatives of Salmonella choleraesuis in pigs. Infect. Immun. 1999, 67, 4628–4636. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.Y.; Dozois, C.M.; Tinge, S.A.; Lee, T.H.; Curtiss, R. Transduction-Mediated Transfer of Unmarked Deletion and Point Mutations through Use of Counterselectable Suicide Vectors. J. Bacteriol. 2002, 184, 307–312. [Google Scholar] [CrossRef] [Green Version]
- Xie, Q.; Mei, W.; Ye, X.; Zhou, P.; Islam, S.; Elbassiony, K.R.A.; Yuan, C.; Li, J.; Zhou, Y. The two-component regulatory system CpxA/R is required for the pathogenesis of Aeromonas hydrophila. FEMS Microbiol. Lett. 2018, 365, fny218. [Google Scholar] [CrossRef]
- Balebona, M.C.; Andreu, M.J.; Bordas, M.A.; Zorrilla, I.; Moriñigo, M.A.; Borrego, J.J. Pathogenicity of Vibrio alginolyticusfor Cultured Gilt-Head Sea Bream (Sparus aurata L.). Appl. Environ. Microbiol. 1998, 64, 4269–4275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Ma, S.; Woo, N.Y.S. Vaccination of Silver Sea Bream (Sparus sarba) against Vibrio alginolyticus: Protective Evaluation of Different Vaccinating Modalities. Int. J. Mol. Sci. 2015, 17, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galindo, C.; Sha, J.; Fadl, A.; Pillai, L.; Chopra, A. Host Immune Responses to Aeromonas Virulence Factors. Curr. Immunol. Rev. 2006, 2, 13–26. [Google Scholar] [CrossRef]
- Heuzenroeder, M.W.; Wong, C.Y.; Flower, R.L. Distribution of two hemolytic toxin genes in clinical and environmental isolates of Aeromonasspp: Correlation with virulence in a suckling mouse model. FEMS Microbiol. Lett. 1999, 174, 131–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilera-Arreola, M.G.; Hernández-Rodríguez, C.; Zúñiga, G.; Figueras, M.J.; Garduño, R.A.; Castro-Escarpulli, G. Virulence potential and genetic diversity of Aeromonas caviae, Aeromonas veronii, and Aeromonas hydrophila clinical isolates from Mexico and Spain: a comparative study. Can. J. Microbiol. 2007, 53, 877–887. [Google Scholar] [CrossRef]
- Sha, J.; Kozlova, E.V.; Chopra, A.K. Role of Various Enterotoxins in Aeromonas hydrophila-Induced Gastroenteritis: Generation of Enterotoxin Gene-Deficient Mutants and Evaluation of Their Enterotoxic Activity. Infect. Immun. 2002, 70, 1924–1935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merino, S.; Shaw, J.G.; Tomã¡s, J.M. Bacterial lateral flagella: an inducible flagella system. FEMS Microbiol. Lett. 2006, 263, 127–135. [Google Scholar] [CrossRef]
- Minor, P.D. Live attenuated vaccines: Historical successes and current challenges. Virology 2015, 479, 379–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, X.; Zhang, C.; Zhao, Y.; Kong, X.; Pei, C.; Li, L.; Nie, G.; Li, X. Immune effects of the vaccine of live attenuated Aeromonas hydrophila screened by rifampicin on common carp (Cyprinus carpio L.). Vaccine 2016, 34, 3087–3092. [Google Scholar] [CrossRef]
- Liu, Y.; Bi, Z. Potential use of a transposon Tn916-generated mutant of Aeromonas hydrophila J-1 defective in some exoproducts as a live attenuated vaccine. Prev. Vet. Med. 2007, 78, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Swain, P.; Behera, T.; Mohapatra, D.; Nanda, P.K.; Nayak, S.K.; Meher, P.K.; Das, B.K. Derivation of rough attenuated variants from smooth virulent Aeromonas hydrophila and their immunogenicity in fish. Vaccine 2010, 28, 4626–4631. [Google Scholar] [CrossRef]
- Chopra, A.K.; Xu, X.J.; Ribardo, D.; Gonzalez, M.; Kuhl, K.; Peterson, J.W.; Houston, C.W. The cytotoxic enterotoxin of Aeromonas hydrophila induces proinflammatory cytokine production and activates arachidonic acid metabolism in macrophages. Infect. Immun. 2000, 68, 2808–2818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, S.-M.; Compans, R.W. Host responses from innate to adaptive immunity after vaccination: Molecular and cellular events. Mol. Cells 2009, 27, 5–14. [Google Scholar] [CrossRef]
- Schmitz, M.L.; Weber, A.; Roxlau, T.; Gaestel, M.; Kracht, M. Signal integration, crosstalk mechanisms and networks in the function of inflammatory cytokines. Biochim. Biophys. Acta Bioenerg. 2011, 1813, 2165–2175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.; Lin, C.-N.; Wang, Y.-C.; Liu, F.-Y.; Chuang, Y.-J.; Lan, C.-Y.; Hsieh, W.-P.; Chen, B.-S. The role of TGF-β signaling and apoptosis in innate and adaptive immunity in zebrafish: A systems biology approach. BMC Syst. Biol. 2014, 8, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Embregts, C.W.; Forlenza, M. Oral vaccination of fish: Lessons from humans and veterinary species. Dev. Comp. Immunol. 2016, 64, 118–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munang’Andu, H.M.; Mutoloki, S.; Evensen, Ø. A Review of the Immunological Mechanisms Following Mucosal Vaccination of Finfish. Front. Immunol. 2015, 6, 427. [Google Scholar] [CrossRef] [Green Version]
- Bøgwald, J.; Dalmo, R.A. Review on Immersion Vaccines for Fish: An Update 2019. Microorganisms 2019, 7, 627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shoemaker, C.A.; Mohammed, H.H.; Bader, T.J.; Peatman, E.; Beck, B.H. Immersion vaccination with an inactivated virulent Aeromonas hydrophila bacterin protects hybrid catfish (Ictalurus punctatus X Ictalurus furcatus) from motile Aeromonas septicemia. Fish Shellfish Immunol. 2018, 82, 239–242. [Google Scholar] [CrossRef] [PubMed]
- Acar, Ü.; Parrino, V.; Kesbiç, O.S.; Paro, G.L.; Saoca, C.; Abbate, F.; Yılmaz, S.; Fazio, F. Effects of Different Levels of Pomegranate Seed Oil on Some Blood Parameters and Disease Resistance Against Yersinia ruckeri in Rainbow Trout. Front. Physiol. 2018, 9, 596. [Google Scholar] [CrossRef] [PubMed]
- Parrino, V.; Kesbiç, O.S.; Acar, Ü.; Fazio, F. Hot pepper (Capsicum sp.) oil and its effects on growth performance and blood parameters in rainbow trout (Oncorhynchus mykiss). Nat. Prod. Res. 2020, 34, 3226–3230. [Google Scholar] [CrossRef]
- Song, X.; Zhao, J.; Bo, Y.; Liu, Z.; Wu, K.; Gong, C. Aeromonas hydrophila induces intestinal inflammation in grass carp (Ctenopharyngodon idella): An experimental model. Aquaclture 2014, 434, 171–178. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains and Plasmids | Description | Source |
|---|---|---|
| Strains | ||
| Aeromonas hydrophila | ||
| ZYAH72 | Wild type | Lab collection |
| AHFGDS | ZYAH72 derivative, ΔastΔaltΔaphΔhlyΔaerA | This work |
| Escherichia coli | ||
| χ7213 | thr-1 leuB6 fhuA21 lacY1 glnV44 recA1 ΔasdA4 Δ(zhf-2::Tn10) thi-1 | [22] |
| Plasmids | ||
| pRE112 | Suicide vector, sacB, mob-(RP4)R6K ori, Cmr | [23] |
| pRE112-ast | pRE112 derivative, designed for knockout of ast, Cmr | This work |
| pRE112-alt | pRE112 derivative, designed for knockout of alt, Cmr | This work |
| pRE112-aph | pRE112 derivative, designed for knockout of aph, Cmr | This work |
| pRE112-hly | pRE112 derivative, designed for knockout of hly, Cmr | This work |
| pRE112-aerA | pRE112 derivative, designed for knockout of aerA, Cmr | This work |
| Dose of Challenge CFU | Number of Death/Total | Survival Rate (%) | ||
|---|---|---|---|---|
| ZYAH72 | AHFGDS | ZYAH72 | AHFGDS | |
| 4.3 × 107 | - | 10/10 | - | 0 |
| 8.6 × 106 | - | 8/10 | - | 20 |
| 1.7 × 106 | - | 4/10 | - | 60 |
| 3.4 × 105 | 10/10 | 0/10 | 0 | 100 |
| 6.8 × 104 | 10/10 | 0/10 | 0 | 100 |
| 1.4 × 104 | 4/10 | - | 60 | - |
| 2.7 × 103 | 2/10 | - | 80 | - |
| 5.4 × 102 | 0/10 | - | 100 | - |
| LD50 * | 1.2 × 104 | 2.8 × 106 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Ma, S.; Li, Z.; Yu, W.; Zhou, P.; Ye, X.; Islam, M.S.; Zhang, Y.-A.; Zhou, Y.; Li, J. Construction and Characterization of an Aeromonas hydrophila Multi-Gene Deletion Strain and Evaluation of Its Potential as a Live-Attenuated Vaccine in Grass Carp. Vaccines 2021, 9, 451. https://doi.org/10.3390/vaccines9050451
Li J, Ma S, Li Z, Yu W, Zhou P, Ye X, Islam MS, Zhang Y-A, Zhou Y, Li J. Construction and Characterization of an Aeromonas hydrophila Multi-Gene Deletion Strain and Evaluation of Its Potential as a Live-Attenuated Vaccine in Grass Carp. Vaccines. 2021; 9(5):451. https://doi.org/10.3390/vaccines9050451
Chicago/Turabian StyleLi, Jihong, Shilin Ma, Zhi Li, Wei Yu, Peng Zhou, Xiang Ye, Md. Sharifull Islam, Yong-An Zhang, Yang Zhou, and Jinquan Li. 2021. "Construction and Characterization of an Aeromonas hydrophila Multi-Gene Deletion Strain and Evaluation of Its Potential as a Live-Attenuated Vaccine in Grass Carp" Vaccines 9, no. 5: 451. https://doi.org/10.3390/vaccines9050451