Distinct miRNA Profile of Cellular and Extracellular Vesicles Released from Chicken Tracheal Cells Following Avian Influenza Virus Infection

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Avian Influenza Virus (AIV)
2.2. Toll-Like Receptor (TLR) Ligands
2.3. Tracheal Organ Culture (TOC)
2.4. TOC Infection with AIV (H4N6) and Stimulation with TLR Ligands
2.5. Extracellular Vesicle Isolation
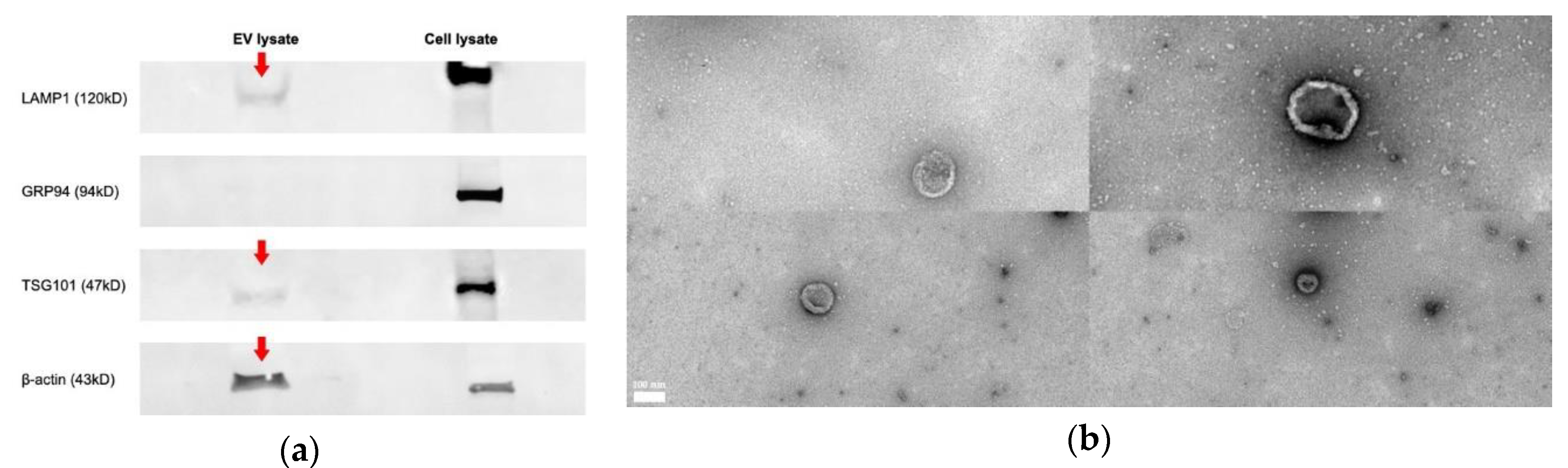
2.6. Western Blot
2.7. Negative Staining and Transmission Electron Microscopy (TEM)
2.8. MiRNA Isolation
2.9. Small RNA Library Preparation and Sequencing
2.10. MiRNA Expression Analysis
2.11. In Silico Target Gene Prediction and Pathway Analysis
3. Results
3.1. Chicken Tracheal Cells Release Extracellular Vesicles
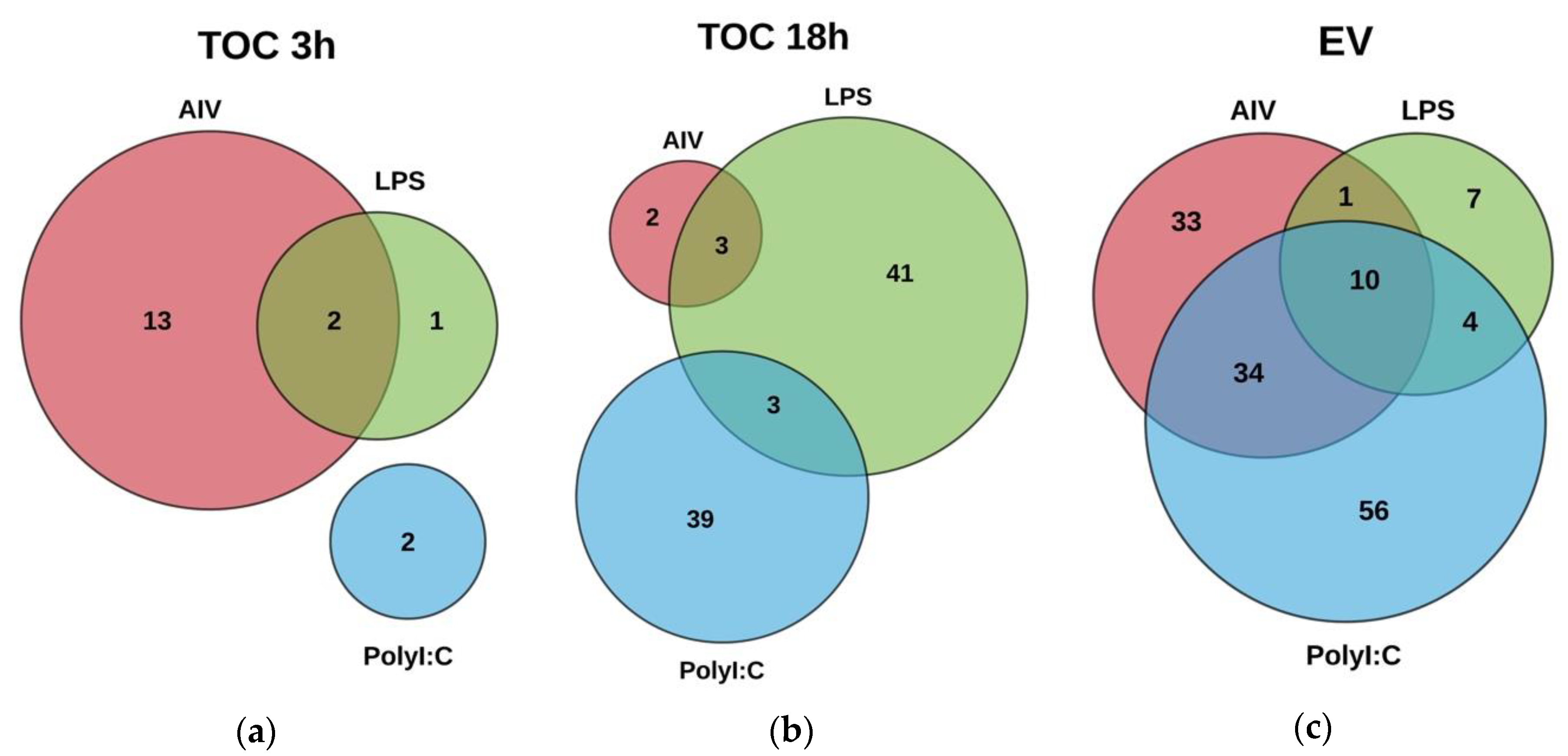
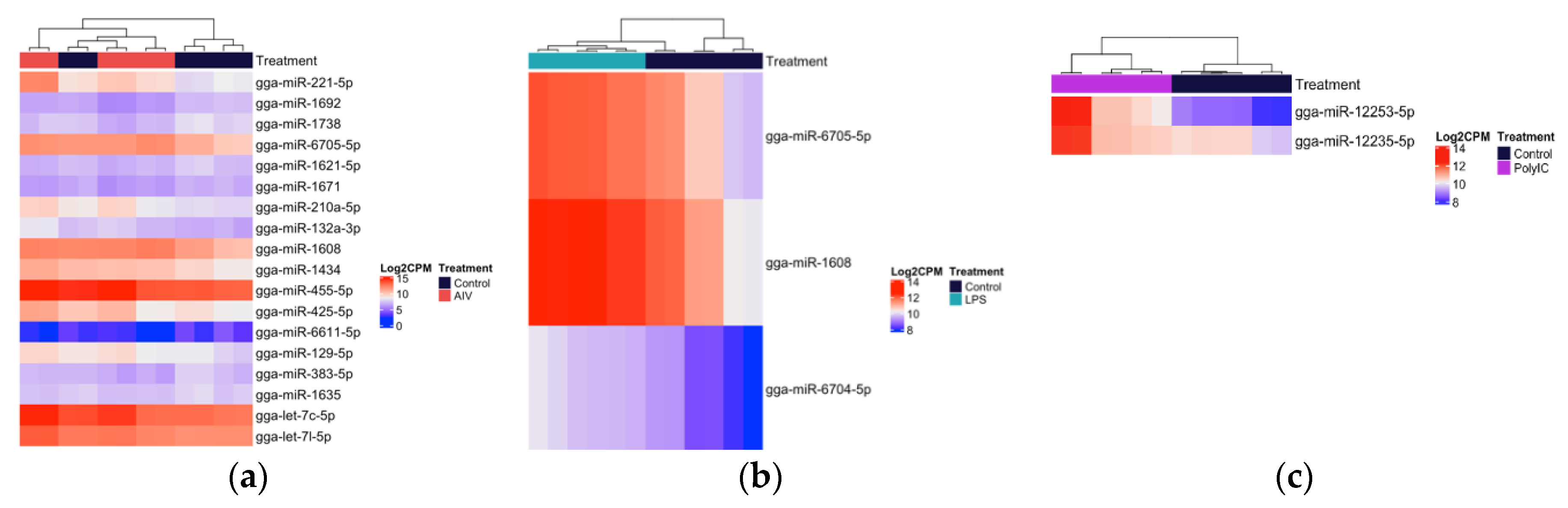
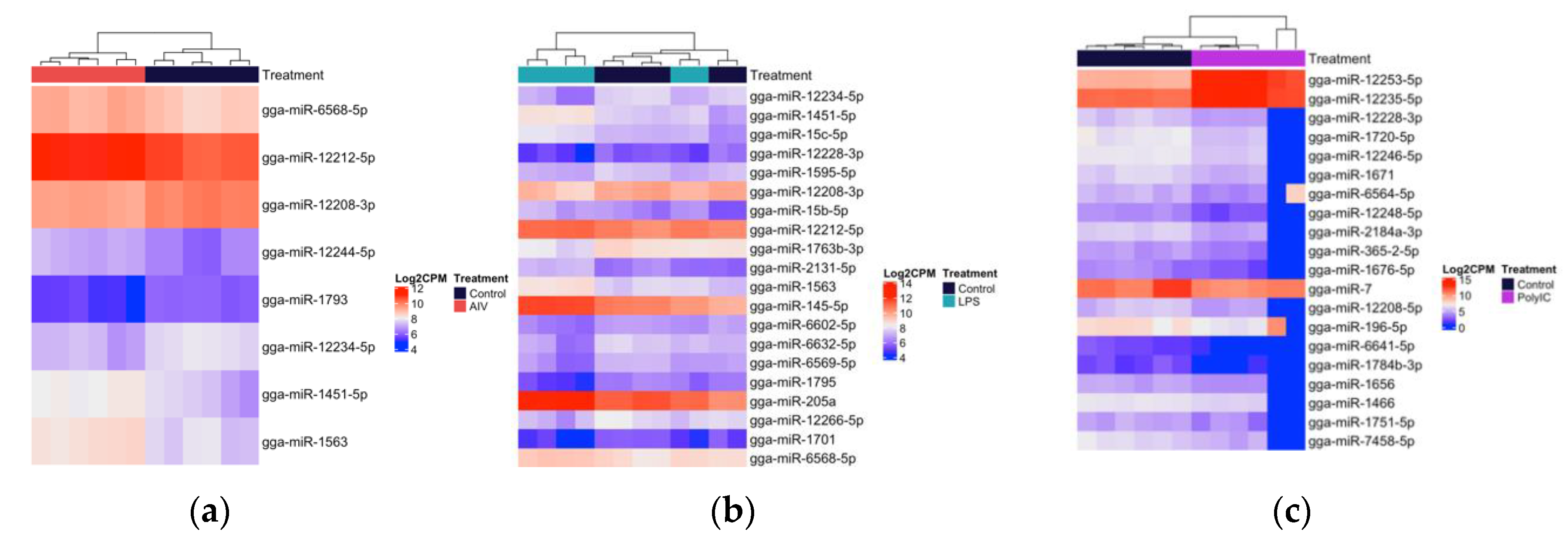
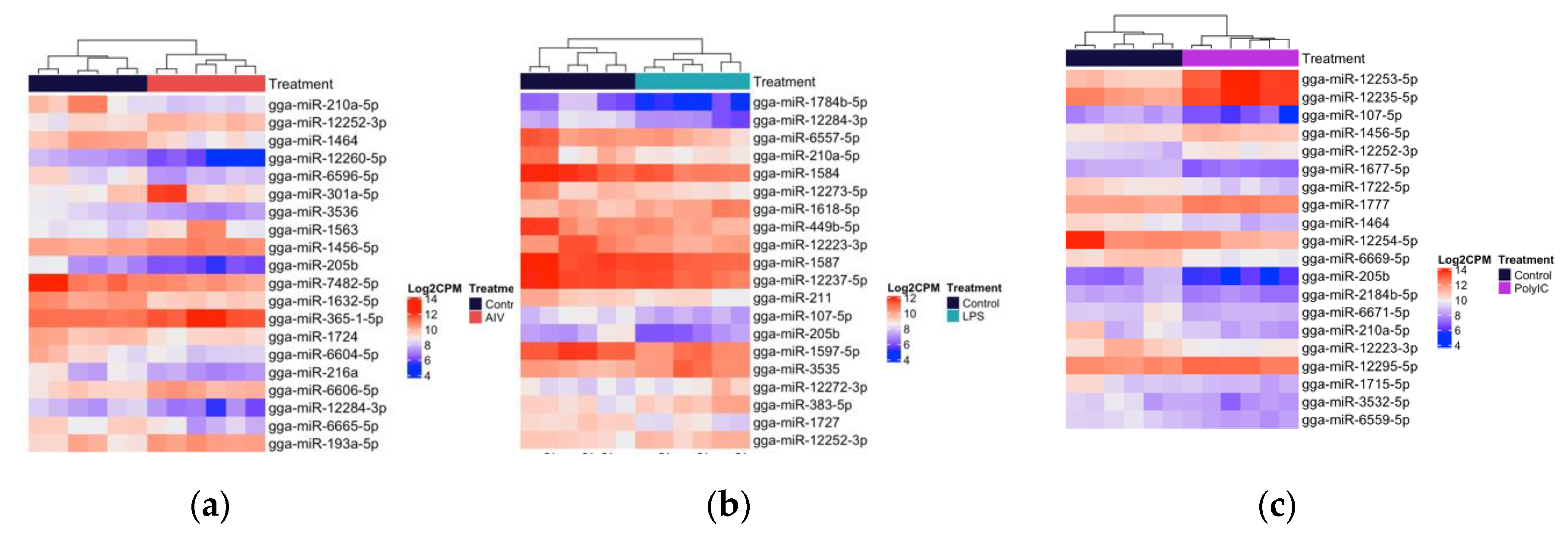
3.2. Cellular and EV Treatment Groups Have Distinct miRNAs Expression Profiles
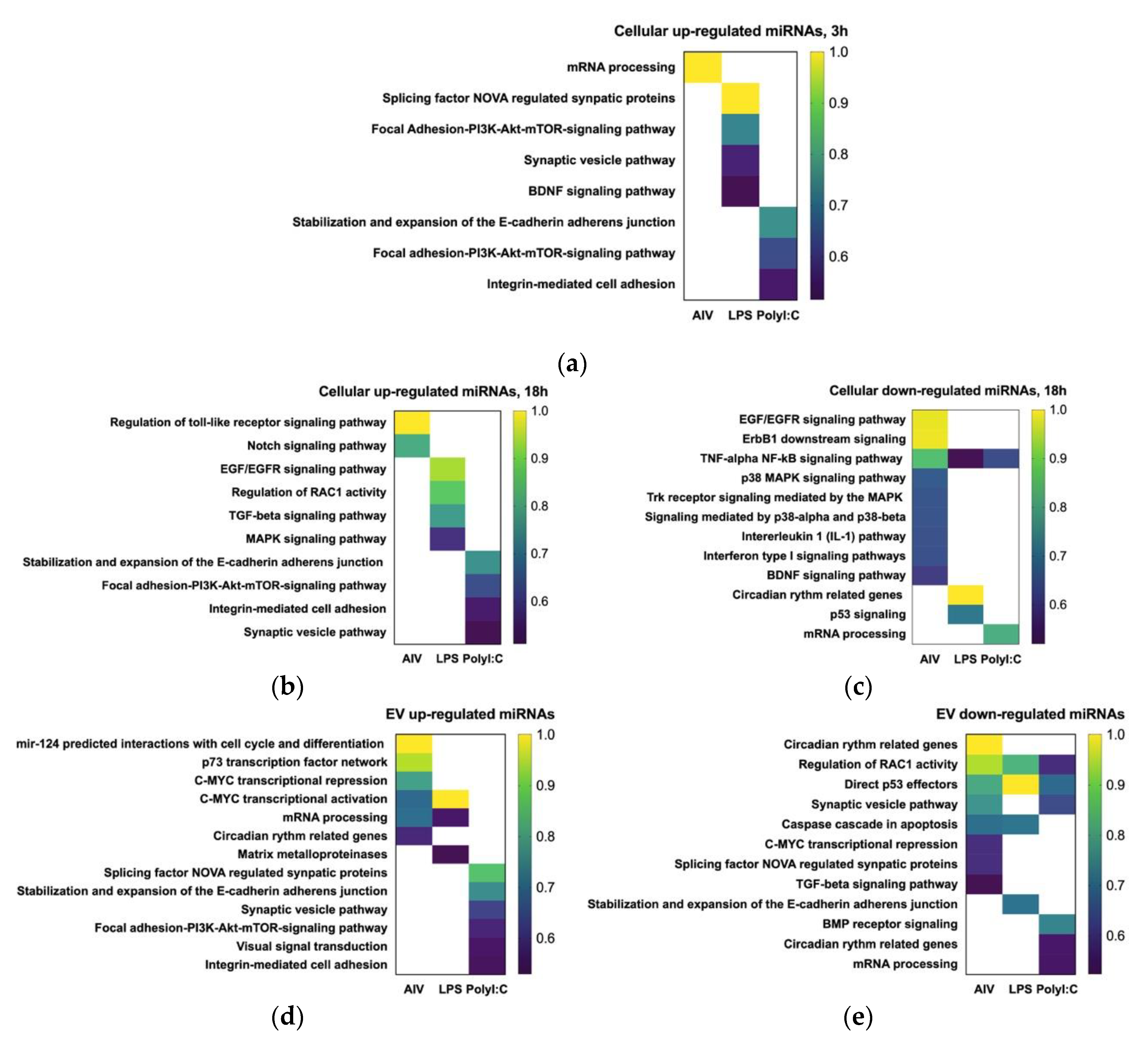
3.3. Target Gene Prediction and Functional Annotation Reveals DE miRNAs Target Multiple Pathways
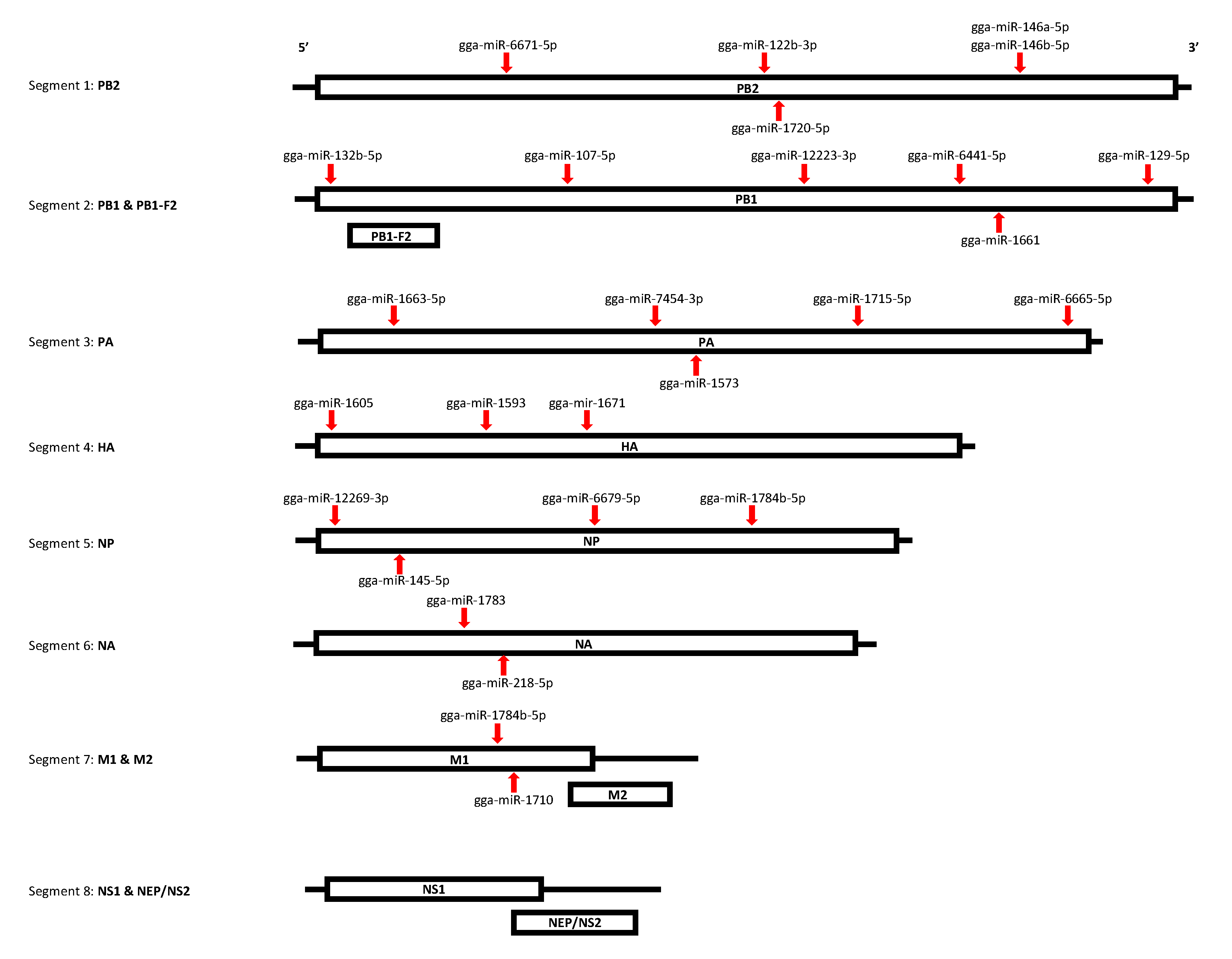
3.4. The Functional Annotation Reveals DE miRNAs Target Multiple Segments of the AIV Viral Genome
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Barjesteh, N.; Shojadoost, B.; Brisbin, J.T.; Emam, M.; Hodgins, D.C.; Nagy, É.; Sharif, S. Reduction of avian influenza virus shedding by administration of Toll-like receptor ligands to chickens. Vaccine 2015, 33, 4843–4849. [Google Scholar] [CrossRef] [PubMed]
- Barjesteh, N.; Alkie, T.N.; Hodgins, D.C.; Nagy, É.; Sharif, S. Local innate responses to TLR ligands in the chicken trachea. Viruses 2016, 8, 207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, M.S.; Mallick, A.I.; Read, L.R.; Villanueva, A.I.; Parvizi, P.; Abdul-Careem, M.F.; Nagy, É.; Sharif, S. Prophylactic treatment with Toll-like receptor ligands enhances host immunity to avian influenza virus in chickens. Vaccine 2012, 30, 4524–4531. [Google Scholar] [CrossRef] [PubMed]
- Rivera, A.; Siracusa, M.C.; Yap, G.S.; Gause, W.C. Innate cell communication kick-starts pathogen-specific immunity. Nat. Immunol. 2016, 17, 356–363. [Google Scholar] [CrossRef]
- McCoy-Simandle, K.; Hanna, S.J.; Cox, D. Exosomes and nanotubes: Control of immune cell communication. Int. J. Biochem. Cell Biol. 2016, 71, 44–54. [Google Scholar] [CrossRef] [Green Version]
- Willms, E.; Johansson, H.J.; Mäger, I.; Lee, Y.; Blomberg, K.E.M.; Sadik, M.; Alaarg, A.; Smith, C.I.E.; Lehtiö, J.; El Andaloussi, S.; et al. Cells release subpopulations of exosomes with distinct molecular and biological properties. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Y.; Liu, H.; Tang, W.H. Exosomes: Biogenesis, biologic function and clinical potential. Cell Biosci. 2019, 9, 1–18. [Google Scholar] [CrossRef]
- Chahar, H.S.; Corsello, T.; Kudlicki, A.S.; Komaravelli, N.; Casola, A. Respiratory syncytial virus infection changes cargo composition of exosome released from airway epithelial cells. Sci. Rep. 2018, 8, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Chettimada, S.; Lorenz, D.R.; Misra, V.; Dillon, S.T.; Reeves, R.K.; Manickam, C.; Morgello, S.; Kirk, G.D.; Mehta, S.H.; Gabuzda, D. Exosome markers associated with immune activation and oxidative stress in HIV patients on antiretroviral therapy. Sci. Rep. 2018, 8, 1–16. [Google Scholar] [CrossRef]
- Petrik, J. Immunomodulatory effects of exosomes produced by virus-infected cells. Transfus. Apher. Sci. 2016, 55, 84–91. [Google Scholar] [CrossRef]
- Yao, Z.; Qiao, Y.; Li, X.; Chen, J.; Ding, J.; Bai, L.; Shen, F.; Shi, B.; Liu, J.; Peng, L.; et al. Exosomes exploit the virus entry machinery and pathway to transmit alpha interferon-induced antiviral activity. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brennecke, J.; Stark, A.; Russell, R.B.; Cohen, S.M. Principles of microRNA-target recognition. PLoS Biol. 2005, 3. [Google Scholar] [CrossRef] [PubMed]
- Catalanotto, C.; Cogoni, C.; Zardo, G. MicroRNA in control of gene expression: An overview of nuclear functions. Int. J. Mol. Sci. 2016, 17, 1712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, B.P.; Burge, C.B.; Bartel, D.P. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell 2005, 120, 15–20. [Google Scholar] [CrossRef] [Green Version]
- Felekkis, K.; Touvana, E.; Stefanou, C.; Deltas, C. MicroRNAs: A newly described class of encoded molecules that play a role in health and disease. Hippokratia 2010, 14, 236–240. [Google Scholar]
- Sohel, M.H. Extracellular/circulating microRNAs: Release mechanisms, functions and challenges. Achiev. Life Sci. 2016, 10, 175–186. [Google Scholar] [CrossRef] [Green Version]
- Buggele, W.A.; Johnson, K.E.; Horvath, C.M. Influenza A virus infection of human respiratory cells induces primary microRNA expression. J. Biol. Chem. 2012, 287, 31027–31040. [Google Scholar] [CrossRef] [Green Version]
- Buggele, W.A.; Krause, K.E.; Horvath, C.M. Small RNA profiling of influenza A virus-infected cells identifies miR-449b as a regulator of histone deacetylase 1 and interferon beta. PLoS ONE 2013, 8, e76560. [Google Scholar] [CrossRef] [Green Version]
- Yarbrough, M.L.; Zhang, K.; Sakthivel, R.; Forst, C.V.; Posner, B.A.; Barber, G.N.; White, M.A.; Fontoura, B.M.A. Primate-specific miR-576-3p sets host defense signalling threshold. Nat. Commun. 2014, 5, 4963. [Google Scholar] [CrossRef]
- McCaskill, J.L.; Ressel, S.; Alber, A.; Redford, J.; Power, U.F.; Schwarze, J.; Dutia, B.M.; Buck, A.H. Broad-spectrum inhibition of respiratory virus infection by microRNA mimics targeting p38 MAPK signaling. Mol. Ther. Nucleic Acids 2017, 7, 256–266. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Song, Y.; He, L.; Wan, X.; Lai, L.; Dai, F.; Liu, Y.; Wang, Q. MicroRNA-223 promotes type i interferon production in antiviral innate immunity by targeting forkhead box protein O3 (FOXO3). J. Biol. Chem. 2016, 291, 14706–14716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, L.; Liu, H.; Gao, S.; Jiang, W.; Huang, W. Cellular microRNAs inhibit replication of the H1N1 influenza A virus in infected cells. J. Virol. 2010, 84, 8849–8860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Z.; Ke, X.; Wang, M.; He, S.; Li, Q.; Zheng, C.; Zhang, Z.; Liu, Y.; Wang, H. Human microRNA hsa-miR-296-5p suppresses enterovirus 71 replication by targeting the viral genome. J. Virol. 2013, 87, 5645–5656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trobaugh, D.W.; Klimstra, W.B. MicroRNA regulation of RNA virus replication and pathogenesis. Trends Mol. Med. 2017, 23, 80–93. [Google Scholar] [CrossRef] [PubMed]
- Ingle, H.; Kumar, S.; Raut, A.A.; Mishra, A.; Kulkarni, D.D.; Kameyama, T.; Takaoka, A.; Akira, S.; Kumar, H. The microRNA miR-485 targets host and influenza virus transcripts to regulate antiviral immunity and restrict viral replication. Sci. Signal. 2015, 8, ra126. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.J.; Yang, J.; Fan, X.L.; Zhao, H.B.; Hu, W.; Li, Z.P.; Yu, G.C.; Ding, X.R.; Wang, J.Z.; Bo, X.C.; et al. Cellular microRNA let-7c inhibits M1 protein expression of the H1N1 influenza A virus in infected human lung epithelial cells. J. Cell. Mol. Med. 2012, 16, 2539–2546. [Google Scholar] [CrossRef]
- Khongnomnan, K.; Makkoch, J.; Poomipak, W.; Poovorawan, Y.; Payungporn, S. Human miR-3145 inhibits influenza A viruses replication by targeting and silencing viral PB1 gene. Exp. Biol. Med. 2015, 240, 1630–1639. [Google Scholar] [CrossRef] [Green Version]
- Jia, X.; Nie, Q.; Zhang, X.; Nolan, L.K.; Lamont, S.J. Novel microRNA involved in host response to avian pathogenic Escherichia coli identified by deep sequencing and integration analysis. Infect. Immun. 2017, 85. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Liu, W.; Xu, H.; Liu, J.; Deng, Y.; Cheng, H.; Zhu, S.; Pei, Y.; Hu, J.; Hu, Z.; et al. MicroRNA expression profiling in Newcastle disease virus-infected DF-1 cells by deep sequencing. Front. Microbiol. 2019, 10, 1659. [Google Scholar] [CrossRef] [Green Version]
- Mu, J.; Liu, X.; Yu, X.; Li, J.; Fei, Y.; Ding, Z.; Yin, R. Cellular microRNA expression profile of chicken macrophages infected with newcastle disease virus vaccine strain LaSota. Pathogens 2019, 8, 123. [Google Scholar] [CrossRef] [Green Version]
- Lian, L.; Qu, L.; Chen, Y.; Lamont, S.J.; Yang, N. A systematic analysis of miRNA transcriptome in Marek’s disease virus-induced lymphoma reveals novel and differentially expressed miRNAs. PLoS ONE 2012, 7, e51003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Brahmakshatriya, V.; Lupiani, B.; Reddy, S.M.; Soibam, B.; Benham, A.L.; Gunaratne, P.; Liu, H.C.; Trakooljul, N.; Ing, N.; et al. Integrated analysis of microRNA expression and mRNA transcriptome in lungs of avian influenza virus infected broilers. BMC Genom. 2012, 13, 278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Brahmakshatriya, V.; Zhu, H.; Lupiani, B.; Reddy, S.M.; Yoon, B.J.; Gunaratne, P.H.; Kim, J.H.; Chen, R.; Wang, J.; et al. Identification of differentially expressed miRNAs in chicken lung and trachea with avian influenza virus infection by a deep sequencing approach. BMC Genom. 2009, 10, 512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szretter, K.J.; Balish, A.L.; Katz, J.M. Influenza: Propagation, quantification, and storage. Curr. Protoc. Microbiol. 2006, 3. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Manual on Animal Influenza Diagnosis and Surveillance, 2nd ed.; WHO Press: Geneva, Switzerland, 2002; pp. 1–94. [Google Scholar]
- Barjesteh, N.; Taha-Abdelaziz, K.; Kulkarni, R.R.; Sharif, S. Innate antiviral responses are induced by TLR3 and TLR4 ligands in chicken tracheal epithelial cells: Communication between epithelial cells and macrophages. Virology 2019, 534, 132–142. [Google Scholar] [CrossRef]
- Hossain, M.J.; Mori, I.; Dong, L.; Liu, B.; Kimura, Y. Fetal calf serum inhibits virus genome expression in Madin-Darby canine kidney cells persistently infected with influenza A virus. Med. Microbiol. Immunol. 2008, 197, 21–27. [Google Scholar] [CrossRef]
- Barjesteh, N.; Behboudi, S.; Brisbin, J.T.; Villanueva, A.I.; Nagy, É.; Sharif, S. TLR ligands induce antiviral responses in chicken macrophages. PLoS ONE 2014, 9, e105713. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Andrews, S. FastQC a quality control tool for high throughput sequence data. Babraham Bioinforma. 2010. [Google Scholar] [CrossRef]
- Ewels, P.; Magnusson, M.; Lundin, S.; Käller, M. MultiQC: Summarize analysis results for multiple tools and samples in a single report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef] [Green Version]
- Griffiths-Jones, S.; Saini, H.K.; Van Dongen, S.; Enright, A.J. MiRBase: Tools for microRNA genomics. Nucleic Acids Res. 2008, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedländer, M.R.; MacKowiak, S.D.; Li, N.; Chen, W.; Rajewsky, N. MiRDeep2 accurately identifies known and hundreds of novel microRNA genes in seven animal clades. Nucleic Acids Res. 2012, 40, 37–52. [Google Scholar] [CrossRef] [PubMed]
- Di Tommaso, P.; Chatzou, M.; Floden, E.W.; Barja, P.P.; Palumbo, E.; Notredame, C. Nextflow enables reproducible computational workflows. Nat. Biotechnol. 2017, 35, 316–319. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2009, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Bache, S.M. Magrittr. 2014. Available online: https://cran.r-project.org/web/packages/magrittr/vignettes/magrittr.html (accessed on 9 April 2020).
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef] [Green Version]
- Lex, A.; Gehlenborg, N.; Strobelt, H.; Vuillemot, R.; Pfister, H. UpSet: Visualization of intersecting sets. IEEE Trans. Vis. Comput. Graph. 2014, 20, 1983–1992. [Google Scholar] [CrossRef]
- Agarwal, V.; Bell, G.W.; Nam, J.-W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. eLife 2015, 4, e05005. [Google Scholar] [CrossRef]
- Wong, N.; Wang, X. MiRDB: An online resource for microRNA target prediction and functional annotations. Nucleic Acids Res. 2015, 43, D146–D152. [Google Scholar] [CrossRef]
- Blatti, C.; Emad, A.; Berry, M.J.; Gatzke, L.; Epstein, M.; Lanier, D.; Rizal, P.; Ge, J.; Liao, X.; Sobh, O.; et al. Knowledge-guided analysis of “omics” data using the KnowEnG cloud platform. PLoS Biol. 2020. [Google Scholar] [CrossRef] [Green Version]
- Mi, H.; Muruganujan, A.; Ebert, D.; Huang, X.; Thomas, P.D. PANTHER version 14: More genomes, a new PANTHER GO-slim and improvements in enrichment analysis tools. Nucleic Acids Res. 2019, 47, D419–D426. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blatti, C.; Sinha, S. Characterizing gene sets using discriminative random walks with restart on heterogeneous biological networks. Bioinformatics 2016, 32, 2167–2175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motulsky, H. Analyzing Data with GraphPad Prism; GraphPad Software, Inc: San Diego, CA, USA, 1999; ISBN 8584573909. [Google Scholar]
- Bao, Y.; Bolotov, P.; Dernovoy, D.; Kiryutin, B.; Zaslavsky, L.; Tatusova, T.; Ostell, J.; Lipman, D. The influenza virus resource at the National Center for Biotechnology Information. J. Virol. 2008, 82, 596–601. [Google Scholar] [CrossRef] [Green Version]
- Simonsen, L.; Bernabe, G.; Lacourciere, K.; Taylor, R.J.; Giovanni, M.Y. The NIAID influenza genome sequencing project. In National Institute of Allergy and Infectious Diseases, NIH; Humana Press: Totowa, NJ, USA, 2008; pp. 109–113. [Google Scholar]
- Hsu, P.W.C. MiRNAMap: Genomic maps of microRNA genes and their target genes in mammalian genomes. Nucleic Acids Res. 2006, 34, D135–D139. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.Y.; Chien, C.H.; Jen, K.H.; Huang, H.D. RegRNA: An integrated web server for identifying regulatory RNA motifs and elements. Nucleic Acids Res. 2006, 34. [Google Scholar] [CrossRef] [Green Version]
- Krüger, J.; Rehmsmeier, M. RNAhybrid: MicroRNA target prediction easy, fast and flexible. Nucleic Acids Res. 2006, 34. [Google Scholar] [CrossRef]
- Lötvall, J.; Hill, A.F.; Hochberg, F.; Buzás, E.I.; Di Vizio, D.; Gardiner, C.; Gho, Y.S.; Kurochkin, I.V.; Mathivanan, S.; Quesenberry, P.; et al. Minimal experimental requirements for definition of extracellular vesicles and their functions: A position statement from the International Society for Extracellular Vesicles. J. Extracell. Vesicles 2014, 3, 328. [Google Scholar] [CrossRef]
- Abcam Datasheet Product Anti-LAMP1 antibody—Lysosome marker ab24170. Available online: https://www.abcam.com/lamp1-antibody-lysosome-marker-ab24170.html (accessed on 3 May 2020).
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of microRNA biogenesis, mechanisms of actions, and circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [Green Version]
- Girardi, E.; López, P.; Pfeffer, S. On the importance of host microRNAs during viral infection. Front. Genet. 2018, 9, 439. [Google Scholar] [CrossRef] [Green Version]
- Li, S.P.; Lin, Z.X.; Jiang, X.Y.; Yu, X.Y. Exosomal cargo-loading and synthetic exosome-mimics as potential therapeutic tools. Acta Pharmacol. Sin. 2018, 39, 542–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hessvik, N.P.; Llorente, A. Current knowledge on exosome biogenesis and release. Cell. Mol. Life Sci. 2018, 75, 193–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, B.; Zhou, J.; Wang, H. Host microRNAs and exosomes that modulate influenza virus infection. Virus Res. 2020, 279, 197885. [Google Scholar] [CrossRef] [PubMed]
- Urbanelli, L.; Buratta, S.; Tancini, B.; Sagini, K.; Delo, F.; Porcellati, S.; Emiliani, C. The role of extracellular vesicles in viral infection and transmission. Vaccines 2019, 7, 102. [Google Scholar] [CrossRef] [Green Version]
- Zhong, L.; Simard, M.J.; Huot, J. Endothelial microRNAs regulating the NF-κB pathway and cell adhesion molecules during inflammation. FASEB J. 2018, 32, 4070–4084. [Google Scholar] [CrossRef] [Green Version]
- Dierkes, R.; Warnking, K.; Liedmann, S.; Seyer, R.; Ludwig, S.; Ehrhardt, C. The Rac1 inhibitor NSC23766 exerts anti-influenza virus properties by affecting the viral polymerase complex activity. PLoS ONE 2014, 9, e88520. [Google Scholar] [CrossRef] [Green Version]
- Burnside, J.; Ouyang, M.; Anderson, A.; Bernberg, E.; Lu, C.; Meyers, B.C.; Green, P.J.; Markis, M.; Isaacs, G.; Huang, E.; et al. Deep sequencing of chicken microRNAs. BMC Genom. 2008, 9, 185. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Huang, S.; Ding, J.; Zhao, Y.; Liang, L.; Liu, T.; Zhan, R.; He, X. Multiple microRNAs modulate p21Cip1/Waf1 expression by directly targeting its 3′ untranslated region. Oncogene 2010, 29, 2302–2308. [Google Scholar] [CrossRef] [Green Version]
- Tsitsiou, E.; Lindsay, M.A. MicroRNAs and the immune response. Curr. Opin. Pharmacol. 2009, 9, 514–520. [Google Scholar] [CrossRef]
- Sun, X.; Icli, B.; Wara, A.K.; Belkin, N.; He, S.; Kobzik, L.; Hunninghake, G.M.; Vera, M.P.; Registry, M.; Blackwell, T.S.; et al. MicroRNA-181b regulates NF-κB-mediated vascular inflammation. J. Clin. Investig. 2012, 122, 1973–1990. [Google Scholar] [CrossRef]
- Xie, W.; Li, M.; Xu, N.; Lv, Q.; Huang, N.; He, J.; Zhang, Y. MiR-181a regulates inflammation responses in monocytes and macrophages. PLoS ONE 2013, 8, e58639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galicia, J.C.; Naqvi, A.R.; Ko, C.-C.; Nares, S.; Khan, A.A. MiRNA-181a regulates Toll-like receptor agonist-induced inflammatory response in human fibroblasts. Genes Immun. 2014, 15, 333–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahanda, M.-L.E.; Ruby, T.; Wittzell, H.; Bed’Hom, B.; Chaussé, A.-M.; Morin, V.; Oudin, A.; Chevalier, C.; Young, J.R.; Zoorob, R. Non-coding RNAs revealed during identification of genes involved in chicken immune responses. Immunogenetics 2009, 61, 55–70. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Fontela, C.; Pazos, M.; Delgado, I.; Murk, W.; Mungamuri, S.K.; Lee, S.W.; García-Sastre, A.; Moran, T.M.; Aaronson, S.A. P53 serves as a host antiviral factor that enhances innate and adaptive immune responses to influenza A virus. J. Immunol. 2011, 187, 6428–6436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Neill, L.A.; Sheedy, F.J.; McCoy, C.E. MicroRNAs: The fine-tuners of Toll-like receptor signalling. Nat. Rev. Immunol. 2011, 11, 163–175. [Google Scholar] [CrossRef]
- Li, P.; Fan, W.; Li, Q.; Wang, J.; Liu, R.; Everaert, N.; Liu, J.; Zhang, Y.; Zheng, M.; Cui, H.; et al. Splenic microRNA expression profiles and integration analyses involved in host responses to Salmonella enteritidis infection in chickens. Front. Cell. Infect. Microbiol. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Taganov, K.D.; Boldin, M.P.; Chang, K.J.; Baltimore, D. NF-κB-dependent induction of microRNA miR-146, an inhibitor targeted to signaling proteins of innate immune responses. Proc. Natl. Acad. Sci. USA 2006. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment Group | MiRNA | Log2 Fold Change | Fold Change | |
|---|---|---|---|---|
| TOC 3 h | TOC AIV 3 h (11 miRNAs) | gga-miR-221-5p | 2.23 | 4.690 |
| gga-miR-425-5p | 0.991 | 1.987 | ||
| gga-miR-210a-5p | 0.956 | 1.940 | ||
| gga-miR-455-5p | 0.904 | 1.871 | ||
| gga-miR-6705-5p | 0.845 | 1.796 | ||
| gga-miR-1608 | 0.806 | 1.749 | ||
| gga-let-7c-5p | 0.786 | 1.724 | ||
| gga-miR-132a-3p | 0.767 | 1.701 | ||
| gga-miR-1434 | 0.751 | 1.683 | ||
| gga-miR-129-5p | 0.701 | 1.625 | ||
| gga-let-7l-5p | 0.629 | 1.547 | ||
| TOC LPS 3 h (3 miRNAs) | gga-miR-6705-5p | 1.087 | 2.124 | |
| gga-miR-1608 | 1.053 | 2.075 | ||
| gga-miR-6704-5p | 1.023 | 2.032 | ||
| TOC polyI:C 3 h (2 miRNAs) | gga-miR-12253-5p | 2.716 | 6.568 | |
| gga-miR-12235-5p | 1.055 | 2.078 | ||
| TOC 18 h | TOC AIV 18 h (3 miRNAs) | gga-miR-1563 | 0.742 | 1.673 |
| gga-miR-1451-5p | 0.722 | 1.649 | ||
| gga-miR-12244-5p | 0.702 | 1.627 | ||
| TOC LPS 18 h (32 miRNAs) | gga-miR-1451-5p | 0.97 | 1.959 | |
| gga-miR-425-5p | 0.948 | 1.929 | ||
| gga-miR-221-5p | 0.887 | 1.849 | ||
| gga-miR-191-5p | 0.882 | 1.843 | ||
| gga-miR-145-5p | 0.858 | 1.813 | ||
| gga-miR-30c-5p | 0.851 | 1.804 | ||
| gga-miR-146b-5p | 0.851 | 1.803 | ||
| gga-miR-15b-5p | 0.81 | 1.753 | ||
| gga-miR-30b-5p | 0.808 | 1.750 | ||
| gga-miR-15c-5p | 0.801 | 1.742 | ||
| gga-miR-210a-5p | 0.795 | 1.735 | ||
| gga-miR-2131-5p | 0.794 | 1.734 | ||
| gga-miR-146a-5p | 0.787 | 1.725 | ||
| gga-miR-2184a-5p | 0.781 | 1.719 | ||
| gga-miR-1563 | 0.777 | 1.713 | ||
| gga-miR-455-5p | 0.769 | 1.704 | ||
| gga-miR-205a | 0.767 | 1.702 | ||
| gga-let-7b | 0.76 | 1.694 | ||
| gga-miR-181b-5p | 0.756 | 1.689 | ||
| gga-miR-30a-5p | 0.753 | 1.685 | ||
| gga-miR-30d | 0.753 | 1.685 | ||
| gga-miR-1729-5p | 0.752 | 1.685 | ||
| gga-miR-16c-5p | 0.725 | 1.653 | ||
| gga-miR-17-5p | 0.725 | 1.653 | ||
| gga-let-7c-5p | 0.708 | 1.634 | ||
| gga-miR-10c-5p | 0.708 | 1.633 | ||
| gga-miR-181a-5p | 0.691 | 1.614 | ||
| gga-miR-18a-5p | 0.685 | 1.608 | ||
| gga-miR-10a-5p | 0.671 | 1.592 | ||
| gga-miR-16-5p | 0.651 | 1.570 | ||
| gga-let-7l-5p | 0.63 | 1.547 | ||
| gga-miR-1559-5p | 0.613 | 1.529 | ||
| TOC polyI:C 18 h (2 miRNAs) | gga-miR-12253-5p | 3.565 | 11.831 | |
| gga-miR-12235-5p | 1.503 | 2.834 | ||
| Treatment Group | MiRNA | Log2 Fold Change | Fold Change | |
|---|---|---|---|---|
| TOC 3 h | TOC AIV 3 h (4 miRNAs) | gga-miR-383-5p | −0.591 | −1.507 |
| gga-miR-1738 | −0.783 | −1.721 | ||
| gga-miR-1692 | −0.861 | −1.816 | ||
| gga-miR-6611-5p | −1.016 | −2.022 | ||
| TOC LPS 3 h (0 miRNAs) | NONE | N/A | N/A | |
| TOC polyI:C 3 h (0 miRNAs) | NONE | N/A | N/A | |
| TOC 18 h | TOC AIV 18 h (2 miRNAs) | gga-miR-12234-5p | −0.61 | −1.526 |
| gga-miR-1793 | −0.629 | −1.546 | ||
| TOC LPS 18 h (15 miRNAs) | gga-miR-6602-5p | −0.583 | −1.498 | |
| gga-miR-1809 | −0.617 | −1.533 | ||
| gga-miR-6632-5p | −0.618 | −1.535 | ||
| gga-miR-6569-5p | −0.654 | −1.574 | ||
| gga-miR-12266-5p | −0.666 | −1.587 | ||
| gga-miR-1783 | −0.682 | −1.604 | ||
| gga-miR-7457-5p | −0.699 | −1.624 | ||
| gga-miR-124a-5p | −0.713 | −1.640 | ||
| gga-miR-2184a-3p | −0.716 | −1.643 | ||
| gga-miR-12228-3p | −0.782 | −1.719 | ||
| gga-miR-1761 | −0.789 | −1.728 | ||
| gga-miR-1795 | −0.853 | −1.807 | ||
| gga-miR-12234-5p | −0.915 | −1.886 | ||
| gga-miR-1701 | −0.92 | −1.893 | ||
| gga-miR-12239-3p | −1.002 | −2.003 | ||
| TOC polyI:C 18 h (40 miRNAs) | gga-miR-1735 | −0.582 | −1.497 | |
| gga-miR-1551-5p | −0.606 | −1.522 | ||
| gga-miR-12228-5p | −0.618 | −1.535 | ||
| gga-miR-6706-5p | −0.626 | −1.543 | ||
| gga-miR-7470-5p | −0.628 | −1.545 | ||
| gga-miR-1692 | −0.633 | −1.551 | ||
| gga-miR-12248-3p | −0.663 | −1.584 | ||
| gga-miR-449a | −0.68 | −1.602 | ||
| gga-miR-1561 | −0.712 | −1.638 | ||
| gga-miR-1656 | −0.713 | −1.639 | ||
| gga-miR-6569-5p | −0.729 | −1.658 | ||
| gga-miR-1465 | −0.741 | −1.672 | ||
| gga-let-7g-5p | −0.783 | −1.721 | ||
| gga-miR-7480-5p | −0.803 | −1.745 | ||
| gga-miR-6563-5p | −0.835 | −1.784 | ||
| gga-let-7f-5p | −0.875 | −1.834 | ||
| gga-miR-196-5p | −0.877 | −1.836 | ||
| gga-miR-12246-5p | −0.888 | −1.851 | ||
| gga-miR-1751-5p | −0.913 | −1.883 | ||
| gga-miR-1671 | −0.964 | −1.951 | ||
| gga-miR-1593 | −0.98 | −1.973 | ||
| gga-miR-365-2-5p | −1.012 | −2.017 | ||
| gga-miR-12208-5p | −1.021 | −2.029 | ||
| gga-miR-499-5p | −1.082 | −2.118 | ||
| gga-miR-1750 | −1.085 | −2.122 | ||
| gga-miR-130c-5p | −1.093 | −2.133 | ||
| gga-miR-7458-5p | −1.093 | −2.133 | ||
| gga-miR-1720-5p | −1.1 | −2.144 | ||
| gga-miR-12228-3p | −1.138 | −2.201 | ||
| gga-miR-1c | −1.146 | −2.213 | ||
| gga-miR-7 | −1.24 | −2.362 | ||
| gga-miR-2184a-3p | −1.248 | −2.375 | ||
| gga-miR-6564-5p | −1.286 | −2.439 | ||
| gga-miR-6611-5p | −1.294 | −2.452 | ||
| gga-miR-190a-5p | −1.367 | −2.580 | ||
| gga-miR-1676-5p | −1.569 | −2.967 | ||
| gga-miR-12248-5p | −1.944 | −3.847 | ||
| gga-miR-6641-5p | −3.127 | −8.733 | ||
| gga-miR-1784b-3p | −3.128 | −8.742 | ||
| gga-miR-216c | −3.426 | −10.751 | ||
| Treatment Group | MiRNA | Log2 Fold Change | Fold Change |
|---|---|---|---|
| EV AIV (21 miRNAs) | gga-miR-301a-5p | 1.525 | 2.877 |
| gga-miR-1563 | 1.221 | 2.332 | |
| gga-miR-122-5p | 0.948 | 1.930 | |
| gga-miR-15b-5p | 0.93 | 1.905 | |
| gga-miR-1452 | 0.903 | 1.870 | |
| gga-miR-194 | 0.894 | 1.858 | |
| gga-miR-1575 | 0.812 | 1.756 | |
| gga-miR-6606-5p | 0.807 | 1.749 | |
| gga-miR-193a-5p | 0.802 | 1.744 | |
| gga-miR-12221-5p | 0.766 | 1.701 | |
| gga-miR-6543-5p | 0.727 | 1.655 | |
| gga-miR-12252-3p | 0.723 | 1.650 | |
| gga-miR-12253-5p | 0.678 | 1.600 | |
| gga-miR-92-5p | 0.672 | 1.594 | |
| gga-miR-1670 | 0.672 | 1.593 | |
| gga-miR-12229-5p | 0.67 | 1.591 | |
| gga-miR-365-1-5p | 0.669 | 1.590 | |
| gga-miR-6708-5p | 0.64 | 1.558 | |
| gga-miR-6616-5p | 0.62 | 1.537 | |
| gga-miR-1637 | 0.618 | 1.534 | |
| gga-miR-383-5p | 0.605 | 1.521 | |
| EV LPS (5 miRNAs) | gga-miR-6697-5p | 0.829 | 1.777 |
| gga-miR-3535 | 0.697 | 1.621 | |
| gga-miR-383-5p | 0.649 | 1.568 | |
| gga-miR-1618-5p | 0.602 | 1.518 | |
| gga-miR-12272-3p | 0.601 | 1.517 | |
| EV polyI:C (14 miRNAs) | gga-miR-12253-5p | 2.822 | 7.070 |
| gga-miR-12235-5p | 1.713 | 3.279 | |
| gga-miR-6593-5p | 1.223 | 2.335 | |
| gga-miR-12290-5p | 0.953 | 1.936 | |
| gga-miR-12252-3p | 0.865 | 1.821 | |
| gga-miR-1397-5p | 0.856 | 1.810 | |
| gga-miR-1777 | 0.74 | 1.671 | |
| gga-miR-1456-5p | 0.734 | 1.663 | |
| gga-miR-6606-5p | 0.714 | 1.640 | |
| gga-miR-7471-5p | 0.677 | 1.599 | |
| gga-miR-1608 | 0.651 | 1.570 | |
| gga-miR-1670 | 0.607 | 1.524 | |
| gga-miR-12295-5p | 0.607 | 1.523 | |
| gga-miR-1649-5p | 0.598 | 1.514 |
| Treatment Group | MiRNA | Log2 Fold Change | Fold Change |
|---|---|---|---|
| EV AIV (57 miRNAs) | gga-miR-3532-5p | −0.612 | −1.528 |
| gga-miR-1722-5p | −0.619 | −1.536 | |
| gga-miR-1677-5p | −0.631 | −1.549 | |
| gga-miR-1661 | −0.634 | −1.552 | |
| gga-miR-1306-5p | −0.636 | −1.554 | |
| gga-miR-2184b-5p | −0.65 | −1.569 | |
| gga-miR-107-5p | −0.661 | −1.581 | |
| gga-miR-6516-5p | −0.667 | −1.588 | |
| gga-miR-1724 | −0.679 | −1.601 | |
| gga-miR-449b-5p | −0.687 | −1.610 | |
| gga-miR-1651-5p | −0.691 | −1.614 | |
| gga-miR-301b-5p | −0.696 | −1.620 | |
| gga-miR-128-1-5p | −0.7 | −1.624 | |
| gga-miR-6671-5p | −0.716 | −1.643 | |
| gga-miR-1715-5p | −0.719 | −1.646 | |
| gga-miR-365b-5p | −0.72 | −1.648 | |
| gga-miR-6675-5p | −0.729 | −1.658 | |
| gga-miR-6590-5p | −0.73 | −1.659 | |
| gga-miR-1648-5p | −0.737 | −1.666 | |
| gga-miR-726-3p | −0.746 | −1.677 | |
| gga-miR-1553-5p | −0.754 | −1.687 | |
| gga-miR-1626-5p | −0.768 | −1.703 | |
| gga-miR-1727 | −0.773 | −1.709 | |
| gga-miR-6550-5p | −0.774 | −1.710 | |
| gga-miR-1664-5p | −0.784 | −1.722 | |
| gga-miR-12254-5p | −0.813 | −1.757 | |
| gga-miR-12269-3p | −0.823 | −1.769 | |
| gga-miR-1784-5p | −0.852 | −1.805 | |
| gga-miR-6639-5p | −0.865 | −1.821 | |
| gga-miR-218-5p | −0.877 | −1.836 | |
| gga-miR-12247-3p | −0.912 | −1.882 | |
| gga-miR-6665-5p | −0.913 | −1.883 | |
| gga-miR-6604-5p | −0.914 | −1.885 | |
| gga-miR-1632-5p | −1.015 | −2.022 | |
| gga-miR-1710 | −1.016 | −2.022 | |
| gga-miR-1730-5p | −1.018 | −2.025 | |
| gga-miR-3536 | −1.047 | −2.066 | |
| gga-miR-12244-5p | −1.078 | −2.112 | |
| gga-miR-1784b-5p | −1.089 | −2.128 | |
| gga-miR-1815 | −1.096 | −2.137 | |
| gga-miR-1464 | −1.096 | −2.137 | |
| gga-miR-1801 | −1.1 | −2.143 | |
| gga-miR-216a | −1.101 | −2.145 | |
| gga-miR-6684-5p | −1.112 | −2.161 | |
| gga-miR-1605 | −1.114 | −2.165 | |
| gga-miR-6596-5p | −1.122 | −2.177 | |
| gga-miR-12284-3p | −1.138 | −2.201 | |
| gga-miR-142-5p | −1.141 | −2.205 | |
| gga-miR-7482-5p | −1.142 | −2.206 | |
| gga-miR-7454-3p | −1.153 | −2.224 | |
| gga-miR-7464-3p | −1.256 | −2.388 | |
| gga-miR-7456-5p | −1.262 | −2.398 | |
| gga-miR-3528 | −1.295 | −2.454 | |
| gga-miR-122b-3p | −1.437 | −2.708 | |
| gga-miR-210a-5p | −1.716 | −3.285 | |
| gga-miR-12260-5p | −1.791 | −3.460 | |
| gga-miR-205b | −1.865 | −3.643 | |
| EV LPS (17 miRNAs) | gga-miR-1464 | −0.597 | −1.512 |
| gga-miR-132b-5p | −0.638 | −1.556 | |
| gga-miR-1584 | −0.674 | −1.595 | |
| gga-miR-211 | −0.689 | −1.612 | |
| gga-miR-6665-5p | −0.716 | −1.643 | |
| gga-miR-1727 | −0.743 | −1.674 | |
| gga-miR-12223-3p | −0.762 | −1.696 | |
| gga-miR-1597-5p | −0.783 | −1.721 | |
| gga-miR-7482-5p | −0.79 | −1.729 | |
| gga-miR-107-5p | −0.797 | −1.737 | |
| gga-miR-449b-5p | −0.835 | −1.783 | |
| gga-miR-6557-5p | −0.869 | −1.826 | |
| gga-miR-12273-5p | −0.895 | −1.859 | |
| gga-miR-210a-5p | −1.276 | −2.421 | |
| gga-miR-12284-3p | −1.28 | −2.428 | |
| gga-miR-205b | −1.383 | −2.608 | |
| gga-miR-1784b-5p | −2.111 | −4.318 | |
| EV polyI:C (90 miRNAs) | gga-miR-1465 | −0.584 | −1.499 |
| gga-miR-6582-5p | −0.589 | −1.504 | |
| gga-miR-1553-5p | −0.597 | −1.512 | |
| gga-miR-1795 | −0.6 | −1.516 | |
| gga-miR-490-5p | −0.603 | −1.519 | |
| gga-miR-6707-5p | −0.604 | −1.520 | |
| gga-miR-1755 | −0.627 | −1.545 | |
| gga-miR-210b-5p | −0.629 | −1.546 | |
| gga-miR-302b-5p | −0.63 | −1.547 | |
| gga-miR-211 | −0.632 | −1.550 | |
| gga-miR-12274-5p | −0.644 | −1.562 | |
| gga-miR-1730-5p | −0.644 | −1.562 | |
| gga-miR-1626-5p | −0.65 | −1.569 | |
| gga-miR-23b-5p | −0.653 | −1.572 | |
| gga-miR-1667-5p | −0.657 | −1.577 | |
| gga-miR-1727 | −0.671 | −1.592 | |
| gga-miR-132b-5p | −0.677 | −1.598 | |
| gga-miR-1802 | −0.677 | −1.598 | |
| gga-miR-12274-3p | −0.678 | −1.600 | |
| gga-miR-1805-5p | −0.719 | −1.646 | |
| gga-miR-449a | −0.726 | −1.654 | |
| gga-miR-1597-5p | −0.729 | −1.658 | |
| gga-miR-204 | −0.734 | −1.663 | |
| gga-miR-726-5p | −0.74 | −1.671 | |
| gga-miR-7444-5p | −0.741 | −1.672 | |
| gga-miR-6679-5p | −0.762 | −1.696 | |
| gga-miR-449b-5p | −0.763 | −1.697 | |
| gga-miR-6596-5p | −0.78 | −1.717 | |
| gga-miR-6567-5p | −0.78 | −1.718 | |
| gga-miR-7451-5p | −0.783 | −1.720 | |
| gga-miR-12266-5p | −0.783 | −1.721 | |
| gga-miR-212-5p | −0.789 | −1.727 | |
| gga-miR-1462-5p | −0.789 | −1.728 | |
| gga-miR-1814 | −0.805 | −1.747 | |
| gga-miR-6550-5p | −0.81 | −1.753 | |
| gga-miR-6516-5p | −0.821 | −1.767 | |
| gga-miR-1690-5p | −0.83 | −1.777 | |
| gga-miR-1663-5p | −0.837 | −1.787 | |
| gga-miR-365b-5p | −0.84 | −1.789 | |
| gga-miR-1598 | −0.842 | −1.792 | |
| gga-miR-301b-5p | −0.851 | −1.804 | |
| gga-miR-6598-5p | −0.882 | −1.843 | |
| gga-miR-12247-3p | −0.883 | −1.844 | |
| gga-miR-1306-5p | −0.888 | −1.851 | |
| gga-miR-6665-5p | −0.892 | −1.855 | |
| gga-miR-6604-5p | −0.899 | −1.865 | |
| gga-miR-6559-5p | −0.91 | −1.879 | |
| gga-miR-218-5p | −0.911 | −1.880 | |
| gga-miR-1715-5p | −0.919 | −1.890 | |
| gga-miR-6669-5p | −0.919 | −1.891 | |
| gga-miR-6671-5p | −0.923 | −1.896 | |
| gga-miR-12223-3p | −0.931 | −1.907 | |
| gga-miR-6566-5p | −0.932 | −1.907 | |
| gga-miR-3536 | −0.964 | −1.951 | |
| gga-miR-216b | −0.966 | −1.954 | |
| gga-miR-1632-5p | −1 | −2.000 | |
| gga-miR-3532-5p | −1.006 | −2.008 | |
| gga-miR-2184b-5p | −1.009 | −2.012 | |
| gga-miR-1658-5p | −1.027 | −2.037 | |
| gga-miR-216a | −1.056 | −2.080 | |
| gga-miR-7482-5p | −1.066 | −2.093 | |
| gga-miR-1638 | −1.072 | −2.102 | |
| gga-miR-449d-5p | −1.076 | −2.108 | |
| gga-miR-6639-5p | −1.079 | −2.112 | |
| gga-miR-12269-3p | −1.081 | −2.116 | |
| gga-miR-1722-5p | −1.095 | −2.136 | |
| gga-miR-7456-5p | −1.132 | −2.191 | |
| gga-miR-6675-5p | −1.14 | −2.204 | |
| gga-miR-1605 | −1.155 | −2.227 | |
| gga-miR-12209-3p | −1.169 | −2.248 | |
| gga-miR-12279-3p | −1.176 | −2.260 | |
| gga-miR-12260-5p | −1.199 | −2.295 | |
| gga-miR-7479-5p | −1.207 | −2.309 | |
| gga-miR-1464 | −1.214 | −2.320 | |
| gga-miR-12219-3p | −1.23 | −2.346 | |
| gga-miR-12284-3p | −1.234 | −2.352 | |
| gga-miR-1775-5p | −1.259 | −2.393 | |
| gga-miR-1573 | −1.273 | −2.417 | |
| gga-miR-219a | −1.281 | −2.429 | |
| gga-miR-7473-5p | −1.308 | −2.477 | |
| gga-miR-12254-5p | −1.406 | −2.651 | |
| gga-miR-142-5p | −1.413 | −2.663 | |
| gga-miR-1677-5p | −1.418 | −2.673 | |
| gga-miR-30b-5p | −1.448 | −2.728 | |
| gga-miR-210a-5p | −1.47 | −2.770 | |
| gga-miR-19a-5p | −1.504 | −2.837 | |
| gga-miR-7454-3p | −1.571 | −2.970 | |
| gga-miR-1784b-5p | −1.654 | −3.148 | |
| gga-miR-107-5p | −1.689 | −3.224 | |
| gga-miR-205b | −2.287 | −4.879 |
| Treatment Group | Total DE | Up-Regulated | Down-Regulated | ||||||
|---|---|---|---|---|---|---|---|---|---|
| miRNAs | Target Genes | Target Pathways | miRNAs | Target Genes | Target Pathways | miRNAs | Target Genes | Target Pathways | |
| TOC 3 h AIV | 15 | 105 | 3 | 11 | 94 | 1 | 4 | 11 | 2 |
| TOC 3 h LPS | 3 | 34 | 4 | 3 | 34 | 4 | 0 | N/A | N/A |
| TOC 3 h polyI:C | 2 | 151 | 14 | 2 | 151 | 14 | 0 | N/A | N/A |
| TOC 18 h AIV | 5 | 25 | 17 | 3 | 17 | 2 | 2 | 8 | 15 |
| TOC 18 h LPS | 47 | 578 | 14 | 32 | 447 | 10 | 15 | 112 | 4 |
| TOC 18 h polyI:C | 42 | 384 | 17 | 2 | 146 | 13 | 40 | 233 | 5 |
| EV AIV | 78 | 487 | 16 | 21 | 119 | 9 | 57 | 346 | 9 |
| EV LPS | 22 | 147 | 11 | 5 | 16 | 3 | 17 | 131 | 8 |
| EV polyI:C | 104 | 817 | 23 | 14 | 167 | 17 | 90 | 628 | 9 |
| Treatment Group | Pathway | miRNA(s) | |
|---|---|---|---|
| TOC 3 h | TOC 3 h AIV | mRNA processing | gga-let-7c-5p, gga-let-7l-5p, gga-miR-129-5p |
| TOC 3 h LPS | BDNF signaling pathway | gga-miR-6704-5p | |
| Focal Adhesion-PI3K-Akt-mTOR-signaling pathway | gga-miR-6704-5p | ||
| Splicing factor NOVA regulated synaptic proteins | gga-miR-1608 | ||
| Synaptic vesicle pathway | gga-miR-6704-5p | ||
| TOC 3 h polyI:C | Imatinib resistance in chronic myeloid leukemia | gga-miR-12235-5p | |
| PluriNetWork | gga-miR-12235-5p | ||
| Calcium regulation in the cardiac cell | gga-miR-12235-5p | ||
| SIDS susceptibility pathways | gga-miR-12235-5p | ||
| Gastric cancer network 1 | gga-miR-12235-5p | ||
| Stabilization and expansion of the E-cadherin adherens junction | gga-miR-12235-5p | ||
| Fanconi anemia pathway | gga-miR-12235-5p | ||
| Calcium regulation in the cardiac cell | gga-miR-12235-5p | ||
| Focal adhesion-PI3K-Akt-mTOR-signaling pathway | gga-miR-12235-5p | ||
| Glial cell differentiation | gga-miR-12235-5p | ||
| Validated nuclear estrogen receptor alpha network | gga-miR-12235-5p | ||
| Visual signal transduction: cones | gga-miR-12235-5p | ||
| Integrin-mediated cell adhesion | gga-miR-12235-5p | ||
| Synaptic vesicle pathway | gga-miR-12235-5p | ||
| TOC 18 h | TOC 18 h AIV | Regulation of Toll-like receptor signaling pathway | gga-miR-12244-5p |
| Notch signaling pathway | gga-miR-12244-5p | ||
| TOC 18 h LPS | Adipogenesis | gga-miR-15b-5p, gga-miR-15c-5p, gga-miR-16-5p, gga-miR-16c-5p, gga-miR-2184a-5p | |
| EGF/EGFR signaling pathway | gga-miR-145-5p | ||
| Gastric cancer network 1 | gga-miR-181a-5p, gga-miR-181b-5p | ||
| Insulin signaling | gga-miR-15b-5p, gga-miR-15c-5p, gga-miR-1563, gga-miR-16-5p, gga-miR-16c-5p | ||
| Regulation of nuclear beta catenin signaling and target gene transcription | gga-miR-145-5p | ||
| Regulation of RAC1 activity | gga-miR-205a | ||
| Senescence and autophagy in cancer | gga-miR-17-5p | ||
| TarBasePathway | gga-miR-30a-5p, gga-miR-30b-5p, gga-miR-30c-5p, gga-miR-30d, gga-miR-455-5p | ||
| TGF-beta signaling pathway | gga-miR-15b-5p, gga-miR-15c-5p, gga-miR-16-5p, gga-miR-16c-5p | ||
| XPodNet - protein-protein interactions in the podocyte expanded by STRING | gga-miR-145-5p, gga-miR-1563, gga-miR-17-5p, gga-miR-205a, gga-miR-30a-5p, gga-miR-30b-5p, gga-miR-30c-5p, gga-miR-30d | ||
| TOC 18 h polyI:C | Imatinib resistance in chronic myeloid leukemia | gga-miR-12235-5p | |
| PluriNetWork | gga-miR-12235-5p | ||
| Calcium regulation in the cardiac cell | gga-miR-12235-5p | ||
| SIDS susceptibility pathways | gga-miR-12235-5p | ||
| Gastric cancer network 1 | gga-miR-12235-5p | ||
| Stabilization and expansion of the E-cadherin adherens junction | gga-miR-12235-5p | ||
| Fanconi anemia pathway | gga-miR-12235-5p | ||
| Calcium regulation in the cardiac cell | gga-miR-12235-5p | ||
| Focal adhesion-PI3K-Akt-mTOR-signaling pathway | gga-miR-12235-5p | ||
| Glial cell differentiation | gga-miR-12235-5p | ||
| Validated nuclear estrogen receptor alpha network | gga-miR-12235-5p | ||
| Visual signal transduction: cones | gga-miR-12235-5p | ||
| Synaptic vesicle pathway | gga-miR-12235-5p | ||
| Treatment Group | Pathway | miRNA(s) | |
|---|---|---|---|
| TOC 3 h | TOC 3 h AIV | Ectoderm differentiation | gga-miR-6611-5p |
| Validated targets of C-MYC transcriptional activation | gga-miR-383-5p | ||
| TOC 3 h LPS | N/A | N/A | |
| TOC 3 h polyI:C | N/A | N/A | |
| TOC 18 h | TOC 18 h AIV | Insulin Signaling | gga-miR-1793 |
| EGF/EGFR signaling pathway | gga-miR-1793 | ||
| ErbB1 downstream signaling | gga-miR-1793 | ||
| EGFR1 signaling pathway | gga-miR-1793 | ||
| TNF-alpha NF-kB signaling pathway | gga-miR-1793 | ||
| p38 MAPK signaling pathway | gga-miR-1793 | ||
| MAPK signaling pathway | gga-miR-1793 | ||
| Trk receptor signaling mediated by the MAPK pathway | gga-miR-1793 | ||
| Signaling mediated by p38-alpha and p38-beta | gga-miR-1793 | ||
| Serotonin receptor 4/6/7 and NR3C signaling | gga-miR-1793 | ||
| Structural pathway of interleukin 1 (IL-1) | gga-miR-1793 | ||
| LPA4-mediated signaling events | gga-miR-1793 | ||
| Interferon type I signaling pathways | gga-miR-1793 | ||
| Bladder cancer | gga-miR-1793 | ||
| BDNF signaling pathway | gga-miR-1793 | ||
| TOC 18 h LPS | Circadian rhythm related genes | gga-miR-12239-3p, gga-miR-124a-5p | |
| p53 signaling | gga-miR-124a-5p, gga-miR-1783 | ||
| PluriNetWork | gga-miR-124a-5p | ||
| TNF-alpha NF-kB signaling pathway | gga-miR-7457-5p | ||
| TOC 18 h polyI:C | Ectoderm differentiation | gga-miR-1c, gga-miR-6611-5p, gga-miR-6706-5p | |
| Integrated breast cancer pathway | gga-miR-1c, gga-miR-130c-5p | ||
| mRNA processing | gga-miR-12248-5p, gga-miR-1465, gga-miR-7, gga-let-7f-5p, gga-let-7g-5p | ||
| PluriNetWork | gga-miR-1551-5p, gga-miR-449a | ||
| TNF-alpha NF-kB signaling pathway | gga-miR-6641-5p, gga-miR-7480-5p | ||
| Treatment Group | Pathway | miRNA(s) |
|---|---|---|
| EV AIV | FOXA1 transcription factor network | gga-miR-15b-5p |
| GPCRs, Class A Rhodopsin-like | gga-miR-6616-5p | |
| Insulin signaling | gga-miR-1563, gga-miR-15b-5p | |
| mir-124 predicted interactions with cell cycle and differentiation | gga-miR-92-5p | |
| mRNA processing | gga-miR-15b-5p, gga-miR-1452, gga-miR-6543-5p | |
| p73 transcription factor network | gga-miR-194 | |
| TGF-beta signaling pathway | gga-miR-15b-5p | |
| Validated targets of C-MYC transcriptional activation | gga-miR-383-5p | |
| Validated targets of C-MYC transcriptional repression | gga-miR-6708-5p | |
| EV LPS | Matrix Metalloproteinases | gga-miR-12272-3p |
| mRNA Processing | gga-miR-12272-3p | |
| Validated targets of C-MYC transcriptional activation | gga-miR-383-5p | |
| EV polyI:C | Calcium regulation in the cardiac cell | gga-miR-12235-5p |
| Fanconi anemia pathway | gga-miR-12235-5p | |
| Focal adhesion-PI3K-Akt-mTOR-signaling pathway | gga-miR-12235-5p | |
| Gastric cancer network 1 | gga-miR-12235-5p | |
| Glial cell differentiation | gga-miR-12235-5p | |
| Imatinib resistance in chronic myeloid leukemia | gga-miR-12235-5p | |
| PluriNetWork | gga-miR-12235-5p | |
| SIDS Susceptibility pathways | gga-miR-12235-5p | |
| Splicing factor NOVA regulated synaptic proteins | gga-miR-1608 | |
| Stabilization and expansion of the E-cadherin adherens junction | gga-miR-12235-5p | |
| Synaptic vesicle pathway | gga-miR-12235-5p | |
| Validated nuclear estrogen receptor alpha network | gga-miR-12235-5p | |
| Visual signal transduction: cones | gga-miR-12235-5p | |
| XPodNet - protein-protein interactions in the podocyte expanded by STRING | gga-miR-1456-5p |
| Treatment Group | Pathway | miRNA(s) |
|---|---|---|
| EV AIV | Caspase cascade in apoptosis | gga-miR-1784b-5p, gga-miR-3536 |
| Direct p53 effectors | gga-miR-12247-3p, gga-miR-12284-3p, gga-miR-205b, gga-miR-142-5p | |
| Imatinib resistance in chronic myeloid leukemia | gga-miR-1724, gga-miR-6639-5p | |
| Regulation of RAC1 activity | gga-miR-205b, gga-miR-449b-5p | |
| Splicing factor NOVA regulated synaptic proteins | gga-miR-7456-5p | |
| Synaptic vesicle pathway | gga-miR-1632-5p, gga-miR-3532-5p, gga-miR-6639-5p | |
| TGF-beta signaling pathway | gga-miR-1632-5p, gga-miR-142-5p, gga-miR-1727 | |
| Validated targets of C-MYC transcriptional repression | gga-miR-12247-3p, gga-miR-1626-5p | |
| XPodNet - protein-protein interactions in the podocyte expanded by STRING | gga-miR-301b-5p, gga-miR-1632-5p, gga-miR-6639-5p, gga-miR-205b, gga-miR-218-5p, gga-miR-107-5p | |
| EV LPS | Caspase Cascade in apoptosis | gga-miR-1784b-5p |
| Direct p53 effectors | gga-miR-12284-3p, gga-miR-205b | |
| Globo sphingolipid metabolism | gga-miR-1597-5p, gga-miR-211 | |
| PluriNetWork | gga-miR-449b-5p | |
| PodNet: protein-protein interactions in the podocyte | gga-miR-107-5p, gga-miR-205b | |
| Regulation of RAC1 activity | gga-miR-205b, gga-miR-449b-5p | |
| Stabilization and expansion of the E-cadherin adherens junction | gga-miR-211 | |
| XPodNet - protein-protein interactions in the podocyte expanded by STRING | gga-miR-107-5p, gga-miR-205b, gga-miR-211 | |
| EV polyI:C | BMP receptor signaling | gga-miR-1677-5p, gga-miR-490-5p, gga-miR-7454-3p |
| Circadian rhythm related genes | gga-miR-1632-5p, gga-miR-1755, gga-miR-218-5p, gga-miR-365b-5p, gga-miR-7482-5p | |
| Direct p53 effectors | gga-miR-12247-3p, gga-miR-12284-3p, gga-miR-142-5p, gga-miR-205b, gga-miR-219a | |
| mRNA processing | gga-miR-1465, gga-miR-1638, gga-miR-1663-5p, gga-miR-205b, gga-miR-6598-5p, gga-miR-726-5p | |
| PodNet: protein-protein interactions in the podocyte | gga-miR-107-5p, gga-miR-1632-5p, gga-miR-1658-5p, gga-miR-205b, gga-miR-219a, gga-miR-301b-5p | |
| Regulation of RAC1 activity | gga-miR-205b, gga-miR-449a, gga-miR-449b-5p, gga-miR-449d-5p, gga-miR-7451-5p | |
| Splicing factor NOVA regulated synaptic proteins | gga-miR-30b-5p, gga-miR-302b-5p, gga-miR-7456-5p | |
| Synaptic vesicle pathway | gga-miR-132b-5p, gga-miR-1632-5p, gga-miR-3532-5p, gga-miR-6639-5p, gga-miR-6669-5p | |
| XPodNet – protein-protein interactions in the podocyte expanded by STRING | Gga-miR-107-5p, gga-miR-1632-5p, gga-miR-1658-5p, gga-miR-204, gga-miR-205b, gga-miR-211, gga-miR-218-5p, gga-miR-219a, gga-miR-30b-5p, gga-miR-301b-5p, gga-miR-6598-5p, gga-miR-6639-5p, gga-miR-6669-5p |
| Segment | Protein(s) | miRNA | Position | miRanda score | miRanda Free Energy (kcal/mol) | Expression |
|---|---|---|---|---|---|---|
| Segment 1 | PB2 | gga-miR-122b-3p | 1207–1227 | 170 | −17.54 | Downregulated in EV AIV |
| gga-miR-146a-5p | 1896–1917 | 161 | −17.51 | Upregulated in TOC LPS 18 h | ||
| gga-miR-146b-5p | 1896–1917 | 161 | −17.87 | Upregulated in TOC LPS 18 h | ||
| gga-miR-1720-5p | 1249–1268 | 177 | −32.16 | Downregulated in TOC polyI:C 18 h | ||
| gga-miR-6671-5p | 578–601 | 161 | −22.53 | Downregulated in EV AIV & EV polyI:C | ||
| Segment 2 | PB1, PB1-F2 | gga-miR-107-5p | 774–796 | 163 | −20.95 | Downregulated in EV AIV, EV LPS & EV polyI:C |
| gga-miR-12223-3p | 1328–1349 | 160 | −27.23 | Downregulated in EV LPS & EV polyI:C | ||
| gga-miR-129-5p | 2264–2284 | 179 | −24.92 | Upregulated in TOC AIV 3h | ||
| gga-miR-132b-5p | 91–111 | 160 | −20.56 | Downregulated in EV LPS & EV polyI:C | ||
| gga-miR-1661 | 1822–1844 | 163 | −26.29 | Downregulated in EV AIV | ||
| gga-miR-6641-5p | 1729–1750 | 176 | −16.88 | Downregulated in TOC polyI:C 18 h | ||
| Segment 3 | PA | gga-miR-1573 | 1027–1047 | 166 | −19.85 | Downregulated in EV polyI:C |
| gga-miR-1663-5p | 266–285 | 162 | −26.14 | Downregulated in EV polyI:C | ||
| gga-miR-1715-5p | 1439–1463 | 161 | −24.17 | Downregulated in EV AIV & EV polyI:C | ||
| gga-miR-6665-5p | 2005–2025 | 162 | −20.58 | Downregulated in EV AIV, EV LPS & EV polyI:C | ||
| gga-miR-7454-3p | 944–968 | 160 | −19.88 | Downregulated in EV AIV & EV polyI:C | ||
| Segment 4 | HA | gga-miR-1593 | 500–520 | 161 | −21.16 | Downregulated in TOC polyI:C 18 h |
| gga-miR-1605 | 55–76 | 168 | −20.89 | Downregulated in EV AIV & EV polyI:C | ||
| gga-miR-1671 | 761–783 | 161 | −24.1 | Downregulated in TOC polyI:C 18 h | ||
| Segment 5 | NP | gga-miR-12269-3p | 67–88 | 160 | −22.67 | Downregulated in EV AIV & EV polyI:C |
| gga-miR-145-5p | 291–313 | 167 | −22.82 | Upregulated in TOC LPS 18 h | ||
| gga-miR-1784b-5p | 1163–1186 | 163 | −21.01 | Downregulated in EV AIV, EV LPS & EV polyI:C | ||
| gga-miR-6679-5p | 786–809 | 178 | −20.29 | Downregulated in EV polyI:C | ||
| Segment 6 | NA | gga-miR-1783 | 455–477 | 163 | −20.22 | Downregulated in TOC LPS 18 h |
| gga-miR-218-5p | 535–553 | 160 | −23.78 | Downregulated in EV AIV & EV polyI:C | ||
| Segment 7 | M1, M2 | gga-miR-1710 | 569–590 | 164 | −19.87 | Downregulated in EV AIV |
| gga-miR-1784b-5p | 565–588 | 161 | −18.81 | Downregulated in EV AIV, EV LPS & EV polyI:C | ||
| Segment 8 | NS1, NEP | NONE | N/A | N/A | N/A | N/A |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
O’Dowd, K.; Emam, M.; El Khili, M.R.; Emad, A.; Ibeagha-Awemu, E.M.; Gagnon, C.A.; Barjesteh, N. Distinct miRNA Profile of Cellular and Extracellular Vesicles Released from Chicken Tracheal Cells Following Avian Influenza Virus Infection. Vaccines 2020, 8, 438. https://doi.org/10.3390/vaccines8030438
O’Dowd K, Emam M, El Khili MR, Emad A, Ibeagha-Awemu EM, Gagnon CA, Barjesteh N. Distinct miRNA Profile of Cellular and Extracellular Vesicles Released from Chicken Tracheal Cells Following Avian Influenza Virus Infection. Vaccines. 2020; 8(3):438. https://doi.org/10.3390/vaccines8030438
Chicago/Turabian StyleO’Dowd, Kelsey, Mehdi Emam, Mohamed Reda El Khili, Amin Emad, Eveline M. Ibeagha-Awemu, Carl A. Gagnon, and Neda Barjesteh. 2020. "Distinct miRNA Profile of Cellular and Extracellular Vesicles Released from Chicken Tracheal Cells Following Avian Influenza Virus Infection" Vaccines 8, no. 3: 438. https://doi.org/10.3390/vaccines8030438