Anti-Leishmanial Vaccines: Assumptions, Approaches, and Annulments
by
 , , ,
, , ,
Shubhranshu Zutshi
1 ,
,
Sunil Kumar
1,
Prashant Chauhan
1,
Yashwant Bansode
1,
Arathi Nair
1,
Somenath Roy
2,
Arup Sarkar
3 and
Bhaskar Saha
1,3,* 1
National Centre for Cell Science, Ganeshkhind, Pune 411007, India
2
Department of Human Physiology with Community Health, Vidyasagar University, Midnapore 721102, India
3
Department of Biotechnology, Trident Academy of Creative Technology, Bhubaneswar 751024, India
*
Author to whom correspondence should be addressed.
Vaccines 2019, 7(4), 156; https://doi.org/10.3390/vaccines7040156
Submission received: 27 August 2019
/
Revised: 24 September 2019
/
Accepted: 8 October 2019
/
Published: 18 October 2019
Abstract
:Leishmaniasis is a neglected protozoan parasitic disease that occurs in 88 countries but a vaccine is unavailable. Vaccination with live, killed, attenuated (physically or genetically) Leishmania have met with limited success, while peptide-, protein-, or DNA-based vaccines showed promise only in animal models. Here, we critically assess several technical issues in vaccination and expectation of a host-protective immune response. Several studies showed that antigen presentation during priming and triggering of the same cells in infected condition are not comparable. Altered proteolytic processing, antigen presentation, protease-susceptible sites, and intracellular expression of pathogenic proteins during Leishmania infection may vary dominant epitope selection, MHC-II/peptide affinity, and may deter the reactivation of desired antigen-specific T cells generated during priming. The robustness of the memory T cells and their functions remains a concern. Presentation of the antigens by Leishmania-infected macrophages to antigen-specific memory T cells may lead to change in the T cells’ functional phenotype or anergy or apoptosis. Although cells may be activated, the peptides generated during infection may be different and cross-reactive to the priming peptides. Such altered peptide ligands may lead to suppression of otherwise active antigen-specific T cells. We critically assess these different immunological issues that led to the non-availability of a vaccine for human use.
1. Introduction
The earliest vaccination protocols by Edward Jenner, Louis Pasteur, and Robert Koch demonstrated that killed, live-attenuated, or xenogenic microorganisms induced host-protective immunity [1]. During the next hundred years, the modus operandi of the immune system and the vaccine-induced protective immunity has been gradually deciphered and the central dogma of vaccination has been framed. It holds that an immunogen is capable of inducing long-lived immunological memory and protective immunity against rechallenge with the same pathogen [2]. Although this concept has been successful in generating potent vaccines for many deadly diseases, leishmaniasis caused by the protozoan parasite Leishmania remains a threatening exception.
As Jose Esparaza described in the context of HIV vaccine failures, a paradigm shift from the current concept of vaccinology is essential [3]. The seven decades of failures in developing an anti-leishmanial vaccine fit this proposition very well for an urgent relook at the host–pathogen interaction dynamics [2,4]. Here, we follow the scheme of immune priming, reactivation, and outcome of challenge infection (Figure 1), emphasizing the immune response parameters that are responsible for the failures.
2. Factors in Antigenic Priming That Affect Vaccination Efficacy
2.1. Selection of an Infective-Stage Specific Vaccine Candidate
Leishmanization with live Leishmania and whole-parasite-based vaccines against Leishmania used promastigotes of different Leishmania species as a form of the vaccine but the associated risks and disadvantages prompted developing new vaccination modalities such as protein- or DNA-priming-based vaccines [5]. Due to ease in culture and characterization of promastigotes, early subunit vaccination studies primarily focused on promastigote antigens, e.g., gp63, gp46, LACK, and promastigote surface antigen-2 (PSA-2) [6,7]. As Leishmania amastigotes propagate in humans [8] and as its proteome is available for degradation and presentation by antigen-presenting cells (APCs), an anti-amastigote immune response is vital for the maintenance of long-term immunity. Immunomodulatory Th2 properties of sand fly saliva [9,10,11] prompted the inclusion of genes for sand fly salivary proteins in the vaccine construct. Polarization of Th subsets to Th1 and Th2 in Leishmania-resistant C57BL/6 and Leishmania-susceptible BALB/c mice, respectively, indicates that the Leishmania antigen-specific Th1-clones at later stages of infection, via amastigote degradation, may significantly contribute to protection and disease progression. Many amastigote-specific antigens including A2 protein from Leishmania donovani [12], hydrophilic acylated surface protein B1 (HASPB1) of L. major [13] and L. donovani [14], p27-/-, and LdCen-/-Leishmania [15,16] were discovered as probable vaccine candidates. Table 1 presents a comprehensive compilation of the vaccine approaches and analyses.
2.2. Determinant Selection as a Function of Relative Abundance of Leishmanial Proteins
Availability of an intracellular protein for protease-dependent degradation depends on its abundance and expression inside the APCs. Analysis of 2.5% of the most abundantly expressed mRNAs showed that nearly 41% and 11% of encoded proteins from these mRNA’s contained MHC-I and MHC-II ligands, respectively [115]. High-throughput MS-based analyses of 22,000 HLA-binding peptides across cancer cell lines and primary cells identified a significant positive correlation between a protein’s expression level and its presentation though HLA, where a faster degradation of overexpressed proteins was found to be in part responsible for their preferred presentation to MHC molecules [116]. Therefore, the antigen-specific recall response generated during the infection would depend upon the intracellular expression level of the immunogenic stage-specific protein used for priming, as it may be outnumbered by increased turnover of other proteins in infected macrophages affecting their loading onto MHC-II molecules. Intracellular competition between the potential determinants would thus play a key role in the expansion of the antigen-primed host-protective memory T cells during infection.
2.3. Intracellular Compartmentalization Contributes to the Processing of an Antigen
Even if Leishmania-infected macrophages form phagolysosomes, the availability of antigen for protease-dependent processing would depend on its subcellular localization in Leishmania. Leishmania cysteine protease-specific Th cell lines were better activated by macrophages containing inactivated or killed parasites [117]. Macrophages containing live parasites ectopically expressing Leishmanial membrane-bound acid phosphatases (MAPs) either on parasite’s surface or insoluble form were able to better activate T cells; however, wild-type MAP and cysteine proteases expressing Leishmania-infected macrophages were unable to activate T cells [118]. Amastigotes are able to sequester and protect the protein antigens from uptake and degradation by MHC-II presentation pathway, possibly by modification of cellular trafficking pathways [119]. Splenic DCs efficiently presented and elicited immune responses to surface localized OVA rather than cytoplasmic form when infected with OVA-expressing transgenic Leishmania [120]. Antigen localization plays a crucial role in its uptake and presentation via MHC-II in the altered physiological environment of Leishmania-infected macrophages for the induction of immune responses and reactivation of the antigen-primed memory T cells during exposure to live virulent Leishmania.
3. Leishmania Targeting of Antigen Processing and Presentation Affects Vaccination Efficacy
3.1. Receptor-Mediated Internalization of Leishmania Parasites
Receptor-mediated endocytosis of Leishmania by macrophages involves numerous receptors, e.g., complement receptor 1 (CR1), CR3, Fc-gamma receptors (FCγR), and fibronectin receptors (FnRs), which assist interaction or docking of parasites on macrophage surface [121,122]. The complement components are endogenous adjuvants for vaccine-induced CD8+ T cell responses in Leishmania infection [123]. Due to their phenotypic plasticity, M1 and M2 macrophages change phenotypes during Leishmania infection [124]. As lipophosphoglycan (LPG)-dependent TLR2 activation during Leishmania infection causes M1/M2 polarization of macrophages altering cytokines stoichiometry, macrophage plasticity is attributed to cytokines; particularly, IFN-γ promoted M1 and IL-4, IL-13 driven M2 phenotypes (Figure 2).
M1 macrophages are potent producers of reactive oxygen species (ROS), inducible nitric oxide synthase (iNOS), and reactive nitro-species, and also function as effective APCs, secreting high levels of IL-12 and IL-23 [125]. These observations imply that Fc glycosylation and FcR/CR interactions during priming may be manipulated to establish robust macrophage polarization.
3.2. The Hijacking of Lysosomal Fusion Machinery inside Macrophages
Phagolysosome biogenesis is the major defense mechanism of immune cells from invading pathogens. Synaptotagmin (Syt) Type-1 membrane protein regulates vesicular fusion processes such as exocytosis and phagocytosis. While SytV is a major regulator of phagolysosome biogenesis, SytXI is involved in secretion from Leishmania-infected macrophages in gp63-dependent manner [126]. SytV silencing inhibits the recruitment of cathepsin D and vesicular proton ATPase to endosomal membrane. LPG binds to phagosome membrane microdomains and dissociates SytV hampering the acidification process, phagolysosome formation, and aiding parasite survival [127]. A host-protective role of transcriptional regulator STAT1 in maintaining counter ion conductivity by chloride ion channels, which are important for phagosomal acidification, and macrophage proteasomal system, are reported [128,129]. Hence, phagosomal maturation is a step, which Leishmania targets initially to survive inside macrophages until the time it transforms into acid-resistant amastigote form, inhibiting the antigen presentation process and affecting the vaccine-primed antigen-specific T cells’ reactivation (Figure 3).
3.3. Altered Activity of Proteases
Leishmania infection tweaks the proteasomal machinery to increase the degradation of transcription factor STAT-1α and reduces the release of interferons [128]. Lysosomal proteases like cathepsin generate high-affinity peptides for efficient binding to MHC [130]. Due to their selectivity in cleavage sites, selective inhibition of these proteases during infection can change the type of peptide generated in APCs. While IFN-γ stimulation increased cathepsin D activity, LPS stimulation had the opposite effect on cathepsin D [131]. As LPS works through TLR4, TLR4 activation by Leishmania [132] may alter cathepsin S activity by LPS and IFN-γ mediated alteration in cathepsin S expression [133]. The repertoire of T cells generated during priming and natural infection by the same antigen may thus change due to parasite-induced selective interference in proteolytic processing of antigens.
3.4. Degradation of Antigen Presentation Machinery
During early MHC biosynthesis, I chain is attached to MHC-II αβ dimer to prevent premature peptide loading. Proteolysis and acidic pH induce I chain release and replacement with CLIP (class II-associated invariant chain peptide). When MHC-II is ready for antigen loading in mature or late endosomes, CLIP is dissociated from MHC-II via another glycoprotein DM that induces a conformational change in CLIP molecule [134]. Both MHC-II and H2-DM molecules are taken up by amastigotes within parasitophorous vacuoles resulting in degradation of MHC-II and associated molecules [135], and hindering the antigen presentation to T helper cells affecting their activation and differentiation into cytokine-secreting effectors.
3.5. Immunodominance and Epitope Crypticity: Importance in Anti-Leishmanial Vaccination
In spite of the above-mentioned problems with antigen processing, the selection of the priming epitope may face altered immunodominance during Leishmania infection. Immunodominance refers to the “preferred” or “biased” selection of a few epitopes of a protein or from a whole proteome for MHC-I/II presentation, during the generation of an anti-pathogenic immune response. An immunodominant epitope elicits a higher response when compared to a subdominant epitope. The epitopes that are not “favored” for binding to MHC molecules are termed as cryptic epitopes. There are many factors that can influence the dominance or crypticity of a particular T cell epitope: (1) TCR recognition sites or intrinsic nature of antigen, (2) peptide-MHC-II affinity, (3) abundance of particular antigen, (4) promiscuous binding of antigen, and (5) MHC-guided processing and the flanking regions of peptides [136,137]. Vaccination with LACK antigen from L. major causes a CD8+ T cell-dependent protection against challenge with L. major [78,138,139]. The expression of Pgp-1 glycoprotein enriched antigen-stimulated memory T cells within the periphery [140] but their sustenance requires persistent antigenic stimulation [141]. Thus, the antigenic crypticity or shifting immunodominance during antigen presentation in Leishmania infection plays important roles in vaccine outcome.
3.6. Loading of Antigen to MHC Molecule
L. donovani-infected THP-1 macrophages have a different HLA-I self-peptide repertoire [142] implying that Leishmania infection may affect the antigen-specific T cell repertoire. The incapability of Leishmania-infected macrophages to activate Th cell was reversed by low pH or 2-(1-adamantyl)-ethanol treatment altering intracellular conditions favoring peptide exchange and a significant increase in anti-Leishmanial T cell compartment, suggesting binding of incognate peptides hindered the loading of Leishmanial peptides during the course of infection [143]. Hence, a perturbed MHC compartment inside the Leishmania-infected macrophages can lead to the inefficient presentation of anti-leishmanial peptides (Figure 3).
3.7. Peptide-MHC-II Affinity: A Major Determinant of Immunodominant Epitope Selection
The most significant factor for the competent presentation of a foreign peptide to a Naïve T cell and immunodominance of the selected peptide [144] is a peptide’s affinity for the peptide-binding groove of an MHC molecule. Importance of a single residue in the binding potential of an antigenic peptide was described where substitutions in S. aureus nuclease epitopes can convert immunodominant epitopes into subdominant ones and vice-versa, and this principle is applied to cancer therapy using Pseudomonas exotoxin-A [145,146,147]. These findings have significant implications in vaccine development due to following reasons: (1) Change in amino acid residues of flanking regions of epitopes may render them less susceptible to action of proteases and alter their processing for efficient presentation by APC; (2) substitution in anchor residues of a peptide can decrease the affinity of an otherwise immunodominant epitope and it can become subdominant or cryptic; (3) a set of mutations in the protein sequence may modify its natural conformation, thereby rendering the protease-sensitive regions inaccessible to the enzymatic breakdown (Figure 3). These events change the repertoire of epitopes being presented on APC for T cell activation impairing the recall response to a vaccine candidate during natural infection.
Recognition of specific cleavage sites on epitopes by proteases is an essential step during antigen processing. Schneider et al. increased the presentation efficiency of a subdominant epitope of hen egg lysozyme by adding a dibasic endopeptidase motif in its flanking regions; however, the T cell stimulatory capacity remained unchanged [148]. Substitutions altering processing and presentation of epitopes from Pseudomonas endotoxin-BII domain reduce their immunogenicity [149]. Spacer sequences or various tags added to recombinant proteins during purification steps were shown to distort the natural dominance of specific epitopes [150]. A change at a single site on a peptide can drastically change its presentation by APC. This is crucial in the context of Leishmania as there are copy number variations (CNV) in various genes of protozoan parasite associated with drug resistance [151,152]. This process can obviously generate proteins, which may be functionally similar, albeit possessing different amino acid residues generating from duplicated gene loci. This, in turn, may affect the antigen processing dynamics of a peptide and change the repertoire of physiological immunodominant epitopes of a given antigenic protein.
4. Leishmania Targets Antigen Presentation by APCs and Immune Synapse (IS)
The dynamics of APC–T cell interaction, expression levels of co-stimulatory and co-inhibitory molecules, and the cytokine milieu play decisive roles in the anti-leishmanial immune response. In the lymph node, T cells move through a meshwork of APCs, and on recognizing MHC-bound cognate antigen, forms a contact zone—immune synapse (IS)—at the T cell/APC interface. Exposure to chemokines during extravasation activates the adhesion molecules on T cells, leading to the interdigitation of cell’s glycocalyx. Adhesion occurs mainly through T cells expressed αLβ2 integrin LFA-1, which upon activation interacts with ICAM-1 expressed on endothelial cells. TCR signaling micro-clusters forms at the periphery of IS, which gradually moves inward by the retrograde flow of actin cytoskeleton and coalesce to form the main contact zone or core supramolecular activation cluster (cSMAC). cSMAC comprises of TCR-MHC complex enriched with Src-family kinases like Lck and Fyn, CD28, protein kinase C θ (PKCθ) and zeta-chain-associated protein kinase 70 (ZAP-70). In addition to this, interaction occurs at segregated contact zones or peripheral SMAC (p-SMAC) via LFA-1/ICAM-1, CD2/CD58 interaction [153,154]. pSMAC also has the cytoskeletal linker protein talin. The outermost distal SMAC (dSMAC) has CD43 and CD45 and performs mechano-supportive role. These molecular interactive forces are at peak after 30 min [155]. Once the T cell is in contact with the APC, TCR conveys a stop signal, forming a stable junction IS [156]. As an effect of LFA-1 and ICAM-1 interaction, downstream signaling events result in cytoskeletal remodeling and T cell motility [157] (Figure 4).
On TCR stimulation, several signaling proteins like the adaptor protein Linker of activation of T cells (LAT), which is phosphorylated by ZAP-70 [158], PKCθ [159], and tyrosine kinase Lck [160] are targeted to cholesterol and sphingolipids rich lipid rafts. L. major depletes membrane cholesterol [161] that may affect the signaling component’s segregation in lipid raft and the formation of a stable IS. L. donovani also interferes with the engagement of MHC-peptide complex with TCR [162], possibly via alteration of membrane lipid content, as shown by protection against VL by hypercholesterolemia [163], by cholesterol-enabled formation of a stable IS and cross-presentation and activation of CD8+ T cells [164]. Leishmania alters membrane cholesterol, disrupting the stability of the MHC-peptide complex, MHCII conformation, and affecting antigen presentation [165,166]. A cholesterol-liposome treatment re-established the formation of immune synapse, restored anti-leishmanial T cell repertoire, and immune functions in L. donovani infection [167].
Expression of cell surface molecules and their interaction also dictates the robustness of T cell activation. Leishmania donovani decreases the expression of both MHC-I and MHC-II in macrophages [168] and DCs [169] by direct internalization [135] or by the action of certain extracellular vesicles [170]. Leishmania binds to and affects the functioning of integrins [171] and αMβ2 (Mac-1) [172] on phagocytes, impairing IS formation and altered T cells stimulation. L. amazonensis also impairs the formation of adhesion signaling complexes on the plasma membrane [173].
The expression of co-stimulatory molecules and the threshold and magnitude of their interaction with co-receptors on T cells [174] can influence the generation of antigen-specific T cells. L. donovani lowers the expression of co-stimulatory molecules CD80 and CD86, which are critical for Th2 differentiation [175,176]. It also alters the micro-domain lipid constitution, hence lowering the availability of TCR ligands on the surface of macrophages [162]. CD40 and CD80 play a crucial role in anti-Leishmania T-cell activation, wherein L. major-infected macrophages co-cultured with peripheral blood leukocytes (PBL) showed an IFN–γ- and CD40-mediated increase in the expression of CD40 and CD80 [177,178,179,180,181].
CD4+ and CD8+ T cells express inhibitory cytotoxic T-lymphocyte-associated protein 4 (CTLA-4/CD152), which binds to CD80 (B7-1) and CD86 (B7-2). It terminates T cell response, generation of memory T cells [182], and regulates T cell anergy [183]. The maximum level of expression of CTLA-4 occurs 24–72 h post-activation [184]. Blockade of B7-2 following L. donovani infection resulted in enhanced Th1 and Th2 responses [185]. CTLA-4 blockade increased IFN-γ, IL-4, and CXC chemokine γIP-10 [186]. PD1 (CD279), an inhibitory co-receptor, is recruited to the IS and its engagement with PDL1 or PDL2 leads to the recruitment of tyrosine phosphatase SHP-2, that attenuates TCR signaling by phosphorylating signaling molecules like Vav, Akt, CD3ζ, ZAP70, and ERK [187]. L. donovani-infected macrophages showed a reduced PD1 expression and inhibition of apoptosis [188]. Another immune-inhibitory receptor on macrophage is the signal regulatory protein (SIRP)-α [189] but its role in T cell priming remains unknown.
Transcriptional profiling of genes upregulated during L. infantum infection shows that there is a marked difference in the gene expression in infected mice when compared to control mice. Upregulation of Th1 and M1 macrophage activation markers like Ifng, Stat1, Cxcl9, Cxcl10, Ccr5, Cxcr3, Xcl1, and Ccl3 occurs in early infection. Of the 112 genes studied, 22 showed differential expression in infected vs. uninfected mice. Il12rb2, Il23r, and Ptgs2 were exclusively expressed only in infected mice but not in the control group, suggesting that the immune molecules may vary drastically under the condition when initial priming is done (uninfected) compared to when subsequent infection occurs [190]. Thus, an aim in vaccination is to induce the balanced expression of co-stimulatory and co-inhibitory molecules such that appropriate Th cell priming occurs.
As the IS forms, several cytokines are secreted at the IS. The Th cells secrete cytokines such as IFN-γ, IL-10, and IL-2 [191] towards the antigenic target cells, or cytokines like IL-4, TNF, CCL3, MIP-1α towards target as well as non-target cells [192]. Actin remodeling plays an important role in the secretion of cytokines at the IS as silencing of Cdc42, a regulator of cytoskeletal rearrangement, inhibits the secretion of cytokines like IFN-γ at the IS [193].
During APC–T cell interaction lasting about ~12 h, abundant IL-2 is produced [194]. However, many other cytokines secreted over time, which may not be present at the time of IS formation, may be important in limiting Leishmania growth. Due to limited literature on this topic, one might speculate the potential cytokines trafficking from T-cells in case of Leishmania infected macrophages and vice versa that shapes the immune synapse are yet to be uncovered and many can have profound effects on both cell types and antigen presentation at IS. Since IS formation is a regulated, dynamic, and sequential process [195], it has multiple points of disruption that intracellularly residing parasites from Trypanosomatidae family may exploit to ensure their intracellular survival. MHC-II peptide density plays an important role in driving signals through TCR and subsequent cytokine secretion. The cytokine expression of a particular T cell subset may be determined by the affinity of its TCR for peptide-MHC-ligand [196]. Therefore, the binding strengths may be one of the mechanisms through which the cytokines may get affected at IS. For example, the coinhibitory receptor CTLA-4 has a higher affinity to bind to CD86 [197] than CD28, therefore, it outcompetes CD28 binding at IS and may lead to a suppressed T cell instead of activation [198]. In the case of murine VL, CTLA-4 increases TGF-β levels and apoptosis of CD4+ T cells [199]. The administration of anti-CTLA-4 mAbs to BALB/c mice following one day of infection with L. donovani enhances the frequency of IFN-γ- and IL-4-producing cells in both spleen and liver contributing to resistance against the parasite [186]. Aided with the availability of recombinant cytokines along with the monoclonal Abs, targeting inhibitory receptors provide some promising tools with which the limitations surmounting anti-leishmanial vaccines can be overcome.
5. Various Fates of T Cells during Infection
T cell response to an infection is a dynamic process. Following vaccination, phenotypic changes occur in antigen-specific T cells, which may be further altered during the course of infection [200]. The ‘functional avidity’ determined by the strength of ligand–receptor interactions at the IS or the T cells responsiveness to the titer of cognate antigen in vitro or their activation threshold is another parameter crucial in designing effective vaccines [201,202,203]. Polymorphisms in the co-receptor [204], presence of inhibitory molecules [205], micro RNAs [206], and location of the ligand [207] can influence T cell responses. On impaired antigen presentation and an unstable IS formation, only a few T cells are activated. These T cells can have different fates and can undergo anergy, exhaustion, apoptosis, or can induce a non-robust T cell response due to T cell plasticity.
Plasticity of T Cells
Naïve CD4+ T cells can be induced to differentiate into Th1, Th2, Th9, Th17, Th22, T follicular helper (Tfh), and T regulatory (Treg) cell type, based on the cytokine milieu. IL-12 signals through STAT4 and induces differentiation into Th1, and IL-4 signals through STAT-6 and skews the differentiation into Th2. TGF-β and IL-6 activate Smad family proteins and STAT3, respectively, driving Th17 differentiation, whereas TGF-β and IL-4 towards Th9, IL-6, TNF- α towards IL-22, IL-21 towards Tfh, and TGF-β alone towards Treg. These cells play decisive roles in modulating the efficacy of vaccines. Th17 are inflammatory cells that produce IL-17A and offer protection against intracellular pathogens, whereas Tregs play an immunosuppressive role and dampens the immune response induced by vaccines. TGF-β skews the T cell response to either Th17 or Treg [208]. The high concentration of TGF-β is conducive for Treg, whereas the low concentration of TGF-β, along with cytokines such as IL-21 and IL-6, promotes Th17 responses [209]. Treg cells express high-affinity IL-2 receptors and IL-2 stabilized Foxp3 a transcription factor crucial for maintaining suppression of the immune system [210]. However, the presence of proinflammatory cytokines like IL-21, IL-6, IL-23, and IL-1β, promotes differentiation into Th17 rather than Tregs [211]. Treg-induced immunosuppression is effected by both cytokines and co-inhibitory molecules like IL-10, TGF-β, perforins, granzymes, and CTLA-4 and LAG3 [210]. The Th17 differentiation to induced Treg (iTreg) can be also reprogrammed by small-molecule-like (aminooxy) acetate [212]. The plasticity of T cells, deviating from one subset to another based on the cytokines present during infection, thus induces a non-robust T-cell response.
6. Anergy, Exhaustion, and Apoptosis
Even in case of a robust T cell response, complete parasite elimination may not happen, resulting in chronic dormant infection. Under such an infection, exhaustion or attenuation of effector T cells occurs [213]. T cell exhaustion is characterized by low proliferation, production of IL-2, and inability to produce TNF-α and IFN-γ. A high load of antigen during the course of infection and high-affinity interaction can lead to rapid CD8+ T cells proliferation whereas lack of integration of stimulatory signals can drive CD8+ T cells to exhaustion [214,215], anergy, or apoptosis [216]. Of these integrating stimulatory signals, the cytokine milieu is a major determinant of the proliferation rate of naïve and memory T cells [217]. T cell anergy can occur due to the APC expressed immunomodulatory tryptophan metabolizing enzyme indoleamine-2,3-dioxygenase (IDO) [218] or ATP-catabolizing enzymes CD39 and CD73 [219]. Anergy can also occur when T cells interact with an immature APC, whereby the TCR–peptide–MHC interaction takes place but adequate costimulatory signals are not obtained [220]. Treg cells expressing immuno-modulatory molecules like IDO [221], ATP-metabolizing enzymes [222], and CTLA-4 [223] can result in sustained T cell anergy. The increased expression of IDO [224] or ATP-metabolizing enzymes [225] on DCs during L. major and L. amazonensis infection leads to poor induction of T cells. In L. donovani infection, an increased expression of PD-1 on parasite-specific CD8+ T cells characterizes T cell exhaustion [226]. L. amazonensis-infected macrophages produced TGF-β, a pro-apoptotic, immunosuppressive cytokine that induces T-cell anergy/apoptosis [227].
Parasite-specific T cells that surpass all stages of infection undergo programmed cell death once the infection is cleared, with an exception of a few memory T cells that have an antigen-independent long-term survival [228]. Further increase in the number of antigen-specific CD8+ T cells can be achieved via booster immunization. The nature of antigen used for booster immunization is of prime importance, as boosting with the homologous antigen can lead to its neutralization and dampened CD8+ T cell responses. This can be overcome by the use of a heterologous antigen [229]. However, repeated boosting can lead to the terminal differentiation and unresponsiveness of the memory T cells [230,231]. Repeated immunization has shown to increase the functional diversity of memory T cells and decrease their proliferation rate [232]. The quality of memory T cells generated under such conditions depends on the time duration between the immunization, the inflammatory status, the characteristics of the antigen(s) used for immunization, and the responses of the immune system towards each antigen [233]. As a consequence of re-stimulation of T cells, activation-induced cell death (AICD) or T cell-autonomous death (ACAD) may occur via expression of ligands like FasL, CD95L, TNF, etc. or by increased expression of Bim in T cells [228] leading to apoptosome formation [234]. As the inflammation is high during priming, it leads to the accumulation of effector and memory T cells and antagonistically delays the conversion of early CD8+ cells to late memory T cells [235].
Leishmania-Infected Macrophages Respond Differently to Antigen-Specific T Cells
Activated T cells capable of killing Leishmania-infected macrophages exit the lymph node and reach the site of infection and secrete cytokines like IFN-γ. Many studies discuss that the refractoriness towards IFN-γ abundance may be attributed to IFN-γR inhibition, as IFN-γ leads to classical (M1) macrophage activation generating nitric oxide and other microbicidal free radicals responsible for Leishmania killing. L. donovani amastigotes are capable of drastically reducing the levels of IFN-γ-induced expression of MHC-II and iNOS expression. This causes a fragmented anti-parasitic response via-JAK-STAT signaling. L. donovani amastigotes are also reported to impair IFN-induced STAT1α nuclear translocation by abrogating its association with importin-α5 [236]. L. major and L. mexicana modulate IFN-signaling, by suppression of IFN-γRα and IFN-γRβ subunit expression in addition to the components of this pathway such as Jak-1 and Jak-2 [237]. Apparently, this effect was attributed to the decreased availability of the receptor protein after infection [238]. SHP-1 phosphatase is another regulator of cytokine response in infected macrophages, as deficiency of this phosphatase leads to increased inflammatory cytokine response (TNF-α, IL-1, and IL-6). Leishmania can control SHP-1 activity and can allow its intracellular survival. Leishmania-induced increases in activation of macrophage SHP-1 are associated with impaired IFN-γ-triggered JAK2 activation [239]. In such a condition during the actual course of Leishmania infection, the infected macrophages would be unresponsive to the available pools of IFN-γ or TNF-α. Because a small number of antigen-specific T cells overcome the first barrier of antigen selection in situ, the host-protective anti-leishmanial effects are not abundant. Thus, abrogation of IFN-signaling, macrophage activation, and Jak/STAT signaling [240] affect the vaccine performance and possible accentuation of this pathway may bring therapeutic benefits to anti-leishmanial vaccination programs. A representation of selective-modulation of IS and its components by Leishmania is indicated in Figure 4.
7. Conclusions
Since the demonstration of vaccinating principles by Jenner, researchers have followed the same for protecting vaccines. Although vaccinating with whole organisms—killed or attenuated in many different forms—brought success in many cases, the anti-leishmanial vaccines failed. Similar can be said of recombinant or peptide vaccines. As all the approaches with assumptions that introduction of the antigen(s) to a host will elicit host-protective immunity against the virulent pathogen turned out to be invalid, we took up this analytical review to find out the possible reasons for the failures and the possible approach that may bring success to Leishmania vaccine generation. Firstly, trafficking of T cells between lymph nodes and site of infection is essential for their activation and maturation. Inhibition of CD4+ T cell migration pattern in lymph nodes is reported during Shigella infection, which affects antigenic priming. Similar reports are found in HIV-1 infection, whereby continuous stimulation of immune system impairs the migration ability of circulatory CCR6+ CXCR3+ Th cells from blood to peripheral organs [241,242]. In fact, Leishmania infection impairs the migration of DC’s from the site of infection to lymph node for antigenic priming, which alters host immune response [243,244]. Therefore, thorough assessment of immune priming, circulating and central memory T cells are crucial for vaccination-induced disease protection. Secondly, epigenetic regulation of host-protective signaling in Leishmania-infected macrophages [245] indicates that parasite-induced epigenetic events may also be responsible for Th cell plasticity, altered MHC presentation, and vaccination outcome. Thirdly, there lies ambiguity in the importance of persistent parasites in maintaining central memory T cells. It needs to reassessed from a new perspective as it may lead us to revisit whole-parasite-based vaccines as a potential vaccination strategy [246,247]. In fact, killed-parasite vaccine was found to be inefficacious due to early recruitment of neutrophils, and healed C57BL/6 mice were immune to subsequent sand fly challenge infection [248]. This fortifies the role of additional parameters, such as sand fly salivary proteins in conferring protective immunity as shown in other studies [249]. Also, adoption of sand fly challenge model is the need of the hour for a better correlation of immune response parameters. Glennie et al. showed that TRM (tissue-resident memory) cells reside in tissues distant to Leishmania infection site and are better equipped at controlling subsequent infection by quick recruitment of circulating memory T cells in CXCR-3 dependent manner. However, later observations indicated that protection by TRM cells in Leishmania-immune C57BL/6 mice is mediated by early recruitment of inflammatory monocytes, which eliminate Leishmania via ROS and NO generation. Irrespective of effector mechanism, TRM cells, a novel memory T cell subset, appears to be important in generating protective immunity during the early phase of infection, when parasite immune-escape mechanism are relatively dormant [250]. This needs detailed experimental analysis as the ability to generate TRM by vaccine candidates may be incorporated as a novel parameter for assessing vaccination efficacy. Fourthly, re-enforcing parameters such as dendritic-cell based vaccine, TLR ligands-based adjuvants, and supplementing the vaccination with a blockade of co-inhibitory signal molecules like CTLA-4, PD-1, CD200R, and TIM-3 deserve consideration for inclusion in the vaccination protocol. It may enhance vaccination efficacy and T cell activation by modulating B7 molecules [198]. Finally, the principle of vaccination that focuses on host-protection as the outcome needs to include the principles of immunoregulation so that the elicitation of priming-induced T cell memory should be robust enough to match the immune interfering effects of the exposure to a pathogen. Hence, reassessment of anti-leishmanial immune-correlates in vector-transmission based infection models, incorporating TRM cells in pre-clinical vaccine efficacy studies, and co-inhibitory signal blocking at IS during prophylaxis look to be promising future approaches for reaching the goal of a successful anti-leishmanial vaccination.
Author Contributions
S.Z., S.K., P.C., Y.B., and A.N. composed the manuscript including compilation of the Table. The figures were drawn by P.C., A.S., and B.S., who also conceptualized the review and implemented the project. S.R. helped in the preparation of manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Garçon, N.; Stern, P.L.; Cunningham, A.L. Understanding Modern Vaccines: Perspectives in Vaccinology; Elsevier: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Cambronero, M.R.; Prado-Cohrs, D.; Sanroma, M.L. Conceptos Inmunológicos Básicos Aplicados a La Vacunología. Vacunas 2017, 18, 49–58. [Google Scholar] [CrossRef]
- Esparza, J. A New Scientific Paradigm May Be Needed to Finally Develop an HIV Vaccine. Front. Immunol. 2015, 6, 124. [Google Scholar] [CrossRef] [PubMed]
- Cobey, S. Pathogen Evolution and the Immunological Niche. Ann. N. Y. Acad. Sci. 2014, 1320, 1. [Google Scholar] [CrossRef] [PubMed]
- Coler, R.N.; Reed, S.G. Second-Generation Vaccines against Leishmaniasis. Trends Parasitol. 2005, 21, 244–249. [Google Scholar] [CrossRef]
- Fernandes, A.P.; Coelho, E.A.F.; Machado-Coelho, G.L.L.; Junior, G.G.; Gazzinelli, R.T. Making an Anti-Amastigote Vaccine for Visceral Leishmaniasis: Rational, Update and Perspectives. Curr. Opin. Microbiol. 2012, 15, 476–485. [Google Scholar] [CrossRef]
- Kedzierski, L. Leishmaniasis Vaccine: Where Are We Today? J. Glob. Infect. Dis. 2010, 2, 177–185. [Google Scholar] [CrossRef]
- Burza, S.; Croft, S.L.; Boelaert, M. Leishmaniasis. Lancet 2018, 392, 951–970. [Google Scholar] [CrossRef]
- Titus, R.G. The Immunomodulatory Factors of Bloodfeeding Arthropod Saliva. Parasite Immunol. 2000, 22, 319–331. [Google Scholar]
- Zahedifard, F.; Gholami, E.; Taheri, T.; Taslimi, Y.; Doustdari, F.; Seyed, N.; Torkashvand, F.; Meneses, C.; Papadopoulou, B.; Kamhawi, S. Enhanced Protective Efficacy of Nonpathogenic Recombinant Leishmania Tarentolae Expressing Cysteine Proteinases Combined with a Sand Fly Salivary Antigen. PLoS Negl. Trop. Dis. 2014, 8, e2751. [Google Scholar] [CrossRef]
- Sacks, D.; Noben-Trauth, N. The Immunology of Susceptibility and Resistance to Leishmania Major in Mice. Nat. Rev. Immunol. 2002, 2, 845–858. [Google Scholar] [CrossRef]
- Zhang, W.-W.; Charest, H.; Ghedin, E.; Matlashewski, G. Identification and Overexpression of the A2 Amastigote-Specific Protein in Leishmania Donovani. Mol. Biochem. Parasitol. 1996, 78, 79–90. [Google Scholar] [CrossRef]
- Flinn, H.M.; Rangarajan, D.; Smith, D.F. Expression of a Hydrophilic Surface Protein in Infective Stages of Leishmania Major. Mol. Biochem. Parasitol. 1994, 65, 259–270. [Google Scholar] [CrossRef]
- Alce, T.M.; Gokool, S.; McGhie, D.; Stäger, S.; Smith, D.F. Expression of Hydrophilic Surface Proteins in Infective Stages of Leishmania Donovani. Mol. Biochem. Parasitol. 1999, 1, 191–196. [Google Scholar] [CrossRef]
- Dey, R.; Dagur, P.K.; Selvapandiyan, A.; McCoy, J.P.; Salotra, P.; Duncan, R.; Nakhasi, H.L. Live Attenuated Leishmania Donovani P27 Gene Knockout Parasites Are Nonpathogenic and Elicit Long-Term Protective Immunity in BALB/c Mice. J. Immunol. 2013, 190, 2138–2149. [Google Scholar] [CrossRef]
- Selvapandiyan, A.; Debrabant, A.; Duncan, R.; Muller, J.; Salotra, P.; Sreenivas, G.; Salisbury, J.L.; Nakhasi, H.L. Centrin Gene Disruption Impairs Stage-Specific Basal Body Duplication and Cell Cycle Progression in Leishmania. J. Biol. Chem. 2004, 279, 25703–25710. [Google Scholar] [CrossRef]
- Saljoughian, N.; Taheri, T.; Rafati, S. Live Vaccination Tactics: Possible Approaches for Controlling Visceral Leishmaniasis. Front. Immunol. 2014, 5, 134. [Google Scholar] [CrossRef] [Green Version]
- Bruhn, K.W.; Birnbaum, R.; Haskell, J.; Vanchinathan, V.; Greger, S.; Narayan, R.; Chang, P.L.; Tran, T.A.; Hickerson, S.M.; Beverley, S.M.; et al. Killed but Metabolically Active Leishmania Infantum as a Novel Whole-Cell Vaccine for Visceral Leishmaniasis. Clin. Vaccine Immunol. 2012, 19, 490–498. [Google Scholar] [CrossRef]
- Musa, A.M.; Khalil, E.A.G.; Mahgoub, F.A.E.; Elgawi, S.H.H.; Modabber, F.; Elkadaru, A.E.M.Y.; Aboud, M.H.; Noazin, S.; Ghalib, H.W.; El-Hassan, A.M. Immunochemotherapy of Persistent Post-Kala-Azar Dermal Leishmaniasis: A Novel Approach to Treatment. Trans. R. Soc. Trop. Med. Hyg. 2008, 102, 58–63. [Google Scholar] [CrossRef]
- Palatnik-de-Sousa, C.B.; Moreno, M.; Paraguai-de-Souza, E.; Borojevic, R. The FML Vaccine (Fucose-Manose Ligand) Protects Hamsters from Experimental Kala-Azar. Cienc. Cult. 1994, 64, 290–296. [Google Scholar]
- Santos, W.R.; Paraguai de Souza, E.; Palatnik, M.; Palatnik de Sousa, C.B.; de Souza, E.P.; Palatnik, M.; de Sousa, C.B.P. Vaccination of Swiss Albino Mice against Experimental Visceral Leishmaniasis with the FML Antigen of Leishmaniadonovani. Vaccine 1999, 17, 2554–2561. [Google Scholar] [CrossRef]
- Nogueira, F.S.; Moreira, M.A.B.; Borja-Cabrera, G.P.; Santos, F.N.; Menz, I.; Parra, L.E.; Xu, Z.; Chu, H.J.; Palatnik-de-Sousa, C.B.; Luvizotto, M.C.R. Leishmune® Vaccine Blocks the Transmission of Canine Visceral Leishmaniasis: Absence of Leishmania Parasites in Blood, Skin and Lymph Nodes of Vaccinated Exposed Dogs. Vaccine 2005, 23, 4805–4810. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, E.M.; Barbosa, A.D.F.; Santos, F.N.; Borja-Cabrera, G.P.; Nico, D.; Souza, L.O.P.; Mendes-Aguiar, C.D.O.; De Souza, E.P.; Fampa, P.; Parra, L.E.; et al. The FML-Vaccine (Leishmune®) against Canine Visceral Leishmaniasis: A Transmission Blocking Vaccine. Vaccine 2006, 24, 2423–2431. [Google Scholar] [CrossRef] [PubMed]
- Afrin, F.; Ali, N. Adjuvanticity and Protective Immunity Elicited by Leishmania Donovani Antigens Encapsulated in Positively Charged Liposomes. Infect. Immun. 1997, 65, 2371–2377. [Google Scholar]
- Bhowmick, S.; Ravindran, R.; Ali, N. Leishmanial Antigens in Liposomes Promote Protective Immunity and Provide Immunotherapy against Visceral Leishmaniasis via Polarized Th1 Response. Vaccine 2007, 25, 6544–6556. [Google Scholar] [CrossRef] [PubMed]
- Mazumder, S.; Ravindran, R.; Banerjee, A.; Ali, N. Non-Coding PDNA Bearing Immunostimulatory Sequences Co-Entrapped with Leishmanial Antigens in Cationic Liposomes Elicits Almost Complete Protection against Experimental Visceral Leishmaniasis in BALB/c Mice. Vaccine 2007, 25, 8771–8781. [Google Scholar] [CrossRef] [PubMed]
- Ravindran, R.; Maji, M.; Ali, N. Vaccination with Liposomal Leishmanial Antigens Adjuvanted with Monophosphoryl Lipid-Trehalose Dicorynomycolate (MPL-TDM) Confers Long-Term Protection against Visceral Leishmaniasis through a Human Administrable Route. Mol. Pharm. 2012, 9, 59–70. [Google Scholar] [CrossRef] [PubMed]
- De Souza, E.P.; Bernardo, R.R.; Palatnik, M.; de Sousa, C.B.P. Vaccination of Balb/c Mice against Experimental Visceral Leishmaniasis with the GP36 Glycoprotein Antigen of Leishmania Donovani. Vaccine 2001, 19, 3104–3115. [Google Scholar] [CrossRef]
- Bhowmick, S.; Ali, N. Identification of Novel Leishmania Donovani Antigens That Help Define Correlates of Vaccine-Mediated Protection in Visceral Leishmaniasis. PLoS ONE 2009, 4, e5820. [Google Scholar] [CrossRef] [PubMed]
- Jaffe, C.L.; Rachamim, N.; Sarfstein, R. Characterization of Two Proteins from Leishmania Donovani and Their Use for Vaccination against Visceral Leishmaniasis. J. Immunol. 1990, 144, 699–706. [Google Scholar] [PubMed]
- Tewary, P.; Mehta, J.; Sukumaran, B.; Madhubala, R. Vaccination with Leishmania Soluble Antigen and Immunostimulatory Oligodeoxynucleotides Induces Specific Immunity and Protection against Leishmania Donovani Infection. FEMS Immunol. Med. Microbiol. 2004, 42, 241–248. [Google Scholar] [CrossRef]
- Iborra, S.; Carrión, J.; Anderson, C.; Alonso, C.; Sacks, D.; Soto, M. Vaccination with the Leishmania Infantum Acidic Ribosomal P0 Protein plus CpG Oligodeoxynucleotides Induces Protection against Cutaneous Leishmaniasis in C57BL/6 Mice but Does Not Prevent Progressive Disease in BALB/c Mice. Infect. Immun. 2005, 73, 5842–5852. [Google Scholar] [CrossRef] [PubMed]
- Garg, R.; Dube, A. Animal Models for Vaccine Studies for Visceral Leishmaniasis. Indian J. Med. Res. 2006, 123, 439–454. [Google Scholar] [PubMed]
- Kumari, S.; Samant, M.; Misra, P.; Khare, P.; Sisodia, B.; Shasany, A.K.; Dube, A. Th1-Stimulatory Polyproteins of Soluble Leishmania Donovani Promastigotes Ranging from 89.9 to 97.1 KDa Offers Long-Lasting Protection against Experimental Visceral Leishmaniasis. Vaccine 2008, 26, 5700–5711. [Google Scholar] [CrossRef] [PubMed]
- Kumari, S.; Samant, M.; Khare, P.; Sundar, S.; Sinha, S.; Dube, A. Induction of Th1-Type Cellular Responses in Cured/Exposed Leishmania-Infected Patients and Hamsters against Polyproteins of Soluble Leishmania Donovani Promastigotes Ranging from 89.9 to 97.1 KDa. Vaccine 2008, 26, 4813–4818. [Google Scholar] [CrossRef]
- Karanja, R.; Ingonga, J.; Mwangi, M.; Mwala, D.; Lugalia, R.; Magambo, J.; Tonui, W. Immunization with a Combination of Leishmania Major Lipophosphoglycan (LPG) and Phlebotomus Duboscqi Salivary Gland Lysates (SGLs) Abrogates Protective Effect of LPG against L. Major in BALB/c Mice. Afr. J. Health Sci. 2011, 18, 1–5. [Google Scholar]
- Tonui, W.K.; Mbati, P.A.; Anjili, C.O.; Orago, A.S.; Turco, S.J.; Githure, J.I.; Keoch, D.K. Transmission Blocking Vaccine Studies in Leishmaniasis: II Effect of Immunisation Using Leshmania Major Derived 63 Kilodalton on the Course of L. Major Infection in BALB/c Mice. East Afr. Med. J. 2001, 78, 90–92. [Google Scholar] [CrossRef]
- Bhowmick, S.; Ravindran, R.; Ali, N. Gp63 in Stable Cationic Liposomes Confers Sustained Vaccine Immunity to Susceptible BALB/c Mice Infected with Leishmania Donovani. Infect. Immun. 2008, 76, 1003–1015. [Google Scholar] [CrossRef]
- Handman, E.; Button, L.L.; McMaster, R.W. Leishmania Major: Production of Recombinant Gp63, Its Antigenicity and Immunogenicity in Mice. Exp. Parasitol. 1990, 70, 427–435. [Google Scholar] [CrossRef]
- Abdelhak, S.; Louzir, H.; Timm, J.; Blel, L.; Benlasfar, Z.; Lagranderie, M.; Gheorghiu, M.; Dellagi, K.; Gicquel, B. Recombinant BCG Expressing the Leishmania Surface Antigen Gp63 Induces Protective Immunity against Leishmania Major Infection in BALB/c Mice. Microbiology 1995, 141, 1585–1592. [Google Scholar] [CrossRef]
- Badiee, A.; Jaafari, M.R.; Khamesipour, A.; Samiei, A.; Soroush, D.; Kheiri, M.T.; Barkhordari, F.; McMaster, W.R.; Mahboudi, F. Enhancement of Immune Response and Protection in BALB/c Mice Immunized with Liposomal Recombinant Major Surface Glycoprotein of Leishmania (Rgp63): The Role of Bilayer Composition. Colloids Surf. B Biointerfaces 2009, 74, 37–44. [Google Scholar] [CrossRef]
- Mazumder, S.; Maji, M.; Das, A.; Ali, N. Potency, Efficacy and Durability of DNA/DNA, DNA/Protein and Protein/Protein Based Vaccination Using Gp63 against Leishmania Donovani in BALB/c Mice. PLoS ONE 2011, 6, e14644. [Google Scholar] [CrossRef] [PubMed]
- Chenik, M.; Louzir, H.; Ksontini, H.; Dilou, A.; Abdmouleh, I.; Dellagi, K. Vaccination with the Divergent Portion of the Protein Histone H2B of Leishmania Protects Susceptible BALB/c Mice against a Virulent Challenge with Leishmania Major. Vaccine 2006, 24, 2521–2529. [Google Scholar] [CrossRef] [PubMed]
- Meddeb-Garnaoui, A.; Toumi, A.; Ghelis, H.; Mahjoub, M.; Louzir, H.; Chenik, M. Cellular and Humoral Responses Induced by Leishmania Histone H2B and Its Divergent and Conserved Parts in Cutaneous and Visceral Leishmaniasis Patients, Respectively. Vaccine 2010, 28, 1881–1886. [Google Scholar] [CrossRef] [PubMed]
- Dole, V.S.; Raj, V.S.; Ghosh, A.; Madhubala, R.; Myler, P.J.; Stuart, K.D. Immunization with Recombinant LD1 Antigens Protects against Experimental Leishmaniasis. Vaccine 2000, 19, 423–430. [Google Scholar] [CrossRef]
- Tewary, P.; Sukumaran, B.; Saxena, S.; Madhubala, R. Immunostimulatory Oligodeoxynucleotides Are Potent Enhancers of Protective Immunity in Mice Immunized with Recombinant ORFF Leishmanial Antigen. Vaccine 2004, 22, 3053–3060. [Google Scholar] [CrossRef]
- Sharma, A.; Madhubala, R. Ubiquitin Conjugation of Open Reading Frame F DNA Vaccine Leads to Enhanced Cell-Mediated Immune Response and Induces Protection against Both Antimony-Susceptible and-Resistant Strains of Leishmania Donovani. J. Immunol. 2009, 183, 7719–7731. [Google Scholar] [CrossRef]
- Kushawaha, P.K.; Gupta, R.; Sundar, S.; Sahasrabuddhe, A.A.; Dube, A. Elongation Factor-2, a Th1 Stimulatory Protein of Leishmania Donovani, Generates Strong IFN-γ and IL-12 Response in Cured Leishmania -Infected Patients/Hamsters and Protects Hamsters against Leishmania Challenge. J. Immunol. 2011, 187, 6417–6427. [Google Scholar] [CrossRef]
- Bhardwaj, S.; Vasishta, R.K.; Arora, S.K. Vaccination with a Novel Recombinant Leishmania Antigen plus MPL Provides Partial Protection against L. Donovani Challenge in Experimental Model of Visceral Leishmaniasis. Exp. Parasitol. 2009, 121, 29–37. [Google Scholar] [CrossRef]
- Stäger, S.; Smith, D.F.; Kaye, P.M. Immunization with a Recombinant Stage-Regulated Surface Protein from Leishmania Donovani Induces Protection against Visceral Leishmaniasis. J. Immunol. 2000, 165, 7064–7071. [Google Scholar] [CrossRef]
- Nagill, R.; Kaur, S. Enhanced Efficacy and Immunogenicity of 78 KDa Antigen Formulated in Various Adjuvants against Murine Visceral Leishmaniasis. Vaccine 2010, 28, 4002–4012. [Google Scholar] [CrossRef]
- Streit, J.A.; Recker, T.J.; Donelson, J.E.; Wilson, M.E. BCG Expressing LCR1 of Leishmania Chagasi Induces Protective Immunity in Susceptible Mice. Exp. Parasitol. 2000, 94, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Serezani, C.H.C.; Franco, A.R.; Wajc, M.; Yokoyama-Yasunaka, J.K.U.; Wunderlich, G.; Borges, M.M.; Uliana, S.R.B. Evaluation of the Murine Immune Response to Leishmania Meta 1 Antigen Delivered as Recombinant Protein or DNA Vaccine. Vaccine 2002, 20, 3755–3763. [Google Scholar] [CrossRef]
- Gomez-Rodriguez, J.; Wohlfert, E.A.; Handon, R.; Meylan, F.; Wu, J.Z.; Anderson, S.M.; Kirby, M.R.; Belkaid, Y.; Schwartzberg, P.L. Itk-Mediated Integration of T Cell Receptor and Cytokine Signaling Regulates the Balance between Th17 and Regulatory T Cells. J. Exp. Med. 2014, 211, 529–543. [Google Scholar] [CrossRef] [PubMed]
- Morris, R.V.; Shoemaker, C.B.; David, J.R.; Lanzaro, G.C.; Titus, R.G. Sandfly Maxadilan Exacerbates Infection with Leishmania Major and Vaccinating against It Protects against L. Major Infection. J. Immunol. 2001, 167, 5226–5230. [Google Scholar] [CrossRef] [PubMed]
- Gomes, R.; Teixeira, C.; Teixeira, M.J.; Oliveira, F.; Menezes, M.J.; Silva, C.; De Oliveira, C.I.; Miranda, J.C.; Elnaiem, D.E.; Kamhawi, S.; et al. Immunity to a Salivary Protein of a Sand Fly Vector Protects against the Fatal Outcome of Visceral Leishmaniasis in a Hamster Model. Proc. Natl. Acad. Sci. USA 2008, 105, 7845–7850. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Zhang, W.W.; Matlashewski, G. Immunization with A2 Protein Results in a Mixed Th1/Th2 and a Humoral Response Which Protects Mice against Leishmania Donovani Infections. Vaccine 2001, 20, 59–66. [Google Scholar] [CrossRef]
- Henriquez, F.L.; Campbell, S.A.; Roberts, C.W.; Mullen, A.B.; Burchmore, R.; Carter, K.C. Vaccination with Recombinant Leishmania Donovani Gamma-Glutamylcysteine Synthetase Fusion Protein Protects Against L. Donovani Infection. J. Parasitol. 2010, 96, 929–936. [Google Scholar] [CrossRef]
- Carcelen, J.; Iniesta, V.; Fernandez-Cotrina, J.; Serrano, F.; Parejo, J.C.; Corraliza, I.; Gallardo-Soler, A.; Marañón, F.; Soto, M.; Alonso, C.; et al. The Chimerical Multi-Component Q Protein from Leishmania in the Absence of Adjuvant Protects Dogs against an Experimental Leishmania Infantum Infection. Vaccine 2009, 27, 5964–5973. [Google Scholar] [CrossRef]
- Zadeh-Vakili, A.; Taheri, T.; Taslimi, Y.; Doustdari, F.; Salmanian, A.H.; Rafati, S. Immunization with the Hybrid Protein Vaccine, Consisting of Leishmania Major Cysteine Proteinases Type I (CPB) and Type II (CPA), Partially Protects against Leishmaniasis. Vaccine 2004, 22, 1930–1940. [Google Scholar] [CrossRef]
- Kronenberg, K.; Brosch, S.; Butsch, F.; Tada, Y.; Shibagaki, N.; Udey, M.C.; Von Stebut, E. Vaccination with TAT-Antigen Fusion Protein Induces Protective, CD8+ T Cell-Mediated Immunity against Leishmania Major. J. Investig. Dermatol. 2010, 130, 2602–2610. [Google Scholar] [CrossRef]
- Skeiky, Y.A.W.; Coler, R.N.; Brannon, M.; Stromberg, E.; Greeson, K.; Crane, R.T.; Campos-Neto, A.; Reed, S.G. Protective Efficacy of a Tandemly Linked, Multi-Subunit Recombinant Leishmanial Vaccine (Leish-111f) Formulated in MPL® Adjuvant. Vaccine 2002, 20, 3292–3303. [Google Scholar] [CrossRef]
- Coler, R.N.; Goto, Y.; Bogatzki, L.; Raman, V.; Reed, S.G. Leish-111f, a Recombinant Polyprotein Vaccine That Protects against Visceral Leishmaniasis by Elicitation of CD4+ T Cells. Infect. Immun. 2007, 75, 4648–4654. [Google Scholar] [CrossRef] [PubMed]
- Chakravarty, J.; Kumar, S.; Trivedi, S.; Rai, V.K.; Singh, A.; Ashman, J.A.; Laughlin, E.M.; Coler, R.N.; Kahn, S.J.; Beckmann, A.M.; et al. A Clinical Trial to Evaluate the Safety and Immunogenicity of the LEISH-F1+MPL-SE Vaccine for Use in the Prevention of Visceral Leishmaniasis. Vaccine 2011, 29, 3531–3537. [Google Scholar] [CrossRef] [PubMed]
- Titus, R.G.; Gueiros-Filho, F.J.; De Freitas, L.A.R.; Beverley, S.M. Development of a Safe Live Leishmania Vaccine Line by Gene Replacement. Proc. Natl. Acad. Sci. USA 1995, 92, 10267–10271. [Google Scholar] [CrossRef] [PubMed]
- Veras, P.S.T.; Brodskyn, C.I.; Balestieri, F.M.P.; De Freitas, L.A.R.; Ramos, A.P.S.; Queiroz, A.R.P.; Barral, A.; Beverley, S.M.; Barral-Netto, M. A Dhfr-Ts- Leishmania Major Knockout Mutant Cross-Protects against Leishmania Amazonensis. Mem. Inst. Oswaldo Cruz 1999, 94, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Selvapandiyan, A.; Dey, R.; Nylen, S.; Duncan, R.; Sacks, D.; Nakhasi, H.L. Intracellular Replication-Deficient Leishmania Donovani Induces Long Lasting Protective Immunity against Visceral Leishmaniasis. J. Immunol. 2009, 183, 1813–1820. [Google Scholar] [CrossRef]
- Papadopoulou, B.; Roy, G.; Breton, M.; Kündig, C.; Dumas, C.; Fillion, I.; Singh, A.K.; Olivier, M.; Ouellette, M. Reduced Infectivity of a Leishmania Donovani Biopterin Transporter Genetic Mutant and Its Use as an Attenuated Strain for Vaccination. Infect. Immun. 2002, 70, 62–68. [Google Scholar] [CrossRef]
- Silvestre, R.; Cordeiro-Da-Silva, A.; Santarém, N.; Vergnes, B.; Sereno, D.; Ouaissi, A. SIR2-Deficient Leishmania Infantum Induces a Defined IFN-γ/IL-10 Pattern That Correlates with Protection. J. Immunol. 2007, 179, 3161–3170. [Google Scholar] [CrossRef]
- Dey, R.; Meneses, C.; Salotra, P.; Kamhawi, S.; Nakhasi, H.L.; Duncan, R. Characterization of a Leishmania Stage-Specific Mitochondrial Membrane Protein That Enhances the Activity of Cytochrome c Oxidase and Its Role in Virulence. Mol. Microbiol. 2010, 77, 399–414. [Google Scholar] [CrossRef]
- Carrión, J.; Folgueira, C.; Soto, M.; Fresno, M.; Requena, J.M. Leishmania Infantum HSP70-II Null Mutant as Candidate Vaccine against Leishmaniasis: A Preliminary Evaluation. Parasites Vectors 2011, 4, 150. [Google Scholar] [CrossRef]
- Solana, J.C.; Ramírez, L.; Corvo, L.; de Oliveira, C.I.; Barral-Netto, M.; Requena, J.M.; Iborra, S.; Soto, M. Vaccination with a Leishmania Infantum HSP70-II Null Mutant Confers Long-Term Protective Immunity against Leishmania Major Infection in Two Mice Models. PLoS Negl. Trop. Dis. 2017, 11, e0005644. [Google Scholar] [CrossRef] [PubMed]
- Kébaïer, C.; Uzonna, J.E.; Beverley, S.M.; Scott, P. Immunization with Persistent Attenuated Δlpg2 Leishmania Major Parasites Requires Adjuvant to Provide Protective Immunity in C57BL/6 Mice. Infect. Immun. 2006, 74, 777–780. [Google Scholar] [CrossRef] [PubMed]
- Uzonna, J.E.; Späth, G.F.; Beverley, S.M.; Scott, P. Vaccination with Phosphoglycan-Deficient Leishmania Major Protects Highly Susceptible Mice from Virulent Challenge without Inducing a Strong Th1 Response. J. Immunol. 2004, 172, 3793–3797. [Google Scholar] [CrossRef] [PubMed]
- Breton, M.; Tremblay, M.J.; Ouellette, M.; Papadopoulou, B. Live Nonpathogenic Parasitic Vector as a Candidate Vaccine against Visceral Leishmaniasis. Infect. Immun. 2005, 73, 6372–6382. [Google Scholar] [CrossRef] [Green Version]
- Mizbani, A.; Taheri, T.; Zahedifard, F.; Taslimi, Y.; Azizi, H.; Azadmanesh, K.; Papadopoulou, B.; Rafati, S. Recombinant Leishmania Tarentolae Expressing the A2 Virulence Gene as a Novel Candidate Vaccine against Visceral Leishmaniasis. Vaccine 2009, 28, 53–62. [Google Scholar] [CrossRef]
- Yam, K.K.; Hugentobler, F.; Pouliot, P.; Stern, A.M.; Lalande, J.D.; Matlashewski, G.; Olivier, M.; Cousineau, B. Generation and Evaluation of A2-Expressing Lactococcus Lactis Live Vaccines against Leishmania Donovani in BALB/c Mice. J. Med. Microbiol. 2011, 60, 1248–1260. [Google Scholar] [CrossRef]
- Gurunathan, S.; Sacks, D.L.; Brown, D.R.; Reiner, S.L.; Charest, H.; Glaichenhaus, N.; Seder, R.A. Vaccination with DNA Encoding the Immunodominant LACK Parasite Antigen Confers Protective Immunity to Mice Infected with Leishmania Major. J. Exp. Med. 1997, 186, 1137–1147. [Google Scholar] [CrossRef]
- Gomes, D.C.d.O.; Pinto, E.F.; de Melo, L.D.B.; Lima, W.P.; Larraga, V.; Lopes, U.G.; Rossi-Bergmann, B.; de Oliveira Gomes, D.C.; Pinto, E.F.; de Melo, L.D.B.; et al. Intranasal Delivery of Naked DNA Encoding the LACK Antigen Leads to Protective Immunity against Visceral Leishmaniasis in Mice. Vaccine 2007, 25, 2168–2172. [Google Scholar] [CrossRef]
- De, D.O.G.; Da, B.S.C.S.; De, H.M.G.; Lopes, U.G.; Rossi-Bergmann, B.; De Oliveira Gomes, D.C.; Da Silva Costa Souza, B.L.; De Matos Guedes, H.L.; Lopes, U.G.; Rossi-Bergmann, B. Intranasal Immunization with LACK-DNA Promotes Protective Immunity in Hamsters Challenged with Leishmania Chagasi. Parasitology 2011, 138, 1892–1897. [Google Scholar] [CrossRef]
- Melby, P.C.; Yang, J.; Zhao, W.; Perez, L.E.; Cheng, J. Leishmania Donovani P36(LACK) DNA Vaccine Is Highly Immunogenic but Not Protective against Experimental Visceral Leishmaniasis. Infect. Immun. 2001, 69, 4719–4725. [Google Scholar] [CrossRef]
- Kaur, T.; Sobti, R.C.; Kaur, S. Cocktail of Gp63 and Hsp70 Induces Protection against Leishmania Donovani in BALB/c Mice. Parasite Immunol. 2011, 33, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Labrecque, S.; Matlashewski, G. Protection against Leishmania Donovani Infection by DNA Vaccination: Increased DNA Vaccination Efficiency through Inhibiting the Cellular P53 Response. Vaccine 2001, 19, 3169–3178. [Google Scholar] [CrossRef]
- Resende, D.M.; Caetano, B.C.; Dutra, M.S.; Penido, M.L.O.; Abrantes, C.F.; Verly, R.M.; Resende, J.M.; Piló-Veloso, D.; Rezende, S.A.; Bruna-Romero, O.; et al. Epitope Mapping and Protective Immunity Elicited by Adenovirus Expressing the Leishmania Amastigote Specific A2 Antigen: Correlation with IFN-γ and Cytolytic Activity by CD8+ T Cells. Vaccine 2008, 26, 4585–4593. [Google Scholar] [CrossRef] [PubMed]
- Sukumaran, B.; Tewary, P.; Saxena, S.; Madhubala, R. Vaccination with DNA Encoding ORFF Antigen Confers Protective Immunity in Mice Infected with Leishmania Donovani. Vaccine 2003, 21, 1292–1299. [Google Scholar] [CrossRef]
- Tewary, P.; Saxena, S.; Madhubala, R. Co-administration of IL-12 DNA with rORFF antigen confers long-term protective immunity against experimental visceral leishmaniaisis. Vaccine 2006, 24, 2409–2416. [Google Scholar] [CrossRef]
- Basu, R.; Bhaumik, S.; Basu, J.M.; Naskar, K.; De, T.; Roy, S. Kinetoplastid Membrane Protein-11 DNA Vaccination Induces Complete Protection against Both Pentavalent Antimonial-Sensitive and -Resistant Strains of Leishmania Donovani That Correlates with Inducible Nitric Oxide Synthase Activity and IL-4 Generation: E. J. Immunol. 2005, 174, 7160–7171. [Google Scholar] [CrossRef]
- Bhaumik, S.; Basu, R.; Sen, S.; Naskar, K.; Roy, S. KMP-11 DNA Immunization Significantly Protects against L. Donovani Infection but Requires Exogenous IL-12 as an Adjuvant for Comparable Protection against L. Major. Vaccine 2009, 27, 1306–1316. [Google Scholar] [CrossRef]
- Carter, K.C.; Henriquez, F.L.; Campbell, S.A.; Roberts, C.W.; Nok, A.; Mullen, A.B.; McFarlane, E. DNA Vaccination against the Parasite Enzyme Gamma-Glutamylcysteine Synthetase Confers Protection against Leishmania Donovani Infection. Vaccine 2007, 25, 4502–4509. [Google Scholar] [CrossRef]
- Saldarriaga, O.A.; Travi, B.L.; Park, W.; Perez, L.E.; Melby, P.C. Immunogenicity of a Multicomponent DNA Vaccine against Visceral Leishmaniasis in Dogs. Vaccine 2006, 24, 1928–1940. [Google Scholar] [CrossRef]
- Samant, M.; Gupta, R.; Kumari, S.; Misra, P.; Khare, P.; Kushawaha, P.K.; Sahasrabuddhe, A.A.; Dube, A. Immunization with the DNA-Encoding N-Terminal Domain of Proteophosphoglycan of Leishmania Donovani Generates Th1-Type Immunoprotective Response against Experimental Visceral Leishmaniasis. J. Immunol. 2009, 183, 470–479. [Google Scholar] [CrossRef]
- Sachdeva, R.; Banerjea, A.C.; Malla, N.; Dubey, M.L. Immunogenicity and Efficacy of Single Antigen Gp63, Polytope and PolytopeHSP70 DNA Vaccines against Visceral Leishmaniasis in Experimental Mouse Model. PLoS ONE 2009, 4, e7880. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Be, I.; da Silva Zardo, R.; de Souza, E.P.; Borja-Cabrera, G.P.; Rosado-Vallado, M.; Mut-Martin, M.; del Rosario García-Miss, M.; de Sousa, C.B.P.; Dumonteil, E. Cross-Protective Efficacy of a Prophylactic Leishmania Donovani DNA Vaccine against Visceral and Cutaneous Murine Leishmaniasis. Infect. Immun. 2005, 73, 812–819. [Google Scholar] [CrossRef] [PubMed]
- Gamboa-León, R.; Paraguai de Souza, E.; Borja-Cabrera, G.P.; Santos, F.N.; Myashiro, L.M.; Pinheiro, R.O.; Dumonteil, E.; Palatnik-de-Sousa, C.B. Immunotherapy against Visceral Leishmaniasis with the Nucleoside Hydrolase-DNA Vaccine of Leishmania Donovani. Vaccine 2006, 24, 4863–4873. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Freier, A.; Boussoffara, T.; Das, S.; Oswald, D.; Losch, F.O.; Selka, M.; Sacerdoti-Sierra, N.; Schönian, G.; Wiesmüller, K.H.; et al. Modular Multiantigen T Cell Epitope-Enriched DNA Vaccine against Human Leishmaniasis. Sci. Transl. Med. 2014, 6, 234ra56. [Google Scholar] [CrossRef]
- Rodríguez-Cortés, A.; Ojeda, A.; López-Fuertes, L.; Timón, M.; Altet, L.; Solano-Gallego, L.; Sánchez-Robert, E.; Francino, O.; Alberola, J. Vaccination with Plasmid DNA Encoding KMPII, TRYP, LACK and GP63 Does Not Protect Dogs against Leishmania Infantum Experimental Challenge. Vaccine 2007, 25, 7962–7971. [Google Scholar] [CrossRef]
- Fragaki, K.; Suffia, I.; Ferrua, B.; Rousseau, D.; Le Fichoux, Y.; Kubar, J. Immunisation with DNA Encoding Leishmania Infantum Protein PapLe22 Decreases the Frequency of Parasitemic Episodes in Infected Hamsters. Vaccine 2001, 19, 1701–1709. [Google Scholar] [CrossRef]
- Guha, R.; Gupta, D.; Rastogi, R.; Vikram, R.; Krishnamurthy, G.; Bimal, S.; Roy, S.; Mukhopadhyay, A. Vaccination with Leishmania Hemoglobin Receptor-Encoding DNA Protects against Visceral Leishmaniasis. Sci. Transl. Med. 2013, 5, 202ra121. [Google Scholar] [CrossRef]
- Lakshmi, B.S.; Wang, R.; Madhubala, R. Leishmania Genome Analysis and High-Throughput Immunological Screening Identifies Tuzin as a Novel Vaccine Candidate against Visceral Leishmaniasis. Vaccine 2014, 32, 3816–3822. [Google Scholar] [CrossRef]
- Tewary, P.; Jain, M.; Sahani, M.H.; Saxena, S.; Madhubala, R. A Heterologous Prime-Boost Vaccination Regimen Using ORFF DNA and Recombinant ORFF Protein Confers Protective Immunity against Experimental Visceral Leishmaniasis. J. Infect. Dis. 2005, 191, 2130–2137. [Google Scholar] [CrossRef]
- Rafati, S.; Zahedifard, F.; Nazgouee, F. Prime-Boost Vaccination Using Cysteine Proteinases Type I and II of Leishmania Infantum Confers Protective Immunity in Murine Visceral Leishmaniasis. Vaccine 2006, 24, 2169–2175. [Google Scholar] [CrossRef]
- Masih, S.; Arora, S.K.; Vasishta, R.K. Efficacy of Leishmania Donovani Ribosomal P1 Gene as DNA Vaccine in Experimental Visceral Leishmaniasis. Exp. Parasitol. 2011, 129, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Saljoughian, N.; Taheri, T.; Zahedifard, F.; Taslimi, Y.; Doustdari, F.; Bolhassani, A.; Doroud, D.; Azizi, H.; Heidari, K.; Vasei, M.; et al. Development of Novel Prime-Boost Strategies Based on a Tri-Gene Fusion Recombinant L. tarentolae Vaccine against Experimental Murine Visceral Leishmaniasis. PLoS Negl. Trop. Dis. 2013, 7, e2174. [Google Scholar] [CrossRef] [PubMed]
- Dondji, B.; Pérez-Jimenez, E.; Goldsmith-Pestana, K.; Esteban, M.; McMahon-Pratt, D. Heterologous Prime-Boost Vaccination with the LACK Antigen Protects against Murine Visceral Leishmaniasis. Infect. Immun. 2005, 73, 5286–5289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dondji, B.; Deak, E.; Goldsmith-Pestana, K.; Pererz-Jimenez, E.; Esteban, M.; Miyake, S.; Yamamura, T.; McMahon-Pratt, D. Intradermal NKT Cell Activation during DNA Priming in Heterologous Prime-Boost Vaccination Enhances T Cell Responses and Protection against Leishmania. Eur. J. Immunol. 2008, 38, 706–719. [Google Scholar] [CrossRef]
- Jayakumar, A.; Castilho, T.M.; Park, E.; Goldsmith-Pestana, K.; Blackwell, J.M.; McMahon-Pratt, D. TLR1/2 Activation during Heterologous Prime-Boost Vaccination (DNA-MVA) Enhances CD8+ T Cell Responses Providing Protection against Leishmania (Viannia). PLoS Negl. Trop. Dis. 2011, 5, e1204. [Google Scholar] [CrossRef]
- Ramos, I.; Alonso, A.; Peris, A.; Marcen, J.M.; Abengozar, M.A.; Alcolea, P.J.; Castillo, J.A.; Larraga, V. Antibiotic Resistance Free Plasmid DNA Expressing LACK Protein Leads towards a Protective Th1 Response against Leishmania Infantum Infection. Vaccine 2009, 27, 6695–6703. [Google Scholar] [CrossRef]
- Bayih, A.G.; Daifalla, N.S.; Gedamu, L. DNA-Protein Immunization Using Leishmania Peroxidoxin-1 Induces a Strong CD4+ T Cell Response and Partially Protects Mice from Cutaneous Leishmaniasis: Role of Fusion Murine Granulocyte-Macrophage Colony-Stimulating Factor DNA Adjuvant. PLoS Negl. Trop. Dis. 2014, 8, e3391. [Google Scholar] [CrossRef]
- Tsagozis, P.; Karagouni, E.; Dotsika, E. Dendritic Cells Pulsed with Peptides of Gp63 Induce Differential Protection against Experimental Cutaneous Leishmaniasis. Int. J. Immunopathol. Pharmacol. 2004, 17, 343–352. [Google Scholar] [CrossRef]
- Remer, K.A.; Apetrei, C.; Schwarz, T.; Linden, C.; Moll, H. Vaccination with Plasmacytoid Dendritic Cells Induces Protection against Infection with Leishmania Major in Mice. Eur. J. Immunol. 2007, 37, 2463–2473. [Google Scholar] [CrossRef]
- Ahuja, S.S.; Reddick, R.L.; Sato, N.; Montalbo, E.; Kostecki, V.; Zhao, W.; Dolan, M.J.; Melby, P.C.; Ahuja, S.K. Dendritic Cell (DC)-Based Anti-Infective Strategies: DCs Engineered to Secrete IL-12 Are a Potent Vaccine in a Murine Model of an Intracellular Infection. J. Immunol. 1999, 163, 3890–3897. [Google Scholar]
- Agallou, M.; Margaroni, M.; Karagouni, E. Cellular Vaccination with Bone Marrow-Derived Dendritic Cells Pulsed with a Peptide of Leishmania Infantum KMP-11 and CpG Oligonucleotides Induces Protection in a Murine Model of Visceral Leishmaniasis. Vaccine 2011, 29, 5053–5064. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, M.; Pal, C.; Ray, M.; Maitra, S.; Mandal, L.; Bandyopadhyay, S. Dendritic Cell-Based Immunotherapy Combined with Antimony-Based Chemotherapy Cures Established Murine Visceral Leishmaniasis. J. Immunol. 2003, 170, 5625–5629. [Google Scholar] [CrossRef] [PubMed]
- Basu, R.; Bhaumik, S.; Haldar, A.K.; Naskar, K.; De, T.; Dana, S.K.; Walden, P.; Roy, S. Hybrid Cell Vaccination Resolves Leishmania Donovani Infection by Eliciting a Strong CD8+ Cytotoxic T-Lymphocyte Response with Concomitant Suppression of Interleukin-10 (IL-10) but Not IL-4 or IL-13. Infect. Immun. 2007, 75, 5956–5966. [Google Scholar] [CrossRef] [PubMed]
- Juncker, A.S.; Larsen, M.V.; Weinhold, N.; Nielsen, M.; Brunak, S.; Lund, O. Systematic Characterisation of Cellular Localisation and Expression Profiles of Proteins Containing MHC Ligands. PLoS ONE 2009, 4, e7448. [Google Scholar] [CrossRef]
- Bassani-Sternberg, M.; Pletscher-Frankild, S.; Jensen, L.J.; Mann, M. Mass Spectrometry of Human Leukocyte Antigen Class I Peptidomes Reveals Strong Effects of Protein Abundance and Turnover on Antigen Presentation. Mol. Cell. Proteom. 2015, 14, 658–673. [Google Scholar] [CrossRef] [Green Version]
- Wolfram, M.; Ilg, T.; Mottram, J.C.; Overath, P. Antigen Presentation by Leishmania Mexicana-infected Macrophages: Activation of Helper T Cells Specific for Amastigote Cysteine Proteinases Requires Intracellular Killing of the Parasites. Eur. J. Immunol. 1995, 25, 1094–1100. [Google Scholar] [CrossRef]
- Wolfram, M.; Fuchs, M.; Wiese, M.; Stierhof, Y.; Overath, P. Antigen Presentation by Leishmania Mexicana-infected Macrophages: Activation of Helper T Cells by a Model Parasite Antigen Secreted into the Parasitophorous Vacuole or Expressed on the Amastigote Surface. Eur. J. Immunol. 1996, 26, 3153–3162. [Google Scholar] [CrossRef]
- Kima, P.E.; Soong, L.; Chicharro, C.; Ruddle, N.H.; McMahon-Pratt, D. Leishmania-infected Macrophages Sequester Endogenously Synthesized Parasite Antigens from Presentation to CD4+ T Cells. Eur. J. Immunol. 1996, 26, 3163–3169. [Google Scholar] [CrossRef]
- Prickett, S.; Gray, P.M.; Colpitts, S.L.; Scott, P.; Kaye, P.M.; Smith, D.F. In Vivo Recognition of Ovalbumin Expressed by Transgenic Leishmania Is Determined by Its Subcellular Localization. J. Immunol. 2006, 176, 4826–4833. [Google Scholar] [CrossRef]
- Ueno, N.; Wilson, M.E. Receptor-Mediated Phagocytosis of Leishmania: Implications for Intracellular Survival. Trends Parasitol. 2012, 28, 335–344. [Google Scholar] [CrossRef]
- Wang, T.T.; Maamary, J.; Tan, G.S.; Bournazos, S.; Davis, C.W.; Krammer, F.; Schlesinger, S.J.; Palese, P.; Ahmed, R.; Ravetch, J.V. Anti-HA Glycoforms Drive B Cell Affinity Selection and Determine Influenza Vaccine Efficacy. Cell 2015, 162, 160–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stäger, S.; Alexander, J.; Kirby, A.C.; Botto, M.; Van Rooijen, N.; Smith, D.F.; Brombacher, F.; Kaye, P.M. Natural Antibodies and Complement Are Endogenous Adjuvants for Vaccine-Induced CD8+ T-Cell Responses. Nat. Med. 2003, 9, 1287–1292. [Google Scholar] [CrossRef] [PubMed]
- Dameshghi, S.; Zavaran-Hosseini, A.; Soudi, S.; Shirazi, F.J.; Nojehdehi, S.; Hashemi, S.M. Mesenchymal Stem Cells Alter Macrophage Immune Responses to Leishmania Major Infection in Both Susceptible and Resistance Mice. Immunol. Lett. 2016, 170, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Verreck, F.A.W.; De Boer, T.; Langenberg, D.M.L.; Hoeve, M.A.; Kramer, M.; Vaisberg, E.; Kastelein, R.; Kolk, A.; De Waal-Malefyt, R.; Ottenhoff, T.H.M. Human IL-23-Producing Type 1 Macrophages Promote but IL-10-Producing Type 2 Macrophages Subvert Immunity to (Myco)Bacteria. Proc. Natl. Acad. Sci. USA 2004, 101, 4560–4565. [Google Scholar] [CrossRef]
- Duque, G.A.; Fukuda, M.; Turco, S.J.; Stäger, S.; Descoteaux, A. Leishmania Promastigotes Induce Cytokine Secretion in Macrophages through the Degradation of Synaptotagmin XI. J. Immunol. 2014, 193, 2363–2372. [Google Scholar] [CrossRef]
- Vinet, A.F.; Fukuda, M.; Turco, S.J.; Descoteaux, A. The Leishmania Donovani Lipophosphoglycan Excludes the Vesicular Proton-ATPase from Phagosomes by Impairing the Recruitment of Synaptotagmin V. PLoS Pathog. 2009, 5, e1000628. [Google Scholar] [CrossRef]
- Forget, G.; Gregory, D.J.; Olivier, M. Proteasome-Mediated Degradation of STAT1α Following Infection of Macrophages with Leishmania Donovani. J. Biol. Chem. 2005, 280, 30542–30549. [Google Scholar] [CrossRef]
- Späth, G.F.; Schlesinger, P.; Schreiber, R.; Beverley, S.M. A Novel Role for Stat1 in Phagosome Acidification and Natural Host Resistance to Intracellular Infection by Leishmania Major. PLoS Pathog. 2009, 5, e1000381. [Google Scholar] [CrossRef]
- Hsieh, C.-S.; deRoos, P.; Honey, K.; Beers, C.; Rudensky, A.Y. A Role for Cathepsin L and Cathepsin S in Peptide Generation for MHC Class II Presentation. J. Immunol. 2002, 168, 2618–2625. [Google Scholar] [CrossRef]
- Rossman, M.D.; Maida, B.T.; Douglas, S.D. Monocyte-Derived Macrophage and Alveolar Macrophage Fibronectin Production and Cathepsin D Activity. Cell. Immunol. 1990, 126, 268–277. [Google Scholar] [CrossRef]
- Kropf, P.; Freudenberg, M.A.; Modolell, M.; Price, H.P.; Herath, S.; Antoniazi, S.; Galanos, C.; Smith, D.F.; Müller, I. Toll-like Receptor 4 Contributes to Efficient Control of Infection with the Protozoan Parasite Leishmania Major. Infect. Immun. 2004, 72, 1920–1928. [Google Scholar] [CrossRef] [PubMed]
- Liuzzo, J.P.; Petanceska, S.S.; Moscatelli, D.; Devi, L.A. Inflammatory Mediators Regulate Cathepsin S in Macrophages and Microglia: A Role in Attenuating Heparan Sulfate Interactions. Mol. Med. 1999, 5, 320–333. [Google Scholar] [CrossRef] [PubMed]
- Blum, J.S.; Wearsch, P.A.; Cresswell, P. Pathways of Antigen Processing. Annu. Rev. Immunol. 2013, 31, 443–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antoine, J.C.; Lang, T.; Prina, E.; Courret, N.; Hellio, R. H-2M Molecules, like MHC Class II Molecules, Are Targeted to Parasitophorous Vacuoles of Leishmania-Infected Macrophages and Internalized by Amastigotes of L. Amazonensis and L. Mexicana. J. Cell Sci. 1999, 112, 2559–2570. [Google Scholar] [PubMed]
- Sercarz, E.E.; Lehmann, P.V.; Ametani, A.; Benichou, G.; Miller, A.; Moudgil, K. Dominance and Crypticity of T Cell Antigenic Determinants. Annu. Rev. Immunol. 1993, 11, 729–766. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; McCluskey, J. Immunodominance and Immunodomination: Critical Factors in Developing Effective CD8+ T-Cell–Based Cancer Vaccines. Adv. Cancer Res. 2006, 95, 203–247. [Google Scholar] [PubMed]
- Luciani, F.; Sanders, M.T.; Oveissi, S.; Pang, K.C.; Chen, W. Increasing Viral Dose Causes a Reversal in CD8 + T Cell Immunodominance during Primary Influenza Infection Due to Differences in Antigen Presentation, T Cell Avidity, and Precursor Numbers. J. Immunol. 2013, 190, 36–47. [Google Scholar] [CrossRef]
- Egui, A.; Ledesma, D.; Pérez-Antón, E.; Montoya, A.; Gómez, I.; Robledo, S.M.; Infante, J.J.; Vélez, I.D.; Lopez, M.C.; Thomas, M.C. Phenotypic and Functional Profiles of Antigen-Specific CD4+ and CD8+ T Cells Associated with Infection Control in Patients with Cutaneous Leishmaniasis. Front. Cell. Infect. Microbiol. 2018, 8, 393. [Google Scholar] [CrossRef]
- Budd, R.C.; Cerottini, J.C.; Horvath, C.; Bron, C.; Pedrazzini, T.; Howe, R.C.; MacDonald, H.R. Distinction of Virgin and Memory T Lymphocytes. Stable Acquisition of the Pgp-1 Glycoprotein Concomitant with Antigenic Stimulation. J. Immunol. 1987, 138, 3120–3129. [Google Scholar]
- Mackay, C.R.; Marston, W.L.; Dudler, L. Naive and Memory T Cells Show Distinct Pathways of Lymphocyte Recirculation. J. Exp. Med. 1990, 171, 801–817. [Google Scholar] [CrossRef]
- Nyambura, L.W.; Jarmalavicius, S.; Walden, P. Impact of Leishmania Donovani Infection on the HLA I Self Peptide Repertoire of Human Macrophages. PLoS ONE 2018, 13, e0200297. [Google Scholar] [CrossRef] [PubMed]
- Roy, K.; Naskar, K.; Ghosh, M.; Roy, S. Class II MHC/Peptide Interaction in Leishmania Donovani Infection: Implications in Vaccine Design. J. Immunol. 2014, 192, 5873–5880. [Google Scholar] [CrossRef] [PubMed]
- Weaver, J.M.; Lazarski, C.A.; Richards, K.A.; Chaves, F.A.; Jenks, S.A.; Menges, P.R.; Sant, A.J. Immunodominance of CD4 T Cells to Foreign Antigens Is Peptide Intrinsic and Independent of Molecular Context: Implications for Vaccine Design. J. Immunol. 2008, 181, 3039–3048. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Williams, K.P.; Chang, Y.-H.; Smith, J.A. Immunodominance: A Single Amino Acid Substitution within an Antigenic Site Alters Intramolecular Selection of T Cell Determinants. J. Immunol. 1993, 151, 1852–1858. [Google Scholar]
- Mazor, R.; Vassall, A.N.; Eberle, J.A.; Beers, R.; Weldon, J.E.; Venzon, D.J.; Tsang, K.Y.; Benhar, I.; Pastan, I. Identification and Elimination of an Immunodominant T-Cell Epitope in Recombinant Immunotoxins Based on Pseudomonas Exotoxin A. Proc. Natl. Acad. Sci. USA 2012, 109, E3597–E3603. [Google Scholar] [CrossRef] [PubMed]
- Mazor, R.; Eberle, J.A.; Hu, X.; Vassall, A.N.; Onda, M.; Beers, R.; Lee, E.C.; Kreitman, R.J.; Lee, B.; Baker, D. Recombinant Immunotoxin for Cancer Treatment with Low Immunogenicity by Identification and Silencing of Human T-Cell Epitopes. Proc. Natl. Acad. Sci. USA 2014, 111, 8571–8576. [Google Scholar] [CrossRef] [PubMed]
- Schneider, S.C.; Ohmen, J.; Fosdick, L.; Gladstone, B.; Guo, J.; Ametani, A.; Sercarz, E.E.; Deng, H. Cutting Edge: Introduction of an Endopeptidase Cleavage Motif into a Determinant Flanking Region of Hen Egg Lysozyme Results in Enhanced T Cell Determinant Display. J. Immunol. 2000, 165, 20–23. [Google Scholar] [CrossRef] [Green Version]
- Moss, D.L.; Park, H.-W.; Mettu, R.R.; Landry, S.J. Deimmunizing Substitutions in Pseudomonas Exotoxin Domain III Perturb Antigen Processing without Eliminating T-Cell Epitopes. J. Biol. Chem. 2019, 294, 4667–4681. [Google Scholar] [CrossRef]
- Kim, A.; Boronina, T.N.; Cole, R.N.; Darrah, E.; Sadegh-Nasseri, S. Distorted Immunodominance by Linker Sequences or Other Epitopes from a Second Protein Antigen during Antigen-Processing. Sci. Rep. 2017, 7, 46418. [Google Scholar] [CrossRef]
- Laffitte, M.-C.N.; Leprohon, P.; Papadopoulou, B.; Ouellette, M. Plasticity of the Leishmania Genome Leading to Gene Copy Number Variations and Drug Resistance. F1000Research 2016, 5, 2350. [Google Scholar] [CrossRef]
- Iantorno, S.A.; Durrant, C.; Khan, A.; Sanders, M.J.; Beverley, S.M.; Warren, W.C.; Berriman, M.; Sacks, D.L.; Cotton, J.A.; Grigg, M.E. Gene Expression in Leishmania Is Regulated Predominantly by Gene Dosage. MBio 2017, 8, e01393-17. [Google Scholar] [CrossRef]
- Monks, C.R.F.; Freiberg, B.A.; Kupfer, H.; Sciaky, N.; Kupfer, A. Three-Dimensional Segregation of Supramolecular Activation Clusters in T Cells. Nature 1998, 395, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Grakoui, A.; Bromley, S.K.; Sumen, C.; Davis, M.M.; Shaw, A.S.; Allen, P.M.; Dustin, M.L. The Immunological Synapse: A Molecular Machine Controlling T Cell Activation. Science 1999, 285, 221–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosseini, B.H.; Louban, I.; Djandji, D.; Wabnitz, G.H.; Deeg, J.; Bulbuc, N.; Samstag, Y.; Gunzer, M.; Spatz, J.P.; Hämmerling, G.J. Immune Synapse Formation Determines Interaction Forces between T Cells and Antigen-Presenting Cells Measured by Atomic Force Microscopy. Proc. Natl. Acad. Sci. USA 2009, 106, 17852–17857. [Google Scholar] [CrossRef] [PubMed]
- Dustin, M.L.; Bromley, S.K.; Kan, Z.; Peterson, D.A.; Unanue, E.R. Antigen Receptor Engagement Delivers a Stop Signal to Migrating T Lymphocytes. Proc. Natl. Acad. Sci. USA 1997, 94, 3909–3913. [Google Scholar] [CrossRef]
- Park, E.J.; Peixoto, A.; Imai, Y.; Goodarzi, A.; Cheng, G.; Carman, C.V.; von Andrian, U.H.; Shimaoka, M. Distinct Roles for LFA-1 Affinity Regulation during T-Cell Adhesion, Diapedesis, and Interstitial Migration in Lymph Nodes. Blood 2010, 115, 1572–1581. [Google Scholar] [CrossRef]
- Zhang, W.; Trible, R.P.; Samelson, L.E. LAT Palmitoylation: Its Essential Role in Membrane Microdomain Targeting and Tyrosine Phosphorylation during T Cell Activation. Immunity 1998, 9, 239–246. [Google Scholar] [CrossRef] [Green Version]
- Bi, K.; Tanaka, Y.; Coudronniere, N.; Sugie, K.; Hong, S.; van Stipdonk, M.J.B.; Altman, A. Antigen-Induced Translocation of PKC-θ to Membrane Rafts Is Required for T Cell Activation. Nat. Immunol. 2001, 2, 556–563. [Google Scholar] [CrossRef]
- Molina, T.J.; Kishihara, K.; Siderovskid, D.P.; Van Ewijk, W.; Narendran, A.; Timms, E.; Wakeham, A.; Paige, C.J.; Hartmann, K.U.; Veillette, A.; et al. Profound Block in Thymocyte Development in Mice Lacking P56lck. Nature 1992, 357, 161–164. [Google Scholar] [CrossRef]
- Rub, A.; Dey, R.; Jadhav, M.; Kamat, R.; Chakkaramakkil, S.; Majumdar, S.; Mukhopadhyaya, R.; Saha, B. Cholesterol Depletion Associated with Leishmania Major Infection Alters Macrophage CD40 Signalosome Composition and Effector Function. Nat. Immunol. 2009, 10, 273–280. [Google Scholar] [CrossRef]
- Meier, C.L.; Svensson, M.; Kaye, P.M. Leishmania -Induced Inhibition of Macrophage Antigen Presentation Analyzed at the Single-Cell Level. J. Immunol. 2003, 171, 6706–6713. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, J.; Das, S.; Guha, R.; Ghosh, D.; Naskar, K.; Das, A.; Roy, S. Hyperlipidemia Offers Protection against Leishmania Donovani Infection: Role of Membrane Cholesterol. J. Lipid Res. 2012, 53, 2560–2572. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, I.; Gatfield, J.; Mini, T.; Jeno, P.; Pieters, J. Essential Role for Cholesterol in the Delivery of Exogenous Antigens to the MHC Class I-Presentation Pathway. Int. Immunol. 2006, 18, 755–765. [Google Scholar] [CrossRef] [PubMed]
- Roy, K.; Mandloi, S.; Chakrabarti, S.; Roy, S. Cholesterol Corrects Altered Conformation of MHC-II Protein in Leishmania Donovani Infected Macrophages: Implication in Therapy. PLoS Negl. Trop. Dis. 2016, 10, e0004710. [Google Scholar] [CrossRef]
- Semini, G.; Paape, D.; Paterou, A.; Schroeder, J.; Barrios-Llerena, M.; Aebischer, T. Changes to Cholesterol Trafficking in Macrophages by Leishmania Parasites Infection. Microbiologyopen 2017, 6, e00469. [Google Scholar] [CrossRef]
- Banerjee, S.; Ghosh, J.; Sen, S.; Guha, R.; Dhar, R.; Ghosh, M.; Datta, S.; Raychaudhury, B.; Naskar, K.; Haldar, A.K.; et al. Designing Therapies against Experimental Visceral Leishmaniasis by Modulating the Membrane Fluidity of Antigen-Presenting Cells. Infect. Immun. 2009, 77, 2330–2342. [Google Scholar] [CrossRef] [Green Version]
- Reiner, N.E.; Ng, W.; McMaster, W.R. Parasite-Accessory Cell Interactions in Murine Leishmaniasis. II. Leishmania Donovani Suppresses Macrophage Expression of Class I and Class II Major Histocompatibility Complex Gene Products. J. Immunol. 1987, 138, 1926–1932. [Google Scholar]
- Muraille, E.; De Trez, C.; Pajak, B.; Torrentera, F.A.; De Baetselier, P.; Leo, O.; Carlier, Y. Amastigote Load and Cell Surface Phenotype of Infected Cells from Lesions and Lymph Nodes of Susceptible and Resistant Mice Infected with Leishmania Major. Infect. Immun. 2003, 71, 2704–2715. [Google Scholar] [CrossRef]
- Silverman, J.M.; Clos, J.; Horakova, E.; Wang, A.Y.; Wiesgigl, M.; Kelly, I.; Lynn, M.A.; McMaster, W.R.; Foster, L.J.; Levings, M.K.; et al. Leishmania Exosomes Modulate Innate and Adaptive Immune Responses through Effects on Monocytes and Dendritic Cells. J. Immunol. 2010, 185, 5011–5022. [Google Scholar] [CrossRef] [Green Version]
- Pinheiro, N.F.; Hermida, M.D.R.; Macedo, M.P.; Mengel, J.; Bafica, A.; dos-Santos, W.L.C. Leishmania Infection Impairs Β1-Integrin Function and Chemokine Receptor Expression in Mononuclear Phagocytes. Infect. Immun. 2006, 74, 3912–3921. [Google Scholar] [CrossRef]
- Rosenthal, L.A.; Sutterwala, F.S.; Kehrli, M.E.; Mosser, D.M. Leishmania Major-Human Macrophage Interactions: Cooperation between Mac-1 (CD11b/CD18) and Complement Receptor Type 1 (CD35) in Promastigote Adhesion. Infect. Immun. 1996, 64, 2206–2215. [Google Scholar] [PubMed]
- de Menezes, J.P.B.; Koushik, A.; Das, S.; Guven, C.; Siegel, A.; Laranjeira-Silva, M.F.; Losert, W.; Andrews, N.W. Leishmania Infection Inhibits Macrophage Motility by Altering F-Actin Dynamics and the Expression of Adhesion Complex Proteins. Cell. Microbiol. 2017, 19, e12668. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, M.F.; Sebzda, E.; Kündig, T.M.; Shahinian, A.; Speiser, D.E.; Mak, T.W.; Ohashi, P.S. T Cell Responses Are Governed by Avidity and Co-Stimulatory Thresholds. Eur. J. Immunol. 1996, 26, 2017–2022. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.A.; Titus, R.G.; Nabavi, N.; Glimcher, L.H. Blockade of CD86 Ameliorates Leishmania Major Infection by Down- Regulating the Th2 Response. J. Infect. Dis. 1996, 174, 1303–1308. [Google Scholar] [CrossRef]
- Li, J.G.; Du, Y.M.; Yan, Z.D.; Yan, J.; Zhuansun, Y.X.; Chen, R.; Zhang, W.; Feng, S.L.; Ran, P.X. CD80 and CD86 Knockdown in Dendritic Cells Regulates Th1/Th2 Cytokine Production in Asthmatic Mice. Exp. Ther. Med. 2016, 11, 878–884. [Google Scholar] [CrossRef]
- Brodskyn, C.I.; DeKrey, G.K.; Titus, R.G. Influence of Costimulatory Molecules on Immune Response to Leishmania Major by Human Cells in Vitro. Infect. Immun. 2001, 69, 665–672. [Google Scholar] [CrossRef]
- Freedman, A.S.; Freeman, G.J.; Rhynhart, K.; Nadler, L.M. Selective Induction of B7/BB-1 on Interferon-γ Stimulated Monocytes: A Potential Mechanism for Amplification of T Cell Activation through the CD28 Pathway. Cell. Immunol. 1991, 137, 429–437. [Google Scholar] [CrossRef]
- Kiener, P.A.; Moran-Davis, P.; Rankin, B.M.; Wahl, A.F.; Aruffo, A.; Hollenbaugh, D. Stimulation of CD40 with Purified Soluble Gp39 Induces Proinflammatory Responses in Human Monocytes. J. Immunol. 1995, 155, 4917–4925. [Google Scholar]
- Caux, C.; Massacrier, C.; Vanbervliet, B.; Dubois, B.; Van Kooten, C.; Durand, I.; Banchereau, J. Activation of Human Dendritic Cells through CD40 Cross-Linking. J. Exp. Med. 1994, 180, 1263–1272. [Google Scholar] [CrossRef]
- Saha, B.; Das, G.; Vohra, H.; Ganguly, N.K.; Mishra, G.C. Macrophage-T Cell Interaction in Experimental Visceral Leishmaniasis: Failure to Express Costimulatory Molecules on Leishmania-infected Macrophages and Its Implication in the Suppression of Cell-mediated Immunity. Eur. J. Immunol. 1995, 25, 2492–2498. [Google Scholar] [CrossRef]
- Walunas, T.L.; Lenschow, D.J.; Bakker, C.Y.; Linsley, P.S.; Freeman, G.J.; Green, J.M.; Thompson, C.B.; Bluestone, J.A. CTLA-4 Can Function as a Negative Regulator of T Cell Activation. Immunity 1994, 1, 405–413. [Google Scholar] [CrossRef]
- Perez, V.L.; Van Parijs, L.; Biuckians, A.; Zheng, X.X.; Strom, T.B.; Abbas, A.K. Induction of Peripheral T Cell Tolerance in Vivo Requires CTLA-4 Engagement. Immunity 1997, 6, 411–417. [Google Scholar] [CrossRef]
- Lindsten, T.; Lee, K.P.; Harris, E.S.; Petryniak, B.; Craighead, N.; Reynolds, P.J.; Lombard, D.B.; Freeman, G.J.; Nadler, L.M.; Gray, G.S. Characterization of CTLA-4 Structure and Expression on Human T Cells. J. Immunol. 1993, 151, 3489–3499. [Google Scholar] [PubMed]
- Kaye, P.M.; Murphy, M.L.; Engwerda, C.R.; Gorak, P.M.A. B7-2 Blockade Enhances T Cell Responses to Leishmania Donovani. J. Immunol. 1997, 159, 4460–4466. [Google Scholar]
- Murphy, M.L.; Cotterell, S.E.; Gorak, P.M.; Engwerda, C.R.; Kaye, P.M. Blockade of CTLA-4 Enhances Host Resistance to the Intracellular Pathogen, Leishmania Donovani. J. Immunol. 1998, 161, 4153–4160. [Google Scholar]
- Sheppard, K.A.; Fitz, L.J.; Lee, J.M.; Benander, C.; George, J.A.; Wooters, J.; Qiu, Y.; Jussif, J.M.; Carter, L.L.; Wood, C.R.; et al. PD-1 Inhibits T-Cell Receptor Induced Phosphorylation of the ZAP70/CD3ζ Signalosome and Downstream Signaling to PKCθ. FEBS Lett. 2004, 574, 37–41. [Google Scholar] [CrossRef]
- Roy, S.; Gupta, P.; Palit, S.; Basu, M.; Ukil, A.; Das, P.K. The Role of PD-1 in Regulation of Macrophage Apoptosis and Its Subversion by Leishmania Donovani. Clin. Transl. Immunol. 2017, 6, e137. [Google Scholar] [CrossRef]
- Abe, T.; Tanaka, Y.; Piao, J.; Tanimine, N.; Oue, N.; Hinoi, T.; Garcia, N.V.; Miyasaka, M.; Matozaki, T.; Yasui, W.; et al. Signal Regulatory Protein Alpha Blockade Potentiates Tumoricidal Effects of Macrophages on Gastroenterological Neoplastic Cells in Syngeneic Immunocompetent Mice. Ann. Gastroenterol. Surg. 2018, 2, 451–462. [Google Scholar] [CrossRef]
- Ontoria, E.; Hernández-Santana, Y.E.; González-García, A.C.; López, M.C.; Valladares, B.; Carmelo, E. Transcriptional Profiling of Immune-Related Genes in Leishmania Infantum-Infected Mice: Identification of Potential Biomarkers of Infection and Progression of Disease. Front. Cell. Infect. Microbiol. 2018, 8, 197. [Google Scholar] [CrossRef]
- Sanderson, N.S.R.; Puntel, M.; Kroeger, K.M.; Bondale, N.S.; Swerdlow, M.; Iranmanesh, N.; Yagita, H.; Ibrahim, A.; Castro, M.G.; Lowenstein, P.R. Cytotoxic Immunological Synapses Do Not Restrict the Action of Interferon-γ to Antigenic Target Cells. Proc. Natl. Acad. Sci. USA 2012, 109, 7835–7840. [Google Scholar] [CrossRef]
- Huse, M.; Lillemeier, B.F.; Kuhns, M.S.; Chen, D.S.; Davis, M.M. T Cells Use Two Directionally Distinct Pathways for Cytokine Secretion. Nat. Immunol. 2006, 7, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Chemin, K.; Bohineust, A.; Dogniaux, S.; Tourret, M.; Guégan, S.; Miro, F.; Hivroz, C. Cytokine Secretion by CD4+ T Cells at the Immunological Synapse Requires Cdc42-Dependent Local Actin Remodeling but Not Microtubule Organizing Center Polarity. J. Immunol. 2012, 189, 2159–2168. [Google Scholar] [CrossRef] [PubMed]
- Fooksman, D.R.; Vardhana, S.; Vasiliver-Shamis, G.; Liese, J.; Blair, D.A.; Waite, J.; Sacristán, C.; Victora, G.D.; Zanin-Zhorov, A.; Dustin, M.L. Functional Anatomy of T Cell Activation and Synapse Formation. Annu. Rev. Immunol. 2010, 28, 79–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Philipsen, L.; Engels, T.; Schilling, K.; Gurbiel, S.; Fischer, K.D.; Tedford, K.; Schraven, B.; Gunzer, M.; Reichardt, P. Multimolecular Analysis of Stable Immunological Synapses Reveals Sustained Recruitment and Sequential Assembly of Signaling Clusters. Mol. Cell. Proteom. 2013, 12, 2551–2567. [Google Scholar] [CrossRef] [Green Version]
- Malherbe, L.; Filippi, C.; Julia, V.; Foucras, G.; Moro, M.; Appel, H.; Wucherpfennig, K.; Guéry, J.C.; Glaichenhaus, N. Selective Activation and Expansion of High-Affinity CD4+ T Cells in Resistant Mice upon Infection with Leishmania Major. Immunity 2000, 13, 771–782. [Google Scholar] [CrossRef]
- Pentcheva-Hoang, T.; Egen, J.G.; Wojnoonski, K.; Allison, J.P. B7-1 and B7-2 Selectively Recruit CTLA-4 and CD28 to the Immunological Synapse. Immunity 2004, 21, 401–413. [Google Scholar] [CrossRef] [Green Version]
- Gannavaram, S.; Bhattacharya, P.; Ismail, N.; Kaul, A.; Singh, R.; Nakhasi, H.L. Modulation of Innate Immune Mechanisms to Enhance Leishmania Vaccine-Induced Immunity: Role of Coinhibitory Molecules. Front. Immunol. 2016, 7, 187. [Google Scholar] [CrossRef] [Green Version]
- Gomes, N.A.; Gattass, C.R.; Barreto-de-Souza, V.; Wilson, M.E.; DosReis, G.A. TGF-β Mediates CTLA-4 Suppression of Cellular Immunity in Murine Kalaazar. J. Immunol. 2000, 164, 2001–2008. [Google Scholar] [CrossRef]
- He, X.; Holmes, T.H.; Mahmood, K.; Kemble, G.W.; Dekker, C.L.; Arvin, A.M.; Greenberg, H.B. Phenotypic Changes in Influenza-Specific CD8 + T Cells after Immunization of Children and Adults with Influenza Vaccines. J. Infect. Dis. 2008, 197, 803–811. [Google Scholar] [CrossRef]
- Viganò, S.; Utzschneider, D.T.; Perreau, M.; Pantaleo, G.; Zehn, D.; Harari, A. Functional Avidity: A Measure to Predict the Efficacy of Effector T Cells? Clin. Dev. Immunol. 2012, 2012. [Google Scholar] [CrossRef]
- Betts, M.R.; Price, D.A.; Brenchley, J.M.; Loré, K.; Guenaga, F.J.; Smed-Sorensen, A.; Ambrozak, D.R.; Migueles, S.A.; Connors, M.; Roederer, M.; et al. The Functional Profile of Primary Human Antiviral CD8 + T Cell Effector Activity Is Dictated by Cognate Peptide Concentration. J. Immunol. 2004, 172, 6407–6417. [Google Scholar] [CrossRef] [PubMed]
- Langenkamp, A.; Casorati, G.; Garavaglia, C.; Dellabona, P.; Lanzavecchia, A.; Sallusto, F. T Cell Priming by Dendritic Cells: Thresholds for Proliferation, Differentiation and Death and Intraclonal Functional Diversification. Eur. J. Immunol. 2002, 32, 2046–2054. [Google Scholar] [CrossRef]
- Wooldridge, L.; Lissina, A.; Vernazza, J.; Gostick, E.; Laugel, B.; Hutchinson, S.L.; Mirza, F.; Dunbar, P.R.; Boulter, J.M.; Glick, M.; et al. Enhanced Immunogenicity of CTL Antigens through Mutation of the CD8 Binding MHC Class I Invariant Region. Eur. J. Immunol. 2007, 37, 1323–1333. [Google Scholar] [CrossRef] [PubMed]
- Azzam, H.S.; Grinberg, A.; Lui, K.; Shen, H.; Shores, E.W.; Love, P.E. CD5 Expression Is Developmentally Regulated by T Cell Receptor (TCR) Signals and TCR Avidity. J. Exp. Med. 1998, 188, 2301–2311. [Google Scholar] [CrossRef] [PubMed]
- Ebert, P.J.R.; Jiang, S.; Xie, J.; Li, Q.-J.; Davis, M.M. An Endogenous Positively Selecting Peptide Enhances Mature T Cell Responses and Becomes an Autoantigen in the Absence of MicroRNA MiR-181a. Nat. Immunol. 2009, 10, 1162. [Google Scholar] [CrossRef] [PubMed]
- Pircher, H.; Rohrer, U.H.; Moskophidis, D.; Zinkernagel, R.M.; Hengartner, H. Lower Receptor Avidity Required for Thymic Clonal Deletion than for Effector T-Cell Function. Nature 1991, 351, 482–485. [Google Scholar] [CrossRef]
- Liu, H.; Cao, A.T.; Feng, T.; Li, Q.; Zhang, W.; Yao, S.; Dann, S.M.; Elson, C.O.; Cong, Y. TGF-β Converts Th1 Cells into Th17 Cells through Stimulation of Runx1 Expression. Eur. J. Immunol. 2015, 45, 1010–1018. [Google Scholar] [CrossRef]
- Li, M.O.; Flavell, R.A. TGF-β: A Master of All T Cell Trades. Cell 2008, 134, 392–404. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, S.; Yamaguchi, T.; Nomura, T.; Ono, M. Regulatory T Cells and Immune Tolerance. Cell 2008, 133, 775–787. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, S.F.; Buckner, J.H. FOXP3 and the Regulation of Treg/Th17 Differentiation. Microbes Infect. 2009, 11, 594–598. [Google Scholar] [CrossRef]
- Xu, T.; Stewart, K.M.; Wang, X.; Liu, K.; Xie, M.; Ryu, J.K.; Li, K.; Ma, T.; Wang, H.; Ni, L. Metabolic Control of T H 17 and Induced T Reg Cell Balance by an Epigenetic Mechanism. Nature 2017, 548, 228. [Google Scholar] [CrossRef] [PubMed]
- Wherry, E.J. T Cell Exhaustion. Nat. Immunol. 2011, 12, 492–499. [Google Scholar] [CrossRef]
- Barber, D.L.; Wherry, E.J.; Masopust, D.; Zhu, B.; Allison, J.P.; Sharpe, A.H.; Freeman, G.J.; Ahmed, R. Restoring Function in Exhausted CD8 T Cells during Chronic Viral Infection. Nature 2006, 439, 682. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, S.D.; Shin, H.; Haining, W.N.; Zou, T.; Workman, C.J.; Polley, A.; Betts, M.R.; Freeman, G.J.; Vignali, D.A.A.; Wherry, E.J. Coregulation of CD8+ T Cell Exhaustion by Multiple Inhibitory Receptors during Chronic Viral Infection. Nat. Immunol. 2009, 10, 29. [Google Scholar] [CrossRef] [PubMed]
- Mescher, M.F.; Curtsinger, J.M.; Agarwal, P.; Casey, K.A.; Gerner, M.; Hammerbeck, C.D.; Popescu, F.; Xiao, Z. Signals Required for Programming Effector and Memory Development by CD8+ T Cells. Immunol. Rev. 2006, 211, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Tough, D.F.; Sprent, J. Turnover of Naive-and Memory-Phenotype T Cells. J. Exp. Med. 1994, 179, 1127–1135. [Google Scholar] [CrossRef]
- Munn, D.H.; Sharma, M.D.; Baban, B.; Harding, H.P.; Zhang, Y.; Ron, D.; Mellor, A.L. GCN2 Kinase in T Cells Mediates Proliferative Arrest and Anergy Induction in Response to Indoleamine 2, 3-Dioxygenase. Immunity 2005, 22, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Sitkovsky, M.V. T Regulatory Cells: Hypoxia-Adenosinergic Suppression and Re-Direction of the Immune Response. Trends Immunol. 2009, 30, 102–108. [Google Scholar] [CrossRef]
- Mueller, D.L. Mechanisms Maintaining Peripheral Tolerance. Nat. Immunol. 2010, 11, 21. [Google Scholar] [CrossRef]
- Fallarino, F.; Grohmann, U.; Puccetti, P. Indoleamine 2,3-Dioxygenase: From Catalyst to Signaling Function. Eur. J. Immunol. 2012, 42, 1932–1937. [Google Scholar] [CrossRef]
- Deaglio, S.; Dwyer, K.M.; Gao, W.; Friedman, D.; Usheva, A.; Erat, A.; Chen, J.F.; Enjyoji, K.; Linden, J.; Oukka, M.; et al. Adenosine Generation Catalyzed by CD39 and CD73 Expressed on Regulatory T Cells Mediates Immune Suppression. J. Exp. Med. 2007, 204, 1257–1265. [Google Scholar] [CrossRef] [PubMed]
- Wing, K.; Onishi, Y.; Prieto-Martin, P.; Yamaguchi, T.; Miyara, M.; Fehervari, Z.; Nomura, T.; Sakaguchi, S. CTLA-4 Control over Foxp3+ Regulatory T Cell Function. Science 2008, 322, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Makala, L.H.C.; Baban, B.; Lemos, H.; El-Awady, A.R.; Chandler, P.R.; Hou, D.Y.; Munn, D.H.; Mellor, A.L. Leishmania Major Attenuates Host Immunity by Stimulating Local Indoleamine 2,3-Dioxygenase Expression. J. Infect. Dis. 2011, 203, 715–725. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, A.B.; Serafim, T.D.; Marques-da-Silva, E.A.; Meyer-Fernandes, J.R.; Afonso, L.C.C. Leishmania Amazonensis Impairs DC Function by Inhibiting CD40 Expression via A 2B Adenosine Receptor Activation. Eur. J. Immunol. 2012, 42, 1203–1215. [Google Scholar] [CrossRef] [PubMed]
- Joshi, T.; Rodriguez, S.; Perovic, V.; Cockburn, I.A.; Stäger, S. B7-H1 Blockade Increases Survival of Dysfunctional CD8+ T Cells and Confers Protection against Leishmania Donovaniinfections. PLoS Pathog. 2009, 5, e1000431. [Google Scholar] [CrossRef]
- Pinheiro, R.O.; Pinto, E.F.; Benedito, A.B.; Lopes, U.G.; Rossi-Bergmann, B. The T-Cell Anergy Induced by Leishmania Amazonensis Antigens Is Related with Defective Antigen Presentation and Apoptosis. An. Acad. Bras. Cienc. 2004, 76, 519–527. [Google Scholar] [CrossRef]
- Krammer, P.H.; Arnold, R.; Lavrik, I.N. Life and Death in Peripheral T Cells. Nat. Rev. Immunol. 2007, 7, 532–542. [Google Scholar] [CrossRef]
- Woodland, D.L. Jump-Starting the Immune System: Prime-Boosting Comes of Age. Trends Immunol. 2004, 25, 98–104. [Google Scholar] [CrossRef]
- Masopust, D.; Ha, S.-J.; Vezys, V.; Ahmed, R. Stimulation History Dictates Memory CD8 T Cell Phenotype: Implications for Prime-Boost Vaccination. J. Immunol. 2006, 177, 831–839. [Google Scholar] [CrossRef] [Green Version]
- Wirth, T.C.; Xue, H.H.; Rai, D.; Sabel, J.T.; Bair, T.; Harty, J.T.; Badovinac, V.P. Repetitive Antigen Stimulation Induces Stepwise Transcriptome Diversification but Preserves a Core Signature of Memory CD8+ T Cell Differentiation. Immunity 2010, 33, 128–140. [Google Scholar] [CrossRef] [Green Version]
- Sallusto, F.; Lanzavecchia, A.; Araki, K.; Ahmed, R. From Vaccines to Memory and Back. Immunity 2010, 33, 451–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butler, N.S.; Nolz, J.C.; Harty, J.T. Immunologic Considerations for Generating Memory CD8 T Cells through Vaccination. Cell. Microbiol. 2011, 13, 925–933. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, V.; Cordeiro-da-Silva, A.; Laforge, M.; Ouaissi, A.; Akharid, K.; Silvestre, R.; Estaquier, J. Impairment of T Cell Function in Parasitic Infections. PLoS Negl. Trop. Dis. 2014, 8, e2567. [Google Scholar] [CrossRef] [PubMed]
- Haring, J.S.; Badovinac, V.P.; Harty, J.T. Inflaming the CD8+ T Cell Response. Immunity 2006, 25, 19–29. [Google Scholar] [CrossRef] [Green Version]
- Matte, C.; Descoteaux, A. Leishmania Donovani Amastigotes Impair Gamma Interferon-Induced STAT1α Nuclear Translocation by Blocking the Interaction between STAT1α and Importin-A5. Infect. Immun. 2010, 78, 3736–3743. [Google Scholar] [CrossRef]
- Bhardwaj, N.; Rosas, L.E.; Lafuse, W.P.; Satoskar, A.R. Leishmania Inhibits STAT1-Mediated IFN-γ Signaling in Macrophages: Increased Tyrosine Phosphorylation of Dominant Negative STAT1β by Leishmania Mexicana. Int. J. Parasitol. 2005, 35, 75–82. [Google Scholar] [CrossRef]
- Ray, M.; Gam, A.A.; Boykins, R.A.; Kenney, R.T. Inhibition of Interferon-γ Signaling by Leishmania Donovani. J. Infect. Dis. 2000, 181, 1121–1128. [Google Scholar] [CrossRef]
- Blanchette, J.; Racette, N.; Faure, R.; Siminovitch, K.A.; Olivier, M. Leishmania-Induced Increases in Activation of Macrophage SHP-1 Tyrosine Phosphatase Are Associated with Impaired IFN-γ-Triggered JAK2 Activation. Eur. J. Immunol. 1999, 29, 3737–3744. [Google Scholar] [CrossRef]
- Nandan, D.; Reiner, N.E. Attenuation of Gamma Interferon-Induced Tyrosine Phosphorylation in Mononuclear Phagocytes Infected with Leishmania Donovani: Selective Inhibition of Signaling through Janus Kinases and Stat1. Infect. Immun. 1995, 63, 4495–4500. [Google Scholar]
- Cecchinato, V.; Bernasconi, E.; Speck, R.F.; Proietti, M.; Sauermann, U.; D’Agostino, G.; Danelon, G.; Jost, T.R.; Grassi, F.; Raeli, L. Impairment of CCR6+ and CXCR3+ Th Cell Migration in HIV-1 Infection Is Rescued by Modulating Actin Polymerization. J. Immunol. 2017, 198, 184–195. [Google Scholar] [CrossRef]
- Salgado-Pabón, W.; Celli, S.; Arena, E.T.; Nothelfer, K.; Roux, P.; Sellge, G.; Frigimelica, E.; Bousso, P.; Sansonetti, P.J.; Phalipon, A. Shigella Impairs T Lymphocyte Dynamics in Vivo. Proc. Natl. Acad. Sci. USA 2013, 110, 4458–4463. [Google Scholar] [CrossRef] [PubMed]
- Steigerwald, M.; Moll, H. Leishmania Major Modulates Chemokine and Chemokine Receptor Expression by Dendritic Cells and Affects Their Migratory Capacity. Infect. Immun. 2005, 73, 2564–2567. [Google Scholar] [CrossRef] [PubMed]
- Hermida, M.D.R.; Doria, P.G.; Taguchi, A.M.P.; Mengel, J.O.; dos-Santos, W.L.C. Leishmania Amazonensis Infection Impairs Dendritic Cell Migration from the Inflammatory Site to the Draining Lymph Node. BMC Infect. Dis. 2014, 14, 450. [Google Scholar] [CrossRef] [PubMed]
- Marr, A.K.; MacIsaac, J.L.; Jiang, R.; Airo, A.M.; Kobor, M.S.; McMaster, W.R. Leishmania Donovani Infection Causes Distinct Epigenetic DNA Methylation Changes in Host Macrophages. PLoS Pathog. 2014, 10, e1004419. [Google Scholar] [CrossRef]
- Uzonna, J.E.; Wei, G.; Yurkowski, D.; Bretscher, P. Immune Elimination of Leishmania Major in Mice: Implications for Immune Memory, Vaccination, and Reactivation Disease. J. Immunol. 2001, 167, 6967–6974. [Google Scholar] [CrossRef]
- Zaph, C.; Uzonna, J.; Beverley, S.M.; Scott, P. Central Memory T Cells Mediate Long-Term Immunity to Leishmania Major in the Absence of Persistent Parasites. Nat. Med. 2004, 10, 1104. [Google Scholar] [CrossRef]
- Peters, N.C.; Kimblin, N.; Secundino, N.; Kamhawi, S.; Lawyer, P.; Sacks, D.L. Vector Transmission of Leishmania Abrogates Vaccine-Induced Protective Immunity. PLoS Pathog. 2009, 5, e1000484. [Google Scholar] [CrossRef]
- Gomes, R.; Oliveira, F.; Teixeira, C.; Meneses, C.; Gilmore, D.C.; Elnaiem, D.-E.; Kamhawi, S.; Valenzuela, J.G. Immunity to Sand Fly Salivary Protein LJM11 Modulates Host Response to Vector-Transmitted Leishmania Conferring Ulcer-Free Protection. J. Investig. Dermatol. 2012, 132, 2735–2743. [Google Scholar] [CrossRef] [Green Version]
- Glennie, N.D.; Yeramilli, V.A.; Beiting, D.P.; Volk, S.W.; Weaver, C.T.; Scott, P. Skin-Resident Memory CD4+ T Cells Enhance Protection against Leishmania Major Infection. J. Exp. Med. 2015, 212, 1405–1414. [Google Scholar] [CrossRef]
Figure 1.
Central Dogma of Vaccination maintains that the host-protective T cells elicited by optimal immunization protocol protect the host from developing disease upon exposure to the pathogen. However, in the case of Leishmania, all the protocols have failed so far in protecting human vaccines.
Figure 1.
Central Dogma of Vaccination maintains that the host-protective T cells elicited by optimal immunization protocol protect the host from developing disease upon exposure to the pathogen. However, in the case of Leishmania, all the protocols have failed so far in protecting human vaccines.

Figure 2.
(A) Cytokines secreted by macrophages and their effects on immune system; (B) M1 and M2 type macrophages polarization in Leishmania infection and its implication on disease pathogenesis.
Figure 2.
(A) Cytokines secreted by macrophages and their effects on immune system; (B) M1 and M2 type macrophages polarization in Leishmania infection and its implication on disease pathogenesis.

Figure 3.
Leishmania-dependent factors affecting antigen priming leading to inefficacy of vaccination. Upon entry into the macrophages, (A) Leishmanial lipophosphoglycan (LPG) inhibits fusion of phagosome with lysosome via inhibition of SytV recruitment. (B) It also inhibits STAT-1, which is responsible for maintenance of conductivity of Cl- ion channels for phagosomal acidification following phagolysosome formation. (C) Amastigote-expressing proteins, which have high turnover during infection, can trigger intracellular competition among the peptides and influence determinant selection. (D) Cytoplasmic and secretory proteins of parasites have better chances of being processed and presented, inducing a bias towards the antigens to be predominantly presented to T cells. (E) TLR4-induced inhibition of Cathepsin D activity may reduce protein processing ability. (F) MHC-II and H2-DM molecules are taken up by amastigotes inside phagolysosome and degraded. (G) Infection-induced loading of incognate peptides instead of Leishmanial peptides can affect antigenic loading on MHC molecule. (H,I) Leishmanial antigens that may have substitutions in anchor residues or protease cleavage sites may either bind weakly to MHC-II, become resistant to action of proteases, or may function as altered peptide ligands, if selected for presentation to T cells.
Figure 3.
Leishmania-dependent factors affecting antigen priming leading to inefficacy of vaccination. Upon entry into the macrophages, (A) Leishmanial lipophosphoglycan (LPG) inhibits fusion of phagosome with lysosome via inhibition of SytV recruitment. (B) It also inhibits STAT-1, which is responsible for maintenance of conductivity of Cl- ion channels for phagosomal acidification following phagolysosome formation. (C) Amastigote-expressing proteins, which have high turnover during infection, can trigger intracellular competition among the peptides and influence determinant selection. (D) Cytoplasmic and secretory proteins of parasites have better chances of being processed and presented, inducing a bias towards the antigens to be predominantly presented to T cells. (E) TLR4-induced inhibition of Cathepsin D activity may reduce protein processing ability. (F) MHC-II and H2-DM molecules are taken up by amastigotes inside phagolysosome and degraded. (G) Infection-induced loading of incognate peptides instead of Leishmanial peptides can affect antigenic loading on MHC molecule. (H,I) Leishmanial antigens that may have substitutions in anchor residues or protease cleavage sites may either bind weakly to MHC-II, become resistant to action of proteases, or may function as altered peptide ligands, if selected for presentation to T cells.

Figure 4.
Represents formation of immune synapse (IS) between Leishmania infected APC and T cells and selective modulation of IS components by Leishmania. (A) On recognizing an APC with MHC bound cognate antigen, T cells form a contact zone called immune synapse (IS). IS comprises of the core supramolecular activation cluster –cSMAC, the peripheral contact zone-pSMAC, and the distal contact zone, each of which is characterized by the presence of unique cluster of differentiation (CD) molecules, and signaling intermediates like Zap70, each having distinct physiological roles in IS. (B) T cell activation occurs via reception of three signals: (1) Through the triad of TCR-MHC-Peptide-CD4, (2) via costimulatory molecules like CD28, and (3) by the action of cytokines. (C) Leishmania alters the membrane lipid content of infected APCs and may affect the clustering of signaling molecules. It targets costimulatory (CD28, CD80, CD86) molecules and coinhibitory molecules (PD-1, CTLA-4). By selective activation or deactivation of the pathways associated with these molecules, Leishmania can induce anergy, exhaustion, and apoptosis in target lymphocytes.
Figure 4.
Represents formation of immune synapse (IS) between Leishmania infected APC and T cells and selective modulation of IS components by Leishmania. (A) On recognizing an APC with MHC bound cognate antigen, T cells form a contact zone called immune synapse (IS). IS comprises of the core supramolecular activation cluster –cSMAC, the peripheral contact zone-pSMAC, and the distal contact zone, each of which is characterized by the presence of unique cluster of differentiation (CD) molecules, and signaling intermediates like Zap70, each having distinct physiological roles in IS. (B) T cell activation occurs via reception of three signals: (1) Through the triad of TCR-MHC-Peptide-CD4, (2) via costimulatory molecules like CD28, and (3) by the action of cytokines. (C) Leishmania alters the membrane lipid content of infected APCs and may affect the clustering of signaling molecules. It targets costimulatory (CD28, CD80, CD86) molecules and coinhibitory molecules (PD-1, CTLA-4). By selective activation or deactivation of the pathways associated with these molecules, Leishmania can induce anergy, exhaustion, and apoptosis in target lymphocytes.


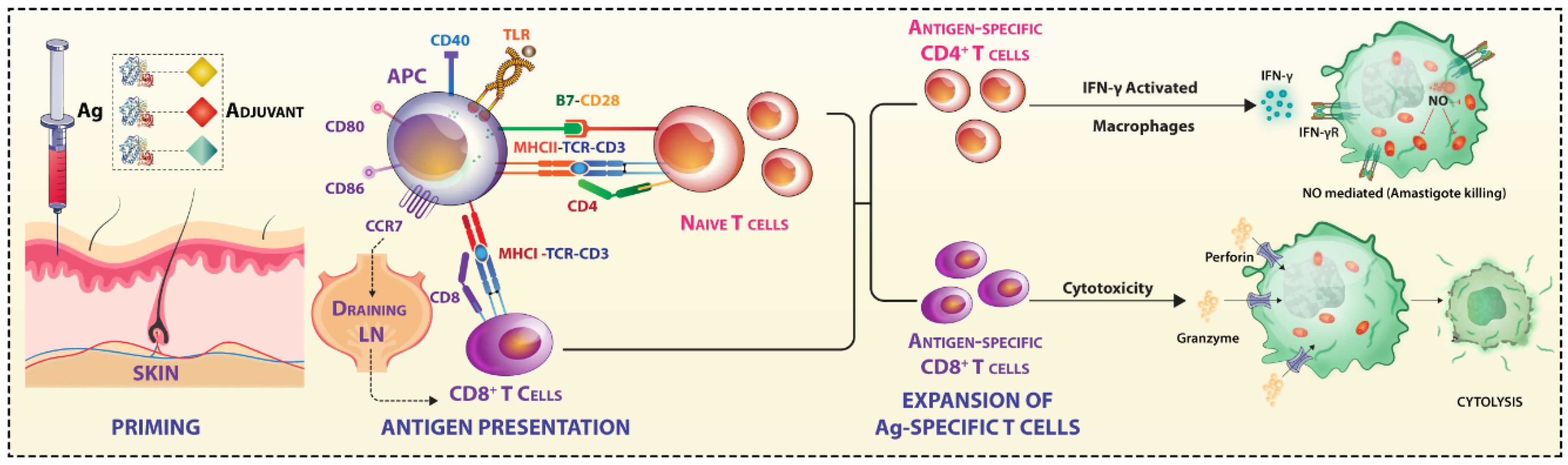{kind=link}
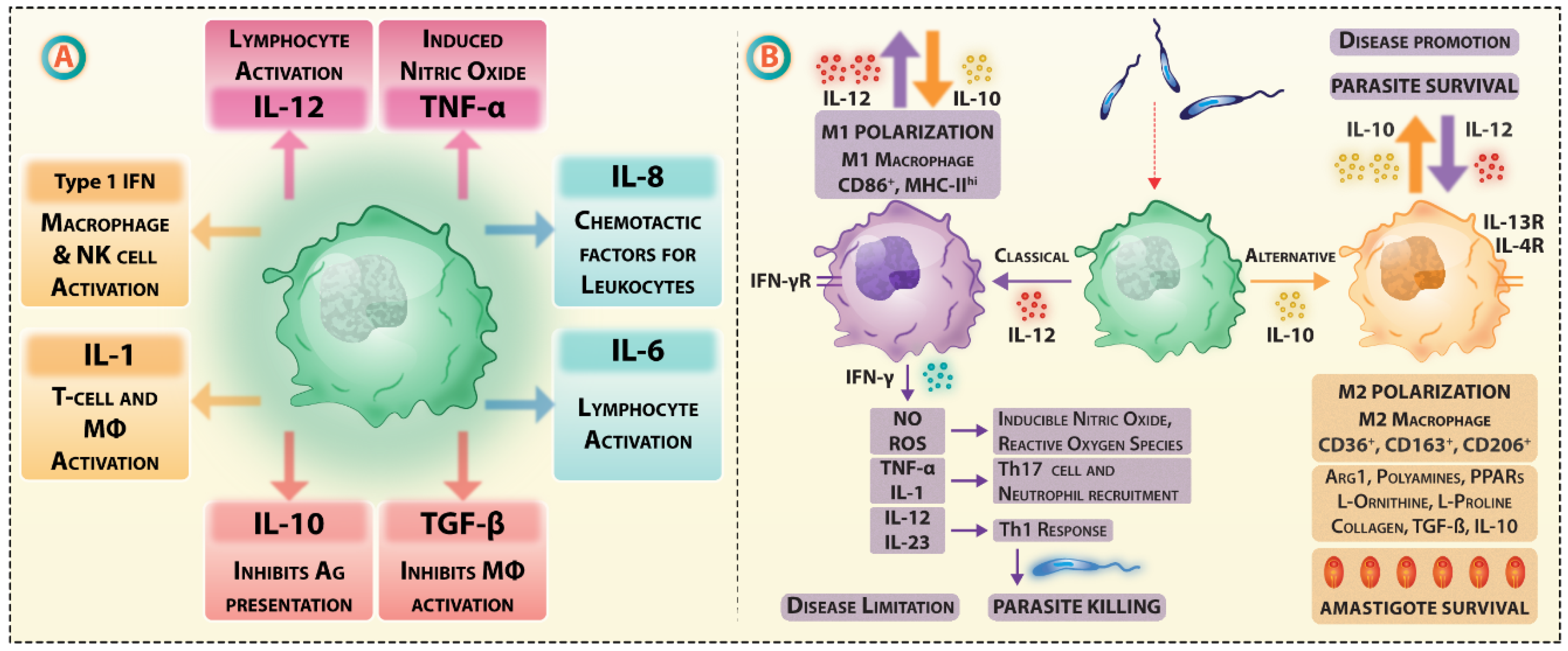{kind=link}
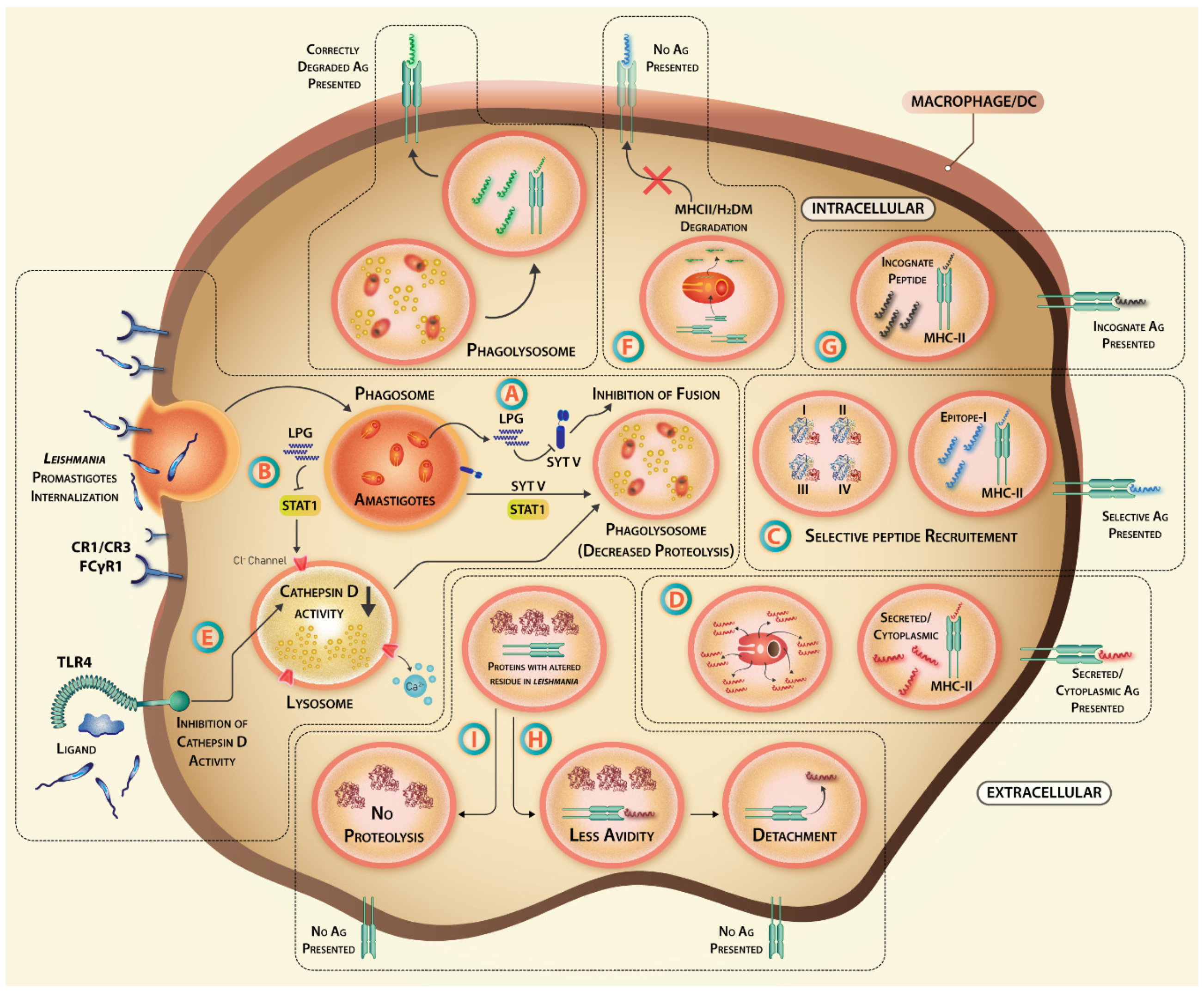{kind=link}
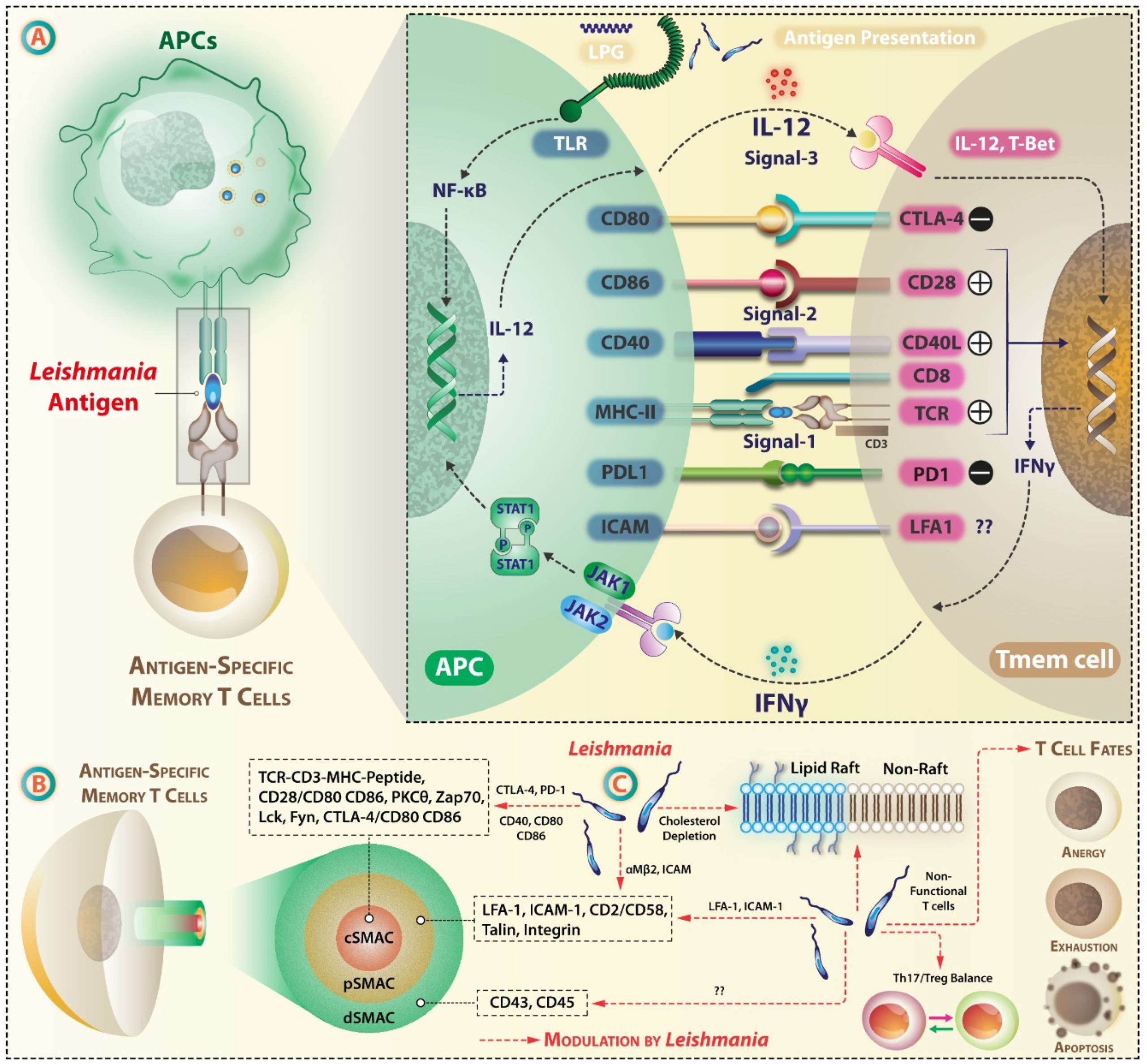{kind=link}
Table 1.
A comprehensive compilation of the vaccine approaches and analyses tested against Leishmania.
Table 1.
A comprehensive compilation of the vaccine approaches and analyses tested against Leishmania.
| VACCINE CANDIDATE | STRAIN (CHALLENGE) | ROI | ADJUVANT | OUTCOME | DRAWBACK(S) | REF |
|---|---|---|---|---|---|---|
| 1ST GENERATION VACCINES | ||||||
| Leishmanization (LZ) | L. major | S.C | -- | Higher success rate in Uzbekistan & Iran | Loss of infectivity, safety & ethical issues | [17] |
| Killed but metabolically active | L. major & L. infantum chagasi | S.C | -- | Treating them with amotosalen and low dose of UV radiation | Shown promise in murine model but failed in human trials | [18] |
| Killed but metabolically active | L. major (Alum ppt. Autoclaved L. major) | -- | BCG | Shown promise in VL and PKDL patients | Chances of relapse | [19] |
| 2ND GENERATION VACCINES | ||||||
| FML (Fucose Mannose Ligand) | L. donovani | S.C, I.P | Saponin, Al(OH)3, QuilA IL-12, BCG | Protective efficacy in BALB/C, Swiss albino, Hamster & dogs | Not shown promise in human | [20,21] |
| Leishmune | L. chagasi | S.C | -- | No parasites were detected in blood, Skin and Lymph node | No trials in humans | [22] |
| Leishmune® | L. donovani L. chagasi | S.C | Saponin | Have transmission blocking property | Partial protection | [23] |
| LAG (L. donovani promastigote membrane antigens) | L. donovani | I.P | Liposome | Shown promise in mice and hamster | Failed in higher animal models | [24] |
| SLA (Soluble Leishmanial Antigen) | L. donovani | I.P, S.C | Liposome, MPL-TDM, non-coding pDNA+ISS | Prophylactic and therapeutic effect in mice model | Free or negatively charged liposome confers partial protection | [25,26,27] |
| gp36 | L. donovani | S.C | Saponin | Protective effect in murine model | Not reached human trials | [28] |
| Ld91, Ld72, Ld51, Ld31 | L. donovani | I.P | Liposome | Reduce parasite burden in visceral organ | Protective efficacy only in mice models | [29] |
| dp72 and gp70-2 | L. donovani | I.P | Corynebacterium parvum | Decrease in parasitemia in visceral organ | Gp70-2 were not promising in mice | [30] |
| SA (Soluble Antigen) | L. donovani | I.M | CpG-ODN | Decrease in parasite load | Ag alone show mixed response in mice models | [31] |
| Ribosomal P0 protein | L. major | S.C | CpG-ODN | Shown promise in C57BL/6 mice | Failure in BALB/C mice | [32] |
| F2 subfraction (97.4-68 kda) | L. donovani | I.D | BCG | Th1 mediated cellular response in cured VL patients and hamsters | Not tested in clinical studies | [33] |
| F2 sub-fraction (89.9-97.1 kDa) | L. donovani | I.D | BCG | 99% parasite inhibition were observed in hamsters | Specific characterization of antigen is needed for higher trials | [34,35] |
| LPG and Phlebotomus duboscqi salivary gland lysates (SGLs) | L. major | S.C | -- | LPG alone provide protection in BALB/C mice | LPG+SGLs failed to provide protection | [36] |
| LPG | Phlebotomus duboscqi sandflies | -- | -- | Block the transmission of Leishmania | -- | [37] |
| gp63 (leishmanolysin) | L. donovani | I.P | Cationic (DSPC) liposomes | Protective effect in BALB/C mice | Mixed responses | [38] |
| Recombinant.gp63 | L. major | -- | -- | Recognition of fusion protein | Fusion protein did not protect the mice | [39] |
| gp63 | L. major | I.V, S.C | -- | BCG expressing gp63 only provide Protection | Limited up to mice only | [40] |
| gp63 | L. major | S.C | CpG ODN + Liposome | Th1 type response | Adjuvant is needed for protection | [41] |
| gp63 | L. donovani | I.M, S.C | CpG ODN | Evoke cellular and humoral response | No trials on higher animal | [42] |
| H2B (Histone protein) | L. major | S.C | CpG | Th1 type response | Only amino-terminal region confer protection | [43] |
| H2B | L. major, L. infantum patients | -- | -- | Whole H2B protein induces Th1 profile in individuals | Not reached clinical trials | [44] |
| rORFF + BT1 | L. donovani | S.C | CFA, ICFA | Detection of high levels of anti-ORFF and anti-BT1 antibodies | Partial protection | [45] |
| rORFF | L. donovani | I.M | CpG ODN | Combination of CpG ODN with rORFF induces reduction in parasite load | Partial protection | [46] |
| ORFF | L. donovani (antimony sensitive & resistant) | I.M | -- | UBQ-ORFF induces cellular & humoral response | Ubiquitin is required for protective effect | [47] |
| Elongation factor 2 (LelF-2) | L. donovani | I.D | -- | Shown promise in Hamster | Not reach clinical trials | [48] |
| rF14 | L. donovani | I.M | MPL | Reduce in parasite burden in spleen and liver | Partial protection | [49] |
| HASPB1 | L. donovani | S.C | IL-12 | Induces DC to produce IL-12 | rHASPB1 immunization induces Th2 response | [50] |
| 78kDa | L. donovani | S.C | MPL-A, Liposome rIL-12, ALD, FCA | Enhanced DTH response, reduction in infection rate of peritoneal macrophages | Antigen alone induces IgG1 isotypes | [51] |
| LCR1 | L. chagasi | I.P, S.C | -- | Protective effect in BALB/C mice | BCG-LCR1 I.P administration did not show protection | [52] |
| Meta1 antigen | L. major | I.M, S.C | IFA | MCP-3/meta 1 fusion partially shift towards Th1 type | Rec. protein Meta1 antigen induces Th2 response | [53] |
| LirCyP1 | L. infantum | I.P | IL-12 | Promote differentiation of helper and memory T cell | Partial protection | [54] |
| Maxadilan (MAX) | L. major | S.C, I.P | CFA IFA | Vaccination against Max elicits Th1 type response | MAX enhance blood flow and inhibit the immune response of the host | [55] |
| LJM19 | L. infantum chagasi | I.D | -- | Induces low parasite load and high IFN-γ/TGF-β ratio | Not reached clinical trials | [56] |
| A2 protein | L. donovani | I.P | Propianibacterium acnes | Production of IFN-ϒ response | Mixed Responses | [57] |
| rLdγGCS | L. donovani | S.C | NIV | Induces humoral response | Mixed Responses | [58] |
| FUSION, HYBRID AND POLYPROTEIN VACCINES | ||||||
| Chimeric Q protein | L. infantum | S.C | -- | Clearance of parasites from majority of the dogs | Studies during natural infection condition are required | [59] |
| Amastigote cysteine proteases | L. major | S.C | -- | Hybrid CPA/B elicits protective response | Partial protection | [60] |
| TAT-antigen fusion protein | L. major | I.D | CpG ODN 1826 | CD8+ T cells rapidly proliferate, activate Leishmania-specific Tc1 cells | Protective effect is observed in C57BL/6 mice | [61] |
| rLeish-111f | L. major | S.C | rIL-12 MPL-SE | Protective efficacy in BALB/C mice | Mixed IgG2a/IgG1 Ab response | [62] |
| Leish-111f | L. infantum | S.C | MPL-SE | 99.6% reductions in parasite loads, completed phase 1 and 2 safety human trial | -- | [63] |
| LEISH-F1 | L. donovani | S.C | MPL-SE | Vaccine is safe, immunogenic in healthy subjects | Further trials are needed | [64] |
| LIVE, ATTENUATED (GENETICALLY) VACCINES | ||||||
| dhfr-ts− null mutant | L. major | I.V, S.C, I.M | -- | Unable to cause disease in susceptible and immune deficient (nu/nu) BALB/C mice | Need to be tested on animal higher models | [65] |
| dhfr-ts− L. major knock out mutant | L. amazonensis | S.C, I.V | -- | Cross- Protection was observed in both BALB/C and C57BL/6 mice | Partial degree of protection | [66] |
| LdCen1(−/−) L. donovani centrin null mutants | L. donovani L. braziliensis | I.M I.C | -- | No parasites were seen in spleen, liver in BALB/C, SCID mice and Hamster, Cross protection against L. braziliensis | Not reach clinical trials | [67] |
| (BT1−/−) Biopterin transporter null mutant | L. donovani | I.V | -- | Th1 mediated response were observed in BALB/C mice | Need to be tested on higher animal models | [68] |
| LiSIR2(+/−) L. infantum SIR2 single knockout | L. infantum | I.P | -- | Reversion of T cell anergy, increased IFN-γ/IL-10 ratio | Mixed response in BALB/C mice | [69] |
| Ldp27(−/−) | L. donovani L. infantum chagasi | I.C | -- | less COX (Cytochrome C oxidase complex) activity and ATP synthesis | Safety efficacy need to be tested on higher animal models | [70] |
| ΔHSP70-II | L. major | I.P, I.V, S.C | -- | Promise protection in BALB/C model, consider safe in SCID mice and hamster | Need to be tested on higher animal models | [71] |
| LiΔHSP70-II null mutant | L. major | I.V, S.C | -- | Promise protection in BALB/C and C57BL/6 mice | Limited only to mice models | [72] |
| Δlpg2 | L. major | S.C | CPG-ODN | Shown promise in C57BL/6 mice | Antigen alone is not sufficient to provide protection | [73] |
| lpg2− | L. major | Foot pad | -- | Suppress IL-4 and IL-10 responses | Very low IFN-ϒ response were observed | [74] |
| Leishmania tarentolae (Non-pathogenic) | L. donovani | I.P | -- | Activates DC, Induces IFN-ϒ, Th1 phenotype in BALB/C mice | Further testing is needed | [75] |
| A2-recombinant L. tarentolae | L. infantum | I.P, I.V | -- | Th1 response in BALB/C mice | I.V route elicits Th2 response | [76] |
| A2-expressing Lactococcus lactis | L. donovani | S.C | -- | Antigen-specific humoral response was observed | Only in BALB/C mice | [77] |
| 3RD GENERATION VACCINES | ||||||
| LACK | L. major | S.C | +rIL-12 -rIL-12 | Protective response in BALB/C mice | Need to be tested on higher animal models | [78] |
| pCIneo-LACK | L. chagasi | I.N Intranasal | -- | Reduce parasite burden in liver and spleen | Limited up to BALB/C mice | [79] |
| LACK-DNA | L. chagasi | I.N | -- | Cellular and humoral response were observed | Not reached clinical trials | [80] |
| p36 (LACK) DNA | L. donovani | I.D, S.C | IL-12 | Robust Th1 response, highly immunogenic | No protection | [81] |
| gp63 and Hsp70 | L. donovani | S.C | -- | Significantly reduce the parasite burden | Need to be tested on higher animal models | [82] |
| A2+E6 | L. donovani | I.M | -- | Induce cellular and humoral response, inhibit cellular p53 response | Limited to mice model | [83] |
| A2 (Expressed in adenovirus) | L. chagasi | S.C | -- | Reduced parasitism in spleen and liver, production of high level of IFN-ϒ | Low antibody response | [84] |
| ORFF | L. donovani | I.M | -- | Induces both cellular and humoral response | Limited to BALB/C mice | [85] |
| rORFF | L. donovani | I.M | IL-12 | Higher levels of IFN-γ were observed | Need to be tested on higher animal models | [86] |
| KMP-11 | L. donovani | I.M | -- | Reversal of T-cell anergy with functional IL-2 generation | Mixed response | [87] |
| KMP-11 | L. major | I.M, S.C | IL-12 | Robust IFN-ϒ production, polarized Th1 response | Require adjuvant for complete protection | [88] |
| ϒ GCS | L. donovani | I.M | -- | Elevated humoral and cell mediated response | Limited to mice model only | [89] |
| H2A, H2B, H3, H4, LACK | L. donovani | I.M, I.D | IL-12 GM-CSF | Induce DTH response, reduction in parasite burden | Unable to induce parasite-specific antibody response | [90] |
| N-terminal domain of proteophosphoglycan (PPG) | L. donovani | I.M | -- | Induce IFN-ϒ, TNF-α, IL-12 and downregulate TGF-β, IL-4, IL-10 | Need further trials ahead of hamster model | [91] |
| Single antigen Gp63, polytope and polytope HSP70 | L. donovani | I.M, I.P | -- | Strong Th1 response | Limited to mice only | [92] |
| NH36 | L. chagasi L. mexicana | I.M, S.C | -- | Reduction in parasite load, Th1 mediated response, cross-protection | No differences were observed after infection with L. mexicana | [93] |
| NH36 | L. chagasi | I.M, I.P | -- | Reduction in parasite burden in liver | Clinical trials not done | [94] |
| KMP11, TSA, elongation factor P74, CPA and CPB, HASPB, A2 LEISHDNAVAX | L. donovani | I.D | -- | Induce CD4+ and CD8+ T cell responses in genetically diverse human populations | Require clinical trials | [95] |
| KMPII, TRYP, LACK and GP63 | L. infantum | I.D | -- | Vaccine was safe and well tolerated for dogs | No protection in dogs | [96] |
| papLe22 | L. infantum | I.M | -- | Downregulate Th2 type response in hamsters | Low antibody response | [97] |
| HbR | L. donovani | I.M | -- | Complete protection in both BALB/C and Hamsters | Clinical trials not done | [98] |
| Tuzin | L. donovani | I.M | -- | higher levels of IFN-γ and IL-12 production | Limited only to BALB/C mice | [99] |
| HETEROLOGOUS PRIME-BOOST (HPB) VACCINES | ||||||
| ORFF | L. donovani | I.M | Alum | 75%–80% reduction in parasite load, enhanced production of IgG2a and IFN-γ in BALB/C mice | Need to be tested on higher animal models | [100] |
| Cysteine proteinases type I and II | L. infantum | S.C | CpG ODN and Montanide 720 | Antigen-specific immune response was observed | Need to be tested on higher animal model | [101] |
| rLdP1 | L. donovani | I.M | FCA | 75.68% decrease in splenic parasite burden in hamsters, | Challenge by other Leishmania species needed for evaluation | [102] |
| A2-CPA-CPB (CTE) recombinant L. tarentolae | L. infantum | Foot pad | cationic solid lipid nanoparticle (cSLN) | High NO production, and low parasite burden | Mixed responses | [103] |
| LACK-WR or LACK-MVA | L. infantum | I.D, I.P | -- | High levels of protection in the draining lymph node | Low protection observed in Spleen and Liver | [104] |
| AlphaGalCer + DNAp36+ VVp36 | L. major | I.D, I.P | αGalCer | 20-fold reductions in parasite burdens | Limited to mice only | [105] |
| DNA/MVA TRYP | L. Viannia panamensis | I.D | α-GalCer, LPS, CpG, Pam3CSK4, MALP-2 | TLR1/2 activation, antigen specific CD8 cells | Need to be tested on higher animal model | [106] |
| pORT-LACK/MVA-LACK | L. infantum | S.C | -- | increase in expression of Th1 type cytokines in PBMC and target organs | Limited to canine model only | [107] |
| LdPxn-1 (L. donovani Peroxidoxin-1) | L. major | I.M S.C | m-GMCSF | Induction of multipotent CD4+ cells | Mixed responses | [108] |
| DENDRITIC CELL(DC)-BASED VACCINES | ||||||
| DCs pulsed with peptide L1 of gp63 (SPL) & pulsing with peptide L2 of gp63 | L. major | I.V | -- | Reduce lesion and parasite load, Th1 phenotype in BALB/C mice with L1 peptide | Partial protection with L1 peptide, whereas L2 peptide switch towards Th2 profile | [109] |
| Plasmacytoid DCs | L. major | I.V | CpG-ODN | Induce T cell-mediated protection | IFN-α production could not be detected | [110] |
| SLDA-pulsed DCs | L. donovani | I.V | IL-12 | Live parasites were not detected in the liver of mice, 1-3 log lower parasite burdens | Granuloma formation were found in SLDA-pulsed, non-transduced DCs BALB/C mice | [111] |
| BM-DCs pulsed with KMP-11 (12–31aa) peptide | L. infantum | I.V | CpG-ODN | Induced the differentiation of peptide-specific Th17 cells | Mixed responses | [112] |
| SLDA-pulsed syngeneic BM-DCs | L. donovani | I.V, I.M | -- | Complete clearance of parasites from spleen and liver | Need to be tested on higher animal models | [113] |
| Hybrid cell vaccine (HCV) | L. donovani | I.V | -- | Strong antigen-specific CTL generation | Expression of IL-4 and IL-13 get elevated at both transcription and translational levels | [114] |
ABBREVIATIONS: ALD (Autoclaved Leishmania antigen), BCG (Bacille Calmette Guerin), CFA (Complete Freund’s Adjuvant), CpG-ODN (CpG- oligodeoxynucleotides), DSPC (Distearoyl phosphatidylcholine), FCA (Freund’s adjuvant), I.D (Intradermal), I.M (Intramuscular), I.N (Intranasal), I.P (Intraperitoneal), I.V (Intravenous), ICFA (Incomplete Freund’s adjuvant), IFA (Incomplete Freund’s Adjuvant), ISS (Immunostimulatory sequences), LPS (Lipopolysaccharide), m-GMCSF (murine granulocyte-macrophage colony-stimulating factor), MPL-SE (Monophosphoryl lipid A plus squalene), MPL-TDM (Monophosphoryl lipid-trehalose dicorynomycolate), NIV (non-ionic surfactant vesicle), S.C (Subcutaneous).
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zutshi, S.; Kumar, S.; Chauhan, P.; Bansode, Y.; Nair, A.; Roy, S.; Sarkar, A.; Saha, B. Anti-Leishmanial Vaccines: Assumptions, Approaches, and Annulments. Vaccines 2019, 7, 156. https://doi.org/10.3390/vaccines7040156
AMA Style
Zutshi S, Kumar S, Chauhan P, Bansode Y, Nair A, Roy S, Sarkar A, Saha B. Anti-Leishmanial Vaccines: Assumptions, Approaches, and Annulments. Vaccines. 2019; 7(4):156. https://doi.org/10.3390/vaccines7040156
Chicago/Turabian StyleZutshi, Shubhranshu, Sunil Kumar, Prashant Chauhan, Yashwant Bansode, Arathi Nair, Somenath Roy, Arup Sarkar, and Bhaskar Saha. 2019. "Anti-Leishmanial Vaccines: Assumptions, Approaches, and Annulments" Vaccines 7, no. 4: 156. https://doi.org/10.3390/vaccines7040156
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.