
Harnessing NK Cells to Control Metastasis

1
Biotherapy Institute of Japan Inc., 2-4-8 Edagawa, Koto-ku, Tokyo 135-0051, Japan
2
N2 Clinic Yotsuya, 5F 2-6 Samon-cho, Shinjuku-ku, Tokyo 160-0017, Japan
*
Author to whom correspondence should be addressed.
Vaccines 2022, 10(12), 2018; https://doi.org/10.3390/vaccines10122018
Submission received: 2 November 2022
/
Revised: 21 November 2022
/
Accepted: 23 November 2022
/
Published: 25 November 2022
Abstract
:In recent years, tumor immunotherapy has produced remarkable results in tumor treatment. Nevertheless, its effects are severely limited in patients with low or absent pre-existing T cell immunity. Accordingly, metastasis remains the major cause of tumor-associated death. On the other hand, natural killer (NK) cells have the unique ability to recognize and rapidly act against tumor cells and surveil tumor cell dissemination. The role of NK cells in metastasis prevention is undisputable as an increase in the number of these cells mostly leads to a favorable prognosis. Hence, it is reasonable to consider that successful metastasis involves evasion of NK-cell-mediated immunosurveillance. Therefore, harnessing NK cells to control metastasis is promising. Circulating tumor cells (CTCs) are the seeds for distant metastasis, and the number of CTCs detected in the blood of patients with tumor is associated with a worse prognosis, whereas NK cells can eliminate highly motile CTCs especially in the blood. Here, we review the role of NK cells during metastasis, particularly the specific interactions of NK cells with CTCs, which may provide essential clues on how to harness the power of NK cells against tumor metastasis. As a result, a new way to prevent or treat metastatic tumor may be developed.
1. Introduction
Although tumor treatment has improved, most metastatic tumors remain incurable [1]. Metastasis is the major cause of tumor-associated death [2]. Hence, concerted efforts have been made to develop therapies that can control metastasis in patients with tumor [3]. Tumors are complex ecosystems composed of neoplastic cells, extracellular matrix (ECM), and “accessory” non-neoplastic cells, which include resident mesenchymal cells, endothelial cells, and immune cells. Accessory cells crosstalk with tumor cells, which fuels and shapes tumor development [4]. Immune cells are essential players in tumor, impacting tumor fate in different stages and therapeutic interventions [4,5]. Over the past decades, immunotherapy has revolutionized tumor treatment [6]. The emergence of immune checkpoint inhibitors (ICIs) targeting programmed cell death protein 1 (PD-1), cytotoxic T lymphocyte-associated protein (CTLA-4), and programmed cell death ligand 1 (PD-L1) has rendered many aggressive tumors treatable and even curable [6]. However, ICIs are successfully used in only a fraction of patients. In particular, their effects are severely limited in patients with low or absent pre-existing T cell immunity [7]. Thus, the initiation of a de novo tumor-specific immune response is required, a process that is dependent on the actions of innate immune cells, among which natural killer (NK) cells play essential roles in tumor immunosurveillance and antitumor immunity owing to their unique ability to identify and kill tumor cells [8]. Indeed, a higher infiltration of NK cells was found to be associated with the better response to anti-PD-1 therapy [9]. NK cells might play an important role in the tumor subsets that exhibit loss of neoantigen presentation due to the downregulation of major histocompatibility complex class I (MHC-I) molecules [10]. Currently, tumor immunotherapy based on NK cells is a very hot topic in oncology and generates considerable interest from the scientific community and pharmaceutical industry. Consequently, there are many studies in which the therapeutic value of NK cells is being evaluated [11,12]. However, the efficacy of NK cell therapy against solid tumor is hampered by inadequate tumor infiltration and immunosuppressive tumor microenvironment. Since NK cells play a more important role in the elimination of metastatic tumor than primary solid tumors [13], we will focus on the interactions of NK cells with metastatic tumor cells, the prominent role of NK cells in the control of metastasis, and the evasion of metastatic tumor cells from NK-cell-mediated immunosurveillance. Understanding tumor cell resistance to NK cells, particularly the specific interactions of circulating tumor cells (CTCs) with NK cells may provide essential clues on how to harness NK cell power to maximize their antitumor potential and may pave the way to the development of novel therapeutic strategies for metastatic tumor.
2. Tumor Metastasis
Metastasis, which is the gravest stage of tumor, occurs when tumor cells acquire invasive features [14] and the ability to evade immunosurveillance [15]. For the successful outgrowth of tumor cells to distant metastatic sites, several critical steps and obstacles need to be overcome. This multistep process, known as the metastatic cascade, involves the detachment of tumor cells from their neighbors and local invasion of surrounding tissues to enter the circulatory system as CTCs in the form of either single (monoclonal) or multiple (polyclonal) tumor cells, until they lodge at secondary sites and enter into a pre-existing or neo-formed vasculature and remain there as disseminated tumor cells (DTCs) or micrometastatic sites (Figure 1). DTCs remain dormant until they resume proliferation and establish detectable metastatic lesions, giving rise to overt macrometastasis and organ colonization [16]. During this process, mesenchymal-like phenotype tumor cells have highly mobile and invasive properties, which occur during the so-called epithelial-to-mesenchymal transition (EMT) [17]. Activation of the EMT program impinges on the immunomodulatory properties and immunogenicity of tumor cells [18,19,20]. CTCs must perform well in multiple events for successful metastasis, especially for successful in survival in the circulation [21] (Figure 1). Most of the CTCs are shed from the primary tumor site, which has become an immunosuppressive microenvironment that protects them from immune attacks, and die during their transport in blood vessels because of hemodynamic shear force, oxidative stress, and susceptibility to immune effector cells [22,23] (Figure 1). Ultimately, only a very small fraction of CTCs survive and become seeds for metastasis. Among tumor-extrinsic factors for metastasis, antitumor immunity, which is a major hindrance to metastatic colonization of CTCs and DTCs [22,23] (Figure 1). Polyclonal CTCs, which have higher metastatic potential [24,25] because they are associated with decreased expression levels of NK-cell-activating ligands that exhibit higher resistance to killing by NK cells [26]. Conventionally, metastasis has been considered to occur in later stages of tumor progression. However, accumulating evidence has also shown metastatic tumor cell dissemination during early tumor formation [27]. The metastatic cascade and immunosurveillance involving NK cells in metastasis are illustrated in Figure 1.
3. NK Cells Are Highly Involved in for the Immunosurveillance of Metastasis
NK cells are effector cells that constitute a key part of the innate immune system and represent up to 5–20% of circulating lymphocyte [28,29]. NK cells show an absence of CD3 molecule but highly express CD56 or CD16, are predominantly defined as CD3−CD56+. There are two major types of NK cell: CD56dimCD16bright/+ (CD56dim) and CD56brightCD16dim/− (CD56bright) NK cells. The CD56dim NK cells are the major subset in peripheral blood, have high cytotoxicity, whereas CD56bright NK cells are predominantly in secondary lymphoid organs as cytokine producing NK cells [28,29]. NK cells have the unique ability to differentiate between normal and transformed cells, and they can recognize and rapidly act against malignant cells without prior sensitization [28,29]. They possess various activating and inhibitory receptors, and the net functional outcome is a complex integration of signals among these activating and inhibitory receptors [28,29]. These inhibitory receptors, such as the inhibitory isoforms of killer-cell immunoglobulin receptors (KIRs), and CD94/NKG2A heterodimers recognize various forms of MHC-I molecules. Thus, the decrease in the expression levels or the absence of MHC-I molecules on tumor cells reduces the strength of inhibitory signals delivered to NK cells, thus promoting NK cell activation. NK cell activation also results from the engagement of activating receptors, such as the activating isoforms of KIRs, the signaling lymphocyte-activating molecule-related receptors NKG2D, DNAX accessory molecule-1 (DNAM-1), and the natural cytotoxicity receptors NKp30, NKp44, and NKp46, which recognize stress-inducible ligands on tumor cells that are scarcely expressed in healthy cells. Therefore, NK cells have a well-documented antitumor effect, including antibody-dependent cellular cytotoxicity (ADCC) through specific IgG antibodies to target antigens [28,29,30,31]. Indeed, an epidemiological study has shown that a reduced NK cell function was associated with an increased tumor incidence in humans [32].
When NK cells recognize aberrant cells, such as tumor cells, they are activated and transport specific lytic granules such as perforins and granzymes toward immunological synapses to induce apoptosis of target cells [33]. Perforins are cytolytic proteins that are inserted into the plasma membrane of a target cell and induce osmotic lysis in a Ca2+-dependent manner [34]. Granzymes are serine proteases that activate caspase signaling, leading to the apoptosis of the target cell [35]. Perforins are critical in controlling tumor metastasis [36,37]. NK cells form multiple contacts with target cells and can sequentially kill several tumor cells in a time-dependent manner [38]. Interestingly, upon a single encounter, an NK cell releases only one tenth of its cytotoxic lytic granules, but it has been determined that even a single granule is sufficient to induce tumor cell death [34]. NK cells have been shown to shift from inducing fast granzyme B-mediated cell death to slow death mediated by receptor ligands (Fas ligand and tumor necrosis factor (TNF)-related apoptosis-inducing ligand (TRAIL)) in later stages and can serially kill up to more than 30 tumor cells [39].
Although the role of NK cells in the control of primary tumors remains controversial, their robust antimetastatic effect in vivo has been demonstrated in various experimental models [40,41,42,43,44]. Tumor stem cells (TSCs) [45] and EMT trans-differentiated cells [17] are considered responsible for tumor metastasis. Meanwhile, NK cells can target TSCs and EMT cells [30,46,47,48]. Human and mouse breast TSCs induce NK cell activation and expansion in vivo, which correlate with the inhibition of TSCs metastatic spread [46]. EMT induces E-cadherin- and cell adhesion molecule 1 (CADM1)-mediated NK cell susceptibility. Thus, a higher CADM1 expression level correlates with improved survival and reduced metastasis in patients with lung and breast adenocarcinomas [48]. In human tumors, there is accumulating evidence of correlations among the number of infiltrating NK cells, metastasis, and the prognosis of various tumors, such as esophageal tumor (ET) [49], gastric carcinomas (GCs) [50], gastrointestinal sarcoma tumors [51,52], renal cell carcinomas (RCCs) [53], colorectal carcinomas (CRCs) [54], and prostate tumor (PT-1) [55]. In addition, the presence of highly effective NK cells indicates a good prognosis in metastatic PT-1 [56]. Overall, an increased number of NK cells is highly beneficial for the survival of tumor patients [8]. Importantly, however, CTCs are considered to be indicators of an increased risk of metastasis and poorer outcomes in tumor patients. In particular, NK cells are present in blood and lymph nodes and may participate in the immunosurveillance of CTCs [22,44]. In an experimental model, NK cells destroy CTCs before the extravasation and thus suppressing tumor metastasis [42,44,57]. Thus, a short period of an elevated number of NK cells is correlated with the reduction in the number of CTCs [58,59]. Indeed, NK cells were found to eliminate disseminated tumor cells from the lungs within 24 h of arrival, but not thereafter [44]. Moreover, the cytotoxicity of NK cells from CTC-positive patients is lower than that from CTC-negative patients with metastatic breast tumor (BT), CRC, and PT-1, showing a close correlation between peripheral blood CTC number and NK cell antitumor activity [60,61]. Hence, a decreased circulating NK cell activity is associated with increased risk of metastasis in patients with pharyngeal carcinoma [62]. In addition, a decreased NK cell activity is a parameter for predicting distant metastasis following curative surgery for CRC [63,64,65]. Moreover, surgical stress induces NK cell dysfunction in animal models [66,67,68], and human tumors [69,70,71,72,73], leading to metastasis [66,67,68]. Thus, enhancing NK cell function by inhibiting the induced NK cell dysfunction can prevent postoperative metastasis [66,67,68,74]. Taken together, these results indicate that NK cells eliminate CTCs and play a prominent role in the control of metastasis [13].
4. Evasion of NK-Cell-Mediated Immunosurveillance in Metastasis
The immune system plays a major role in every step of tumor progression [4,5]. The escape of tumor cells from immunosurveillance is a prerequisite for metastasis [15,23]. Since NK cells are highly involved in for immunosurveillance in metastasis [13], it is therefore plausible that for successful metastasis, tumor cells have to evade NK-cell-mediated immunosurveillance. Indeed, several mechanisms of NK-cell-mediated tumor cell escape have been described [23,26,75]. These mechanisms involve tumor cell intrinsic features and tumor-driven extrinsic microenvironmental factors (Figure 2). Herein, we explore these escape mechanisms of NK cells in metastasis. The intrinsic features endow tumor cells with distinct metastatic potential through epigenetic and genetic alterations, including the downregulation of NK cell activating receptor ligands due to, for example, the reduction in the expression levels of MHC-I polypeptide-related sequence A (MICA)/MICB and UL16-binding protein 2 (ULBP2) caused by the aberrant expression of oncogenic microRNA, miR-20a, and miR-34a/c in tumor cells [76,77]. MICA/MICB and ULBP2 are the ligands of the NK cell activating receptor NKG2D. Along similar lines, tumor cells expressing miR-296-3p inhibit the expression of intercellular adhesion molecule 1 (ICAM-1), which is the ligand of β2 integrin lymphocyte function-associated antigen 1 (LFA-1), providing activating signals to NK cells [58]. In addition, NK cell inhibitory receptor ligands, such as human leukocyte antigen-G (HLA-G) and PD-L1, are up-regulated. HLA-G is the non-classic MHC-I gene and the ligand for the inhibitory receptor of KIRs, which are highly expressed in numerous tumor cells [78]. HLA-G exerts its function through its binding to immunoglobulin-like transcript 2 (ILT2) and KIRs on NK cells to protect tumor cells from NK cell cytotoxicity [79]. Thus, HLA-G expression is associated with tumor metastasis [80]. Moreover, plasma soluble HLA-G (sHLA-G) in extracellular vesicles was associated with the presence of CTCs and disease progression, indicating that CTCs exploit sHLA-G in the blood to evade NK-cell-mediated immunosurveillance [81]. Moreover, NK cell functions are affected by the inhibitory ligand PD-L1 expressed on tumor cells, which provide inhibitory signals to NK cells through the PD-1/PD-L1 axis, and reduce NK cell cytotoxicity [82]. In addition, cell death receptor FAS expression on tumor cells is down-regulated, leading to escape from NK-cell-related apoptosis [83].
In contrast, the tumor cell-driven extrinsic microenvironmental factors suppress NK cell immunosurveillance through modulation of the recruitment, cell surface molecules, or the release of immunosuppressive soluble factors such as TGF-β1 [84,85,86] (Figure 2). NK cells in lung, breast, or prostate tumor tissues displaying altered receptor expression with impaired cytotoxicity compared with NK cells in control tissues [84,85,86]. These alterations have also been observed in peripheral NK cells and associated with disease progression [85,86]. The expression levels of activating receptors on NK cells were decreased: for example, such as CD16 in BT, multiple myeloma (MM), and CRC [86,87,88]; NKp46 in pancreatic tumor (PT-2), GC, CRC, acute myeloid leukemia (AML), and cervical tumor (CC-1) [89,90,91]; NKp44 in BT and AML [92,93]; NKp30 in BT, hepatocellular carcinoma (HCC), PT-2, GC, CRC, chronic lymphocytic leukemia, and CC-1 [86,89,90,91,94]; CD94/NKG2C in AML [91]; NKG2D in BT, lung tumor (LT), CRC, colon carcinoma (CC-2), PT-2, GC, and CC-1 [86,89,90,95,96]; CD244 in AML [91]; DNAM-1 in BT, CC-2, and AML [86,91]. On the other hand, the expression levels of inhibitory receptors on NK cells were increased: for example, KIR3DL1 in PT-2, GC, and CRC [84]; KIR2DL2/L3 in melanoma [97]; NKG2A in BT, LT, and CRC [86,98,99]; PD-1 in ET, HCC, CRC, GC, biliary tumor, and LT [100,101]; TIM-3 in LT, melanomas, and GC [102,103,104]. TGF-β1 in the tumor environment is involved in the phenotype alteration and functional impairment of NK cells [85,86]. In addition, TGF-β1 and other immunosuppressive factors produced by tumor cells have been shown to suppress NK cell effector functions [105] (Figure 2) and recruit immunosuppressive cells to tumor tissue, such as regulatory T cells (Tregs) [106], myeloid-derived suppressor cells (MDSCs) [107], CD11b+Ly6G+ neutrophils [108], and indoleamine 2, 3-dioxygenase 1 (IDO1)-expressing dendritic cells (DCs) [109]. They also contribute to the phenotype alteration and functional impairment of NK cells to prepare for distant microenvironments for metastatic niches (Figure 1). Moreover, neutrophils can cooperate with metastatic tumor cells to enhance both the dissemination of tumor cells out of the primary tumor and their subsequent intravasation into the lung vasculature through the secretion of G-CSF to attract neutrophils, which suppress NK cell activity through ROS signaling and shield intraluminal metastatic tumor cells from being cleared by NK cells (Figure 2), and ultimately enhance metastatic outgrowth [109,110]. Furthermore, tumor cells may reprogram NK cells through DNA methyltransferases (Dnmt1, Dnmt3a, and Dnmt3b), thereby increasing the expression level of inhibitory receptors TIGIT and KLRG1 on NK cells and promoting metastatic outgrowth [111].
Platelets are small, anucleated cell fragments that have a characteristic discoid shape and diameters ranging from 1 to 3 μm [112]. The main roles of platelets are the maintenance of the hemostasis of the vascular system and the promotion of wound healing at sites of vascular injury [113]. However, having a high platelet count was identified as a risk factor for adverse outcomes in numerous different tumors [114]. Platelets may affect many components of antitumor immunity [115]. An increased coagulability of platelets facilitates vascular evasion and the establishment of metastasis [114,116], whereas the abrogation of platelet function results in reduced metastasis, depending on the presence of NK cells [117]. Platelets help tumor cells evade NK cell immunosurveillance in the blood stream, supporting CTCs to establish metastasis [114,116] (Figure 1). Platelets promote metastasis by coating CTCs traveling through the blood, thereby physically shielding them from shear force. Moreover, a ‘pseudonormal’ phenotype is conferred to CTCs by the transfer of platelet-derived MHC-I molecules, which causes CTCs to mimic host cells and protects them from recognition by NK cells. Moreover, platelet-induced NK cell inhibitory ligands, such as TNF family members of glucocorticoid-induced TNF receptor-related (GITR) ligands, and NKG2D ligands shedding, particularly MICA and MICB on CTCs, protect CTCs from NK cell immunosurveillance [118,119,120,121] (Figure 2). In addition, platelets may play a prometastatic role by promoting thrombin activation resulting in shedding the DNAM-1 ligand Necl5 from the metastatic tumor cells to evade NK cell surveillance [44]. Moreover, platelets release soluble factors upon activation when adhesion to CTCs. TGF-β1 is one of the platelet-derived soluble factors that inhibit NK cell function partially mediated by the downregulation of the activating receptor NKG2D on NK cells [122] (Figure 2). TGF-β1 initiates and maintains the EMT phenotype alteration of CTCs [123]. Therefore, platelets affect NK cell function on several different levels, and both soluble and membrane-bound factors are involved in the evasion of CTCs from NK cell immunosurveillance in vivo.
Taken together, these results exemplify the obstacles posed by NK cell immunosurveillance in metastasis (Figure 2). The characterization of diverse mechanisms contributing to the dysfunction of NK cells in different tumors may pave the way for the development of novel therapeutic strategies by harnessing NK cells in the control of metastasis.
5. Harnessing NK Cells in Control of Metastasis
A breakthrough in tumor immunotherapy comes from the clinical successes of ICI and chimeric antigen receptor (CAR) T cell therapies, proving that these treatments have great promise for tumor patients [6,124]. NK-cell-based tumor therapy currently constitutes a major area of immunotherapy innovation and has grown exponentially [11,12,125]. To cure tumors, unleashing the full antitumor potential of NK cells is an attractive option (Figure 3) (Table 1), since NK cells play a more important role in the elimination of metastasis [13], which is the major cause of tumor-related death [2]. Most of these strategies are very often reviewed in detail [11,12,13], and the advantage/disadvantages for targeting metastasis are summarized by Lorenzo-Herrero et al. [126].
5.1. Cytokine-Based Therapy
Cytokines endow NK cells with enhanced effector functions in antitumor immunity and enhance their persistence in vivo [127] (Figure 3). IL-2 and IL-15 are key cytokines that upregulate the activity of NK cells [128]. IL-2 was the first cytokine employed in tumor treatment and the first reproducible and effective human tumor immunotherapy approved by the Food and Drug Administration (FDA) [129]. An improved survival of patients with metastatic RCC and melanomas was achieved by high-dose IL-2 treatment [130]. However, it caused life-threatening toxicities, including vascular leak syndrome [129,130]. In addition, IL-2 therapy induces the proliferation and activation of Tregs and therefore immunosuppression [131]. Thus, it has been speculated that the limited efficacy of IL-2 therapies in vivo is at least partially attributable to the inhibitory effect of Tregs. For these reasons, there is a need for alternatives to IL-2 and a rationale for the evaluation of variant forms of recombinant IL-2 in order to gain a higher affinity to NK cells and lower affinity to Tregs [132]. Over the past decades, IL-15 has emerged as a promising substitute for IL-2. IL-2 and IL-15 are closely related homeostatic cytokines, and both require CD132 and CD122 heterodimers for signaling [128]. Clinical trials with recombinant IL-15 or IL-15/IL-15Rα complexes are being conducted in metastatic tumors [133] (Table 1). In the first in-human phase I clinical trial of IL-15 in patients with metastatic malignant melanoma and metastatic RCC, hyperproliferation and an increase in the number of circulating NK cells were observed [133]. Even though a preliminary antitumor evaluation showed no objective responses, two patients showed clearance of lung metastasis [133]. Other cytokines that activate NK cells without stimulating Tregs, such as IL-12, IL-18, and IL-21, are now being studied [127]. However, cytokine-based-NK cell-activating strategies in the treatment of metastasis remain to be established.
5.2. Adoptive NK Cell Therapy
An alternative approach to the systemic activation of NK cells is to directly introduce activated NK cells to a patient, known as adoptive transfer (Figure 3) (Table 1). As reviewed by Myers et al. [11], a wide variety of sources of therapeutic NK cells are currently being tested clinically, including autologous NK cells, allogenic NK cells, umbilical cord blood (UCB) NK cells, NK cell lines, cytokine-induced memory-like (CIML) NK cells, and CAR NK cells. The adoptive transfer of a patients’ own NK cells (autologous transfer) enables ex vivo stimulation and expansion prior to re-administration as a therapeutic modality. Initial clinical results of adoptive transfer of expanded autologous NK cells in patients with metastatic tumor showed that this treatment strategy is well tolerated, but the clinical response was limited [129,134]. It was considered that the failure of autologous NK cell therapies could be partially attributed to the inability of inhibitory KIRs to recognize self-MHC I on tumor cells. Thus, the adoptive transfer of ‘foreign’ NK cells (allogenic NK cells) for therapy has been examined in patients with various tumors. Allogeneic NK cells derived from healthy donor cells are advantageous because they have higher potential in antitumor activity. A complete remission and disease-free survival have been observed in patients with AML after the adoptive transfer of allogenic KIR-ligand mismatched donor NK cells [135]. The lack of engagement of inhibitory KIR receptors on allogenic NK cells with MHC-I ligands in these patients may be beneficial and contribute to the clinical response [135]. Thus, the transfer of allogeneic NK cells results in a reduction in the CTC number in patients with stage IV non-small cell lung cancer (NSCLC) [136] and recurrent BT [137]. However, the clinical response is limited in metastatic tumor patients in studies of adoptive allogenic NK cell therapy [138,139,140,141,142,143]. To expand the therapeutic use of allogenic NK cells, the use of UCB is considered. NK cells constitute up to 30% of the lymphocytes in UCB, which is a robust source of therapeutic NK cells [144]. The therapeutic efficacy of UCB NK cells is currently being evaluated in clinical trial (Table 1). Clonal NK cell lines, such as NK-92 and KHYG-1, are an alternative source of allogeneic NK cells. The NK-92 cell line has received FDA approval for use in clinical trials and has been extensively tested in clinical trials [145]. However, these cells are aneuploid and therefore genetically unstable, which requires them to be irradiated prior to infusion. NK cells have innate memory, that is, they can remember a prior activation event and consequently respond more robustly when restimulated. NK cell memory has been identified following combined cytokine of IL-12, IL-15, and IL-18 preactivation [146]. The CIML NK cells were shown to have a phenotype distinct from conventional NK cells. They have improved effector functions and induce remission in AML patients [147]. Following the clinical successes achieved with CAR T cell therapies, the use of CAR NK cells represents a promising immunotherapeutic strategy, and CAR NK cells might show greater advantages than CAR T cells, such as the induction of less severe side effects [148]. In this regard, it has recently been shown that CAR NK cells do not cause any serious cytokine storm in patients with lymphoid tumors [149]. In a murine model, CAR NK cell therapy reduced lung metastasis in an RCC by targeting ERbB2/HER2, indicating its potential in the control of disease dissemination [150].
5.3. Agonists of Activating and Inhibitory Receptors
NK cell immunosurveillance may be improved in metastasis by targeting NK-cell-function-related activating and inhibitory receptors (Figure 3) (Table 1). Over the past two decades, monoclonal antibodies (mAbs) have been widely used in tumor treatment, and NK cell activity has been increased by employing tumor-specific antibodies that ligate to CD16 receptors on NK cells to promote ADCC [151]. The CD20-targeting mAb rituximab, the epidermal growth factor receptor (EGFR)-targeting mAb cetuximab, and the erb-b2 receptor tyrosine kinase 2 (ERBB2)-targeting mAb trastuzumb are used in the treatment of solid and hematological tumors [152]. The CD38-targeting mAb daratunumb [153] and the CD139-targeting mAb elotuzumab [154] constitute the treatment arsenal against MM. The clinical response is modulated by the polymorphism in the genes encoding CD16 receptors [155], indicating that ADCC plays a crucial role in the therapeutic activity of these mAbs. Children with neuroblastoma are given allogeneic NK cells following administration of anti-GD2 mAb, which recognizes a surface molecule highly expressed in neuroblastoma cells. Administration of NK cells with ADCC enhanced by anti-GD2 mAb produced a partial or complete response in approximately 40% of patients [156,157]. Trastuzumab was successfully introduced in the treatment of HER2+ BT and GC. In patients with HER2+ BT, even trastuzumab monotherapy has produced a clinical response [158], whereas a clinical study of adoptive infusion of autologous NK cells in addition to trastuzumab in a refractory patient with HER2+ BT showed the reversal of the resistance to trastuzumab [159]. Clinical trials on the combinations of autologous NK cells with mAbs are ongoing, such as the combinations of autologous NK cells with trastuzumab in HER2+ tumors [160] and with cetuximab in recurrent and/or metastatic nasopharyngeal carcinoma [161] (Table 1). The combination of adoptive NK cells and mAbs targeting CTCs via NK-cell-mediated ADCC is a promising therapeutic strategy for the control of metastasis, as this function appears to be poorly affected by the barrier of the tumor microenvironment. Moreover, CD16 can also be engaged with bispecific or trispecific killer-cell engagers (BiKE or TriKE), which bind the CD16 and tumor antigens simultaneously, to trigger NK cell activation through the CD16 receptor, significantly increasing NK cell cytolytic activity and cytokine production against tumor targets [162].
Blocking inhibitory receptors on NK cells is a suitable strategy to increase antitumor activity [11] (Figure 3) (Table 1). The blockade of the anti-KIR antibody lirilumab enhanced NK cell activity through the blockade of NK cell interaction between inhibitory receptors KIR2DL-1, KIR2DL-2, and KIR2DL-3 with HLA-C group 1 and 2 allotypes [163]. A phase I trial in patients with relapsed/refractory MM using lirilumab as a single agent has shown enhanced ex vivo patient-derived NK cell cytotoxicity against MM (Table 1). However, no objective responses were observed [164]. Therefore, the combination of IPH2101 with lenalidomide is under clinical investigation [165] (Table 1). In addition, the humanized anti-NKG2A antibody monalizumab, which blocks NKG2A–HLA-E interaction, enhanced NK cell activity against various tumor cells; its clinical trials are ongoing [166] (Table 1). Monalizumab monotherapy for recurrent metastatic squamous cell carcinoma of the head and neck was much less efficacious with no objective response, and the disease was stable in only 23% of the patients [167]. On the other hand, checkpoint blockade targeting the PD-1/PD-L1 and CTLA-4 inhibitory axis has produced remarkable results in tumor treatment, and the clinical benefits are considered to mainly be based on the reactivation of exhausted T cells [168]. However, the effect of ICIs on NK cells through PD-1/PD-L1 blockade also mediates antitumor immunity [9]. Thus, targeting PD-1 by blocking the PD-1/PD-L1 signaling axis enhances the NK cell immune response against MM [169]. Clinical studies have been conducted to investigate the effect of pembrolizumab (anti-PD-1) combined with autologous [170] or allogeneic [171] NK cells in patients with NSCLC. These studies showed increased NK cell activity in patients receiving NK cells and improved survival compared with patients receiving pembrolizumab alone [170,171]. Additionally, a fully humanized anti-PD-L1 mAb has been shown to block PD-1/PD-L1 interactions and promote NK-cell-mediated ADCC against tumor cells [172]. Furthermore, TIM-3 blockade has been shown to reverse NK cell impairments and increase NK cell antitumor activity in patients with advanced melanoma and lung adenocarcinoma [102,104]. A plethora of anti-TIGIT mAbs are in advanced clinical development for solid tumors, such as domvanalimb, vibostolimb, tiragolumab, and ociperlimab [173] (Table 1). Further investigation, however, is required to elucidate the actual anti-metastatic potential of blocking inhibitory receptors in humans.
TGF-β1 is one of the major suppressive cytokines produced by tumor cells and platelets. It inhibits NK cell effector functions and helps tumor cells evade NK cell immunosurveillance [84,85,86,123] (Figure 2). A pharmacological inhibitor of the TGF-β1 pathway (Figure 3), galunisertib, has been tested in patients with neuroblastoma [174] and HCC [175], and an increased NK cell activity against tumor cells was observed [174].
As platelets can protect tumor cells from NK cell immunosurveillance, targeting the protective interaction of platelets with tumor cells has been suggested to improve the NK cell antitumor activity [114] (Figure 3). The platelet inhibitor ticagrelor specifically inhibits tumor-associated platelets and strongly reduces lung metastasis in a mammary carcinoma mouse model [176]. In addition, silencing tumor-specific tissue factors (TFs) by nanoparticle-mediated delivery of siRNA resulted in reduced platelet adhesion and ultimately the number of lung metastases [177]. Future research will show if targeting the interaction of platelets with tumor cells can improve the efficiency of NK cell immunosurveillance.
6. Conclusions and Future Direction
Tumor immunotherapy by revitalizing immune responses against tumor cells has shifted the paradigm in tumor therapy [6,168]. Nevertheless, the metastatic spread of tumor cells remains the main cause of tumor-related death [2]. NK cells are innate immune cells that can directly and rapidly kill tumor cells without antigen restriction, and they are highly responsible for immunosurveillance in metastasis [13,44]. Hence, tumor cells have to evade NK-cell-mediated immunosurveillance for successful metastasis (Figure 2). Thus, it is promising to harness NK cells for the prevention or treatment of tumor metastasis (Figure 3). As the evading mechanisms involved NK cell immunosurveillance in the metastatic cascade, particularly the specific interactions of CTCs with NK cells in metastasis (Figure 2), an optimal therapeutic window may exist to achieve maximal NK cell antitumor activity. The experimental lung metastasis model showed that NK cells eliminate CTCs from the lung within 24 h of arrival, but not thereafter. Half of NK–tumor cell encounters lead to tumor cell death in the first 4 h after tumor cell arrival [44]. NK cell therapies would be mostly used in the adjuvant setting as adjuvants, such as after surgery [74] and following stem cell transplantation [178] targeting minimal residual disease. Specifically, NK cells may be most effective for targeting CTCs, as their function appears to be poorly affected by the barrier of the tumor microenvironment [179]. Clinical trials are underway to evaluate the efficacy of NK cell immunotherapy by each therapeutic strategy alone or in combination (Figure 3), and further in combination with other strategies, including standard treatments, such as mAbs therapy [160,161] and chemoradiotherapy [180], with encouraging clinical results. Moreover, recent data suggest that molecularly targeted agents [181] and radiotherapy [182] capable of inducing senescence in tumor cells elicit NK-cell-mediated immunosurveillance, facilitating tumor regression, indicating that NK cells are likely to be a key player in future multimodal strategies against tumors. Further studies should be performed in order to fully understand the antitumor and antimetastatic properties of NK cells, particularly when and at which steps of the metastatic cascade NK cells operate, and how many times NK cell immunotherapy is required, which will pave the way to developing novel therapeutic strategies for the prevention or treatment of tumor metastasis.
Author Contributions
Conceptualization, X.D. and H.T.; literature review, X.D.; writing—original draft preparation, X.D.; writing—review and editing, X.D. and H.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing not applicable.
Conflicts of Interest
X.D. and H.T. hold patents for feeder-cell-free NK cell expansion culture technology and are stockholders of Biotherapy Institution of Japan Inc., a biotech startup company developing NK-cell-based immunotherapy and Mesenchymal Stromal Cell (MSC)-based products, such as adipose MSCs, and MSC-derived conditioned medium (CM) concentrates and extracellular vesicle (EV) isolates (StemSup®) for clinical research.
References
- Santucci, C.; Carioli, G.; Bertuccio, P.; Malvezzi, M.; Pastorino, U.; Boffetta, P.; Negri, E.; Bosetti, C.; La Vecchia, C. Progress in cancer mortality, incidence, and survival: A global overview. Eur. J. Cancer Prev. 2020, 29, 367–381. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef]
- Stoletov, K.; Beatty, P.H.; Lewis, J.D. Novel therapeutic targets for cancer metastasis. Expert Rev. Anticancer Ther. 2020, 20, 97–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, H.; Hagerling, C.; Werb, Z. Roles of the immune system in cancer: From tumor initiation to metastatic progression. Genes Dev. 2018, 32, 1267–1284. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, H.; Robles, I.; Werb, Z. Innate and acquired immune surveillance in the postdissemination phase of metastasis. FEBS J. 2018, 285, 654–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van den Bulk, J.; Verdegaal, E.M.; de Miranda, N.F. Cancer immunotherapy: Broadening the scope of targetable tumours. Open Biol. 2018, 8, 180037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teng, M.W.; Ngiow, S.F.; Ribas, A.; Smyth, M.J. Classifying Cancers Based on T-cell Infiltration and PD-L1. Cancer Res. 2015, 75, 2139–2145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bald, T.; Krummel, M.F.; Smyth, M.J.; Barry, K.C. The NK cell-cancer cycle: Advances and new challenges in NK cell-based immunotherapies. Nat. Immunol. 2020, 21, 835–847. [Google Scholar] [CrossRef]
- Hsu, J.; Hodgins, J.J.; Marathe, M.; Nicolai, C.J.; Bourgeois-Daigneault, M.C.; Trevino, T.N.; Azimi, C.S.; Scheer, A.K.; Randolph, H.E.; Thompson, T.W.; et al. Contribution of NK cells to immunotherapy mediated by PD-1/PD-L1 blockade. J. Clin. Investig. 2018, 128, 4654–4668. [Google Scholar] [CrossRef]
- Lee, H.; Quek, C.; Silva, I.; Tasker, A.; Batten, M.; Rizos, H.; Lim, S.Y.; Nur Gide, T.; Shang, P.; Attrill, G.H.; et al. Integrated molecular and immunophenotypic analysis of NK cells in anti-PD-1 treated metastatic melanoma patients. Oncoimmunology 2018, 8, e1537581. [Google Scholar] [CrossRef] [PubMed]
- Myers, J.A.; Miller, J.S. Exploring the NK cell platform for cancer immunotherapy. Nat. Rev. Clin. Oncol. 2021, 18, 85–100. [Google Scholar] [CrossRef] [PubMed]
- Shimasaki, N.; Jain, A.; Campana, D. NK cells for cancer immunotherapy. Nat. Rev. Drug Discov. 2020, 19, 200–218. [Google Scholar] [CrossRef]
- López-Soto, A.; Gonzalez, S.; Smyth, M.J.; Galluzzi, L. Control of Metastasis by NK Cells. Cancer Cell 2017, 32, 135–154. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Ting, D.T.; Stott, S.L.; Wittner, B.S.; Ozsolak, F.; Paul, S.; Ciciliano, J.C.; Smas, M.E.; Winokur, D.; Gilman, A.J.; et al. RNA sequencing of pancreatic circulating tumour cells implicates WNT signalling in metastasis. Nature 2012, 487, 510–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spranger, S.; Bao, R.; Gajewski, T.F. Melanoma-intrinsic β-catenin signalling prevents anti-tumour immunity. Nature 2015, 523, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Massagué, J.; Obenauf, A.C. Metastatic colonization by circulating tumour cells. Nature 2016, 529, 298–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brabletz, T.; Kalluri, R.; Nieto, M.A.; Weinberg, R.A. EMT in cancer. Nat. Rev. Cancer 2018, 18, 128–134. [Google Scholar] [CrossRef]
- Chockley, P.J.; Keshamouni, V.G. Immunological Consequences of Epithelial-Mesenchymal Transition in Tumor Progression. J. Immunol. 2016, 197, 691–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Soto, A.; Zapico, L.H.; Acebes-Huerta, A.; Rodrigo, L.; Gonzalez, S. Regulation of NKG2D signaling during the epithelial-to-mesenchymal transition. Oncoimmunology 2013, 2, e25820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raimondi, C.; Carpino, G.; Nicolazzo, C.; Gradilone, A.; Gianni, W.; Gelibter, A.; Gaudio, E.; Cortesi, E.; Gazzaniga, P. PD-L1 and epithelial-mesenchymal transition in circulating tumor cells from non-small cell lung cancer patients: A molecular shield to evade immune system? Oncoimmunology 2017, 6, e1315488. [Google Scholar] [CrossRef] [PubMed]
- Fidler, I.J. The pathogenesis of cancer metastasis: The ‘seed and soil’ hypothesis revisited. Nat. Rev. Cancer 2003, 3, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Mohme, M.; Riethdorf, S.; Pantel, K. Circulating and disseminated tumour cells—Mechanisms of immune surveillance and escape. Nat. Rev. Clin. Oncol. 2017, 14, 155–167. [Google Scholar] [CrossRef] [PubMed]
- Schaller, J.; Agudo, J. Metastatic Colonization: Escaping Immune Surveillance. Cancers 2020, 12, 3385. [Google Scholar] [CrossRef]
- Cheung, K.J.; Ewald, A.J. A collective route to metastasis: Seeding by tumor cell clusters. Science 2016, 352, 167–169. [Google Scholar] [CrossRef] [Green Version]
- Aceto, N.; Bardia, A.; Miyamoto, D.T.; Donaldson, M.C.; Wittner, B.S.; Spencer, J.A.; Yu, M.; Pely, A.; Engstrom, A.; Zhu, H.; et al. Circulating tumor cell clusters are oligoclonal precursors of breast cancer metastasis. Cell 2014, 158, 1110–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo, H.C.; Xu, Z.; Kim, I.S.; Pingel, B.; Aguirre, S.; Kodali, S.; Liu, J.; Zhang, W.; Muscarella, A.M.; Hein, S.M.; et al. Resistance to natural killer cell immunosurveillance confers a selective advantage to polyclonal metastasis. Nat. Cancer 2020, 1, 709–722. [Google Scholar] [CrossRef]
- Hosseini, H.; Obradović, M.M.S.; Hoffmann, M.; Harper, K.L.; Sosa, M.S.; Werner-Klein, M.; Nanduri, L.K.; Werno, C.; Ehrl, C.; Maneck, M.; et al. Early dissemination seeds metastasis in breast cancer. Nature 2016, 540, 552–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terunuma, H.; Deng, X.; Dewan, Z.; Fujimoto, S.; Yamamoto, N. Potential role of NK cells in the induction of immune responses: Implications for NK cell-based immunotherapy for cancers and viral infections. Int. Rev. Immunol. 2008, 27, 93–110. [Google Scholar] [CrossRef]
- Deng, X.; Terunuma, H.; Nieda, M. Immunosurveillance of cancer and viral infections with regard to alterations of human NK cells originating from lifestyle and aging. Biomedicines 2021, 9, 557. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Terunuma, H.; Nieda, M.; Xiao, W.; Nicol, A. Synergistic cytotoxicity of ex vivo expanded natural killer cells in combination with monoclonal antibody drugs against cancer cells. Int. Immunopharmacol. 2012, 14, 593–605. [Google Scholar] [CrossRef]
- Deng, X.; Terunuma, H.; Terunuma, A.; Takane, T.; Nieda, M. Ex vivo-expanded natural killer cells kill cancer cells more effectively than ex vivo-expanded γδ T cells or αβ T cells. Int. Immunopharmacol. 2014, 22, 486–491. [Google Scholar] [CrossRef] [PubMed]
- Imai, K.; Matsuyama, S.; Miyake, S.; Suga, K.; Nakachi, K. Natural cytotoxic activity of peripheral-blood lymphocytes and cancer incidence: An 11-year follow-up study of a general population. Lancet 2000, 356, 1795–1799. [Google Scholar] [CrossRef] [PubMed]
- Orange, J.S. Formation and function of the lytic NK-cell immunological synapse. Nat. Rev. Immunol. 2008, 8, 713–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gwalani, L.A.; Orange, J.S. Single Degranulations in NK Cells Can Mediate Target Cell Killing. J. Immunol. 2018, 200, 3231–3243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bots, M.; Medema, J.P. Granzymes at a glance. J. Cell Sci. 2006, 119, 5011–5014. [Google Scholar] [CrossRef] [Green Version]
- Smyth, M.J.; Thia, K.Y.; Cretney, E.; Kelly, J.M.; Snook, M.B.; Forbes, C.A.; Scalzo, A.A. Perforin is a major contributor to NK cell control of tumor metastasis. J. Immunol. 1999, 162, 6658–6662. [Google Scholar]
- Kodama, T.; Takeda, K.; Shimozato, O.; Hayakawa, Y.; Atsuta, M.; Kobayashi, K.; Ito, M.; Yagita, H.; Okumura, K. Perforin-dependent NK cell cytotoxicity is sufficient for anti-metastatic effect of IL-12. Eur. J. Immunol. 1999, 29, 1390–1396. [Google Scholar] [CrossRef]
- Bhat, R.; Watzl, C. Serial killing of tumor cells by human natural killer cells--enhancement by therapeutic antibodies. PLoS ONE 2007, 2, e326. [Google Scholar] [CrossRef]
- Prager, I.; Liesche, C.; van Ooijen, H.; Urlaub, D.; Verron, Q.; Sandström, N.; Fasbender, F.; Claus, M.; Eils, R.; Beaudouin, J.; et al. NK cells switch from granzyme B to death receptor-mediated cytotoxicity during serial killing. J. Exp. Med. 2019, 216, 2113–2127. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Iizuka, K.; Aguila, H.L.; Weissman, I.L.; Yokoyama, W.M. In vivo natural killer cell activities revealed by natural killer cell-deficient mice. Proc. Natl. Acad. Sci. USA 2000, 97, 2731–2736. [Google Scholar] [CrossRef] [Green Version]
- Dewan, M.Z.; Terunuma, H.; Takada, M.; Tanaka, Y.; Abe, H.; Sata, T.; Toi, M.; Yamamoto, N. Role of natural killer cells in hormone-independent rapid tumor formation and spontaneous metastasis of breast cancer cells in vivo. Breast Cancer Res. Treat. 2007, 104, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Hanna, N. Inhibition of experimental tumor metastasis by selective activation of natural killer cells. Cancer Res. 1982, 42, 1337–1342. [Google Scholar] [PubMed]
- Aboud, M.; Kingsmore, S.; Segal, S. Role of natural killer cells in controlling local tumor formation and metastatic manifestation of different 3LL Lewis lung carcinoma cell clones. Nat. Immun. 1993, 12, 17–24. [Google Scholar]
- Ichise, H.; Tsukamoto, S.; Hirashima, T.; Konishi, Y.; Oki, C.; Tsukiji, S.; Iwano, S.; Miyawaki, A.; Sumiyama, K.; Terai, K.; et al. Functional visualization of NK cell-mediated killing of metastatic single tumor cells. Elife 2022, 11, e76269. [Google Scholar] [CrossRef] [PubMed]
- Sampieri, K.; Fodde, R. Cancer stem cells and metastasis. Semin. Cancer Biol. 2012, 22, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Tallerico, R.; Conti, L.; Lanzardo, S.; Sottile, R.; Garofalo, C.; Wagner, A.K.; Johansson, M.H.; Cristiani, C.M.; Kärre, K.; Carbone, E.; et al. NK cells control breast cancer and related cancer stem cell hematological spread. Oncoimmunology 2017, 6, e1284718. [Google Scholar] [CrossRef] [Green Version]
- Ames, E.; Canter, R.J.; Grossenbacher, S.K.; Mac, S.; Chen, M.; Smith, R.C.; Hagino, T.; Perez-Cunningham, J.; Sckisel, G.D.; Urayama, S.; et al. NK Cells Preferentially Target Tumor Cells with a Cancer Stem Cell Phenotype. J. Immunol. 2015, 195, 4010–4019. [Google Scholar] [CrossRef] [Green Version]
- Chockley, P.J.; Chen, J.; Chen, G.; Beer, D.G.; Standiford, T.J.; Keshamouni, V.G. Epithelial-mesenchymal transition leads to NK cell-mediated metastasis-specific immunosurveillance in lung cancer. J. Clin. Investig. 2018, 128, 1384–1396. [Google Scholar] [CrossRef] [Green Version]
- Xu, B.; Chen, L.; Li, J.; Zheng, X.; Shi, L.; Wu, C.; Jiang, J. Prognostic value of tumor infiltrating NK cells and macrophages in stage II+III esophageal cancer patients. Oncotarget 2016, 7, 74904–74916. [Google Scholar] [CrossRef] [Green Version]
- Ishigami, S.; Natsugoe, S.; Tokuda, K.; Nakajo, A.; Che, X.; Iwashige, H.; Aridome, K.; Hokita, S.; Aikou, T. Prognostic value of intratumoral natural killer cells in gastric carcinoma. Cancer 2000, 88, 577–583. [Google Scholar] [CrossRef]
- Delahaye, N.F.; Rusakiewicz, S.; Martins, I.; Ménard, C.; Roux, S.; Lyonnet, L.; Paul, P.; Sarabi, M.; Chaput, N.; Semeraro, M.; et al. Alternatively spliced NKp30 isoforms affect the prognosis of gastrointestinal stromal tumors. Nat. Med. 2011, 17, 700–707. [Google Scholar] [CrossRef] [PubMed]
- Rusakiewicz, S.; Semeraro, M.; Sarabi, M.; Desbois, M.; Locher, C.; Mendez, R.; Vimond, N.; Concha, A.; Garrido, F.; Isambert, N.; et al. Immune infiltrates are prognostic factors in localized gastrointestinal stromal tumors. Cancer Res. 2013, 73, 3499–3510. [Google Scholar] [CrossRef] [PubMed]
- Donskov, F.; von der Maase, H. Impact of immune parameters on long-term survival in metastatic renal cell carcinoma. J. Clin. Oncol. 2006, 24, 1997–2005. [Google Scholar] [CrossRef] [PubMed]
- Coca, S.; Perez-Piqueras, J.; Martinez, D.; Colmenarejo, A.; Saez, M.A.; Vallejo, C.; Martos, J.A.; Moreno, M. The prognostic significance of intratumoral natural killer cells in patients with colorectal carcinoma. Cancer 1997, 79, 2320–2328. [Google Scholar] [CrossRef]
- Gannon, P.O.; Poisson, A.O.; Delvoye, N.; Lapointe, R.; Mes-Masson, A.M.; Saad, F. Characterization of the intra-prostatic immune cell infiltration in androgen-deprived prostate cancer patients. J. Immunol. Methods 2009, 348, 9–17. [Google Scholar] [CrossRef]
- Pasero, C.; Gravis, G.; Granjeaud, S.; Guerin, M.; Thomassin-Piana, J.; Rocchi, P.; Salem, N.; Walz, J.; Moretta, A.; Olive, D. Highly effective NK cells are associated with good prognosis in patients with metastatic prostate cancer. Oncotarget 2015, 6, 14360–14373. [Google Scholar] [CrossRef] [Green Version]
- Brodbeck, T.; Nehmann, N.; Bethge, A.; Wedemann, G.; Schumacher, U. Perforin-dependent direct cytotoxicity in natural killer cells induces considerable knockdown of spontaneous lung metastases and computer modelling-proven tumor cell dormancy in a HT29 human colon cancer xenograft mouse model. Mol. Cancer 2014, 13, 244. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Chen, Q.; Yan, J.; Wang, Y.; Zhu, C.; Chen, C.; Zhao, X.; Xu, M.; Sun, Q.; Deng, R.; et al. MiRNA-296-3p-ICAM-1 axis promotes metastasis of prostate cancer by possible enhancing survival of natural killer cell-resistant circulating tumour cells. Cell Death Dis. 2013, 4, e928. [Google Scholar] [CrossRef] [Green Version]
- Uchida, A. The cytolytic and regulatory role of natural killer cells in human neoplasia. Biochim. Biophys. Acta 1986, 865, 329–340. [Google Scholar] [CrossRef]
- Green, T.L.; Cruse, J.M.; Lewis, R.E.; Craft, B.S. Circulating tumor cells (CTCs) from metastatic breast cancer patients linked to decreased immune function and response to treatment. Exp. Mol. Pathol. 2013, 95, 174–179. [Google Scholar] [CrossRef]
- Green, T.L.; Santos, M.F.; Ejaeidi, A.A.; Craft, B.S.; Lewis, R.E.; Cruse, J.M. Toll-like receptor (TLR) expression of immune system cells from metastatic breast cancer patients with circulating tumor cells. Exp. Mol. Pathol. 2014, 97, 44–48. [Google Scholar] [CrossRef] [PubMed]
- Schantz, S.P.; Campbell, B.H.; Guillamondegui, O.M. Pharyngeal carcinoma and natural killer cell activity. Am. J. Surg 1986, 152, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Koda, K.; Saito, N.; Oda, K.; Seike, K.; Kondo, E.; Ishizuka, M.; Takiguchi, N.; Miyazaki, M. Natural killer cell activity and distant metastasis in rectal cancers treated surgically with and without neoadjuvant chemoradiotherapy. J. Am. Coll. Surg. 2003, 197, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Kondo, E.; Koda, K.; Takiguchi, N.; Oda, K.; Seike, K.; Ishizuka, M.; Miyazaki, M. Preoperative natural killer cell activity as a prognostic factor for distant metastasis following surgery for colon cancer. Dig. Surg. 2003, 20, 445–451. [Google Scholar] [CrossRef]
- Koda, K.; Saito, N.; Takiguchi, N.; Oda, K.; Nunomura, M.; Nakajima, N. Preoperative natural killer cell activity: Correlation with distant metastases in curatively research colorectal carcinomas. Int. Surg. 1997, 82, 190–193. [Google Scholar]
- Tai, L.H.; de Souza, C.T.; Bélanger, S.; Ly, L.; Alkayyal, A.A.; Zhang, J.; Rintoul, J.L.; Ananth, A.A.; Lam, T.; Breitbach, C.J.; et al. Preventing postoperative metastatic disease by inhibiting surgery-induced dysfunction in natural killer cells. Cancer Res. 2013, 73, 97–107. [Google Scholar] [CrossRef] [Green Version]
- Tai, L.H.; Zhang, J.; Scott, K.J.; de Souza, C.T.; Alkayyal, A.A.; Ananth, A.A.; Sahi, S.; Adair, R.A.; Mahmoud, A.B.; Sad, S.; et al. Perioperative influenza vaccination reduces postoperative metastatic disease by reversing surgery-induced dysfunction in natural killer cells. Clin. Cancer Res. 2013, 19, 5104–5115. [Google Scholar] [CrossRef] [Green Version]
- Goldfarb, Y.; Sorski, L.; Benish, M.; Levi, B.; Melamed, R.; Ben-Eliyahu, S. Improving postoperative immune status and resistance to cancer metastasis: A combined perioperative approach of immunostimulation and prevention of excessive surgical stress responses. Ann. Surg. 2011, 253, 798–810. [Google Scholar] [CrossRef]
- Angka, L.; Khan, S.T.; Kilgour, M.K.; Xu, R.; Kennedy, M.A.; Auer, R.C. Dysfunctional Natural Killer Cells in the Aftermath of Cancer Surgery. Int. J. Mol. Sci. 2017, 18, 1787. [Google Scholar] [CrossRef] [Green Version]
- Angka, L.; Martel, A.B.; Kilgour, M.; Jeong, A.; Sadiq, M.; de Souza, C.T.; Baker, L.; Kennedy, M.A.; Kekre, N.; Auer, R.C. Natural Killer Cell IFNγ Secretion is Profoundly Suppressed Following Colorectal Cancer Surgery. Ann. Surg. Oncol. 2018, 25, 3747–3754. [Google Scholar] [CrossRef]
- Espí, A.; Arenas, J.; García-Granero, E.; Martí, E.; Lledó, S. Relationship of curative surgery on natural killer cell activity in colorectal cancer. Dis. Colon. Rectum. 1996, 39, 429–434. [Google Scholar] [CrossRef]
- Pollock, R.E.; Lotzová, E.; Stanford, S.D. Surgical stress impairs natural killer cell programming of tumor for lysis in patients with sarcomas and other solid tumors. Cancer 1992, 70, 2192–2202. [Google Scholar] [CrossRef] [PubMed]
- Pollock, R.E.; Lotzová, E.; Stanford, S.D. Mechanism of surgical stress impairment of human perioperative natural killer cell cytotoxicity. Arch. Surg. 1991, 126, 338–342. [Google Scholar] [CrossRef] [PubMed]
- Market, M.; Baxter, K.E.; Angka, L.; Kennedy, M.A.; Auer, R.C. The Potential for Cancer Immunotherapy in Targeting Surgery-Induced Natural Killer Cell Dysfunction. Cancers 2018, 11, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Vito, C.; Mikulak, J.; Zaghi, E.; Pesce, S.; Marcenaro, E.; Mavilio, D. NK cells to cure cancer. Semin. Immunol. 2019, 41, 101272. [Google Scholar] [CrossRef]
- Wang, B.; Wang, Q.; Wang, Z.; Jiang, J.; Yu, S.C.; Ping, Y.F.; Yang, J.; Xu, S.L.; Ye, X.Z.; Xu, C.; et al. Metastatic consequences of immune escape from NK cell cytotoxicity by human breast cancer stem cells. Cancer Res. 2014, 74, 5746–5757. [Google Scholar] [CrossRef] [Green Version]
- Heinemann, A.; Zhao, F.; Pechlivanis, S.; Eberle, J.; Steinle, A.; Diederichs, S.; Schadendorf, D.; Paschen, A. Tumor suppressive microRNAs miR-34a/c control cancer cell expression of ULBP2, a stress-induced ligand of the natural killer cell receptor NKG2D. Cancer Res. 2012, 72, 460–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, A.; Yan, W.H. Human Leukocyte Antigen-G (HLA-G) Expression in Cancers: Roles in Immune Evasion, Metastasis and Target for Therapy. Mol. Med. 2015, 21, 782–791. [Google Scholar] [CrossRef] [PubMed]
- Carosella, E.D.; Rouas-Freiss, N.; Tronik-Le Roux, D.; Moreau, P.; LeMaoult, J. HLA-G: An Immune Checkpoint Molecule. Adv. Immunol. 2015, 127, 33–144. [Google Scholar] [CrossRef]
- Swets, M.; König, M.H.; Zaalberg, A.; Dekker-Ensink, N.G.; Gelderblom, H.; van de Velde, C.J.; van den Elsen, P.J.; Kuppen, P.J. HLA-G and classical HLA class I expression in primary colorectal cancer and associated liver metastases. Hum. Immunol. 2016, 77, 773–779. [Google Scholar] [CrossRef] [PubMed]
- König, L.; Kasimir-Bauer, S.; Hoffmann, O.; Bittner, A.K.; Wagner, B.; Manvailer, L.F.; Schramm, S.; Bankfalvi, A.; Giebel, B.; Kimmig, R.; et al. The prognostic impact of soluble and vesicular HLA-G and its relationship to circulating tumor cells in neoadjuvant treated breast cancer patients. Hum. Immunol. 2016, 77, 791–799. [Google Scholar] [CrossRef] [PubMed]
- Giuliani, M.; Janji, B.; Berchem, G. Activation of NK cells and disruption of PD-L1/PD-1 axis: Two different ways for lenalidomide to block myeloma progression. Oncotarget 2017, 8, 24031–24044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sordo-Bahamonde, C.; Lorenzo-Herrero, S.; Payer, Á.R.; Gonzalez, S.; López-Soto, A. Mechanisms of Apoptosis Resistance to NK Cell-Mediated Cytotoxicity in Cancer. Int. J. Mol. Sci. 2020, 21, 3726. [Google Scholar] [CrossRef] [PubMed]
- Russick, J.; Joubert, P.E.; Gillard-Bocquet, M.; Torset, C.; Meylan, M.; Petitprez, F.; Dragon-Durey, M.A.; Marmier, S.; Varthaman, A.; Josseaume, N.; et al. Natural killer cells in the human lung tumor microenvironment display immune inhibitory functions. J. Immunother. Cancer 2020, 8, e001054. [Google Scholar] [CrossRef] [PubMed]
- Pasero, C.; Gravis, G.; Guerin, M.; Granjeaud, S.; Thomassin-Piana, J.; Rocchi, P.; Paciencia-Gros, M.; Poizat, F.; Bentobji, M.; Azario-Cheillan, F.; et al. Inherent and Tumor-Driven Immune Tolerance in the Prostate Microenvironment Impairs Natural Killer Cell Antitumor Activity. Cancer Res. 2016, 76, 2153–2165. [Google Scholar] [CrossRef] [Green Version]
- Mamessier, E.; Sylvain, A.; Thibult, M.L.; Houvenaeghel, G.; Jacquemier, J.; Castellano, R.; Gonçalves, A.; André, P.; Romagné, F.; Thibault, G.; et al. Human breast cancer cells enhance self tolerance by promoting evasion from NK cell antitumor immunity. J. Clin. Investig. 2011, 121, 3609–3622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Treon, S.P.; Hansen, M.; Branagan, A.R.; Verselis, S.; Emmanouilides, C.; Kimby, E.; Frankel, S.R.; Touroutoglou, N.; Turnbull, B.; Anderson, K.C.; et al. Polymorphisms in FcgammaRIIIA (CD16) receptor expression are associated with clinical response to rituximab in Waldenström’s macroglobulinemia. J. Clin. Oncol. 2005, 23, 474–481. [Google Scholar] [CrossRef] [Green Version]
- Rocca, Y.S.; Roberti, M.P.; Juliá, E.P.; Pampena, M.B.; Bruno, L.; Rivero, S.; Huertas, E.; Sánchez Loria, F.; Pairola, A.; Caignard, A.; et al. Phenotypic and Functional Dysregulated Blood NK Cells in Colorectal Cancer Patients Can Be Activated by Cetuximab Plus IL-2 or IL-15. Front. Immunol. 2016, 7, 413. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.P.; Zhu, Y.; Zhang, J.J.; Xu, Z.K.; Qian, Z.Y.; Dai, C.C.; Jiang, K.R.; Wu, J.L.; Gao, W.T.; Li, Q.; et al. Comprehensive analysis of the percentage of surface receptors and cytotoxic granules positive natural killer cells in patients with pancreatic cancer, gastric cancer, and colorectal cancer. J. Transl. Med. 2013, 11, 262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Iglesias, T.; Del Toro-Arreola, A.; Albarran-Somoza, B.; Del Toro-Arreola, S.; Sanchez-Hernandez, P.E.; Ramirez-Dueñas, M.G.; Balderas-Peña, L.M.; Bravo-Cuellar, A.; Ortiz-Lazareno, P.C.; Daneri-Navarro, A. Low NKp30, NKp46 and NKG2D expression and reduced cytotoxic activity on NK cells in cervical cancer and precursor lesions. BMC Cancer 2009, 9, 186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez-Correa, B.; Morgado, S.; Gayoso, I.; Bergua, J.M.; Casado, J.G.; Arcos, M.J.; Bengochea, M.L.; Duran, E.; Solana, R.; Tarazona, R. Human NK cells in acute myeloid leukaemia patients: Analysis of NK cell-activating receptors and their ligands. Cancer Immunol. Immunother 2011, 60, 1195–1205. [Google Scholar] [CrossRef] [PubMed]
- Amo, L.; Tamayo-Orbegozo, E.; Maruri, N.; Buqué, A.; Solaun, M.; Riñón, M.; Arrieta, A.; Larrucea, S. Podocalyxin-like protein 1 functions as an immunomodulatory molecule in breast cancer cells. Cancer Lett. 2015, 368, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Mercier-Bataille, D.; Sanchez, C.; Baier, C.; Le Treut, T.; Mounier, N.; Mokhtari, S.; Olive, D.; Baumstarck, K.; Sébahoun, G.; Besson, C.; et al. Expression of activating receptors on natural killer cells from AIDS-related lymphoma patients. AIDS Res. Ther. 2014, 11, 38. [Google Scholar] [CrossRef] [PubMed]
- Hoechst, B.; Voigtlaender, T.; Ormandy, L.; Gamrekelashvili, J.; Zhao, F.; Wedemeyer, H.; Lehner, F.; Manns, M.P.; Greten, T.F.; Korangy, F. Myeloid derived suppressor cells inhibit natural killer cells in patients with hepatocellular carcinoma via the NKp30 receptor. Hepatology 2009, 50, 799–807. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Yin, T.; Li, D.; Gao, X.; Wan, Y.; Ma, X.; Ye, T.; Guo, F.; Sun, J.; Lin, Z.; et al. Enhanced interaction between natural killer cells and lung cancer cells: Involvement in gefitinib-mediated immunoregulation. J. Transl. Med. 2013, 11, 186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Kruijf, E.M.; Sajet, A.; van Nes, J.G.; Putter, H.; Smit, V.T.; Eagle, R.A.; Jafferji, I.; Trowsdale, J.; Liefers, G.J.; van de Velde, C.J.; et al. NKG2D ligand tumor expression and association with clinical outcome in early breast cancer patients: An observational study. BMC Cancer 2012, 12, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vuletić, A.; Jurišić, V.; Jovanić, I.; Milovanović, Z.; Nikolić, S.; Konjević, G. Distribution of several activating and inhibitory receptors on CD3(-)CD56(+) NK cells in regional lymph nodes of melanoma patients. J. Surg. Res. 2013, 183, 860–868. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Deng, Y.; Hao, J.W.; Li, Y.; Liu, B.; Yu, Y.; Shi, F.D.; Zhou, Q.H. NK cell phenotypic modulation in lung cancer environment. PLoS ONE 2014, 9, e109976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bossard, C.; Bézieau, S.; Matysiak-Budnik, T.; Volteau, C.; Laboisse, C.L.; Jotereau, F.; Mosnier, J.F. HLA-E/β2 microglobulin overexpression in colorectal cancer is associated with recruitment of inhibitory immune cells and tumor progression. Int. J. Cancer 2012, 131, 855–863. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Cheng, Y.; Xu, Y.; Wang, Z.; Du, X.; Li, C.; Peng, J.; Gao, L.; Liang, X.; Ma, C. Increased expression of programmed cell death protein 1 on NK cells inhibits NK-cell-mediated anti-tumor function and indicates poor prognosis in digestive cancers. Oncogene 2017, 36, 6143–6153. [Google Scholar] [CrossRef] [Green Version]
- Trefny, M.P.; Kaiser, M.; Stanczak, M.A.; Herzig, P.; Savic, S.; Wiese, M.; Lardinois, D.; Läubli, H.; Uhlenbrock, F.; Zippelius, A. PD-1+ natural killer cells in human non-small cell lung cancer can be activated by PD-1/PD-L1 blockade. Cancer Immunol. Immunother. 2020, 69, 1505–1517. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Huang, Y.; Tan, L.; Yu, W.; Chen, D.; Lu, C.; He, J.; Wu, G.; Liu, X.; Zhang, Y. Increased Tim-3 expression in peripheral NK cells predicts a poorer prognosis and Tim-3 blockade improves NK cell-mediated cytotoxicity in human lung adenocarcinoma. Int. Immunopharmacol. 2015, 29, 635–641. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhu, J.; Gu, H.; Yuan, Y.; Zhang, B.; Zhu, D.; Zhou, J.; Zhu, Y.; Chen, W. The Clinical Significance of Abnormal Tim-3 Expression on NK Cells from Patients with Gastric Cancer. Immunol. Investig. 2015, 44, 578–589. [Google Scholar] [CrossRef] [PubMed]
- da Silva, I.P.; Gallois, A.; Jimenez-Baranda, S.; Khan, S.; Anderson, A.C.; Kuchroo, V.K.; Osman, I.; Bhardwaj, N. Reversal of NK-cell exhaustion in advanced melanoma by Tim-3 blockade. Cancer Immunol. Res. 2014, 2, 410–422. [Google Scholar] [CrossRef] [Green Version]
- Viel, S.; Marçais, A.; Guimaraes, F.S.; Loftus, R.; Rabilloud, J.; Grau, M.; Degouve, S.; Djebali, S.; Sanlaville, A.; Charrier, E.; et al. TGF-beta inhibits the activation and functions of NK cells by repressing the mTOR pathway. Sci. Signal 2016, 9, ra19. [Google Scholar] [CrossRef]
- Bozward, A.G.; Warricker, F.; Oo, Y.H.; Khakoo, S.I. Natural Killer Cells and Regulatory T Cells Cross Talk in Hepatocellular Carcinoma: Exploring Therapeutic Options for the Next Decade. Front. Immunol. 2021, 12, 643310. [Google Scholar] [CrossRef]
- Li, H.; Han, Y.; Guo, Q.; Zhang, M.; Cao, X. Cancer-expanded myeloid-derived suppressor cells induce anergy of NK cells through membrane-bound TGF-beta 1. J. Immunol. 2009, 182, 240–249. [Google Scholar] [CrossRef] [Green Version]
- Spiegel, A.; Brooks, M.W.; Houshyar, S.; Reinhardt, F.; Ardolino, M.; Fessler, E.; Chen, M.B.; Krall, J.A.; DeCock, J.; Zervantonakis, I.K.; et al. Neutrophils Suppress Intraluminal NK Cell-Mediated Tumor Cell Clearance and Enhance Extravasation of Disseminated Carcinoma Cells. Cancer Discov. 2016, 6, 630–649. [Google Scholar] [CrossRef] [Green Version]
- Cazzetta, V.; Franzese, S.; Carenza, C.; Della Bella, S.; Mikulak, J.; Mavilio, D. Natural Killer-Dendritic Cell Interactions in Liver Cancer: Implications for Immunotherapy. Cancers 2021, 13, 2184. [Google Scholar] [CrossRef]
- Li, P.; Lu, M.; Shi, J.; Hua, L.; Gong, Z.; Li, Q.; Shultz, L.D.; Ren, G. Dual roles of neutrophils in metastatic colonization are governed by the host NK cell status. Nat. Commun. 2020, 11, 4387. [Google Scholar] [CrossRef]
- Chan, I.S.; Knútsdóttir, H.; Ramakrishnan, G.; Padmanaban, V.; Warrier, M.; Ramirez, J.C.; Dunworth, M.; Zhang, H.; Jaffee, E.M.; Bader, J.S.; et al. Cancer cells educate natural killer cells to a metastasis-promoting cell state. J. Cell Biol. 2020, 219, e202001134. [Google Scholar] [CrossRef] [PubMed]
- Machlus, K.R.; Italiano, J.E., Jr. The incredible journey: From megakaryocyte development to platelet formation. J. Cell Biol. 2013, 201, 785–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sacks, D.; Baxter, B.; Campbell, B.C.V.; Carpenter, J.S.; Cognard, C.; Dippel, D.; Eesa, M.; Fischer, U.; Hausegger, K.; Hirsch, J.A.; et al. Multisociety Consensus Quality Improvement Revised Consensus Statement for Endovascular Therapy of Acute Ischemic Stroke. Int. J. Stroke 2018, 13, 612–632. [Google Scholar] [CrossRef]
- Schmied, L.; Höglund, P.; Meinke, S. Platelet-Mediated Protection of Cancer Cells from Immune Surveillance—Possible Implications for Cancer Immunotherapy. Front. Immunol. 2021, 12, 640578. [Google Scholar] [CrossRef] [PubMed]
- Maouia, A.; Rebetz, J.; Kapur, R.; Semple, J.W. The Immune Nature of Platelets Revisited. Transfus. Med. Rev. 2020, 34, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Placke, T.; Kopp, H.G.; Salih, H.R. Modulation of natural killer cell anti-tumor reactivity by platelets. J. Innate Immun. 2011, 3, 374–382. [Google Scholar] [CrossRef] [PubMed]
- Palumbo, J.S.; Talmage, K.E.; Massari, J.V.; La Jeunesse, C.M.; Flick, M.J.; Kombrinck, K.W.; Jirousková, M.; Degen, J.L. Platelets and fibrin(ogen) increase metastatic potential by impeding natural killer cell-mediated elimination of tumor cells. Blood 2005, 105, 178–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Placke, T.; Örgel, M.; Schaller, M.; Jung, G.; Rammensee, H.G.; Kopp, H.G.; Salih, H.R. Platelet-derived MHC class I confers a pseudonormal phenotype to cancer cells that subverts the antitumor reactivity of natural killer immune cells. Cancer Res. 2012, 72, 440–448. [Google Scholar] [CrossRef] [Green Version]
- Placke, T.; Salih, H.R.; Kopp, H.G. GITR ligand provided by thrombopoietic cells inhibits NK cell antitumor activity. J. Immunol. 2012, 189, 154–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Placke, T.; Kopp, H.G.; Salih, H.R. The wolf in sheep’s clothing: Platelet-derived “pseudo self” impairs cancer cell “missing self” recognition by NK cells. Oncoimmunology 2012, 1, 557–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maurer, S.; Kropp, K.N.; Klein, G.; Steinle, A.; Haen, S.P.; Walz, J.S.; Hinterleitner, C.; Märklin, M.; Kopp, H.G.; Salih, H.R. Platelet-mediated shedding of NKG2D ligands impairs NK cell immune-surveillance of tumor cells. Oncoimmunology 2017, 7, e1364827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopp, H.G.; Placke, T.; Salih, H.R. Platelet-derived transforming growth factor-beta down-regulates NKG2D thereby inhibiting natural killer cell antitumor reactivity. Cancer Res. 2009, 69, 7775–7783. [Google Scholar] [CrossRef] [Green Version]
- Leblanc, R.; Peyruchaud, O. Metastasis: New functional implications of platelets and megakaryocytes. Blood 2016, 128, 24–31. [Google Scholar] [CrossRef]
- Yang, Y. Cancer immunotherapy: Harnessing the immune system to battle cancer. J. Clin. Investig. 2015, 125, 3335–3337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillerey, C.; Huntington, N.D.; Smyth, M.J. Targeting natural killer cells in cancer immunotherapy. Nat. Immunol. 2016, 17, 1025–1036. [Google Scholar] [CrossRef]
- Lorenzo-Herrero, S.; López-Soto, A.; Sordo-Bahamonde, C.; Gonzalez-Rodriguez, A.P.; Vitale, M.; Gonzalez, S. NK Cell-Based Immunotherapy in Cancer Metastasis. Cancers 2018, 11, 29. [Google Scholar] [CrossRef] [Green Version]
- Romee, R.; Leong, J.W.; Fehniger, T.A. Utilizing cytokines to function-enable human NK cells for the immunotherapy of cancer. Scientifica 2014, 2014, 205796. [Google Scholar] [CrossRef] [Green Version]
- Wrangle, J.M.; Patterson, A.; Johnson, C.B.; Neitzke, D.J.; Mehrotra, S.; Denlinger, C.E.; Paulos, C.M.; Li, Z.; Cole, D.J.; Rubinstein, M.P. IL-2 and Beyond in Cancer Immunotherapy. J. Interferon. Cytokine Res. 2018, 38, 45–68. [Google Scholar] [CrossRef]
- Rosenberg, S.A.; Lotze, M.T.; Muul, L.M.; Leitman, S.; Chang, A.E.; Ettinghausen, S.E.; Matory, Y.L.; Skibber, J.M.; Shiloni, E.; Vetto, J.T.; et al. Observations on the systemic administration of autologous lymphokine-activated killer cells and recombinant interleukin-2 to patients with metastatic cancer. N. Engl. J. Med. 1985, 313, 1485–1492. [Google Scholar] [CrossRef]
- Rosenberg, S.A. IL-2: The first effective immunotherapy for human cancer. J. Immunol. 2014, 192, 5451–5458. [Google Scholar] [CrossRef] [Green Version]
- Ahmadzadeh, M.; Rosenberg, S.A. IL-2 administration increases CD4+ CD25(hi) Foxp3+ regulatory T cells in cancer patients. Blood 2006, 107, 2409–2414. [Google Scholar] [CrossRef] [Green Version]
- Sim, G.C.; Liu, C.; Wang, E.; Liu, H.; Creasy, C.; Dai, Z.; Overwijk, W.W.; Roszik, J.; Marincola, F.; Hwu, P.; et al. IL2 Variant Circumvents ICOS+ Regulatory T-cell Expansion and Promotes NK Cell Activation. Cancer Immunol. Res. 2016, 4, 983–994. [Google Scholar] [CrossRef] [Green Version]
- Conlon, K.C.; Lugli, E.; Welles, H.C.; Rosenberg, S.A.; Fojo, A.T.; Morris, J.C.; Fleisher, T.A.; Dubois, S.P.; Perera, L.P.; Stewart, D.M.; et al. Redistribution, hyperproliferation, activation of natural killer cells and CD8 T cells, and cytokine production during first-in-human clinical trial of recombinant human interleukin-15 in patients with cancer. J. Clin. Oncol. 2015, 33, 74–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terunuma, H.; Deng, X.; Nishino, N.; Watanabe, K. NK cell-based autologous immune enhancement therapy (AIET) for cancer. J. Stem Cells Regen. Med. 2013, 9, 9–13. [Google Scholar] [CrossRef]
- Ruggeri, L.; Mancusi, A.; Burchielli, E.; Capanni, M.; Carotti, A.; Aloisi, T.; Aversa, F.; Martelli, M.F.; Velardi, A. NK cell alloreactivity and allogeneic hematopoietic stem cell transplantation. Blood Cells Mol. Dis. 2008, 40, 84–90. [Google Scholar] [CrossRef]
- Lin, M.; Liang, S.Z.; Shi, J.; Niu, L.Z.; Chen, J.B.; Zhang, M.J.; Xu, K.C. Circulating tumor cell as a biomarker for evaluating allogenic NK cell immunotherapy on stage IV non-small cell lung cancer. Immunol. Lett. 2017, 191, 10–15. [Google Scholar] [CrossRef]
- Liang, S.; Xu, K.; Niu, L.; Wang, X.; Liang, Y.; Zhang, M.; Chen, J.; Lin, M. Comparison of autogeneic and allogeneic natural killer cells immunotherapy on the clinical outcome of recurrent breast cancer. Onco. Targets Ther. 2017, 10, 4273–4281. [Google Scholar] [CrossRef] [Green Version]
- Qin, Z.; Chen, J.; Zeng, J.; Niu, L.; Xie, S.; Wang, X.; Liang, Y.; Wu, Z.; Zhang, M. Effect of NK cell immunotherapy on immune function in patients with hepatic carcinoma: A preliminary clinical study. Cancer Biol. Ther. 2017, 18, 323–330. [Google Scholar] [CrossRef] [Green Version]
- Alnaggar, M.; Lin, M.; Mesmar, A.; Liang, S.; Qaid, A.; Xu, K.; Chen, J.; Niu, L.; Yin, Z. Allogenic Natural Killer Cell Immunotherapy Combined with Irreversible Electroporation for Stage IV Hepatocellular Carcinoma: Survival Outcome. Cell Physiol. Biochem. 2018, 48, 1882–1893. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.; Liang, S.; Wang, X.; Liang, Y.; Zhang, M.; Chen, J.; Niu, L.; Xu, K. Short-term clinical efficacy of percutaneous irreversible electroporation combined with allogeneic natural killer cell for treating metastatic pancreatic cancer. Immunol. Lett. 2017, 186, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Adotevi, O.; Godet, Y.; Galaine, J.; Lakkis, Z.; Idirene, I.; Certoux, J.M.; Jary, M.; Loyon, R.; Laheurte, C.; Kim, S.; et al. In situ delivery of allogeneic natural killer cell (NK) combined with Cetuximab in liver metastases of gastrointestinal carcinoma: A phase I clinical trial. Oncoimmunology 2018, 7, e1424673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geller, M.A.; Cooley, S.; Judson, P.L.; Ghebre, R.; Carson, L.F.; Argenta, P.A.; Jonson, A.L.; Panoskaltsis-Mortari, A.; Curtsinger, J.; McKenna, D.; et al. A phase II study of allogeneic natural killer cell therapy to treat patients with recurrent ovarian and breast cancer. Cytotherapy 2011, 13, 98–107. [Google Scholar] [CrossRef] [Green Version]
- Lin, M.; Liang, S.Z.; Wang, X.H.; Liang, Y.Q.; Zhang, M.J.; Niu, L.Z.; Chen, J.B.; Li, H.B.; Xu, K.C. Clinical efficacy of percutaneous cryoablation combined with allogenic NK cell immunotherapy for advanced non-small cell lung cancer. Immunol. Res. 2017, 65, 880–887. [Google Scholar] [CrossRef] [PubMed]
- Dalle, J.H.; Menezes, J.; Wagner, E.; Blagdon, M.; Champagne, J.; Champagne, M.A.; Duval, M. Characterization of cord blood natural killer cells: Implications for transplantation and neonatal infections. Pediatr. Res. 2005, 57, 649–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arai, S.; Meagher, R.; Swearingen, M.; Myint, H.; Rich, E.; Martinson, J.; Klingemann, H. Infusion of the allogeneic cell line NK-92 in patients with advanced renal cell cancer or melanoma: A phase I trial. Cytotherapy 2008, 10, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Berrien-Elliott, M.M.; Wagner, J.A.; Fehniger, T.A. Human Cytokine-Induced Memory-Like Natural Killer Cells. J. Innate Immun. 2015, 7, 563–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romee, R.; Rosario, M.; Berrien-Elliott, M.M.; Wagner, J.A.; Jewell, B.A.; Schappe, T.; Leong, J.W.; Abdel-Latif, S.; Schneider, S.E.; Willey, S.; et al. Cytokine-induced memory-like natural killer cells exhibit enhanced responses against myeloid leukemia. Sci. Transl. Med. 2016, 8, 357ra123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, G.; Dong, H.; Liang, Y.; Ham, J.D.; Rizwan, R.R.; Chen, J. CAR-NK cells: A promising cellular immunotherapy for cancer. EBioMedicine 2020, 59, 102975. [Google Scholar] [CrossRef] [PubMed]
- Liu, E.; Marin, D.; Banerjee, P.; Macapinlac, H.A.; Thompson, P.; Basar, R.; Nassif Kerbauy, L.; Overman, B.; Thall, P.; Kaplan, M.; et al. Use of CAR-Transduced Natural Killer Cells in CD19-Positive Lymphoid Tumors. N. Engl. J. Med. 2020, 382, 545–553. [Google Scholar] [CrossRef]
- Schönfeld, K.; Sahm, C.; Zhang, C.; Naundorf, S.; Brendel, C.; Odendahl, M.; Nowakowska, P.; Bönig, H.; Köhl, U.; Kloess, S.; et al. Selective inhibition of tumor growth by clonal NK cells expressing an ErbB2/HER2-specific chimeric antigen receptor. Mol. Ther. 2015, 23, 330–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ochoa, M.C.; Minute, L.; Rodriguez, I.; Garasa, S.; Perez-Ruiz, E.; Inogés, S.; Melero, I.; Berraondo, P. Antibody-dependent cell cytotoxicity: Immunotherapy strategies enhancing effector NK cells. Immunol. Cell Biol. 2017, 95, 347–355. [Google Scholar] [CrossRef]
- Weiner, L.M.; Surana, R.; Wang, S. Monoclonal antibodies: Versatile platforms for cancer immunotherapy. Nat. Rev. Immunol. 2010, 10, 317–327. [Google Scholar] [CrossRef] [Green Version]
- Offidani, M.; Corvatta, L.; Morè, S.; Nappi, D.; Martinelli, G.; Olivieri, A.; Cerchione, C. Daratumumab for the Management of Newly Diagnosed and Relapsed/Refractory Multiple Myeloma: Current and Emerging Treatments. Front. Oncol. 2021, 10, 624661. [Google Scholar] [CrossRef] [PubMed]
- Passey, C.; Sheng, J.; Mora, J.; Tendolkar, A.; Robbins, M.; Dodge, R.; Roy, A.; Bello, A.; Gupta, M. The Clinical Pharmacology of Elotuzumab. Clin. Pharmacokinet. 2018, 57, 297–313. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, J.; Zarate, R.; Bandres, E.; Boni, V.; Hernández, A.; Sola, J.J.; Honorato, B.; Bitarte, N.; García-Foncillas, J. Fc gamma receptor polymorphisms as predictive markers of Cetuximab efficacy in epidermal growth factor receptor downstream-mutated metastatic colorectal cancer. Eur. J. Cancer 2012, 48, 1774–1780. [Google Scholar] [CrossRef] [PubMed]
- Federico, S.M.; McCarville, M.B.; Shulkin, B.L.; Sondel, P.M.; Hank, J.A.; Hutson, P.; Meagher, M.; Shafer, A.; Ng, C.Y.; Leung, W.; et al. A Pilot Trial of Humanized Anti-GD2 Monoclonal Antibody (hu14.18K322A) with Chemotherapy and Natural Killer Cells in Children with Recurrent/Refractory Neuroblastoma. Clin. Cancer Res. 2017, 23, 6441–6449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Modak, S.; Le Luduec, J.B.; Cheung, I.Y.; Goldman, D.A.; Ostrovnaya, I.; Doubrovina, E.; Basu, E.; Kushner, B.H.; Kramer, K.; Roberts, S.S.; et al. Adoptive immunotherapy with haploidentical natural killer cells and Anti-GD2 monoclonal antibody m3F8 for resistant neuroblastoma: Results of a phase I study. Oncoimmunology 2018, 7, e1461305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogel, C.L.; Cobleigh, M.A.; Tripathy, D.; Gutheil, J.C.; Harris, L.N.; Fehrenbacher, L.; Slamon, D.J.; Murphy, M.; Novotny, W.F.; Burchmore, M.; et al. Efficacy and safety of trastuzumab as a single agent in first-line treatment of HER2-overexpressing metastatic breast cancer. J. Clin. Oncol. 2002, 20, 719–726. [Google Scholar] [CrossRef] [PubMed]
- Takada, M.; Terunuma, H.; Deng, X.; Dewan, M.Z.; Saji, S.; Kuroi, K.; Yamamoto, N.; Toi, M. Refractory lung metastasis from breast cancer treated with multidisciplinary therapy including an immunological approach. Breast Cancer 2011, 18, 64–67. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.C.; Shimasaki, N.; Lim, J.S.J.; Wong, A.; Yadav, K.; Yong, W.P.; Tan, L.K.; Koh, L.P.; Poon, M.L.M.; Tan, S.H.; et al. Phase I Trial of Expanded, Activated Autologous NK-cell Infusions with Trastuzumab in Patients with HER2-positive Cancers. Clin. Cancer Res. 2020, 26, 4494–4502. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.M.; Liou, A.; Poon, M.; Koh, L.P.; Tan, L.K.; Loh, K.S.; Petersson, B.F.; Ting, E.; Campana, D.; Goh, B.C.; et al. Phase I study of expanded natural killer cells in combination with cetuximab for recurrent/metastatic nasopharyngeal carcinoma. Cancer Immunol. Immunother. 2022, 71, 2277–2286. [Google Scholar] [CrossRef] [PubMed]
- Gleason, M.K.; Verneris, M.R.; Todhunter, D.A.; Zhang, B.; McCullar, V.; Zhou, S.X.; Panoskaltsis-Mortari, A.; Weiner, L.M.; Vallera, D.A.; Miller, J.S. Bispecific and trispecific killer cell engagers directly activate human NK cells through CD16 signaling and induce cytotoxicity and cytokine production. Mol. Cancer Ther. 2012, 11, 2674–2684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohrt, H.E.; Thielens, A.; Marabelle, A.; Sagiv-Barfi, I.; Sola, C.; Chanuc, F.; Fuseri, N.; Bonnafous, C.; Czerwinski, D.; Rajapaksa, A.; et al. Anti-KIR antibody enhancement of anti-lymphoma activity of natural killer cells as monotherapy and in combination with anti-CD20 antibodies. Blood 2014, 123, 678–686. [Google Scholar] [CrossRef] [PubMed]
- Vey, N.; Karlin, L.; Sadot-Lebouvier, S.; Broussais, F.; Berton-Rigaud, D.; Rey, J.; Charbonnier, A.; Marie, D.; André, P.; Paturel, C.; et al. A phase 1 study of lirilumab (antibody against killer immunoglobulin-like receptor antibody KIR2D; IPH2102) in patients with solid tumors and hematologic malignancies. Oncotarget 2018, 9, 17675–17688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benson, D.M., Jr.; Cohen, A.D.; Jagannath, S.; Munshi, N.C.; Spitzer, G.; Hofmeister, C.C.; Efebera, Y.A.; Andre, P.; Zerbib, R.; Caligiuri, M.A. A Phase I Trial of the Anti-KIR Antibody IPH2101 and Lenalidomide in Patients with Relapsed/Refractory Multiple Myeloma. Clin. Cancer Res. 2015, 21, 4055–4061. [Google Scholar] [CrossRef] [Green Version]
- van Hall, T.; André, P.; Horowitz, A.; Ruan, D.F.; Borst, L.; Zerbib, R.; Narni-Mancinelli, E.; van der Burg, S.H.; Vivier, E. Monalizumab: Inhibiting the novel immune checkpoint NKG2A. J. Immunother. Cancer 2019, 7, 263. [Google Scholar] [CrossRef] [PubMed]
- Galot, R.; Le Tourneau, C.; Saada-Bouzid, E.; Daste, A.; Even, C.; Debruyne, P.; Henry, S.; Zanetta, S.; Rutten, A.; Licitra, L.; et al. A phase II study of monalizumab in patients with recurrent/metastatic squamous cell carcinoma of the head and neck: The I1 cohort of the EORTC-HNCG-1559 UPSTREAM trial. Eur. J. Cancer 2021, 158, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Wolchok, J.D.; Kluger, H.; Callahan, M.K.; Postow, M.A.; Rizvi, N.A.; Lesokhin, A.M.; Segal, N.H.; Ariyan, C.E.; Gordon, R.A.; Reed, K.; et al. Nivolumab plus ipilimumab in advanced melanoma. New Engl. J. Med. 2013, 369, 122–133. [Google Scholar] [CrossRef] [Green Version]
- Benson, D.M., Jr.; Bakan, C.E.; Mishra, A.; Hofmeister, C.C.; Efebera, Y.; Becknell, B.; Baiocchi, R.A.; Zhang, J.; Yu, J.; Smith, M.K.; et al. The PD-1/PD-L1 axis modulates the natural killer cell versus multiple myeloma effect: A therapeutic target for CT-011, a novel monoclonal anti-PD-1 antibody. Blood 2010, 116, 2286–2294. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Kim, Y.M.; Jung, J.S.; Ji, W.; Lee, J.C.; Choi, C.M. Two-year efficacy of SNK01 plus pembrolizumab for non-small cell lung cancer: Expanded observations from a phase I/IIa randomized controlled trial. Thorac. Cancer 2022, 13, 2050–2056. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.; Luo, H.; Liang, S.; Chen, J.; Liu, A.; Niu, L.; Jiang, Y. Pembrolizumab plus allogeneic NK cells in advanced non-small cell lung cancer patients. J. Clin. Investig. 2020, 130, 2560–2569. [Google Scholar] [CrossRef] [PubMed]
- Boyerinas, B.; Jochems, C.; Fantini, M.; Heery, C.R.; Gulley, J.L.; Tsang, K.Y.; Schlom, J. Antibody-Dependent Cellular Cytotoxicity Activity of a Novel Anti-PD-L1 Antibody Avelumab (MSB0010718C) on Human Tumor Cells. Cancer Immunol. Res. 2015, 3, 1148–1157. [Google Scholar] [CrossRef] [Green Version]
- Rotte, A.; Sahasranaman, S.; Budha, N. Targeting TIGIT for Immunotherapy of Cancer: Update on Clinical Development. Biomedicines 2021, 9, 1277. [Google Scholar] [CrossRef] [PubMed]
- Tran, H.C.; Wan, Z.; Sheard, M.A.; Sun, J.; Jackson, J.R.; Malvar, J.; Xu, Y.; Wang, L.; Sposto, R.; Kim, E.S.; et al. TGFbetaR1 Blockade with Galunisertib (LY2157299) Enhances Anti-Neuroblastoma Activity of the Anti-GD2 Antibody Dinutuximab (ch14.18) with Natural Killer Cells. Clin. Cancer Res. 2017, 23, 804–813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giannelli, G.; Santoro, A.; Kelley, R.K.; Gane, E.; Paradis, V.; Cleverly, A.; Smith, C.; Estrem, S.T.; Man, M.; Wang, S.; et al. Biomarkers and overall survival in patients with advanced hepatocellular carcinoma treated with TGF-βRI inhibitor galunisertib. PLoS ONE 2020, 15, e0222259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Wei, J.; Liu, S.; Wang, J.; Han, X.; Qin, H.; Lang, J.; Cheng, K.; Li, Y.; Qi, Y.; et al. Inhibition of platelet function using liposomal nanoparticles blocks tumor metastasis. Theranostics 2017, 7, 1062–1071. [Google Scholar] [CrossRef]
- Liu, S.; Zhang, Y.; Zhao, X.; Wang, J.; Di, C.; Zhao, Y.; Ji, T.; Cheng, K.; Wang, Y.; Chen, L.; et al. Tumor-Specific Silencing of Tissue Factor Suppresses Metastasis and Prevents Cancer-Associated Hypercoagulability. Nano Lett. 2019, 19, 4721–4730. [Google Scholar] [CrossRef] [PubMed]
- Nahi, H.; Chrobok, M.; Meinke, S.; Gran, C.; Marquardt, N.; Afram, G.; Sutlu, T.; Gilljam, M.; Stellan, B.; Wagner, A.K.; et al. Autologous NK cells as consolidation therapy following stem cell transplantation in multiple myeloma. Cell Rep. Med. 2022, 3, 100508. [Google Scholar] [CrossRef]
- Dianat-Moghadam, H.; Mahari, A.; Heidarifard, M.; Parnianfard, N.; Pourmousavi-Kh, L.; Rahbarghazi, R.; Amoozgar, Z. NK cells-directed therapies target circulating tumor cells and metastasis. Cancer Lett. 2021, 497, 41–53. [Google Scholar] [CrossRef]
- Jung, E.K.; Chu, T.H.; Vo, M.C.; Nguyen, H.P.Q.; Lee, D.H.; Lee, J.K.; Lim, S.C.; Jung, S.H.; Yoon, T.M.; Yoon, M.S.; et al. Natural killer cells have a synergistic anti-tumor effect in combination with chemoradiotherapy against head and neck cancer. Cytotherapy 2022, 24, 905–915. [Google Scholar] [CrossRef]
- Ruscetti, M.; Leibold, J.; Bott, M.J.; Fennell, M.; Kulick, A.; Salgado, N.R.; Chen, C.C.; Ho, Y.J.; Sanchez-Rivera, F.J.; Feucht, J.; et al. NK cell-mediated cytotoxicity contributes to tumor control by a cytostatic drug combination. Science 2018, 362, 1416–1422. [Google Scholar] [CrossRef] [Green Version]
- Walle, T.; Kraske, J.A.; Liao, B.; Lenoir, B.; Timke, C.; von Bohlen Und Halbach, E.; Tran, F.; Griebel, P.; Albrecht, D.; Ahmed, A.; et al. Radiotherapy orchestrates natural killer cell dependent antitumor immune responses through CXCL8. Sci. Adv. 2022, 8, eabh4050. [Google Scholar] [CrossRef]
Figure 1.
Metastatic cascade and immunosurveillance. Primary tumor cells undergo epithelial-to-mesenchymal transition (EMT) within tumor-cell-extrinsic factors in nutritional, stromal, and immunological microenvironments endowed with distinct metastatic potential. The EMT trans-differentiated cells detach from the primary tumor and interact with and remodel the extracellular matrix (ECM) for intravasation, leaving the protection of the immunosuppressive tumor microenvironment. They reach the circulation, which make them more vulnerable to attacks by immune effector cells, particularly, NK cells, which can directly and indirectly interact with circulating tumor cells (CTCs) to control metastasis. However, they survive as polyclonal clusters of CTCs and cooperate with platelets and neutrophils, which shield them and alter the function of NK cells as a mode of escape from immunosurveillance until they encounter conditions that are permissive for extravasation and lodge at secondary sites. They can persist in a relatively silent state as disseminated tumor cells (DTCs) or micrometastatic tumor cells for a highly variable period of time, until they resume proliferation to establish a clinically detectable metastatic disease. Abbreviations: TAF, tumor-associated fibroblast; CTC, circulating tumor cell; CTL, cytotoxic T lymphocyte; DC, dendritic cell; DTC, disseminated tumor cell; ECM, extracellular matrix; EMT, epithelial-mesenchymal transition; MDSC, myeloid-derived suppressor cell; NK, natural killer cell; PLT, platelet; TAM, tumor-associated macrophage; Treg, regulatory T cell.
Figure 1.
Metastatic cascade and immunosurveillance. Primary tumor cells undergo epithelial-to-mesenchymal transition (EMT) within tumor-cell-extrinsic factors in nutritional, stromal, and immunological microenvironments endowed with distinct metastatic potential. The EMT trans-differentiated cells detach from the primary tumor and interact with and remodel the extracellular matrix (ECM) for intravasation, leaving the protection of the immunosuppressive tumor microenvironment. They reach the circulation, which make them more vulnerable to attacks by immune effector cells, particularly, NK cells, which can directly and indirectly interact with circulating tumor cells (CTCs) to control metastasis. However, they survive as polyclonal clusters of CTCs and cooperate with platelets and neutrophils, which shield them and alter the function of NK cells as a mode of escape from immunosurveillance until they encounter conditions that are permissive for extravasation and lodge at secondary sites. They can persist in a relatively silent state as disseminated tumor cells (DTCs) or micrometastatic tumor cells for a highly variable period of time, until they resume proliferation to establish a clinically detectable metastatic disease. Abbreviations: TAF, tumor-associated fibroblast; CTC, circulating tumor cell; CTL, cytotoxic T lymphocyte; DC, dendritic cell; DTC, disseminated tumor cell; ECM, extracellular matrix; EMT, epithelial-mesenchymal transition; MDSC, myeloid-derived suppressor cell; NK, natural killer cell; PLT, platelet; TAM, tumor-associated macrophage; Treg, regulatory T cell.

Figure 2.
Circulating tumor cells (CTCs) evade surveilling NK cells during metastasis. For successfully metastasis, tumor cells have to evade from NK-cell-mediated immunosurveillance. Tumor cells have several strategies involving tumor cell intrinsic features and tumor-driven extrinsic microenviroment factors to subvert their recognition and elimination by NK cells. Intrinsic features endow tumor cells with distinct metastatic potential owing to epigenetic and genetic alterations that include the downregulation of NK cell activating receptor ligands and/or the upregulation of NK cell inhibitory receptor ligands. In contrast, the tumor-cell-driven extrinsic microenvironment suppresses NK cell immunosurveillance through modulation of activating receptors and/or inhibitory receptors on NK cells. Platelets and neutrophils may favor the escape of CTCs from NK cell immunosurveillance, thereby supporting CTCs to establish metastasis. (Arrows indicate the change in expression levels of surface receptors).
Figure 2.
Circulating tumor cells (CTCs) evade surveilling NK cells during metastasis. For successfully metastasis, tumor cells have to evade from NK-cell-mediated immunosurveillance. Tumor cells have several strategies involving tumor cell intrinsic features and tumor-driven extrinsic microenviroment factors to subvert their recognition and elimination by NK cells. Intrinsic features endow tumor cells with distinct metastatic potential owing to epigenetic and genetic alterations that include the downregulation of NK cell activating receptor ligands and/or the upregulation of NK cell inhibitory receptor ligands. In contrast, the tumor-cell-driven extrinsic microenvironment suppresses NK cell immunosurveillance through modulation of activating receptors and/or inhibitory receptors on NK cells. Platelets and neutrophils may favor the escape of CTCs from NK cell immunosurveillance, thereby supporting CTCs to establish metastasis. (Arrows indicate the change in expression levels of surface receptors).

Figure 3.
Strategies to harness NK cells in control of metastasis. Each therapeutic strategy is evaluated in isolation or in combination, and even in combination with other strategies including standard treatment. Abbreviations: ADCC, antibody-dependent cellular cytotoxicity; mAbs, monoclonal antibodies. (Arrows indicate enhancement).
Figure 3.
Strategies to harness NK cells in control of metastasis. Each therapeutic strategy is evaluated in isolation or in combination, and even in combination with other strategies including standard treatment. Abbreviations: ADCC, antibody-dependent cellular cytotoxicity; mAbs, monoclonal antibodies. (Arrows indicate enhancement).


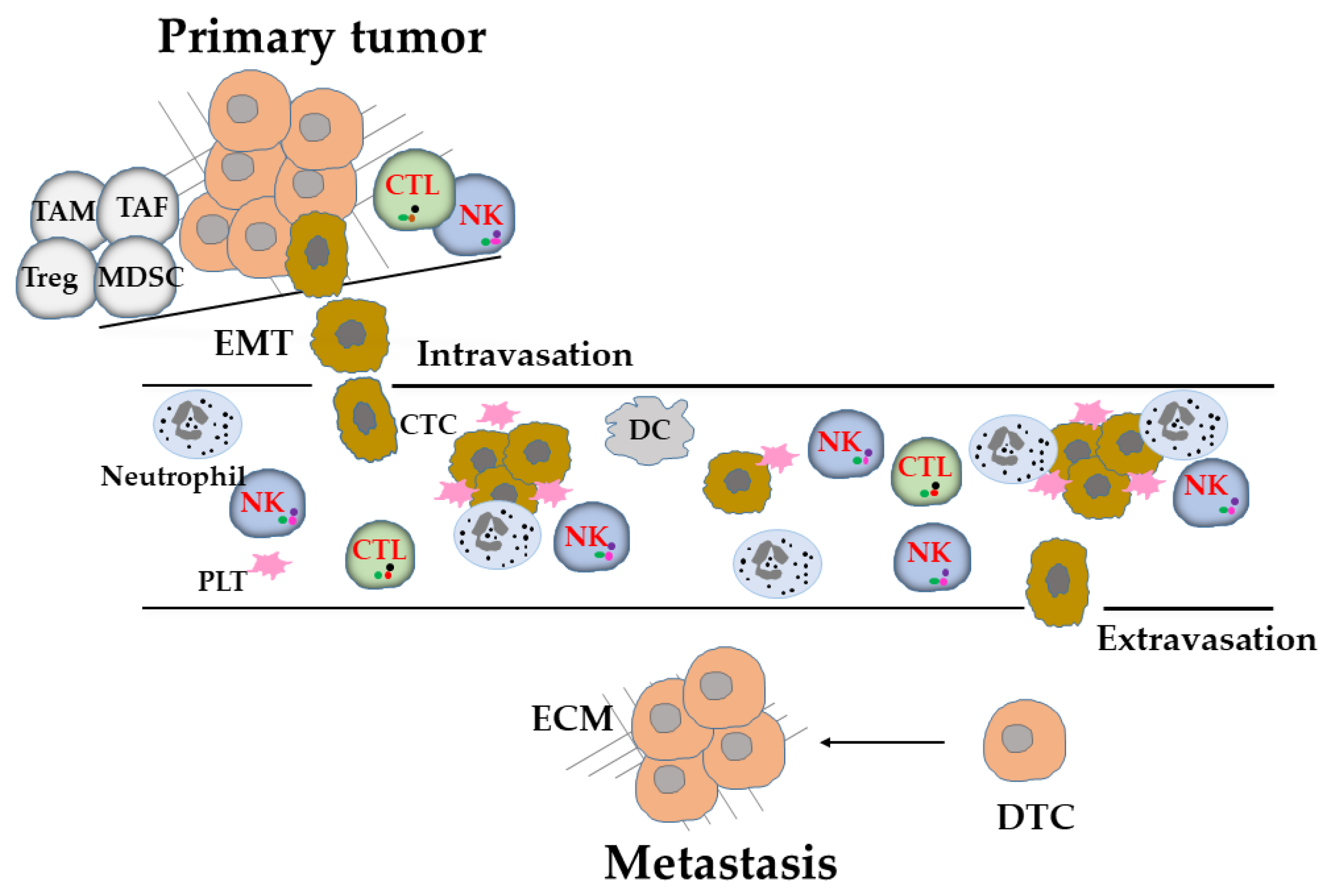{kind=link}
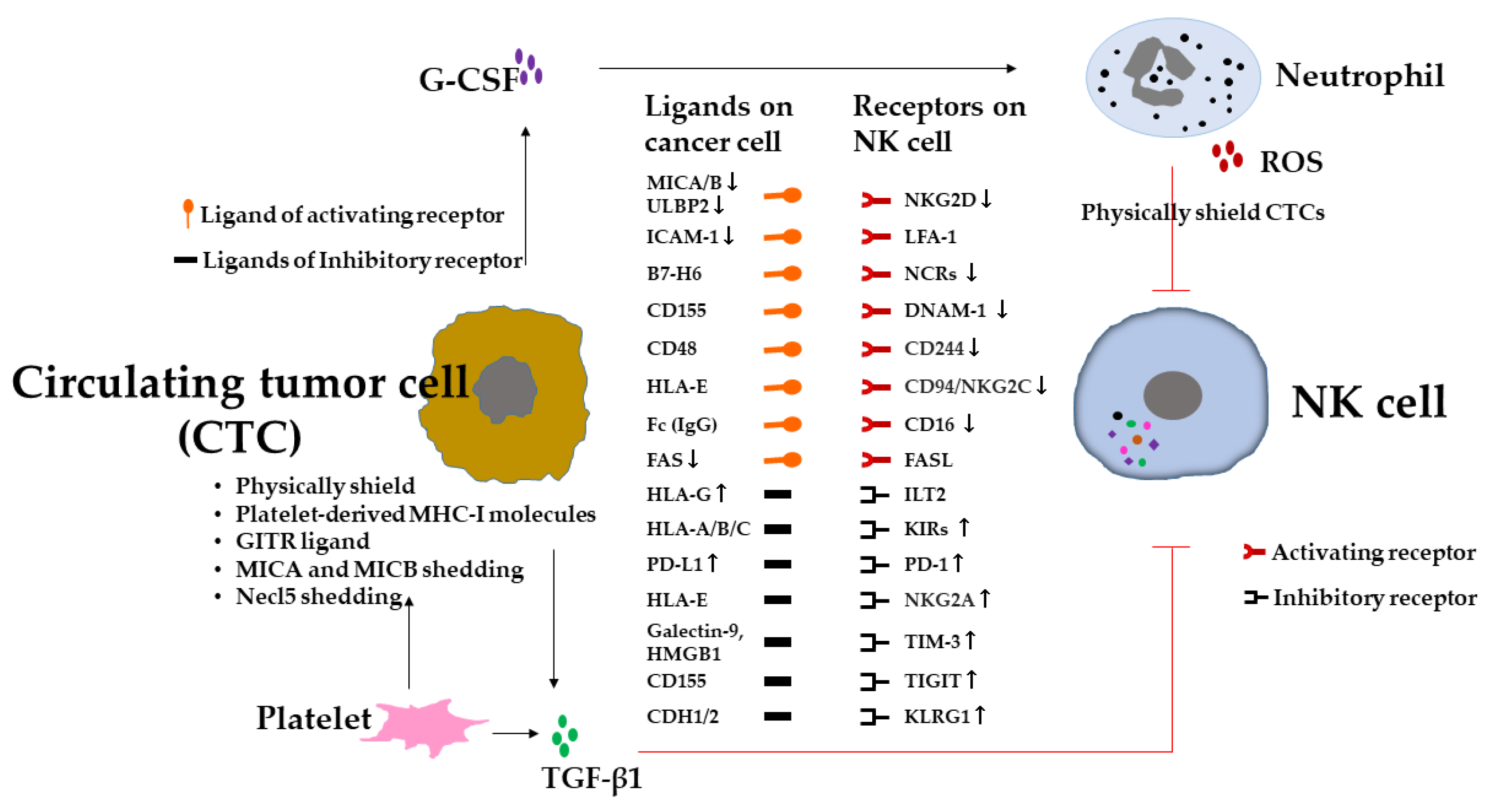{kind=link}
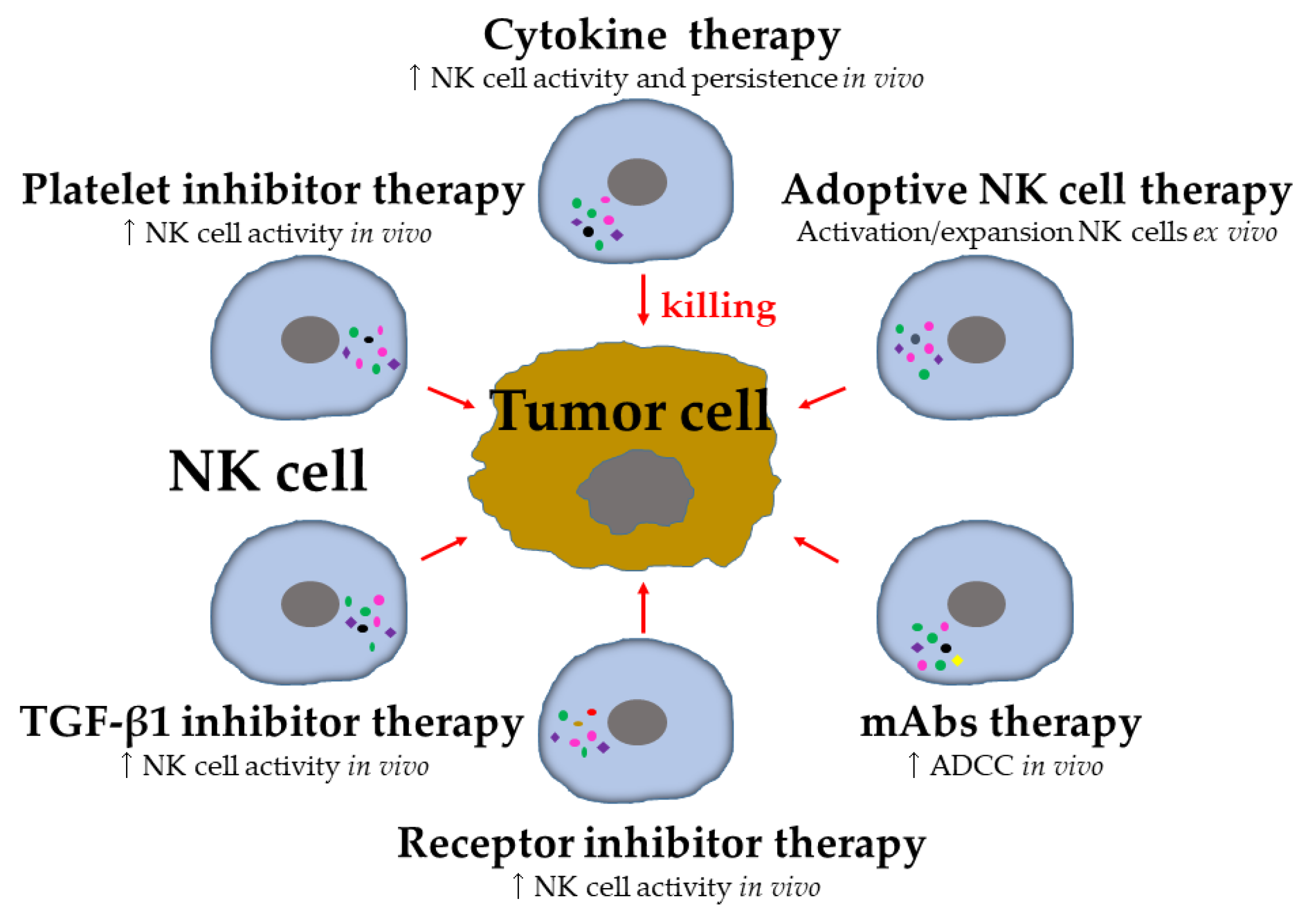{kind=link}
Table 1.
Selected clinical trials of NK-cell-based tumor therapy. Data source ClinicalTrials.gov (www.clinicaltrials.gov, accessed on 18 November 2022).
Table 1.
Selected clinical trials of NK-cell-based tumor therapy. Data source ClinicalTrials.gov (www.clinicaltrials.gov, accessed on 18 November 2022).
| Agent | Approach | Tumor Type | Phase | Trial Identifier |
|---|---|---|---|---|
| Cytokine-based therapy | ||||
| N-803 (IL-15 superagonist) | Monotherapy | Advanced-stage melanoma, NSCLC, RCC, HNSCC | I | NCT01946789 |
| NIZ985 (solube IL-15)/IL-15 receptor α heterodimer (hetIL-15) | In combination with spartalizumb (anti-PD-1 antibody) | Solid tumours, lymphoma or melanoma | I/Ib | NCT04261439 |
| Adoptive NK cell therapy | ||||
| Autologous NK cells | After treatment with bortezomib | Solid and haematological tumor | I | NCT00720785 |
| Allogeneic NK cells | In combination with trastuzumab or cetuximab | Her2+ or EGFR+ solid tumors | I | NCT03319459 |
| UCB NK cells | Before autologous HSCT | NHL | I/II | NCT03579927 |
| NK-92 cells expressing high-affinity variant of CD16 | Combined with N-803 and avelumab | Merkel cell carcinoma that has progressed after ICI | I | NCT03853317 |
| CIML NK cells | Combined with N-803 | R/R AML | II | NCT02782546 |
| iC9/CD19-CAR-CD28-zeta-2A-IL-15 NK cells | In combination with lymphodepleting chemotherapy | CD19+ R/R B cell lymphoma | I/II | NCT03056339 |
| Monoclonal antibody therapy | ||||
| Haploidentical NK cells | After treatment with anti-GD2 mAb | Neuroblastoma | I | NCT02650648 |
| Expanded autologous NK cells | After treatment with cetuximab | EGFR+ NPC or HNSCC | I/II | NCT02507154 |
| Expanded autologous NK cells | After treatment with trastuzumab | HER2+ breast or gastric cancer | I/II | NCT02030561 |
| Receptor inhibitor therapy | ||||
| Lirilumab (anti-KIR antibody) | In combination with elotuzumab or urelumab | Multiple Myeloma | I | NCT02252263 |
| IPH2101 (anti-KIR antibody) | In combination with lenalidomide | Multiple Myeloma | I | NCT01217203 |
| Monalizumb (anti-NKG2A antibody) | In combination with durvalumab | Advanced solid tumors | I/II | NCT02671435 |
| Cobolimab (TIM-3 Inhibitor) | In combination with dostarlimab | Resectable Stage III or Oligometastatic Stage IV Melanoma | II | NCT04139902 |
| Domvanalimab (anti-TIGIT antibody) | Monotherapy or in combination with zimberelimab | Advanced solid tumors | I | NCT03628677 |
| TGF-β1 inhibitor therapy | ||||
| Galunisertib | In combination with nivolumab | Advanced Refractory Solid Tumors | I/II | NCT02423343 |
Owing to the large number of trials in each category, example trials have been selected to illustrate the research and trials mentioned in this Review. Abbreviations: AML, acute myeloid leukemia; CIMI, cytokine-induced memory-like; HNSCC, head and neck squamous cell carcinoma; HSCT, hematopoietic stem cell transplantation; ICI, immune checkpoint inhibitor; NHL, non-Hodgkin lymphoma; NPC, nasopharyngeal carcinoma; NSCLC, non-small cell lung cancer; RCC, renal cell carcinoma; R/R, relapsed and/or refractory; UCB, umbilical cord blood.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Deng, X.; Terunuma, H. Harnessing NK Cells to Control Metastasis. Vaccines 2022, 10, 2018. https://doi.org/10.3390/vaccines10122018
AMA Style
Deng X, Terunuma H. Harnessing NK Cells to Control Metastasis. Vaccines. 2022; 10(12):2018. https://doi.org/10.3390/vaccines10122018
Chicago/Turabian StyleDeng, Xuewen, and Hiroshi Terunuma. 2022. "Harnessing NK Cells to Control Metastasis" Vaccines 10, no. 12: 2018. https://doi.org/10.3390/vaccines10122018
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.