
Evidence of Differences in Cellular Regulation of Wolbachia-Mediated Viral Inhibition between Alphaviruses and Flaviviruses
 , , and
, , and 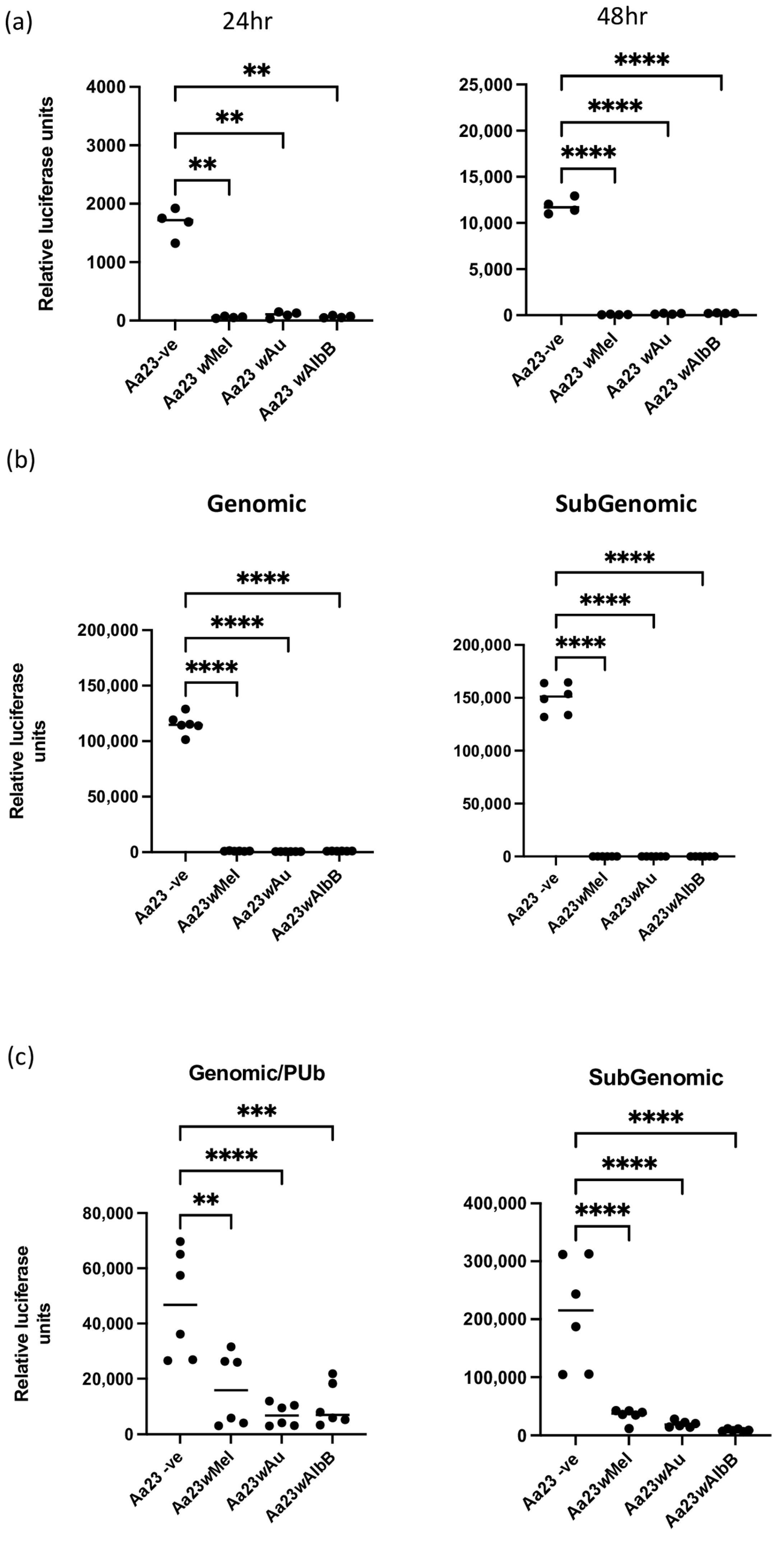{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mosquito Rearing and Cell Work
2.2. Viruses and Viral Clones
2.3. Infection and Transfection of Viruses and Viral Replicon/Trans-Replicases
2.4. qPCR and Luciferase Assay for Measurement of Virus
2.5. Statistical Analysis
3. Results
3.1. Wolbachia Blocks Early SFV Replication/Translation Events in Aedes albopictus Cells
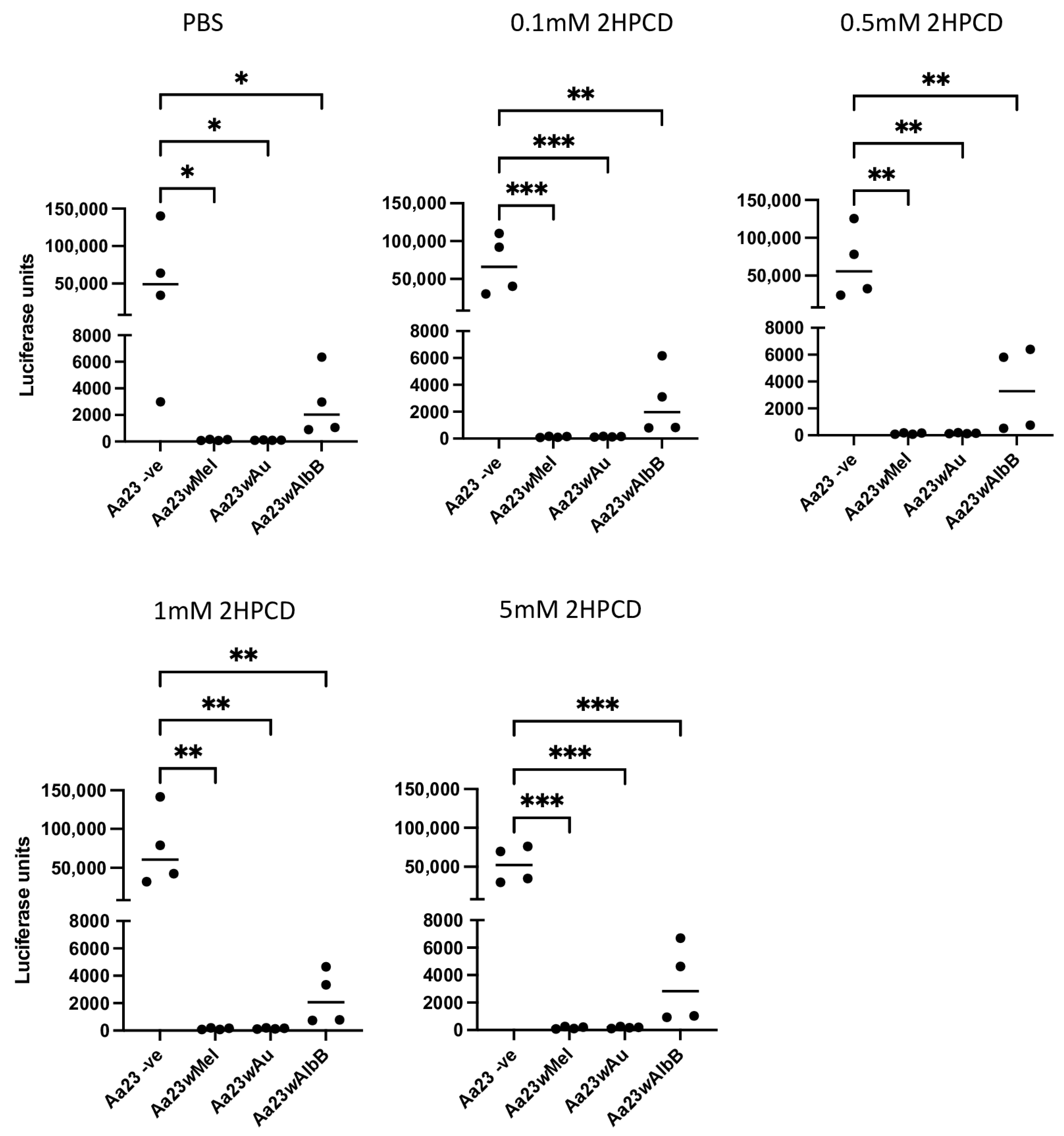
3.2. Wolbachia-Mediated Antiviral Activity against SFV Is Not Rescued by the Addition of 2HPCD
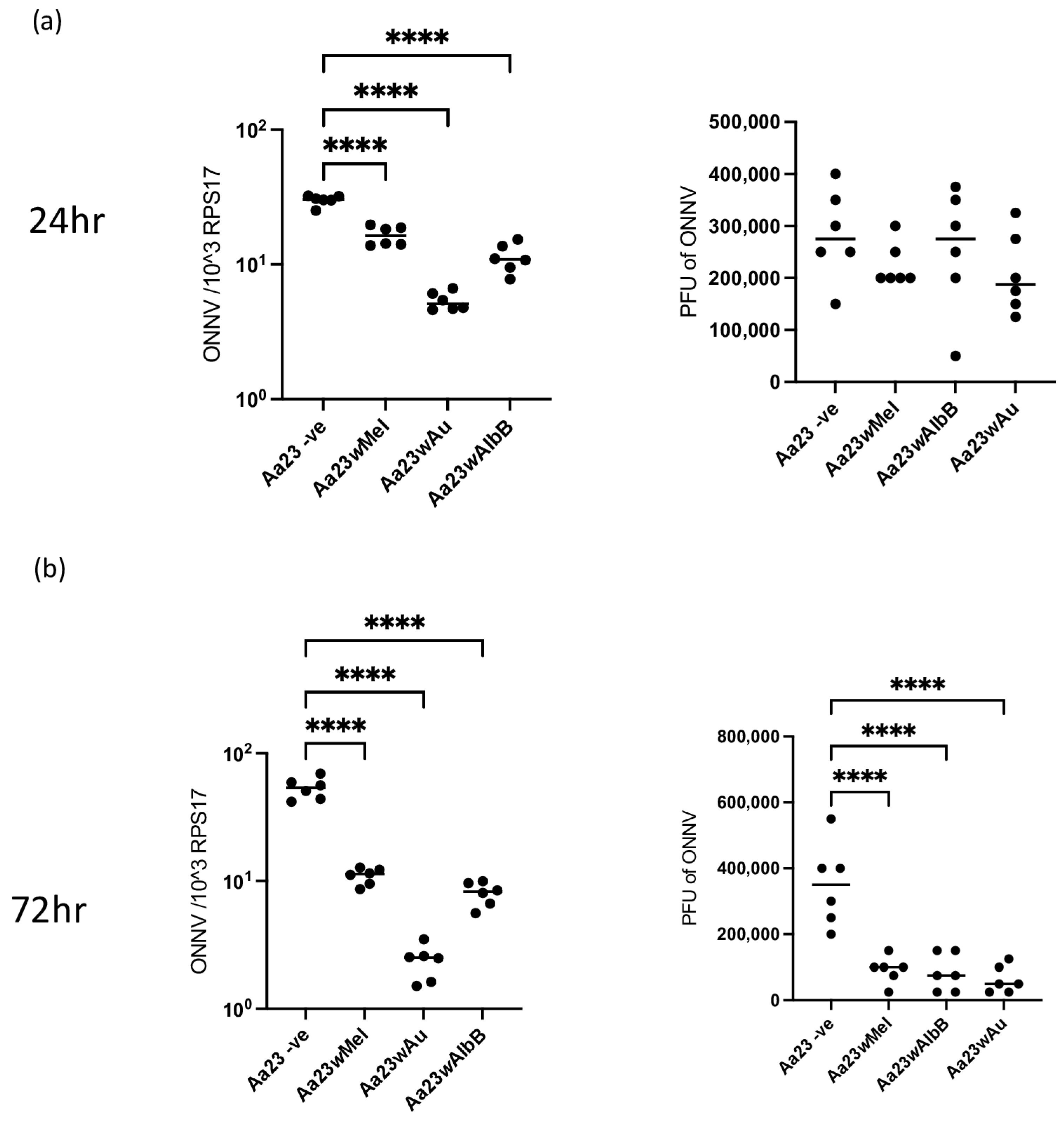
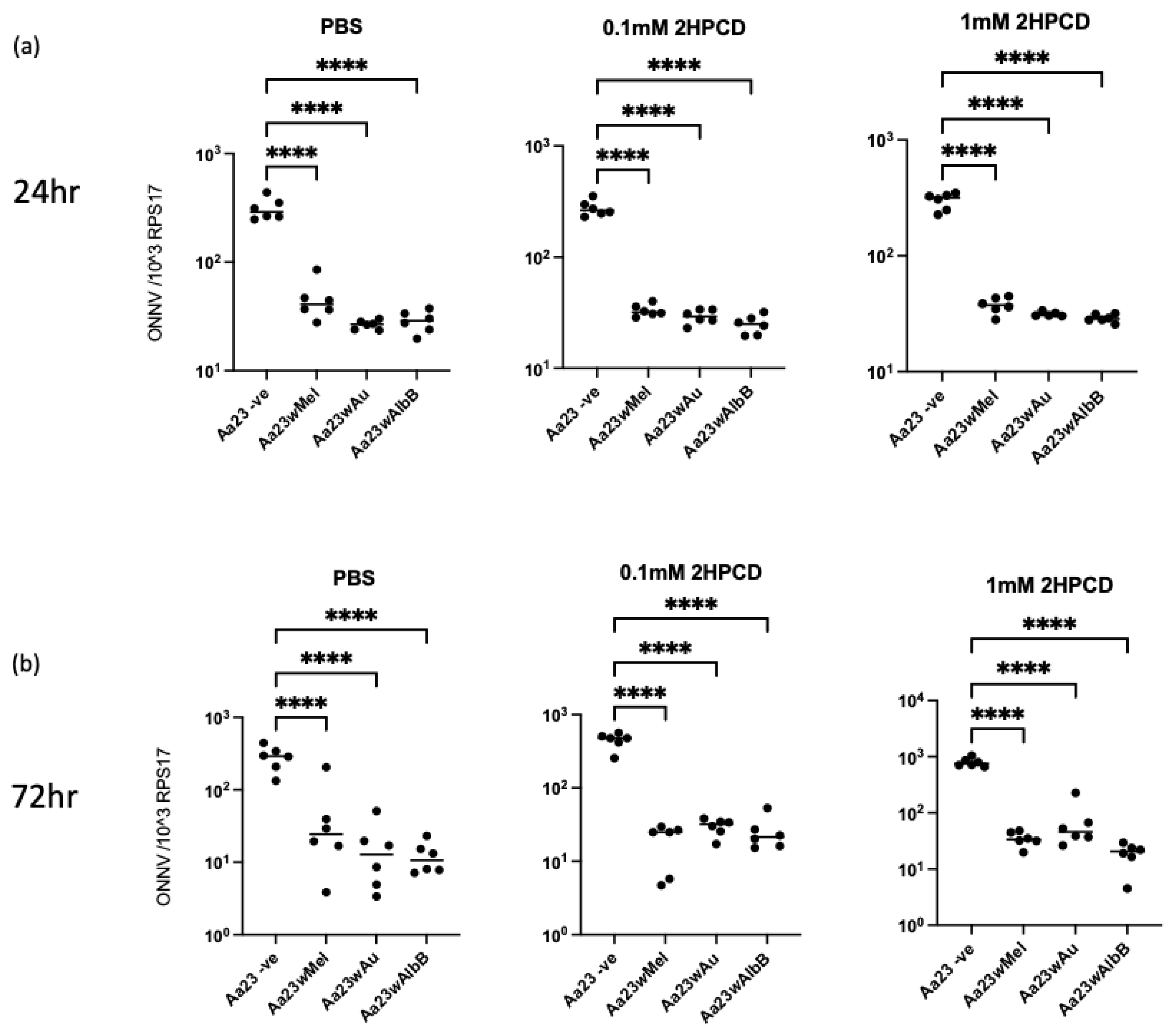
3.3. Wolbachia Strains Block ONNV Replication/Translation and Production of Infectious Particles in Aa23 Cells, and 2HPCD Does Not Rescue Viral Inhibition
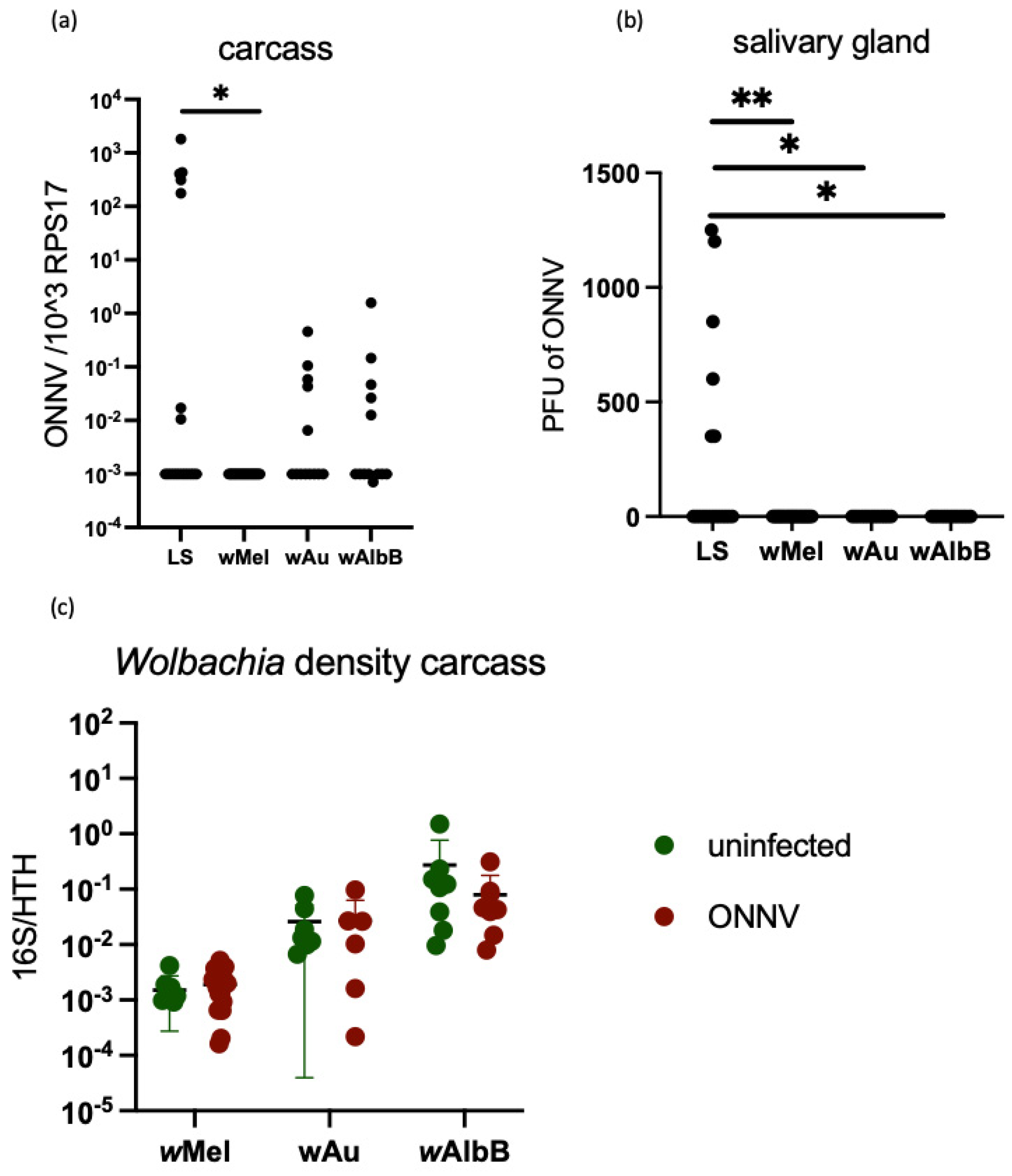
3.4. Stable Transinfection of wMel, wAlbB and wAu in Ae. aegypti Mosquitoes Limits ONNV Replication and Dissemination
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pereira Cabral, B.; da Graça Derengowski Fonseca, M.; Mota, F.B. Long term prevention and vector control of arboviral diseases: What does the future hold? Int. J. Infect. Dis. 2019, 89, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Nazni, W.A.; Hoffmann, A.A.; NoorAfizah, A.; Cheong, Y.L.; Mancini, M.V.; Golding, N.; Kamarul, G.M.R.; Arif, M.A.K.; Thohir, H.; NurSyamimi, H.; et al. Establishment of Wolbachia Strain wAlbB in Malaysian Populations of Aedes aegypti for Dengue Control. Curr. Biol. 2019, 29, 4241–4248.e5. [Google Scholar] [CrossRef] [PubMed]
- Utarini, A.; Indriani, C.; Ahmad, R.A.; Tantowijoyo, W.; Arguni, E.; Ansari, M.R.; Supriyati, E.; Wardana, D.S.; Meitika, Y.; Ernesia, I.; et al. Efficacy of Wolbachia-Infected Mosquito Deployments for the Control of Dengue. N. Engl. J. Med. 2021, 384, 2177–2186. [Google Scholar] [CrossRef]
- Pinto, S.B.; Riback, T.I.S.; Sylvestre, G.; Costa, G.; Peixoto, J.; Dias, F.B.S.; Tanamas, S.K.; Simmons, C.P.; Dufault, S.M.; Ryan, P.A.; et al. Effectiveness of Wolbachia-infected mosquito deployments in reducing the incidence of dengue and other Aedes-borne diseases in Niterói, Brazil: A quasi-experimental study. PLoS Negl. Trop. Dis. 2021, 15, e0009556. [Google Scholar] [CrossRef] [PubMed]
- Rainey, S.M.; Martinez, J.; McFarlane, M.; Juneja, P.; Sarkies, P.; Lulla, A.; Schnettler, E.; Varjak, M.; Merits, A.; Miska, E.A.; et al. Wolbachia Blocks Viral Genome Replication Early in Infection without a Transcriptional Response by the Endosymbiont or Host Small RNA Pathways. PLOS Pathog. 2016, 12, e1005536. [Google Scholar] [CrossRef]
- Durdevic, Z.; Hanna, K.; Gold, B.; Pollex, T.; Cherry, S.; Lyko, F.; Schaefer, M. Efficient RNA virus control in Drosophila requires the RNA methyltransferase Dnmt2. EMBO Rep. 2013, 14, 269–275. [Google Scholar] [CrossRef]
- Bhattacharya, T.; Newton, I.L.G.; Hardy, R.W. Wolbachia elevates host methyltransferase expression to block an RNA virus early during infection. PLoS Pathog. 2017, 13, e1006427. [Google Scholar] [CrossRef]
- Bhattacharya, T.; Yan, L.; Crawford, J.M.; Zaher, H.; Newton, I.L.G.; Hardy, R.W. Differential viral RNA methylation contributes to pathogen blocking in Wolbachia-colonized arthropods. PLoS Pathog. 2022, 18, e1010393. [Google Scholar] [CrossRef]
- Thomas, S.; Verma, J.; Woolfit, M.; O’Neill, S.L. Wolbachia-mediated virus blocking in mosquito cells is dependent on XRN1-mediated viral RNA degradation and influenced by viral replication rate. PLoS Pathog. 2018, 14, e1006879. [Google Scholar] [CrossRef]
- Caragata, E.P.; Rancès, E.; O’Neill, S.L.; McGraw, E.A. Competition for amino acids between Wolbachia and the mosquito host, Aedes aegypti. Microb. Ecol. 2014, 67, 205–218. [Google Scholar] [CrossRef]
- Lindsey, A.R.I.; Bhattacharya, T.; Hardy, R.W.; Newton, I.L.G. Wolbachia and Virus Alter the Host Transcriptome at the Interface of Nucleotide Metabolism Pathways. mBio 2021, 12, e03472-20. [Google Scholar] [CrossRef] [PubMed]
- Geoghegan, V.; Stainton, K.; Rainey, S.M.; Ant, T.H.; Dowle, A.A.; Larson, T.; Hester, S.; Charles, P.D.; Thomas, B.; Sinkins, S.P. Perturbed cholesterol and vesicular trafficking associated with dengue blocking in Wolbachia -infected Aedes aegypti cells. Nat. Commun. 2017, 8, 526. [Google Scholar] [CrossRef] [PubMed]
- Rainey, S.M.; Geoghegan, V.; Lefteri, D.A.; Ant, T.H.; Martinez, J.; McNamara, C.J.; Kamel, W.; de Laurent, Z.R.; Castello, A.; Sinkins, S.P. Differences in proteome perturbations caused by the Wolbachia strain wAu suggest multiple mechanisms of Wolbachia-mediated antiviral activity. Sci. Rep. 2023, 13, 11737. [Google Scholar] [CrossRef] [PubMed]
- Edwards, B.; Ghedin, E.; Voronin, D. Wolbachia interferes with Zika virus replication by hijacking cholesterol metabolism in mosquito cells. Microbiol. Spectr. 2023, 11, e02180-23. [Google Scholar] [CrossRef]
- Jose, J.; Taylor, A.B.; Kuhn, R.J. Spatial and Temporal Analysis of Alphavirus Replication and Assembly in Mammalian and Mosquito Cells. mBio 2017, 8, 10–1128. [Google Scholar] [CrossRef]
- Spuul, P.; Balistreri, G.; Hellström, K.; Golubtsov, A.V.; Jokitalo, E.; Ahola, T. Assembly of Alphavirus Replication Complexes from RNA and Protein Components in a Novel trans-Replication System in Mammalian Cells. J. Virol. 2011, 85, 4739–4751. [Google Scholar] [CrossRef]
- Strauss, J.H.; Strauss, E.G. The alphaviruses: Gene expression, replication, and evolution. Microbiol. Rev. 1994, 58, 491–562. [Google Scholar] [CrossRef]
- Froshauer, S.; Kartenbeck, J.; Helenius, A. Alphavirus RNA replicase is located on the cytoplasmic surface of endosomes and lysosomes. J. Cell Biol. 1988, 107, 2075–2086. [Google Scholar] [CrossRef]
- Takkinen, K.; Peränen, J.; Kääriäinen, L. Proteolytic processing of Semliki Forest virus-specific non-structural polyprotein. J. Gen. Virol. 1991, 72 Pt 7, 1627–1633. [Google Scholar] [CrossRef]
- Lefteri, D.A.; Bryden, S.R.; Pingen, M.; Terry, S.; McCafferty, A.; Beswick, E.F.; Georgiev, G.; Van der Laan, M.; Mastrullo, V.; Campagnolo, P.; et al. Mosquito saliva enhances virus infection through sialokinin-dependent vascular leakage. Proc. Natl. Acad. Sci. USA 2022, 119, e2114309119. [Google Scholar] [CrossRef]
- Pohjala, L.; Utt, A.; Varjak, M.; Lulla, A.; Merits, A.; Ahola, T.; Tammela, P. Inhibitors of alphavirus entry and replication identified with a stable Chikungunya replicon cell line and virus-based assays. PLoS ONE 2011, 6, e28923. [Google Scholar] [CrossRef] [PubMed]
- Utt, A.; Quirin, T.; Saul, S.; Hellström, K.; Ahola, T.; Merits, A. Versatile Trans-Replication Systems for Chikungunya Virus Allow Functional Analysis and Tagging of Every Replicase Protein. PLoS ONE 2016, 11, e0151616. [Google Scholar] [CrossRef]
- Martinez, J.; Sinkins, S.P. Rapid Evolution of Wolbachia Genomes in Mosquito Cell Culture. 2023. Available online: https://www.biorxiv.org/content/10.1101/2023.09.20.558649v1 (accessed on 6 November 2023).
- Blagrove, M.S.C.; Arias-Goeta, C.; Failloux, A.-B.; Sinkins, S.P. Wolbachia strain wMel induces cytoplasmic incompatibility and blocks dengue transmission in Aedes albopictus. Proc. Natl. Acad. Sci. USA 2012, 109, 255–260. [Google Scholar] [CrossRef]
- Ant, T.H.; Herd, C.S.; Geoghegan, V.; Hoffmann, A.A.; Sinkins, S.P. The Wolbachia strain wAu provides highly efficient virus transmission blocking in Aedes aegypti. PLoS Pathog. 2018, 14, e1006815. [Google Scholar] [CrossRef] [PubMed]
- Varjak, M.; Maringer, K.; Watson, M.; Sreenu, V.B.; Fredericks, A.C.; Pondeville, E.; Donald, C.L.; Sterk, J.; Kean, J.; Vazeille, M.; et al. Aedes aegypti Piwi4 Is a Noncanonical PIWI Protein Involved in Antiviral Responses. mSphere 2017, 2, e00144-17. [Google Scholar] [CrossRef] [PubMed]
- Siu, R.W.C.; Fragkoudis, R.; Simmonds, P.; Donald, C.L.; Chase-Topping, M.E.; Barry, G.; Attarzadeh-Yazdi, G.; Rodriguez-Andres, J.; Nash, A.A.; Merits, A.; et al. Antiviral RNA interference responses induced by Semliki Forest virus infection of mosquito cells: Characterization, origin, and frequency-dependent functions of virus-derived small interfering RNAs. J. Virol. 2011, 85, 2907–2917. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.A.E.; Gross, T.L.; Myles, K.M.; Adelman, Z.N. Validation of novel promoter sequences derived from two endogenous ubiquitin genes in transgenic Aedes aegypti. Insect Mol. Biol. 2010, 19, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Phalen, T.; Kielian, M. Cholesterol is required for infection by Semliki Forest virus. J. Cell Biol. 1991, 112, 615–623. [Google Scholar] [CrossRef]
- Ng, C.G.; Coppens, I.; Govindarajan, D.; Pisciotta, J.; Shulaev, V.; Griffin, D.E. Effect of host cell lipid metabolism on alphavirus replication, virion morphogenesis, and infectivity. Proc. Natl. Acad. Sci. USA 2008, 105, 16326–16331. [Google Scholar] [CrossRef]
- Osuna-Ramos, J.F.; Reyes-Ruiz, J.M.; del Ángel, R.M. The Role of Host Cholesterol During Flavivirus Infection. Front. Cell. Infect. Microbiol. 2018, 8, 388. [Google Scholar] [CrossRef]
- Blanc, M.; Hsieh, W.Y.; Robertson, K.A.; Watterson, S.; Shui, G.; Lacaze, P.; Khondoker, M.; Dickinson, P.; Sing, G.; Rodríguez-Martín, S.; et al. Host Defense against Viral Infection Involves Interferon Mediated Down-Regulation of Sterol Biosynthesis. PLoS Biol. 2011, 9, e1000598. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Guo, L.; Li, K.; Xiao, X.; Zhu, W.; Zheng, X.; Hu, J.; Zhang, H.; Cai, J.; Yu, Y.; et al. Inhibition of the mevalonate pathway enhances cancer cell oncolysis mediated by M1 virus. Nat. Commun. 2018, 9, 1524. [Google Scholar] [CrossRef] [PubMed]
- Vial, T.; Marti, G.; Missé, D.; Pompon, J. Lipid Interactions between Flaviviruses and Mosquito Vectors. Front. Physiol. 2021, 12, 763195. [Google Scholar] [CrossRef] [PubMed]
- Urade, R.; Hayashi, Y.; Kito, M. Endosomes differ from plasma membranes in the phospholipid molecular species composition. Biochim. Biophys. Acta 1988, 946, 151–163. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rainey, S.M.; Lefteri, D.A.; Darby, C.; Kohl, A.; Merits, A.; Sinkins, S.P. Evidence of Differences in Cellular Regulation of Wolbachia-Mediated Viral Inhibition between Alphaviruses and Flaviviruses. Viruses 2024, 16, 115. https://doi.org/10.3390/v16010115
Rainey SM, Lefteri DA, Darby C, Kohl A, Merits A, Sinkins SP. Evidence of Differences in Cellular Regulation of Wolbachia-Mediated Viral Inhibition between Alphaviruses and Flaviviruses. Viruses. 2024; 16(1):115. https://doi.org/10.3390/v16010115
Chicago/Turabian StyleRainey, Stephanie M., Daniella A. Lefteri, Christie Darby, Alain Kohl, Andres Merits, and Steven P. Sinkins. 2024. "Evidence of Differences in Cellular Regulation of Wolbachia-Mediated Viral Inhibition between Alphaviruses and Flaviviruses" Viruses 16, no. 1: 115. https://doi.org/10.3390/v16010115