
Molecular Characterization of a Novel Enamovirus Infecting Raspberry
 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. RNA Isolation and cDNA Synthesis
2.3. RACE (Rapid Amplification of cDNA Ends)
2.4. RT-PCR and Sanger Sequencing
2.5. RT-qPCR
2.6. High-Throughput Sequencing and Viral Genome Sequencing
2.7. Aphid Cultures
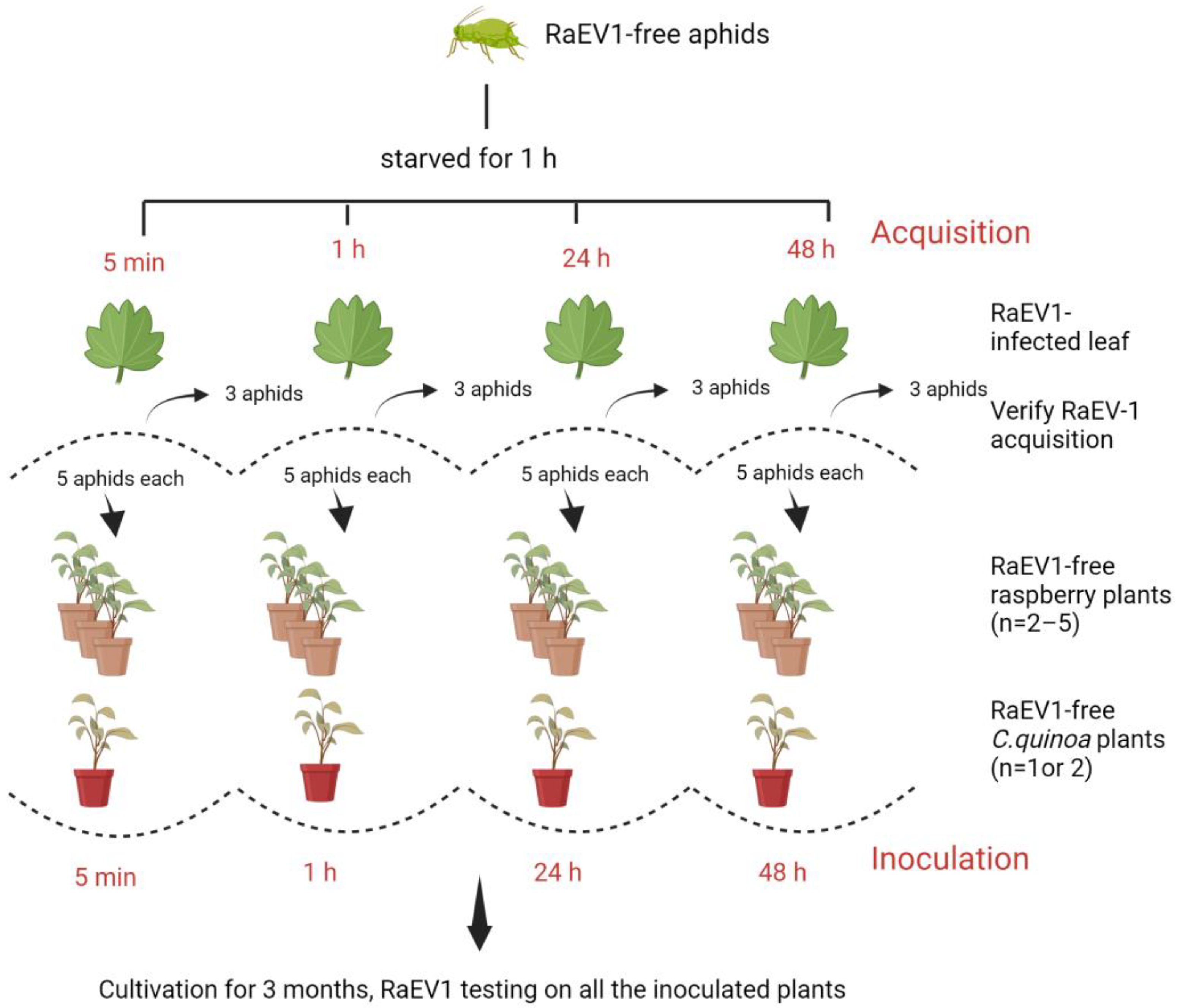
2.8. Virus Transmission Experiment
2.9. Data Analysis
2.10. Phylogenetic Analyses
3. Results and Discussion
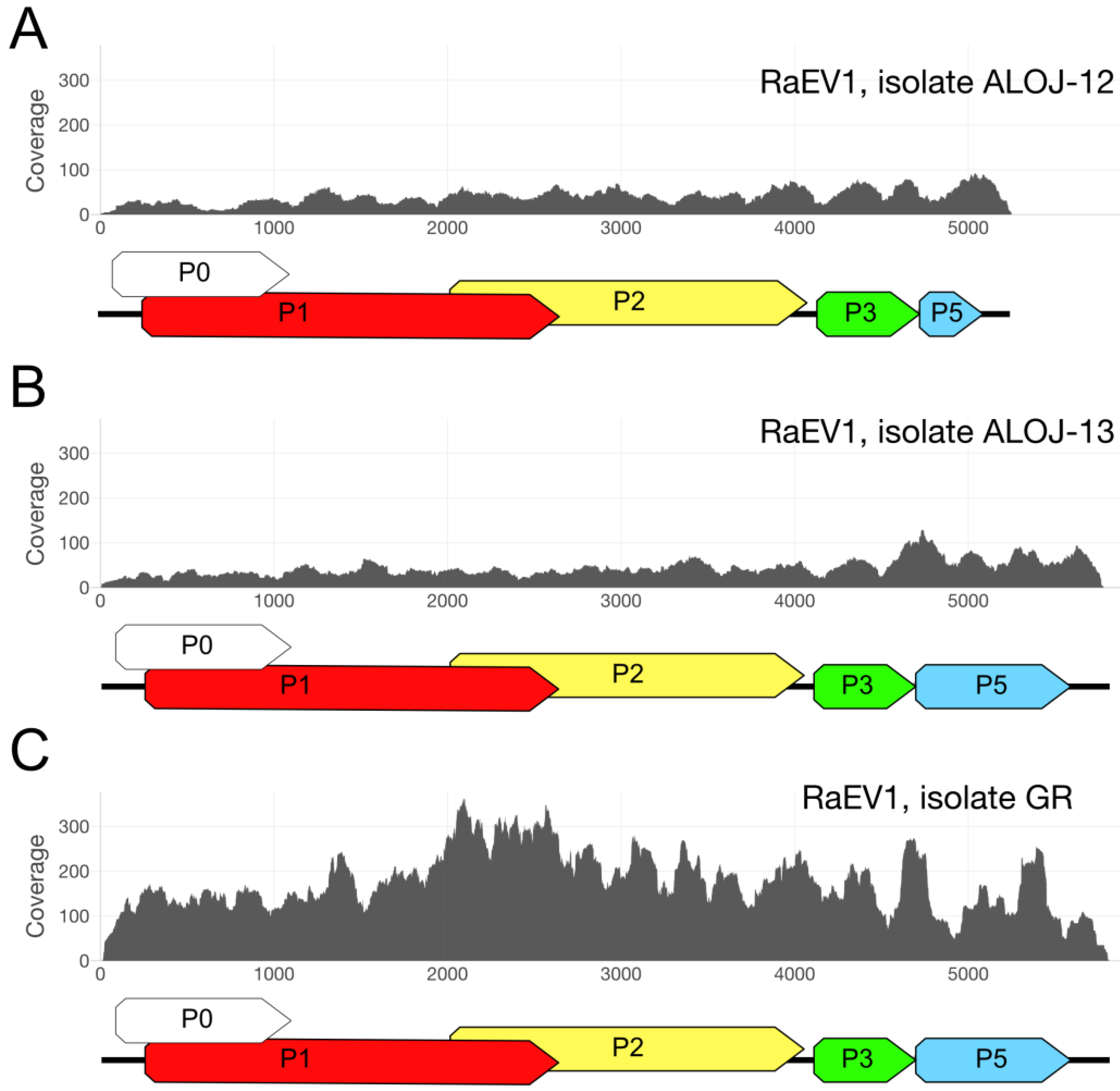
3.1. High-Throughput Sequencing
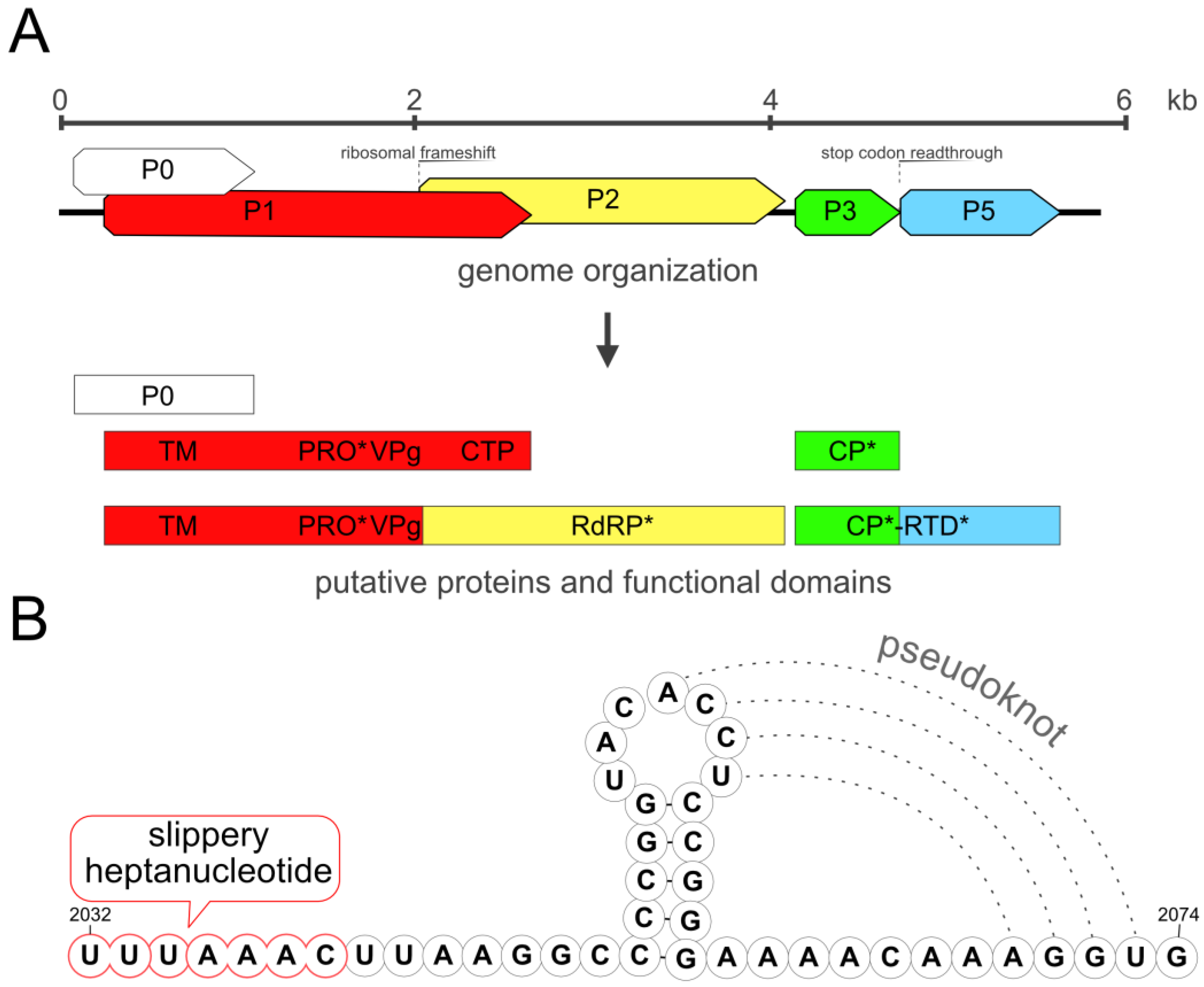
3.2. Genome
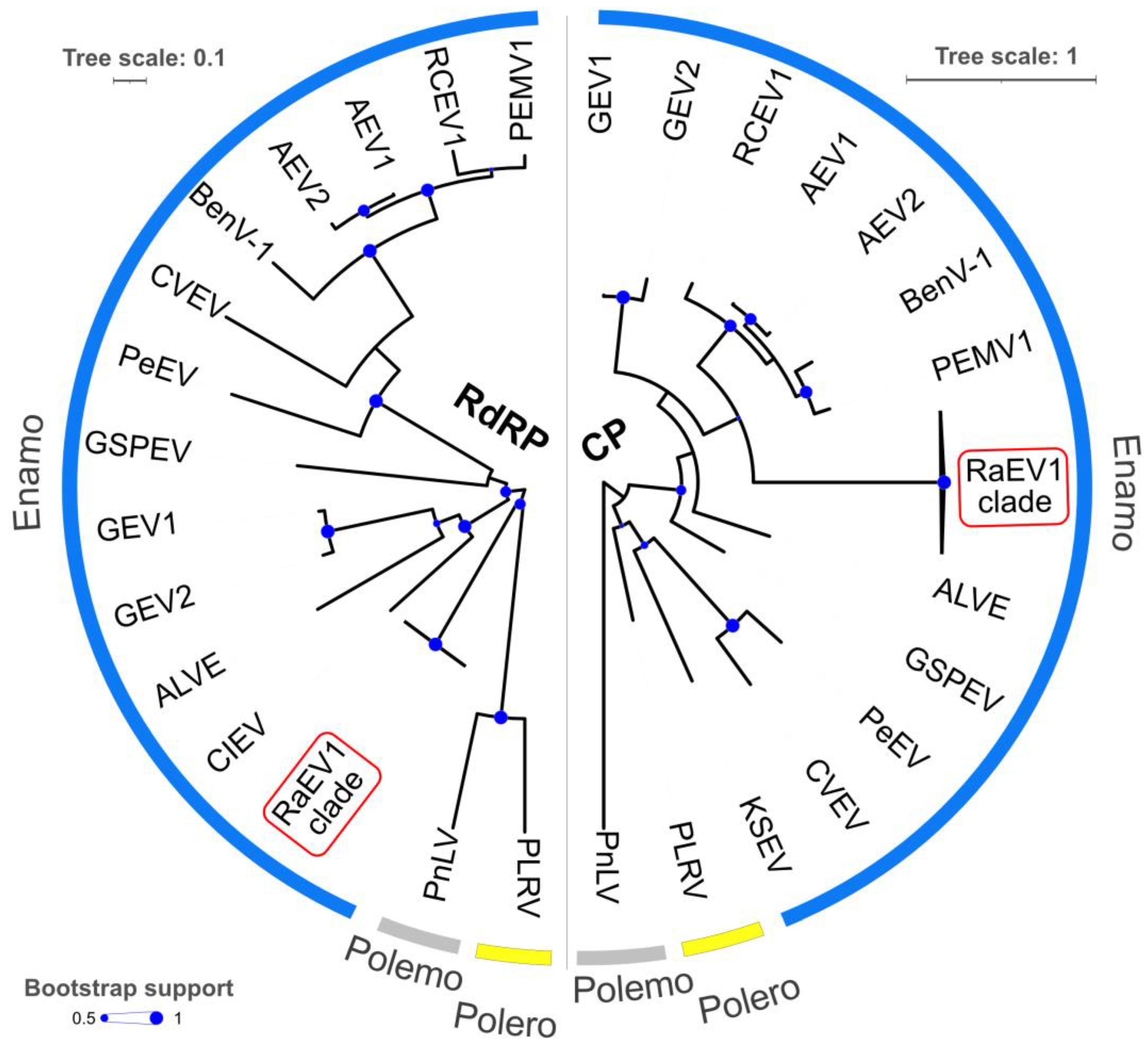
3.3. Phylogeny
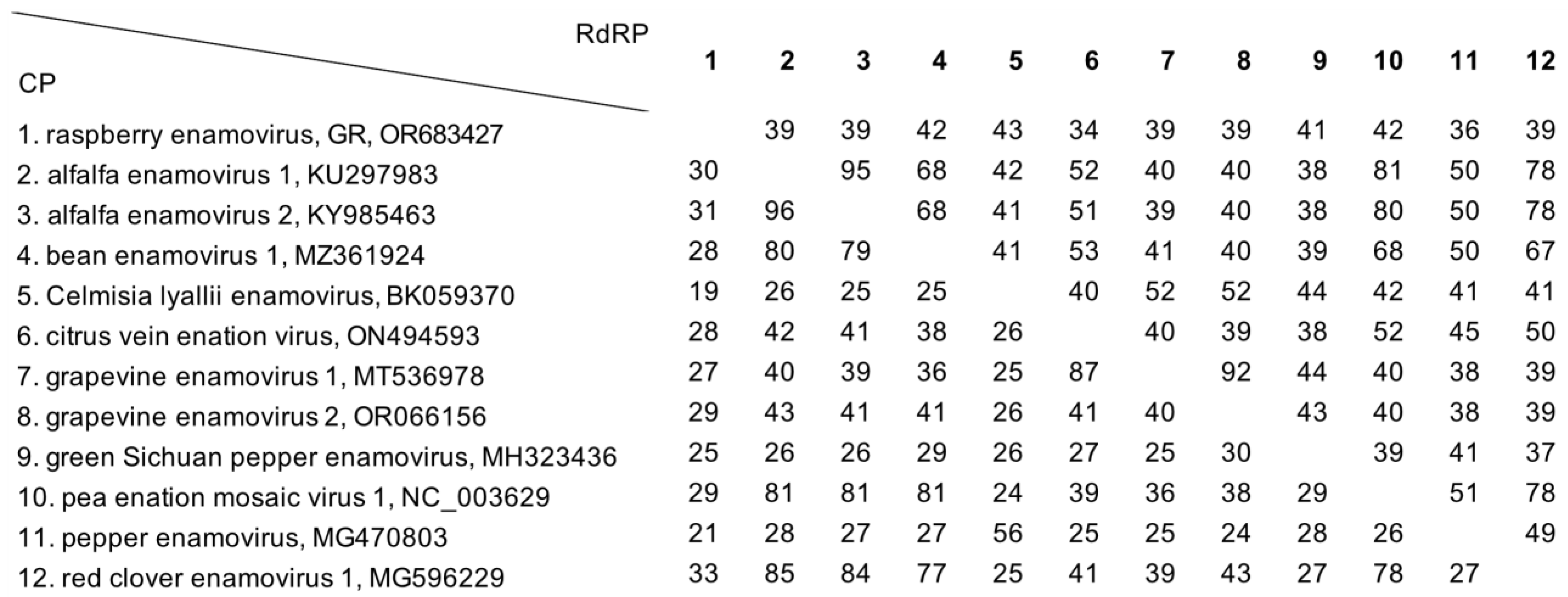
3.4. Shared Protein Identity with Other Enamoviruses
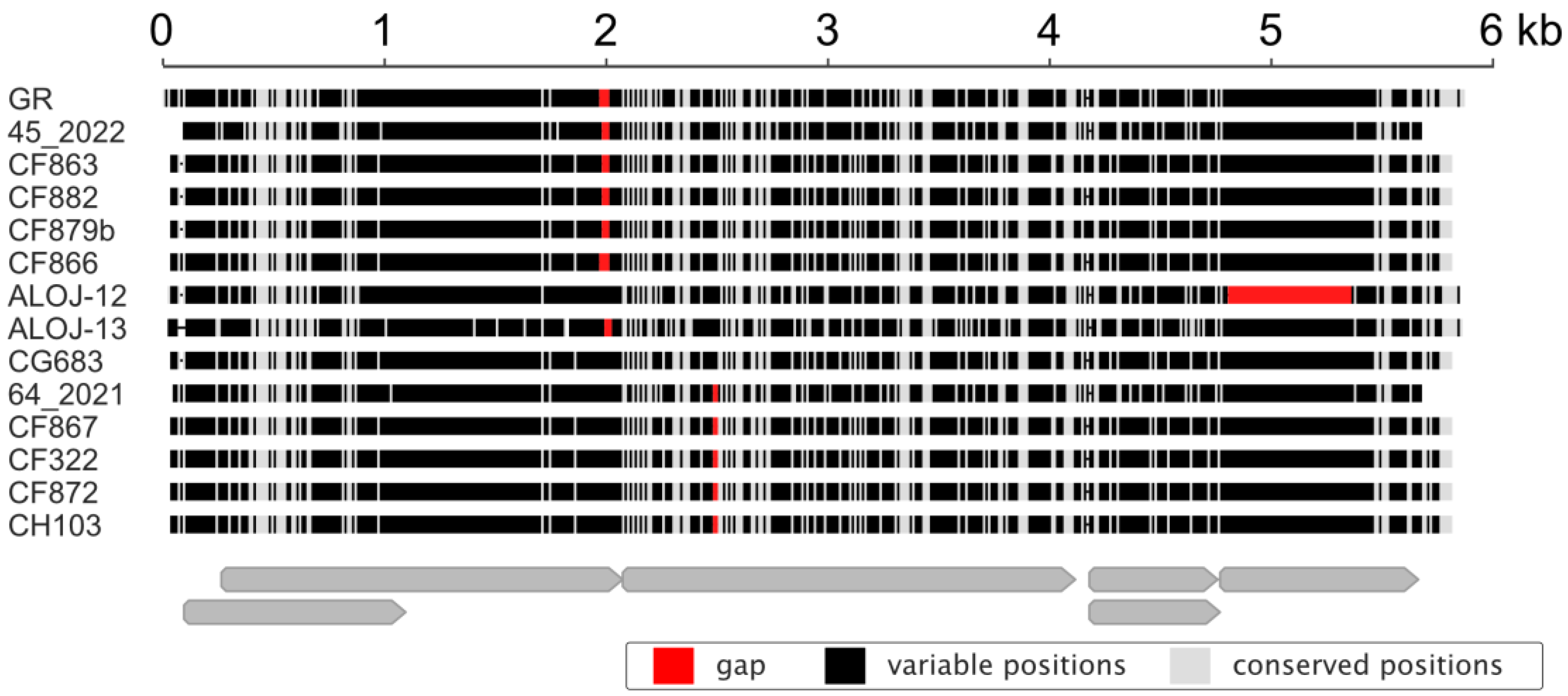
3.5. Isolate Diversity
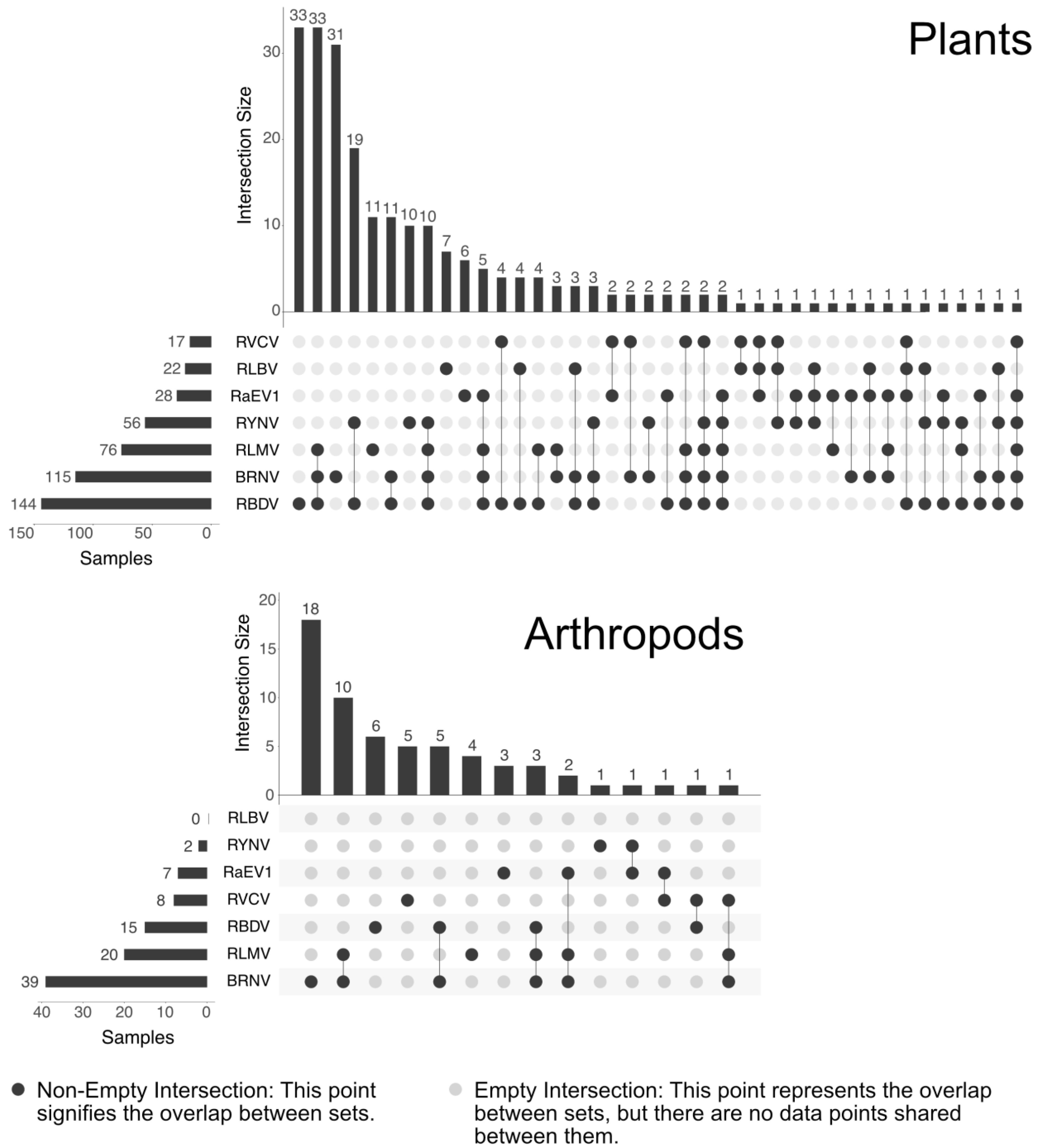
3.6. Prevalence
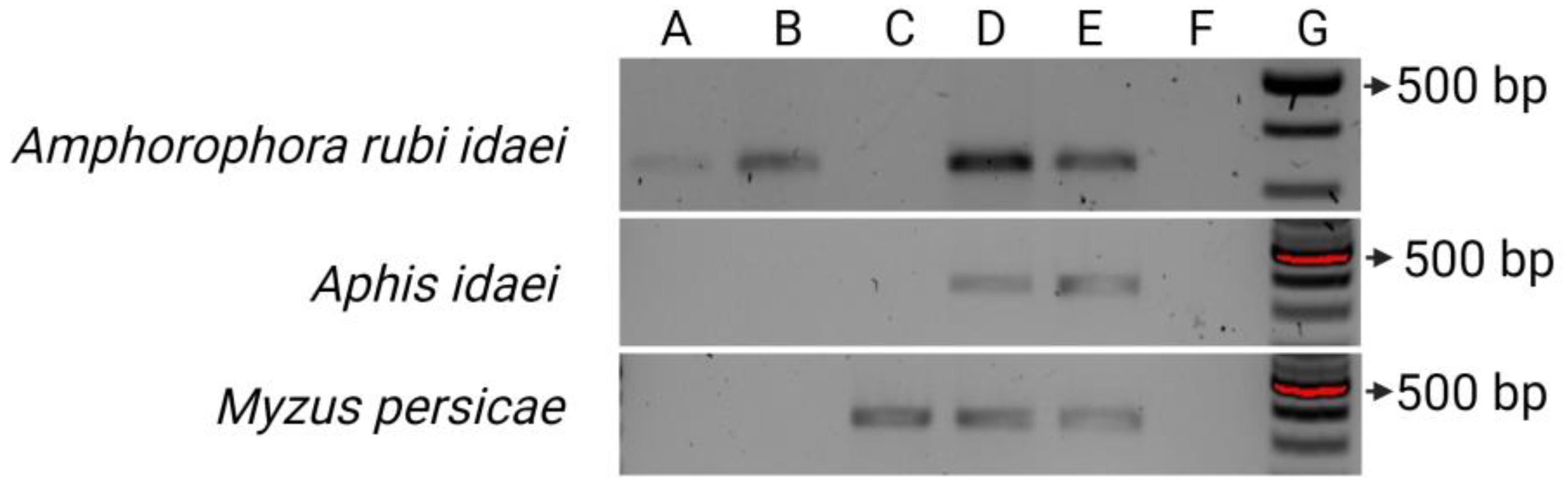
3.7. Transmission Assays
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- EFSA Panel on Plant Health (PLH); Bragard, C.; Dehnen-Schmutz, K.; Gonthier, P.; Jacques, M.; Jaques Miret, J.A.; Justesen, A.F.; MacLeod, A.; Magnusson, C.S.; Milonas, P.; et al. List of non-EU Viruses and Viroids of Cydonia Mill., Fragaria L., Malus Mill., Prunus L., Pyrus L., Ribes L., Rubus L. and Vitis L. EFSA J. 2019, 17, e05501. [Google Scholar] [CrossRef]
- Gazel, M.; Roumi, V.; Ördek, K.; Maclot, F.; Massart, S.; Çağlayan, K. Identification and Molecular Characterization of a Novel Foveavirus from Rubus Spp. in Turkey. Virus Res. 2020, 286, 198078. [Google Scholar] [CrossRef]
- Villamor, D.E.V.; Keller, K.E.; Martin, R.R.; Tzanetakis, I.E. Comparison of High Throughput Sequencing to Standard Protocols for Virus Detection in Berry Crops. Plant Dis. 2022, 106, 518–525. [Google Scholar] [CrossRef]
- Martin, R.R.; MacFarlane, S.; Sabanadzovic, S.; Quito, D.; Poudel, B.; Tzanetakis, I.E. Viruses and Virus Diseases of Rubus. Plant Dis. 2013, 97, 168–182. [Google Scholar] [CrossRef]
- Quito-Avila, D.F.; Lightle, D.; Martin, R.R. Effect of Raspberry Bushy Dwarf Virus, Raspberry Leaf Mottle Virus, and Raspberry Latent Virus on Plant Growth and Fruit Crumbliness in ‘Meeker’ Red Raspberry. Plant Dis. 2014, 98, 176–183. [Google Scholar] [CrossRef]
- Tan, J.L.; Trandem, N.; Fránová, J.; Hamborg, Z.; Blystad, D.-R.; Zemek, R. Known and Potential Invertebrate Vectors of Raspberry Viruses. Viruses 2022, 14, 571. [Google Scholar] [CrossRef]
- Gray, S.; Gildow, F.E. Luteovirus-Aphid Interactions. Annu. Rev. Phytopathol. 2003, 41, 539–566. [Google Scholar] [CrossRef]
- Domier, L.L.; D’Arcy, C.J. Luteoviruses. In Encyclopedia of Virology; Elsevier: Amsterdam, The Netherlands, 2008; pp. 231–238. ISBN 978-0-12-374410-4. [Google Scholar]
- Sõmera, M.; Fargette, D.; Hébrard, E.; Sarmiento, C. ICTV Report Consortium ICTV Virus Taxonomy Profile: Solemoviridae 2021. J. Gen. Virol. 2021, 102, 001707. [Google Scholar] [CrossRef]
- Brault, V.; Van Den Heuvel, J.F.; Verbeek, M.; Ziegler-Graff, V.; Reutenauer, A.; Herrbach, E.; Garaud, J.C.; Guilley, H.; Richards, K.; Jonard, G. Aphid Transmission of Beet Western Yellows Luteovirus Requires the Minor Capsid Read-through Protein P74. EMBO J. 1995, 14, 650–659. [Google Scholar] [CrossRef]
- Van Den Heuvel, J.F.; Bruyère, A.; Hogenhout, S.A.; Ziegler-Graff, V.; Brault, V.; Verbeek, M.; Van Der Wilk, F.; Richards, K. The N-Terminal Region of the Luteovirus Readthrough Domain Determines Virus Binding to Buchnera GroEL and Is Essential for Virus Persistence in the Aphid. J. Virol. 1997, 71, 7258–7265. [Google Scholar] [CrossRef]
- Gray, S.M.; Banerjee, N. Mechanisms of Arthropod Transmission of Plant and Animal Viruses. Microbiol. Mol. Biol. Rev. 1999, 63, 128–148. [Google Scholar] [CrossRef]
- Gray, S.; Cilia, M.; Ghanim, M. Circulative, “Nonpropagative” Virus Transmission. In Advances in Virus Research; Elsevier: Amsterdam, The Netherlands, 2014; Volume 89, pp. 141–199. ISBN 978-0-12-800172-1. [Google Scholar]
- Xu, Y.; Ju, H.-J.; DeBlasio, S.; Carino, E.J.; Johnson, R.; MacCoss, M.J.; Heck, M.; Miller, W.A.; Gray, S.M. A Stem-Loop Structure in Potato Leafroll Virus Open Reading Frame 5 (ORF5) Is Essential for Readthrough Translation of the Coat Protein ORF Stop Codon 700 Bases Upstream. J. Virol. 2018, 92, e01544-17. [Google Scholar] [CrossRef]
- Miller, W.A. Luteovirus (Luteoviridae). In Encyclopedia of Virology; Elsevier: Amsterdam, The Netherlands, 1999; pp. 901–908. ISBN 978-0-12-227030-7. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA Primers for Amplification of Mitochondrial Cytochrome c Oxidase Subunit I from Diverse Metazoan Invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Morris, T.J. Isolation and Analysis of Double-Stranded RNA from Virus-Infected Plant and Fungal Tissue. Phytopathology 1979, 69, 854. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Anisimova, M.; Gascuel, O. Approximate Likelihood-Ratio Test for Branches: A Fast, Accurate, and Powerful Alternative. Syst. Biol. 2006, 55, 539–552. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v3: An Online Tool for the Display and Annotation of Phylogenetic and Other Trees. Nucleic Acids Res. 2016, 44, W242–W245. [Google Scholar] [CrossRef]
- Peter, K.A.; Liang, D.; Palukaitis, P.; Gray, S.M. Small Deletions in the Potato Leafroll Virus Readthrough Protein Affect Particle Morphology, Aphid Transmission, Virus Movement and Accumulation. J. Gen. Virol. 2008, 89, 2037–2045. [Google Scholar] [CrossRef]
- Boissinot, S.; Erdinger, M.; Monsion, B.; Ziegler-Graff, V.; Brault, V. Both Structural and Non-Structural Forms of the Readthrough Protein of Cucurbit Aphid-Borne Yellows Virus Are Essential for Efficient Systemic Infection of Plants. PLoS ONE 2014, 9, e93448. [Google Scholar] [CrossRef]
- Linck, H.; Reineke, A. Rubus Stunt: A Review of an Important Phytoplasma Disease in Rubus spp. J. Plant Dis. Prot. 2019, 126, 393–399. [Google Scholar] [CrossRef]
- Blackman, R.L.; Eastop, V.F. Aphids on the World’s Crops: An Identification and Information Guide, 2nd ed.; Wiley: Chichester, UK; Weinheim, Germany, 2000; ISBN 978-0-471-85191-2. [Google Scholar]
- Whitfield, A.E.; Falk, B.W.; Rotenberg, D. Insect Vector-Mediated Transmission of Plant Viruses. Virology 2015, 479–480, 278–289. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aphid Species | Treatment ID | Acquisition Period | Inoculation Period | No. and Type of Inoculated Plants |
|---|---|---|---|---|
| Aphis idaei | 1 | 5 min | nt | Five individual virus-free raspberry plants in each treatment |
| 2 | 1 h | 1 h | ||
| 3 | 24 h | 24 h | ||
| 4 | 48 h | 48 h | ||
| Amphorophora rubi idaei | 5 | 5 min | 5 min | Two individual virus-free raspberry plants and two Chenopodium quinoa seedling plants in each treatment |
| 6 | 1 h | 1 h | ||
| 7 | 24 h | 24 h | ||
| 8 | 48 h | 48 h | ||
| Myzus persicae | 9 | 5 min | 5 min | Two individual virus-free raspberry plants and one C. quinoa seedling plant in each treatment |
| 10 | 1 h | 1 h | ||
| 11 | 24 h | 24 h | ||
| 12 | 48 h | 48 h |
| Virus | Sample | ||
|---|---|---|---|
| GR | ALOJ-12 | ALOJ-13 | |
| Raspberry enamovirus 1 | 6883 (0.0032%) | 1482 (0.0019%) | 1759 (0.002%) |
| Raspberry leaf blotch virus | 0 | 362,327 (0.45864%) | 0 |
| Black raspberry necrosis virus | 0 | 213,144 (0.26980%) | 70 (0.00008%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koloniuk, I.; Fránová, J.; Přibylová, J.; Sarkisova, T.; Špak, J.; Tan, J.L.; Zemek, R.; Čmejla, R.; Rejlová, M.; Valentová, L.; et al. Molecular Characterization of a Novel Enamovirus Infecting Raspberry. Viruses 2023, 15, 2281. https://doi.org/10.3390/v15122281
Koloniuk I, Fránová J, Přibylová J, Sarkisova T, Špak J, Tan JL, Zemek R, Čmejla R, Rejlová M, Valentová L, et al. Molecular Characterization of a Novel Enamovirus Infecting Raspberry. Viruses. 2023; 15(12):2281. https://doi.org/10.3390/v15122281
Chicago/Turabian StyleKoloniuk, Igor, Jana Fránová, Jaroslava Přibylová, Tatiana Sarkisova, Josef Špak, Jiunn Luh Tan, Rostislav Zemek, Radek Čmejla, Martina Rejlová, Lucie Valentová, and et al. 2023. "Molecular Characterization of a Novel Enamovirus Infecting Raspberry" Viruses 15, no. 12: 2281. https://doi.org/10.3390/v15122281