
Electrostatic Screening, Acidic pH and Macromolecular Crowding Increase the Self-Assembly Efficiency of the Minute Virus of Mice Capsid In Vitro
 , and
, and 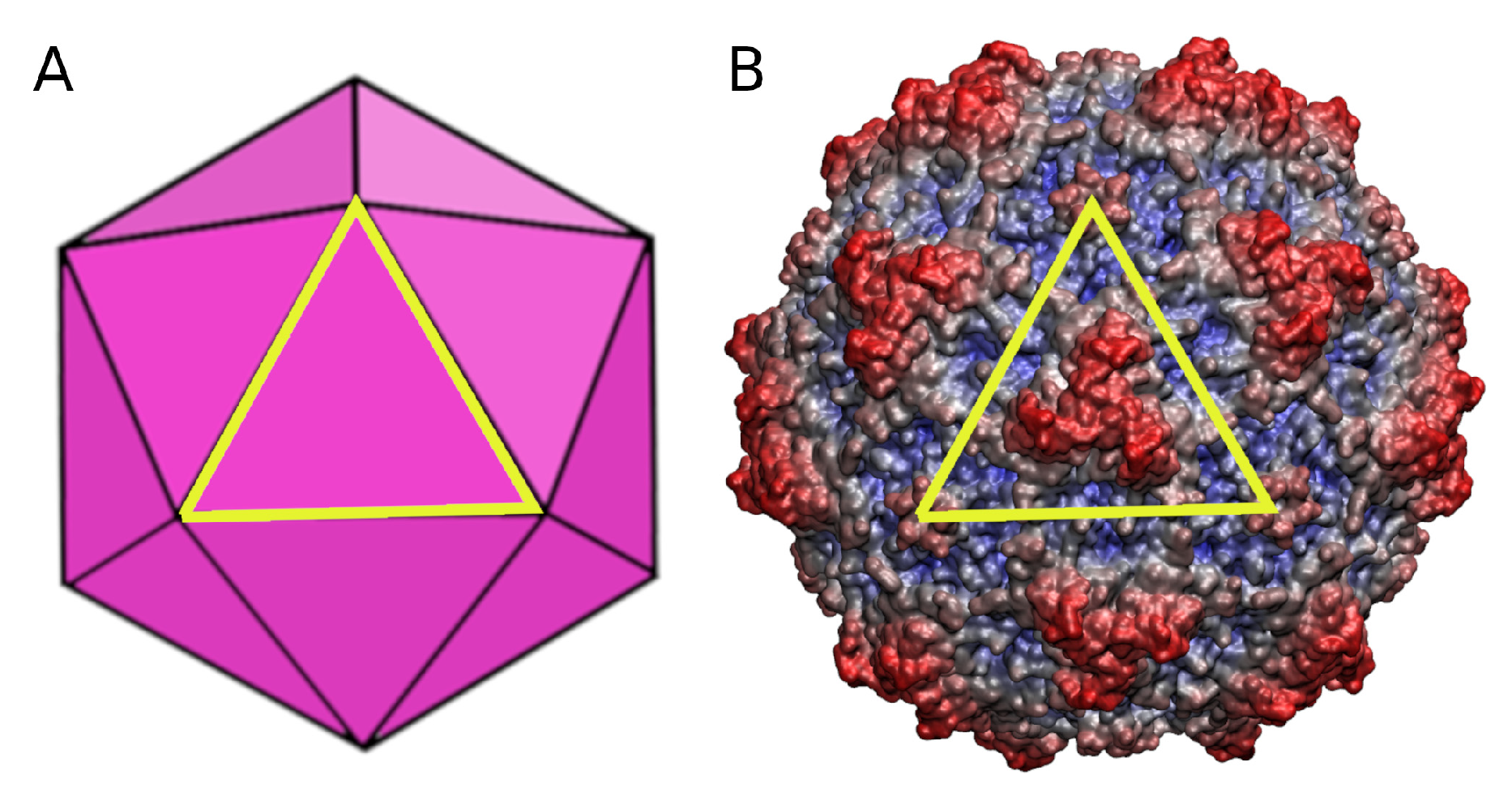{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Expression and Purification of MVM VLPs
2.2. Transmission Electron Microscopy (TEM)
2.3. VLP Disassembly and Reassembly
2.4. VLP Reassembly Quantification and Reassembly Efficiency Calculation
2.5. VLP Kinetic Stability Assay
2.6. Molecular Graphics and Electrostatic Calculations
3. Results
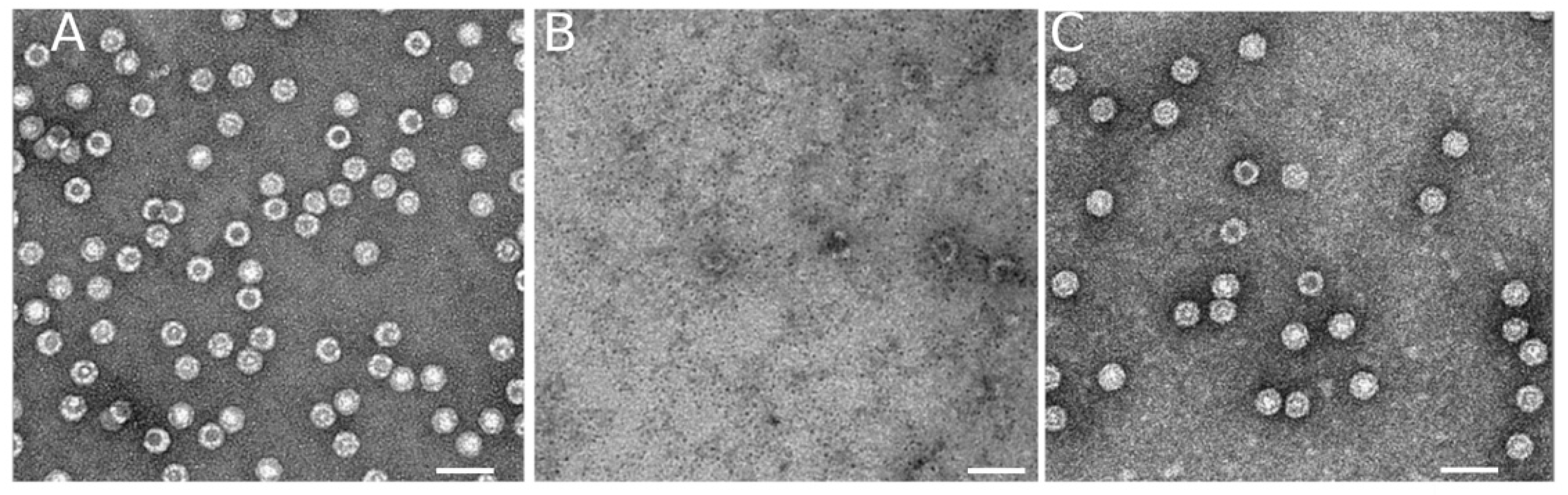
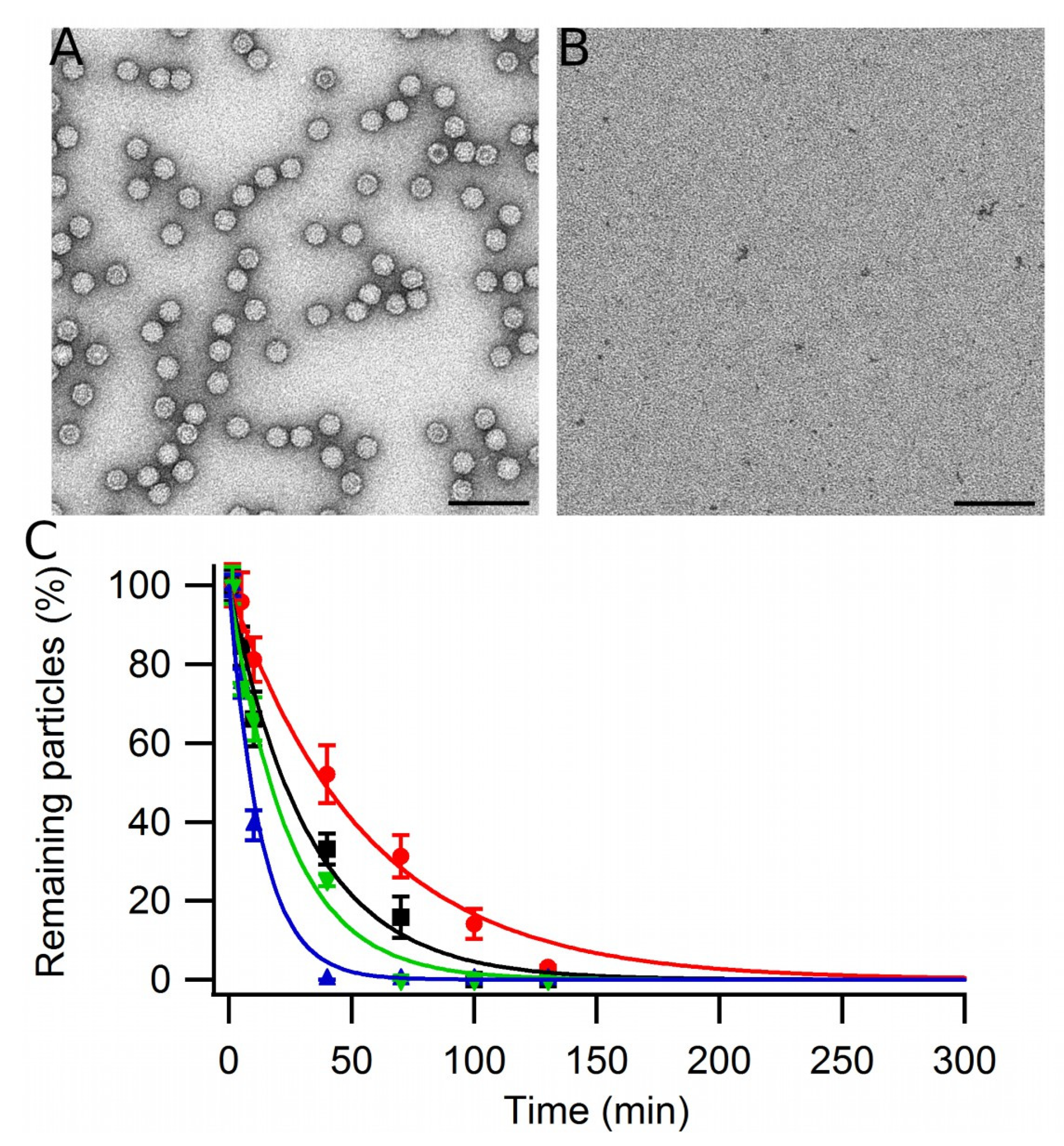
3.1. Disassembly and Reassembly of MVM VLPs
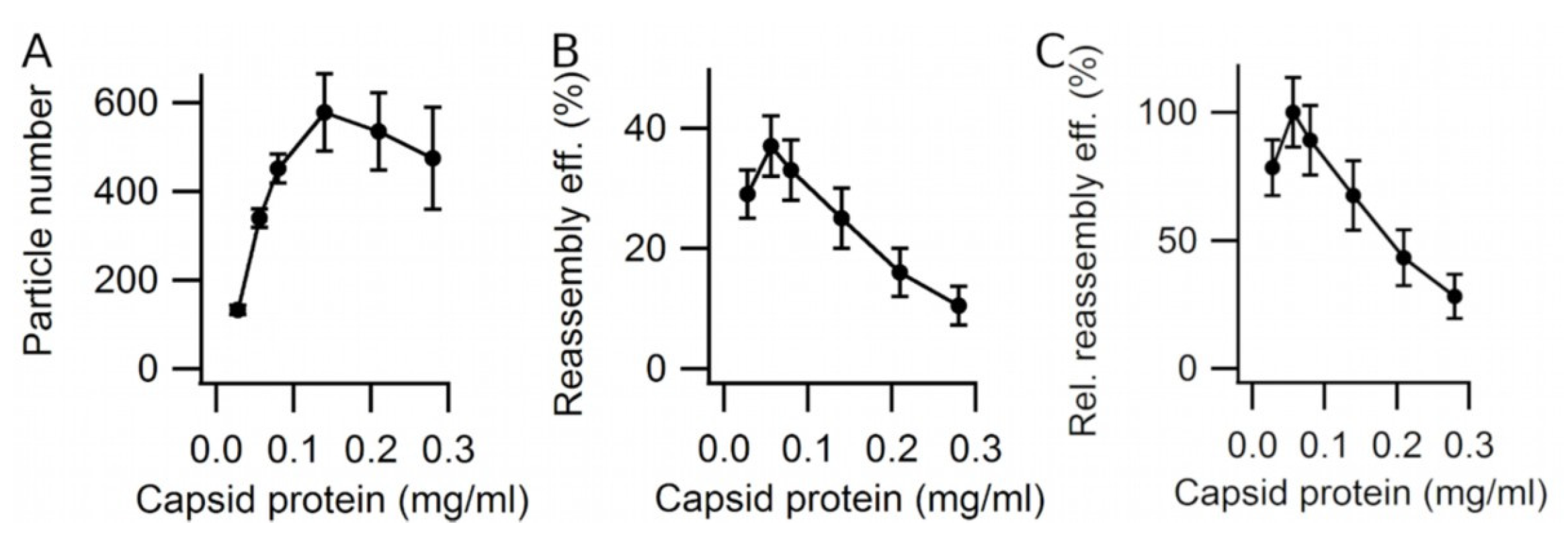
3.2. Effect of Viral Capsid Protein Concentration on VLP Reassembly Efficiency
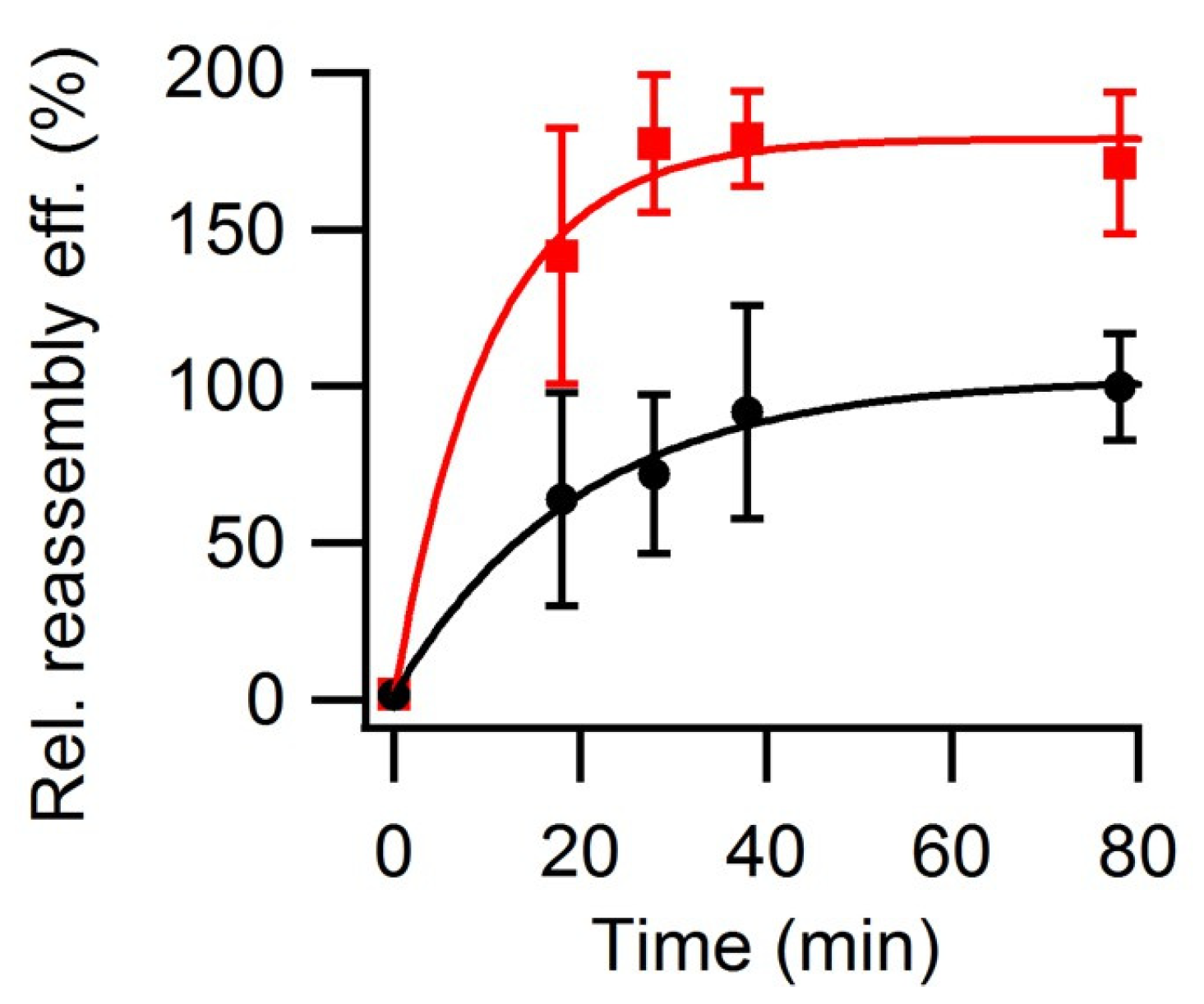
3.3. Effect of Macromolecular Crowding on VLP Reassembly Efficiency
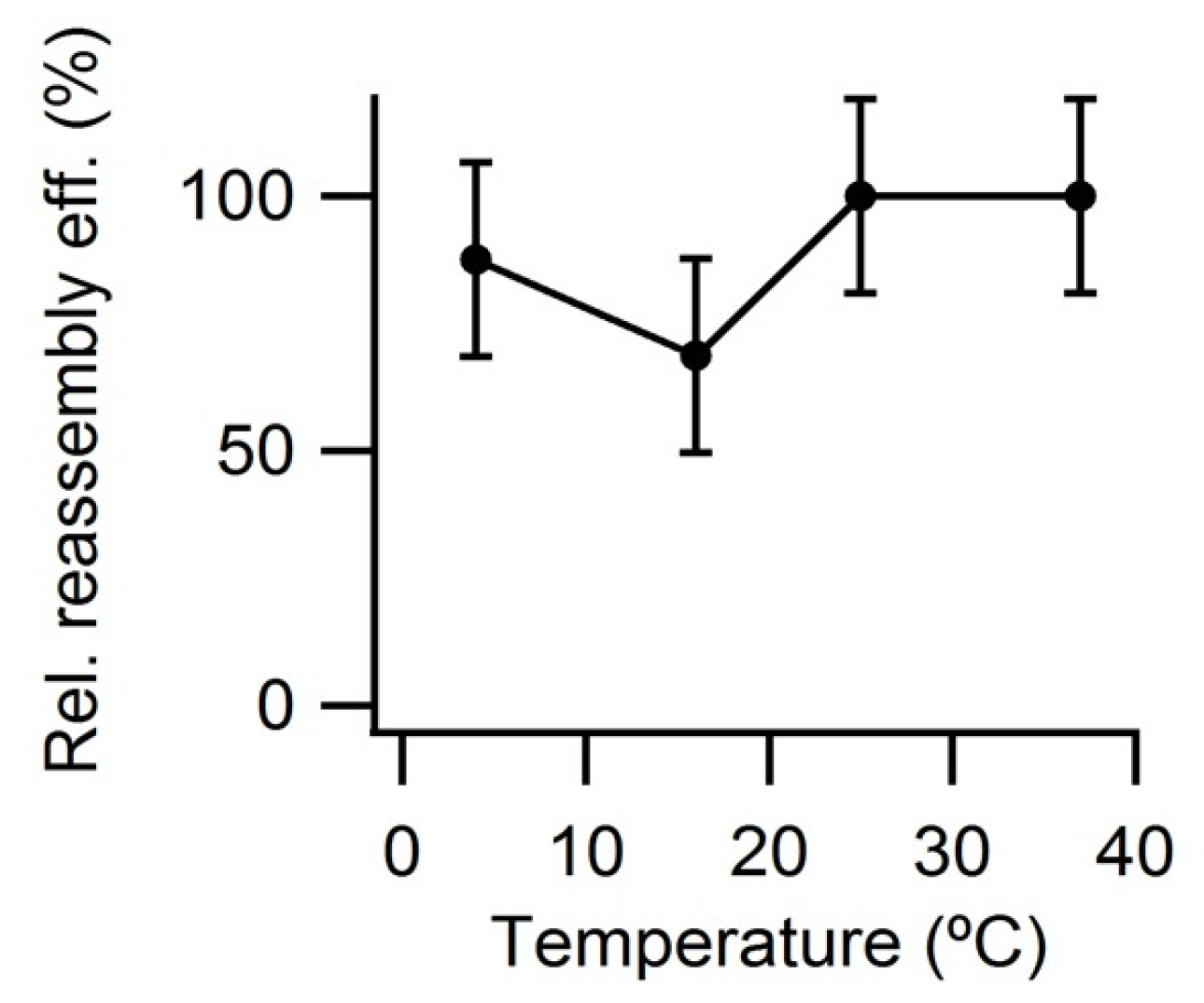
3.4. Effect of Temperature on VLP Reassembly Efficiency
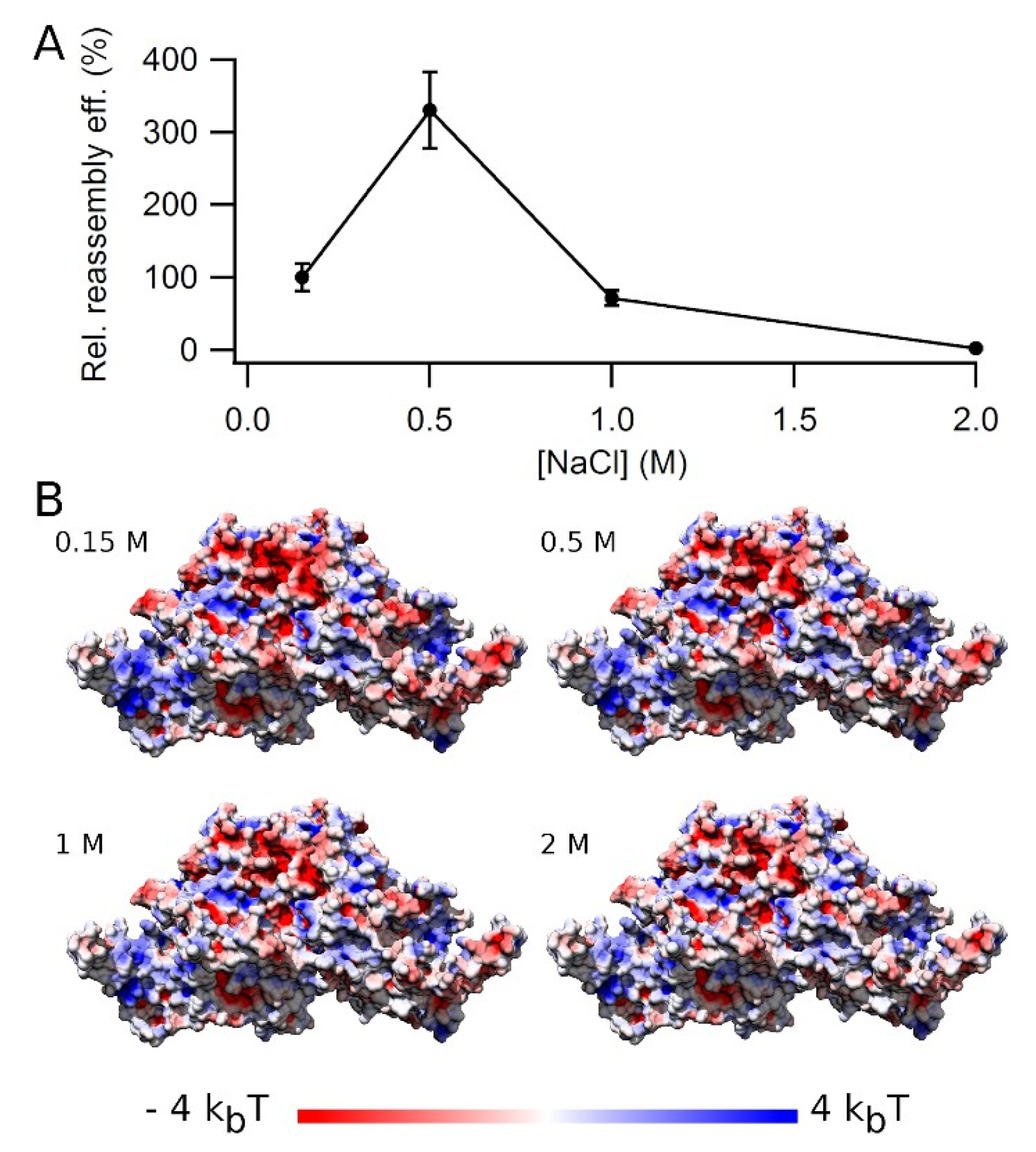
3.5. Effect of Ionic Strength on VLP Reassembly Efficiency
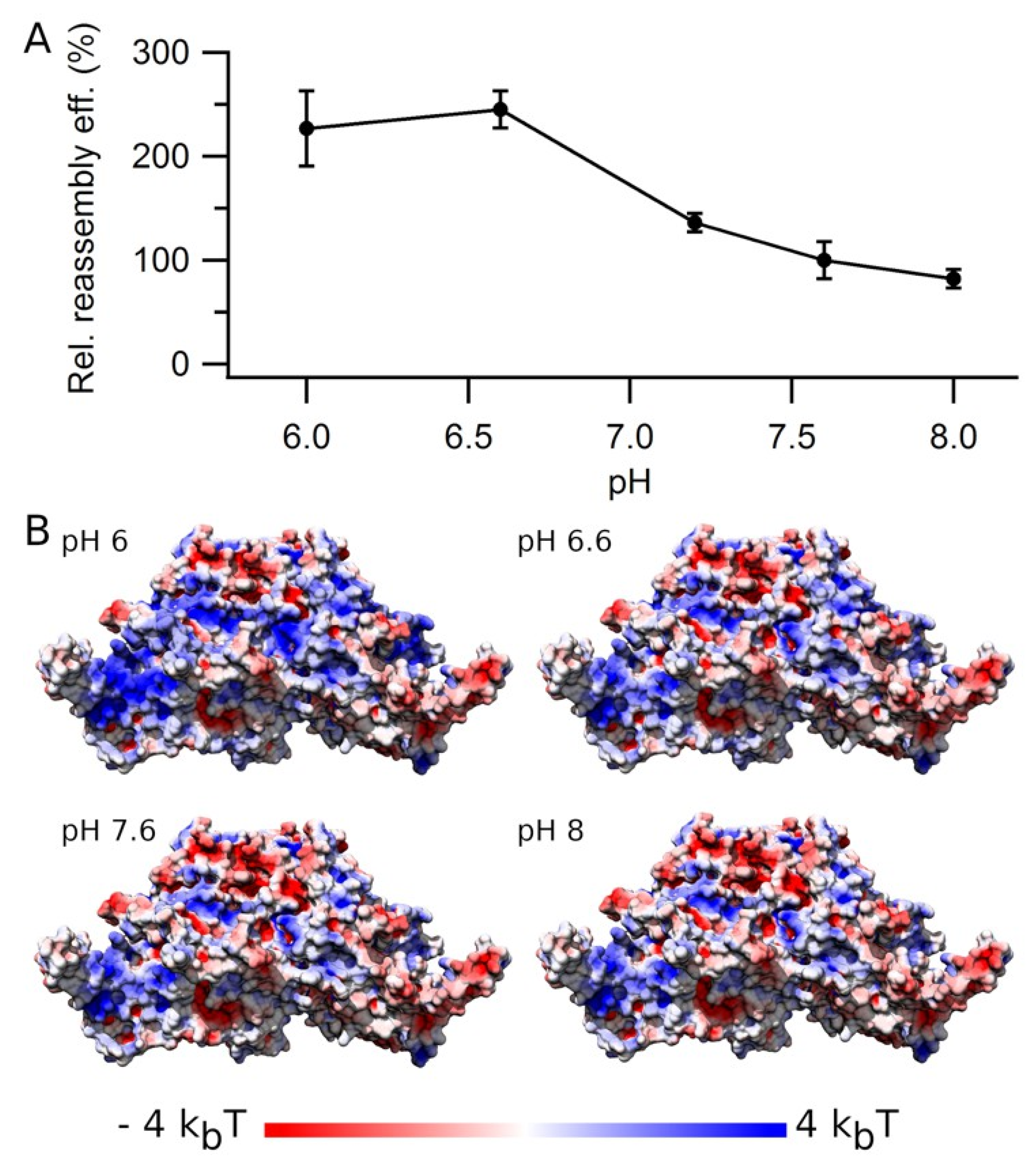
3.6. Effect of pH on VLP Reassembly Efficiency
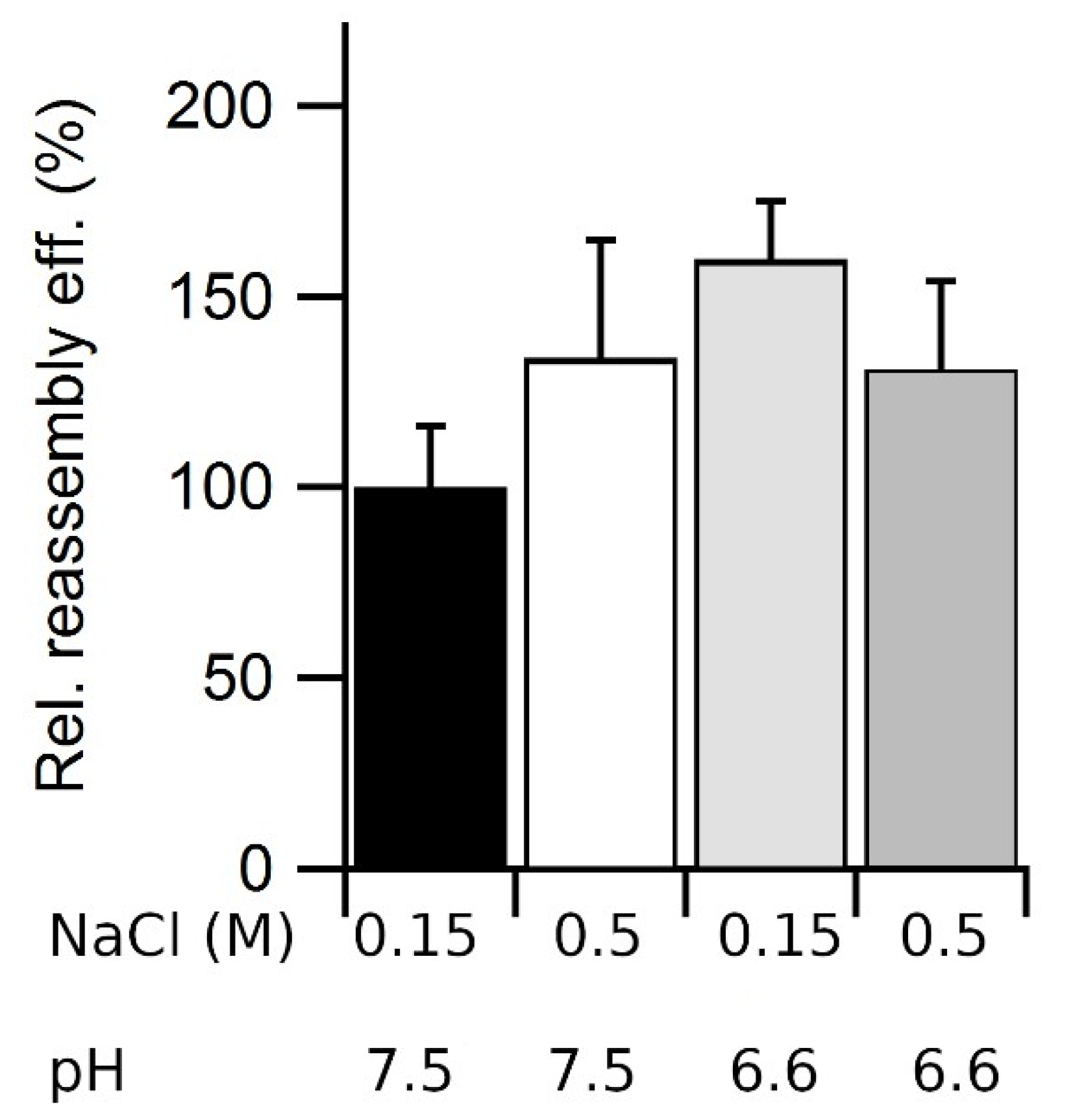
3.7. Combined Effects of Ionic Strength and pH on VLP Reassembly Efficiency
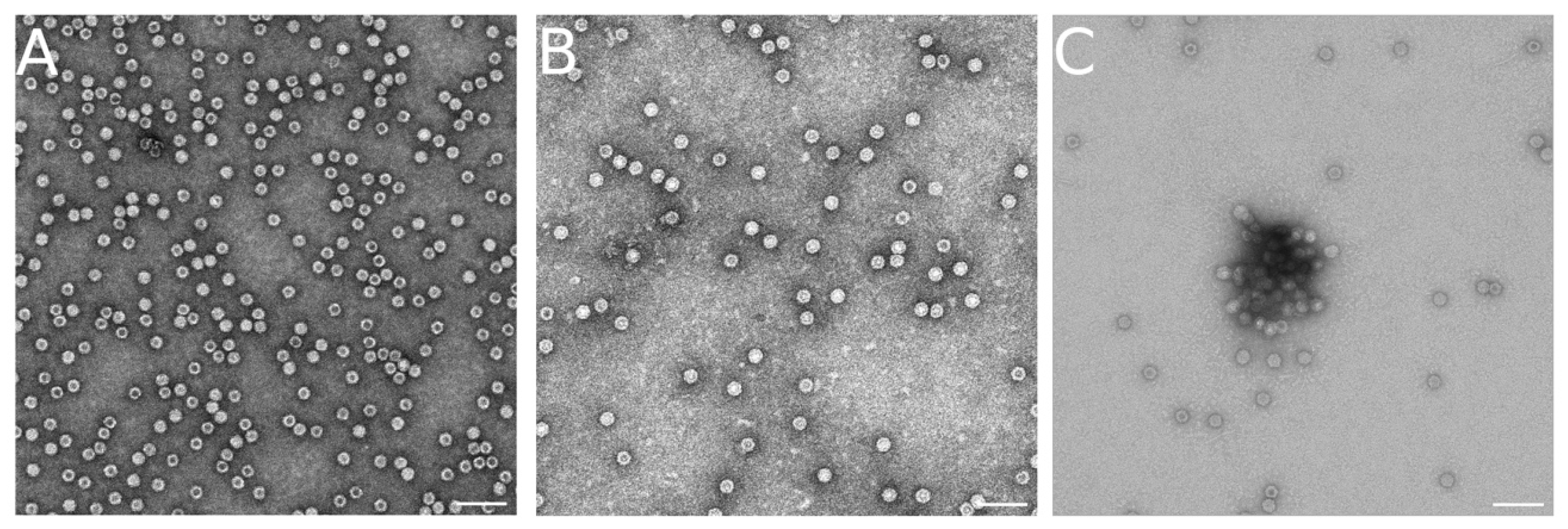
3.8. Dissociation of VLP Aggregates
3.9. Effect of Conditions on Assembly Fidelity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cuervo, A.; Daudén, M.I.; Carrascosa, J.L. Nucleic acid packaging in viruses. Subcell. Biochem. 2013, 68, 361–394. [Google Scholar] [PubMed]
- Mateu, M.G. Assembly, stability and dynamics of virus capsids. Arch. Biochem. Biophys. 2013, 531, 65–79. [Google Scholar] [CrossRef] [PubMed]
- Twarock, R.; Stockley, P.G. RNA-Mediated Virus Assembly: Mechanisms and Consequences for Viral Evolution and Therapy. Annu. Rev. Biophys. 2019, 48, 495–514. [Google Scholar] [CrossRef] [PubMed]
- Bancroft, J.B.; Hiebert, E.; Bracker, C.E. The effects of various polyanions on shell formation of some spherical viruses. Virology 1969, 39, 924–930. [Google Scholar] [CrossRef]
- Adolph, K.W.; Butler, P.J. Studies on the assembly of a spherical plant virus. I. States of aggregation of the isolated protein. J. Mol. Biol. 1974, 88, 327–341. [Google Scholar] [CrossRef]
- Lavelle, L.; Michel, J.-P.; Gingery, M. The disassembly, reassembly and stability of CCMV protein capsids. J. Virol. Methods 2007, 146, 311–316. [Google Scholar] [CrossRef]
- Douglas, T.; Young, M. Viruses: Making friends with old foes. Science 2006, 312, 873–875. [Google Scholar] [CrossRef]
- Glasgow, J.; Tullman-Ercek, D. Production and applications of engineered viral capsids. Appl. Microbiol. Biotechnol. 2014, 98, 5847–5858. [Google Scholar] [CrossRef]
- McNeale, D.; Dashti, N.; Cheah, L.C.; Sainsbury, F. Protein cargo encapsulation by virus-like particles: Strategies and applications. WIREs Nanomed. Nanobiotechnol. 2022, e1869. [Google Scholar] [CrossRef]
- Mateu, M.G. Assembly, Engineering and Applications of Virus-Based Protein Nanoparticles. Adv. Exp. Med. Biol. 2016, 940, 83–120. [Google Scholar]
- Ikwuagwu, B.; Tullman-Ercek, D. Virus-like particles for drug delivery: A review of methods and applications. Curr. Opin. Biotechnol. 2022, 78, 102785. [Google Scholar] [CrossRef] [PubMed]
- Wijesundara, Y.H.; Herbert, F.C.; Kumari, S.; Howlett, T.; Koirala, S.; Trashi, O.; Trashi, I.; Al-Kharji, N.M.; Gassensmith, J.J. Rip it, stitch it, click it: A Chemist’s guide to VLP manipulation. Virology 2022, 577, 105–123. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, M.P.; White, W.I.; Palmer-Hill, F.; Koenig, S.; Suzich, J.A. Quantitative disassembly and reassembly of human papillomavirus type 11 viruslike particles in vitro. J. Virol. 1998, 72, 32–41. [Google Scholar] [CrossRef]
- Mach, H.; Volkin, D.B.; Troutman, R.D.; Wang, B.; Luo, Z.; Jansen, K.U.; Shi, L. Disassembly and reassembly of yeast-derived recombinant human papillomavirus virus-like particles (HPV VLPs). J. Pharm. Sci. 2006, 95, 2195–2206. [Google Scholar] [CrossRef]
- Ehrlich, L.S.; Agresta, B.E.; Carter, C.A. Assembly of recombinant human immunodeficiency virus type 1 capsid protein in vitro. J. Virol. 1992, 66, 4874–4883. [Google Scholar] [CrossRef]
- Campbell, S.; Vogt, V.M. Self-assembly in vitro of purified CA-NC proteins from Rous sarcoma virus and human immunodeficiency virus type 1. J. Virol. 1995, 69, 6487–6497. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; DuFort, C.; Daniel, M.-C.; Murali, A.; Chen, C.; Gopinath, K.; Stein, B.; De, M.; Rotello, V.M.; Holzenburg, A.; et al. Core-controlled polymorphism in virus-like particles. Proc. Natl. Acad. Sci. USA 2007, 104, 1354–1359. [Google Scholar] [CrossRef]
- Bajaj, S.; Banerjee, M. In vitro assembly of polymorphic virus-like particles from the capsid protein of a nodavirus. Virology 2016, 496, 106–115. [Google Scholar] [CrossRef]
- De Ruiter, M.V.; van der Hee, R.M.; Driessen, A.J.M.; Keurhorst, E.D.; Hamid, M.; Cornelissen, J.J.L.M. Polymorphic assembly of virus-capsid proteins around DNA and the cellular uptake of the resulting particles. J. Control. Release Off. J. Control. Release Soc. 2019, 307, 342–354. [Google Scholar] [CrossRef]
- Douglas, C.C.; Thomas, D.; Lanman, J.; Prevelige, P.E. Investigation of N-Terminal Domain Charged Residues on the Assembly and Stability of HIV-1 CA. Biochemistry 2004, 43, 10435–10441. [Google Scholar] [CrossRef]
- Del Alamo, M.; Mateu, M.G. Electrostatic repulsion, compensatory mutations, and long-range non-additive effects at the dimerization interface of the HIV capsid protein. J. Mol. Biol. 2005, 345, 893–906. [Google Scholar] [CrossRef]
- Del Alamo, M.; Rivas, G.; Mateu, M.G. Effect of macromolecular crowding agents on human immunodeficiency virus type 1 capsid protein assembly in vitro. J. Virol. 2005, 79, 14271–14281. [Google Scholar] [CrossRef] [PubMed]
- Ganser-Pornillos Barbie, K.; von Schwedler Uta, K.; Stray Kirsten, M.; Christopher, A.; Sundquist Wesley, I. Assembly Properties of the Human Immunodeficiency Virus Type 1 CA Protein. J. Virol. 2004, 78, 2545–2552. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Perilla, J.R.; Yufenyuy, E.L.; Meng, X.; Chen, B.; Ning, J.; Ahn, J.; Gronenborn, A.M.; Schulten, K.; Aiken, C.; et al. Mature HIV-1 capsid structure by cryo-electron microscopy and all-atom molecular dynamics. Nature 2013, 497, 643–646. [Google Scholar] [CrossRef] [PubMed]
- Cotmore, S.F.; Tattersall, P. Parvoviruses: Small Does Not Mean Simple. Annu. Rev. Virol. 2014, 1, 517–537. [Google Scholar] [CrossRef]
- Mietzsch, M.; Pénzes, J.J.; Agbandje-McKenna, M. Twenty-five years of structural parvovirology. Viruses 2019, 11, 362. [Google Scholar] [CrossRef]
- Singh, P.; Destito, G.; Schneemann, A.; Manchester, M. Canine parvovirus-like particles, a novel nanomaterial for tumor targeting. J. Nanobiotechnol. 2006, 4, 2. [Google Scholar] [CrossRef]
- Singh, P. Tumor targeting using canine parvovirus nanoparticles. Curr. Top. Microbiol. Immunol. 2009, 327, 123–141. [Google Scholar]
- Roldão, A.; Mellado, M.C.M.; Castilho, L.R.; Carrondo, M.J.T.; Alves, P.M. Virus-like particles in vaccine development. Expert Rev. Vaccines 2010, 9, 1149–1176. [Google Scholar] [CrossRef]
- Tu, M.; Liu, F.; Chen, S.; Wang, M.; Cheng, A. Role of capsid proteins in parvoviruses infection. Virol. J. 2015, 12, 114. [Google Scholar] [CrossRef]
- Yan, D.; Wang, B.; Sun, S.; Feng, X.; Jin, Y.; Yao, X.; Cao, S.; Guo, H. Quantum Dots Encapsulated with Canine Parvovirus-Like Particles Improving the Cellular Targeted Labeling. PLoS ONE 2015, 10, e0138883. [Google Scholar] [CrossRef] [PubMed]
- Cayetano-Cruz, M.; Coffeen, C.F.; Valadez-García, J.; Montiel, C.; Bustos-Jaimes, I. Decoration of virus-like particles with an enzymatic activity of biomedical interest. Virus Res. 2018, 255, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Thadani, N.N.; Dempsey, C.; Zhao, J.; Vasquez, S.M.; Suh, J. Reprogramming the Activatable Peptide Display Function of Adeno-Associated Virus Nanoparticles. ACS Nano 2018, 12, 1445–1454. [Google Scholar] [CrossRef]
- Li, C.; Samulski, R.J. Engineering adeno-associated virus vectors for gene therapy. Nat. Rev. Genet. 2020, 21, 255–272. [Google Scholar] [CrossRef] [PubMed]
- Hashemzadeh, M.S.; Gharari, N. Biosynthesis of a VLP-type nanocarrier specific to cancer cells using the BEVS expression system for targeted drug delivery. J. Genet. Eng. Biotechnol. 2023, 21, 20. [Google Scholar] [CrossRef] [PubMed]
- Agbandje-McKenna, M.; Llamas-Saiz, A.L.; Wang, F.; Tattersall, P.; Rossmann, M.G. Functional implications of the structure of the murine parvovirus, minute virus of mice. Structure 1998, 6, 1369–1381. [Google Scholar] [CrossRef]
- Hernando, E.; Llamas-Saiz, A.L.; Foces-Foces, C.; McKenna, R.; Portman, I.; Agbandje-McKenna, M.; Almendral, J.M. Biochemical and physical characterization of parvovirus minute virus of mice virus-like particles. Virology 2000, 267, 299–309. [Google Scholar] [CrossRef]
- Medrano, M.; Fuertes, M.Á.; Valbuena, A.; Carrillo, P.J.P.; Rodríguez-Huete, A.; Mateu, M.G. Imaging and Quantitation of a Succession of Transient Intermediates Reveal the Reversible Self-Assembly Pathway of a Simple Icosahedral Virus Capsid. J. Am. Chem. Soc. 2016, 138, 15385–15396. [Google Scholar] [CrossRef]
- Ros, C.; Bayat, N.; Wolfisberg, R.; Almendral, J.M. Protoparvovirus Cell Entry. Viruses 2017, 9, 313. [Google Scholar] [CrossRef]
- Mattola, S.; Aho, V.; Bustamante-Jaramillo, L.F.; Pizzioli, E.; Kann, M.; Vihinen-Ranta, M. Nuclear entry and egress of parvoviruses. Mol. Microbiol. 2022, 118, 295–308. [Google Scholar] [CrossRef]
- Fuertes, M.A.; Pérez, J.M.; Alonso, C. Small amounts of urea and guanidine hydrochloride can be detected by a far-UV spectrophotometric method in dialysed protein solutions. J. Biochem. Biophys. Methods 2004, 59, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Tseng, Q.; Wang, I.; Duchemin-Pelletier, E.; Azioune, A.; Carpi, N.; Gao, J.; Filhol, O.; Piel, M.; Théry, M.; Balland, M. A new micropatterning method of soft substrates reveals that different tumorigenic signals can promote or reduce cell contraction levels. Lab Chip 2011, 11, 2231. [Google Scholar] [CrossRef]
- Kontou, M.; Govindasamy, L.; Nam, H.J.; Bryant, N.; Llamas-Saiz, A.L.; Foces’Foces, C.; Hernando, E.; Rubio, M.P.; McKenna, R.; Almendral, J.M.; et al. Structural determinants of tissue tropism and in vivo pathogenicity for the parvovirus minute virus of mice. J. Virol. 2005, 79, 10931–10943. [Google Scholar] [CrossRef] [PubMed]
- Holst, M.; Saied, F. Multigrid solution of the Poisson-Boltzmann equation. J. Comput. Chem. 1993, 14, 105–113. [Google Scholar] [CrossRef]
- Baker, N.A.; Sept, D.; Joseph, S.; Holst, M.J.; McCammon, J.A. Electrostatics of Nanosystems: Application to Microtubules and the Ribosome. Proc. Natl. Acad. Sci. USA 2001, 98, 10037–10041. [Google Scholar] [CrossRef]
- Dolinsky, T.J.; Nielsen, J.E.; McCammon, J.A.; Baker, N.A. PDB2PQR: An Automated Pipeline for the Setup of Poisson Boltzmann Electrostatics Calculations. Nucleic Acids Res. 2004, 32, 665–667. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A Visualization System for Exploratory Research and Analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]
- Riolobos, L.; Reguera, J.; Mateu, M.G.; Almendral, J.M. Nuclear transport of trimeric assembly intermediates exerts a morphogenetic control on the icosahedral parvovirus capsid. J. Mol. Biol. 2006, 357, 1026–1038. [Google Scholar] [CrossRef]
- Almendral, J.M. Assembly of simple icosahedral viruses. Subcell. Biochem. 2013, 68, 307–328. [Google Scholar]
- Lombardo, E.; Ramírez, J.C.; Agbandje-McKenna, M.; Almendral, J.M. A beta-stranded motif drives capsid protein oligomers of the parvovirus minute virus of mice into the nucleus for viral assembly. J. Virol. 2000, 74, 3804–3814. [Google Scholar] [CrossRef]
- Lombardo, E.; Ramírez, J.C.; García, J.; Almendral, J.M. Complementary roles of multiple nuclear targeting signals in the capsid proteins of the parvovirus minute virus of mice during assembly and onset of infection. J. Virol. 2002, 76, 7049–7059. [Google Scholar] [CrossRef] [PubMed]
- Riolobos, L.; Valle, N.; Hernando, E.; Maroto, B.; Kann, M.; Almendral, J.M. Viral oncolysis that targets Raf-1 signaling control of nuclear transport. J. Virol. 2010, 84, 2090–2099. [Google Scholar] [CrossRef] [PubMed]
- Maroto, B.; Ramírez, J.C.; Almendral, J.M. Phosphorylation status of the parvovirus minute virus of mice particle: Mapping and biological relevance of the major phosphorylation sites. J. Virol. 2000, 74, 10892–10902. [Google Scholar] [CrossRef] [PubMed]
- Cotmore, S.F.; D’Abramo, A.M., Jr.; Carbonell, L.F.; Bratton, J.; Tattersall, P. The NS2 polypeptide of parvovirus MVM is required for capsid assembly in murine cells. Virology 1997, 231, 267–280. [Google Scholar] [CrossRef] [PubMed]
- Sonntag, F.; Schmidt, K.; Kleinschmidt, J.A. A viral assembly factor promotes AAV2 capsid formation in the nucleolus. Proc. Natl. Acad. Sci. USA 2010, 107, 10220–10225. [Google Scholar] [CrossRef]
- Plevka, P.; Hafenstein, S.; Li, L.; D’Abramo, A.; Cotmore, S.F.; Rossmann, M.G.; Tattersall, P. Structure of a packaging-defective mutant of minute virus of mice indicates that the genome is packaged via a pore at a 5-fold axis. J. Virol. 2011, 85, 4822–4827. [Google Scholar] [CrossRef]
- Yuan, W.; Parrish, C. Canine parvovirus capsid assembly and differences in mammalian and insect cells. Virology 2001, 279, 546–557. [Google Scholar] [CrossRef]
- Maranga, L.; Rueda, P.; Antonis, A.F.; Vela, C.; Langeveld, J.P.; Casal, J.I.; Carrondo, M.J. Large scale production and downstream processing of a recombinant porcine parvovirus vaccine. Appl. Microbiol. Biotechnol. 2002, 59, 45–50. [Google Scholar]
- Steinbach, S.; Wistuba, A.; Bock, T.; Kleinschmidt, J.A. Assembly of adeno-associated virus type 2 capsids in vitro. J. Gen. Virol. 1997, 78, 1453–1462. [Google Scholar] [CrossRef]
- Sánchez-Rodríguez, S.P.; Münch-Anguiano, L.; Echeverría, O.; Vázquez-Nin, G.; Mora-Pale, M.; Dordick, J.S.; Bustos-Jaimes, I. Human parvovirus B19 virus-like particles: In vitro assembly and stability. Biochimie 2012, 94, 870–878. [Google Scholar] [CrossRef] [PubMed]
- Luque, A.; Reguera, D. Theoretical studies on assembly, physical stability and dynamics of viruses. Subcell. Biochem. 2013, 68, 553–595. [Google Scholar] [PubMed]
- Perlmutter, J.D.; Hagan, M.F. Mechanisms of virus assembly. Annu. Rev. Phys. Chem. 2015, 66, 217–239. [Google Scholar] [CrossRef] [PubMed]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fuertes, M.A.; López Mateos, D.; Valiente, L.; Rodríguez Huete, A.; Valbuena, A.; Mateu, M.G. Electrostatic Screening, Acidic pH and Macromolecular Crowding Increase the Self-Assembly Efficiency of the Minute Virus of Mice Capsid In Vitro. Viruses 2023, 15, 1054. https://doi.org/10.3390/v15051054
Fuertes MA, López Mateos D, Valiente L, Rodríguez Huete A, Valbuena A, Mateu MG. Electrostatic Screening, Acidic pH and Macromolecular Crowding Increase the Self-Assembly Efficiency of the Minute Virus of Mice Capsid In Vitro. Viruses. 2023; 15(5):1054. https://doi.org/10.3390/v15051054
Chicago/Turabian StyleFuertes, Miguel Angel, Diego López Mateos, Luis Valiente, Alicia Rodríguez Huete, Alejandro Valbuena, and Mauricio G. Mateu. 2023. "Electrostatic Screening, Acidic pH and Macromolecular Crowding Increase the Self-Assembly Efficiency of the Minute Virus of Mice Capsid In Vitro" Viruses 15, no. 5: 1054. https://doi.org/10.3390/v15051054