
Differences in Pathogenicity and Vaccine Resistance Discovered between Two Epidemic Strains of Marek’s Disease Virus in China

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Clinical Samples
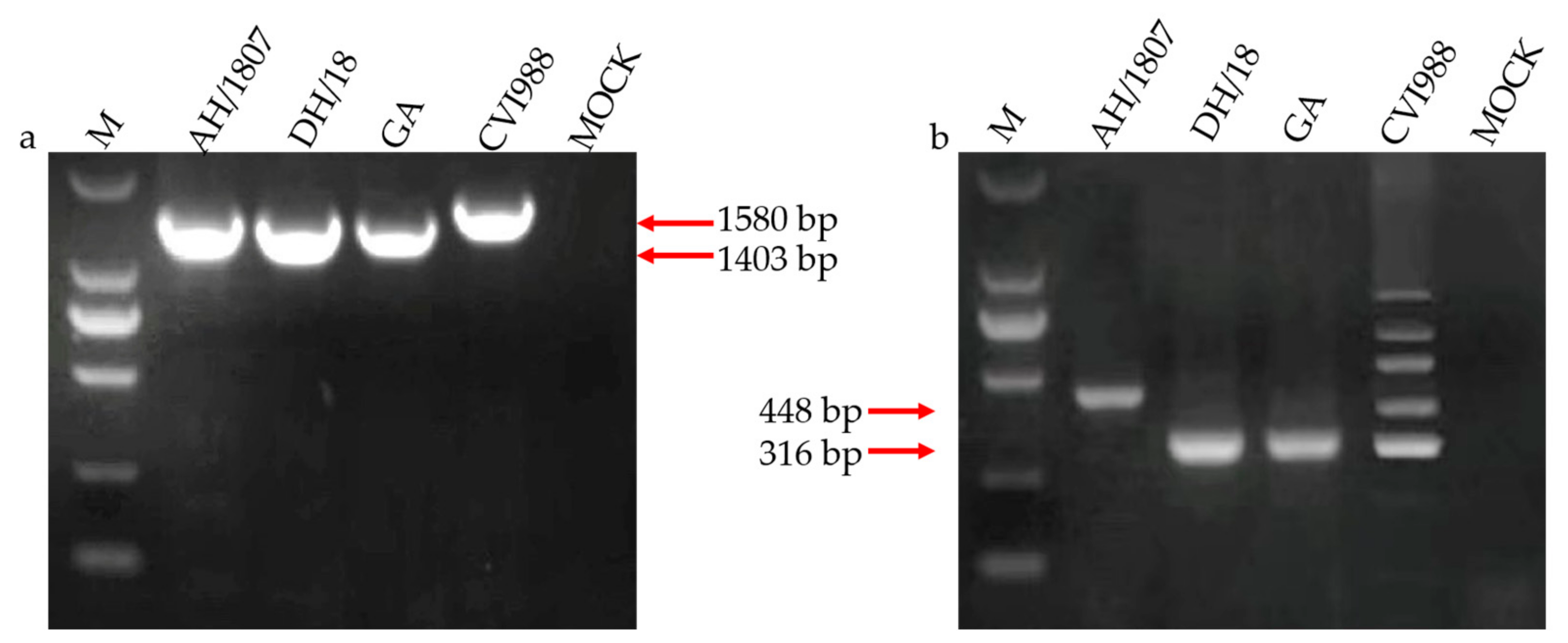
2.2. Viral Isolation and Identification
2.3. Screening of Adventitious Agents
2.4. Pathogenic Processes and Virulence Studies
2.5. DNA and RNA Extraction and Quantitative Real-Time Polymerase Chain Reaction
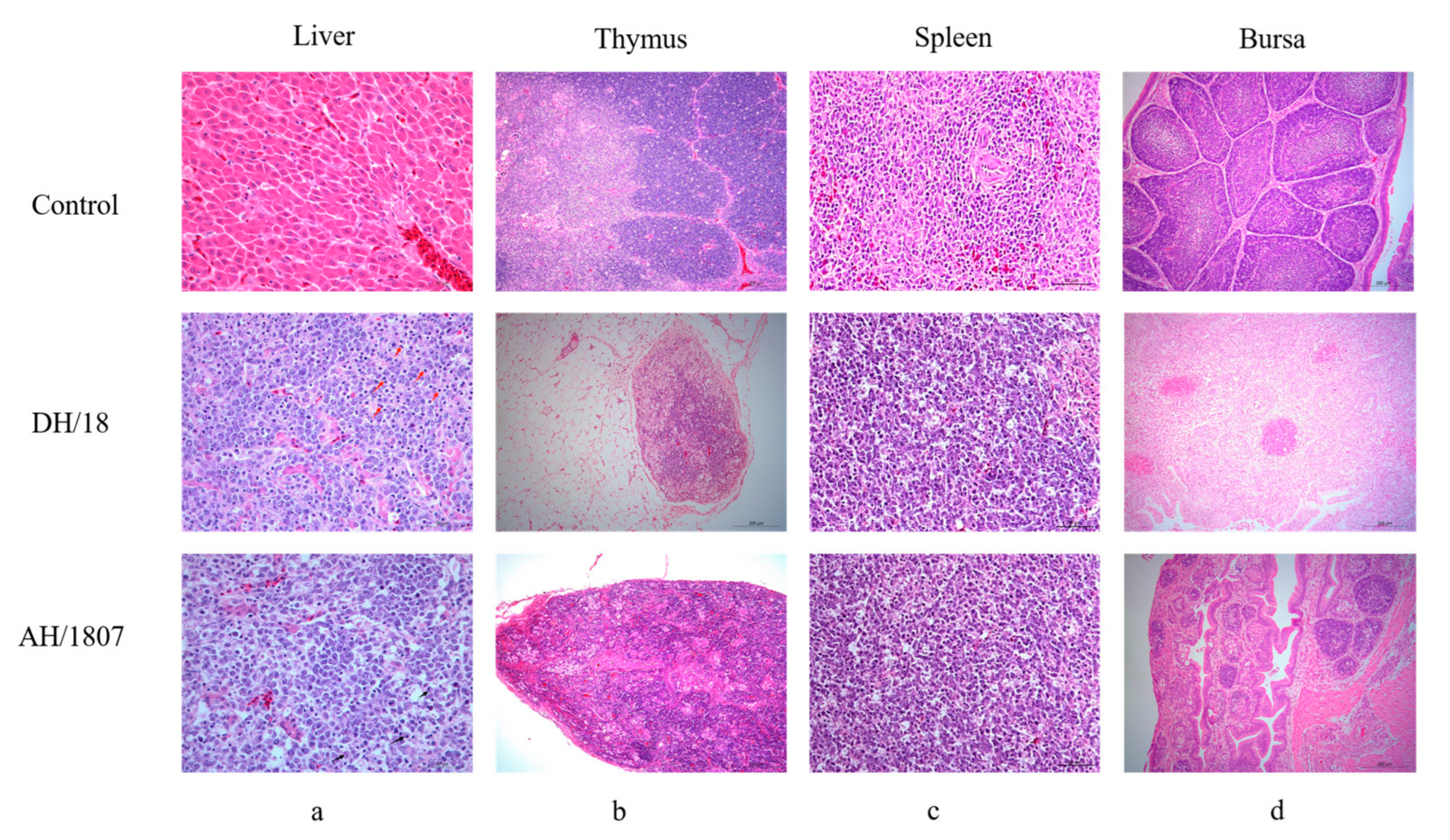
2.6. Histopathological Examination
2.7. Sequence Alignment and Phylogenic Analysis
2.8. Statistical Analysis
3. Results
3.1. Two MDV Strains Isolated in China
3.2. Occurrence Rate and Distribution of MD Tumors
3.3. MD Morbidity and Mortality
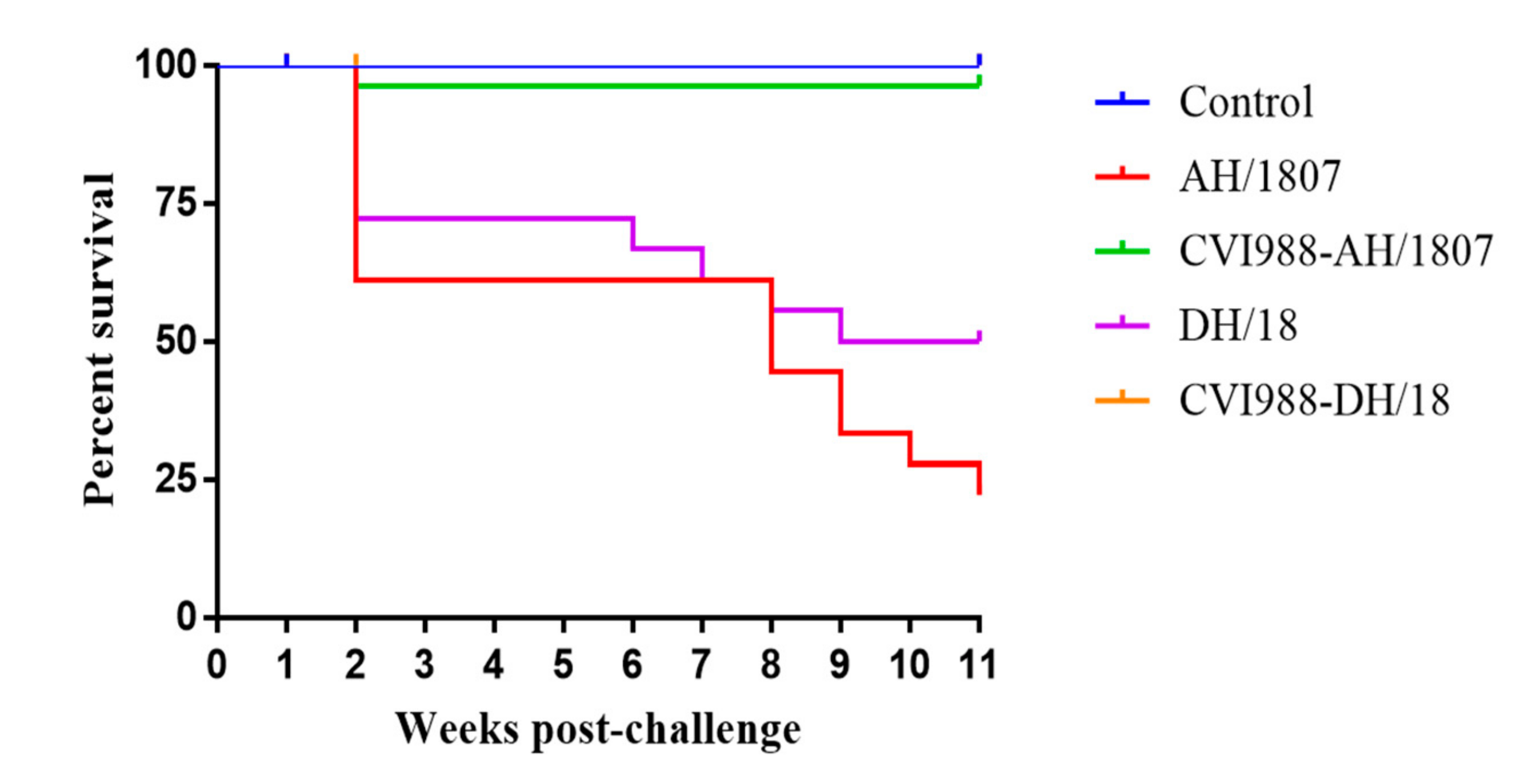
3.4. Survival Analysis
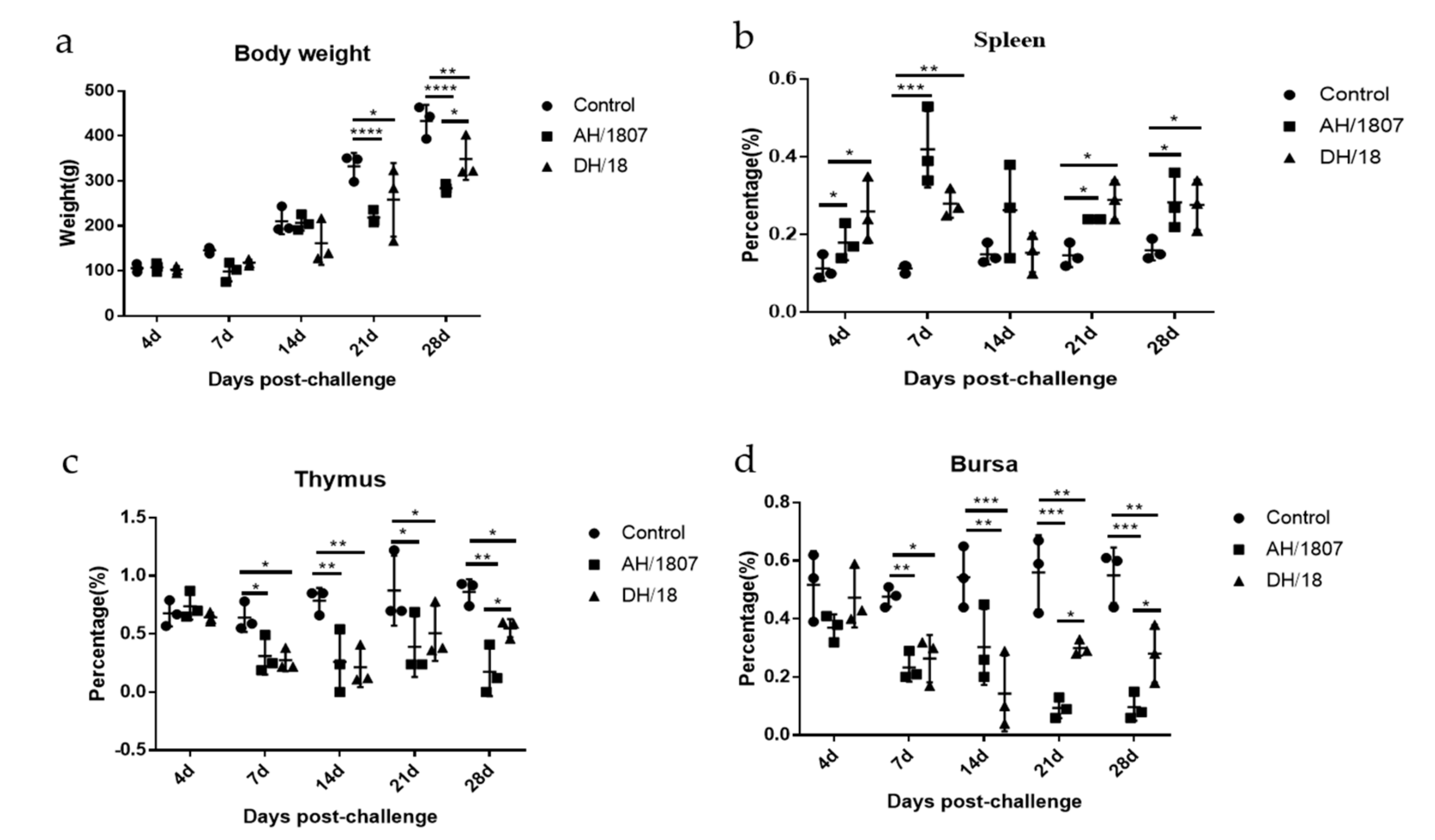
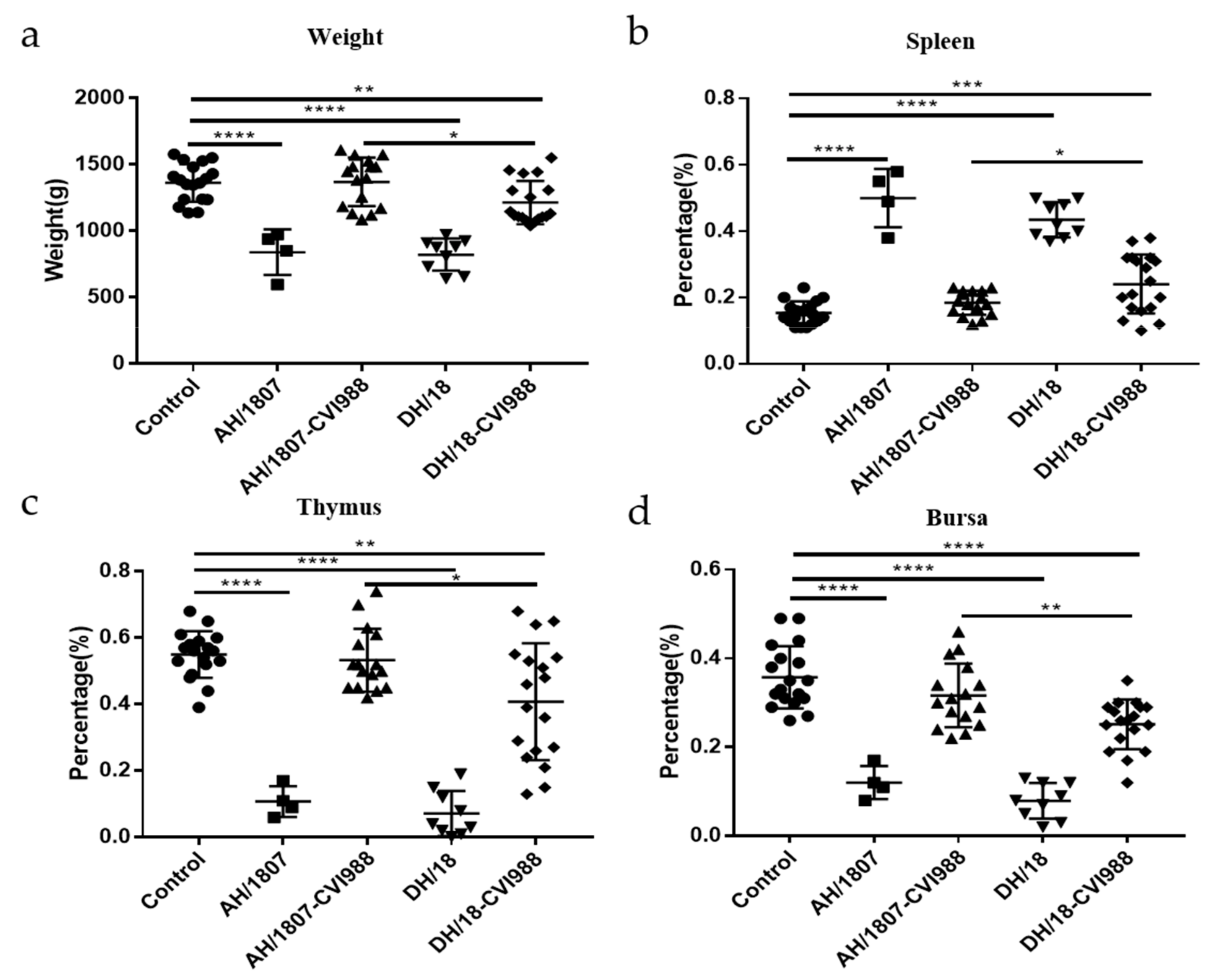
3.5. Developmental Disorders and Immune Organ Damage
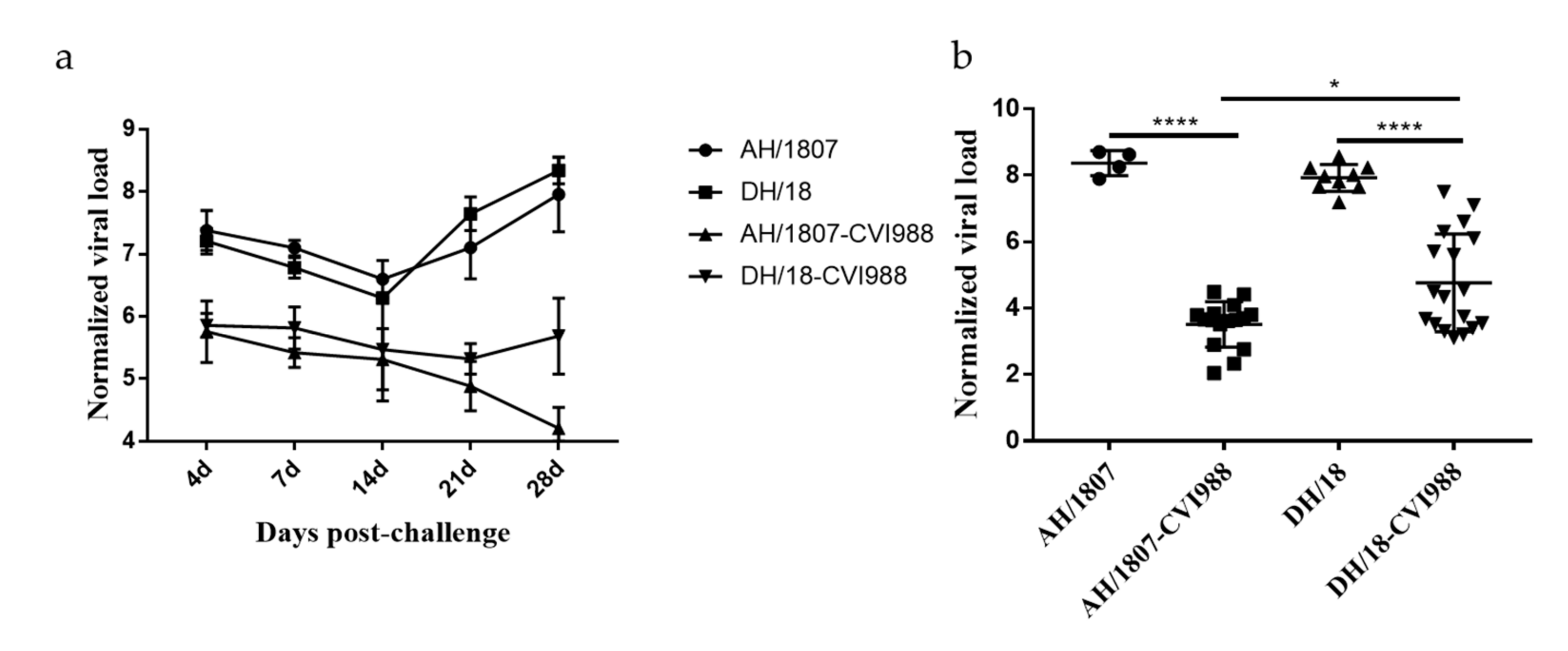
3.6. Kinetics of Viral Replication In Vivo
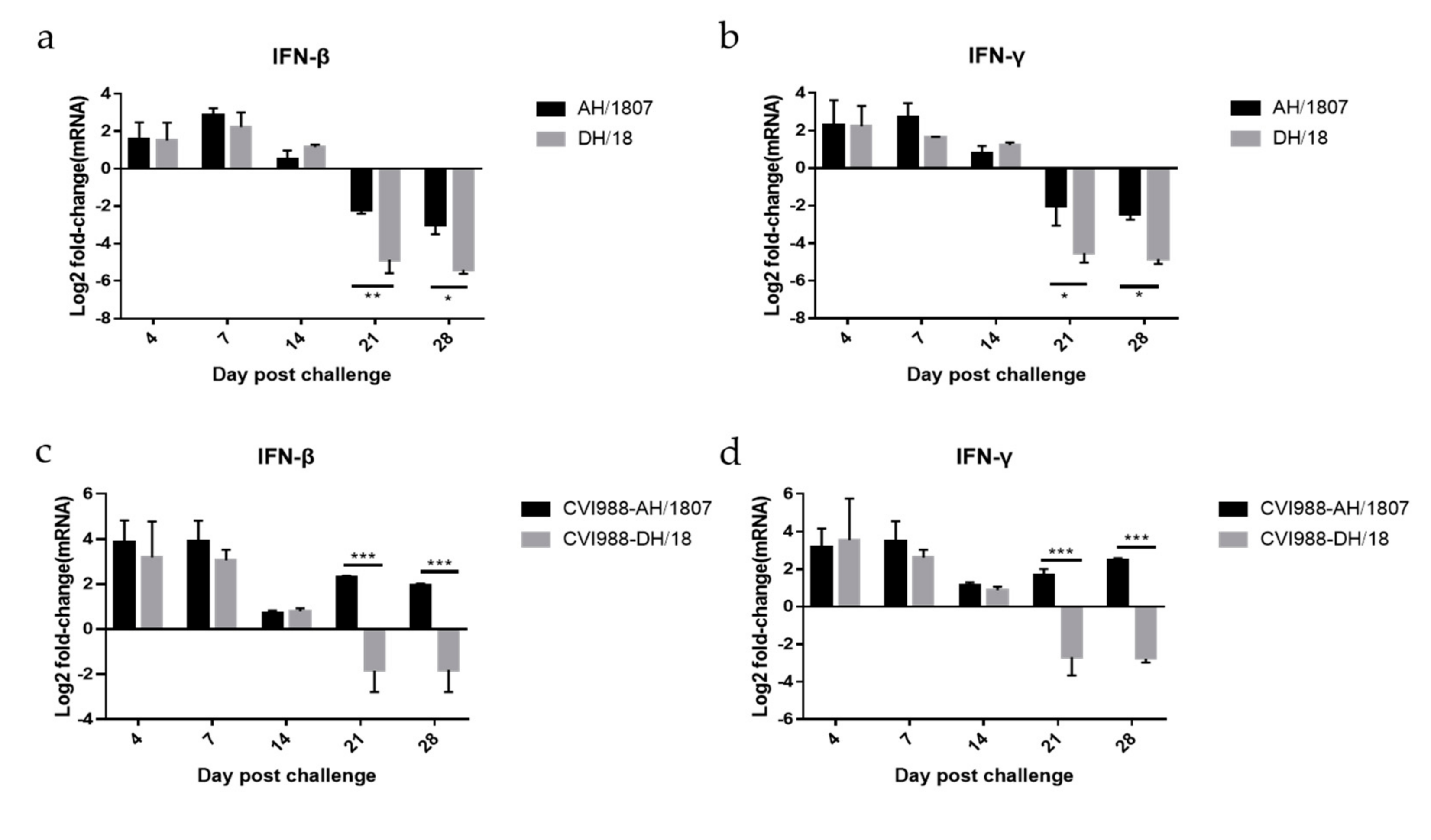
3.7. Expression of IFN-β and IFN-γ mRNA
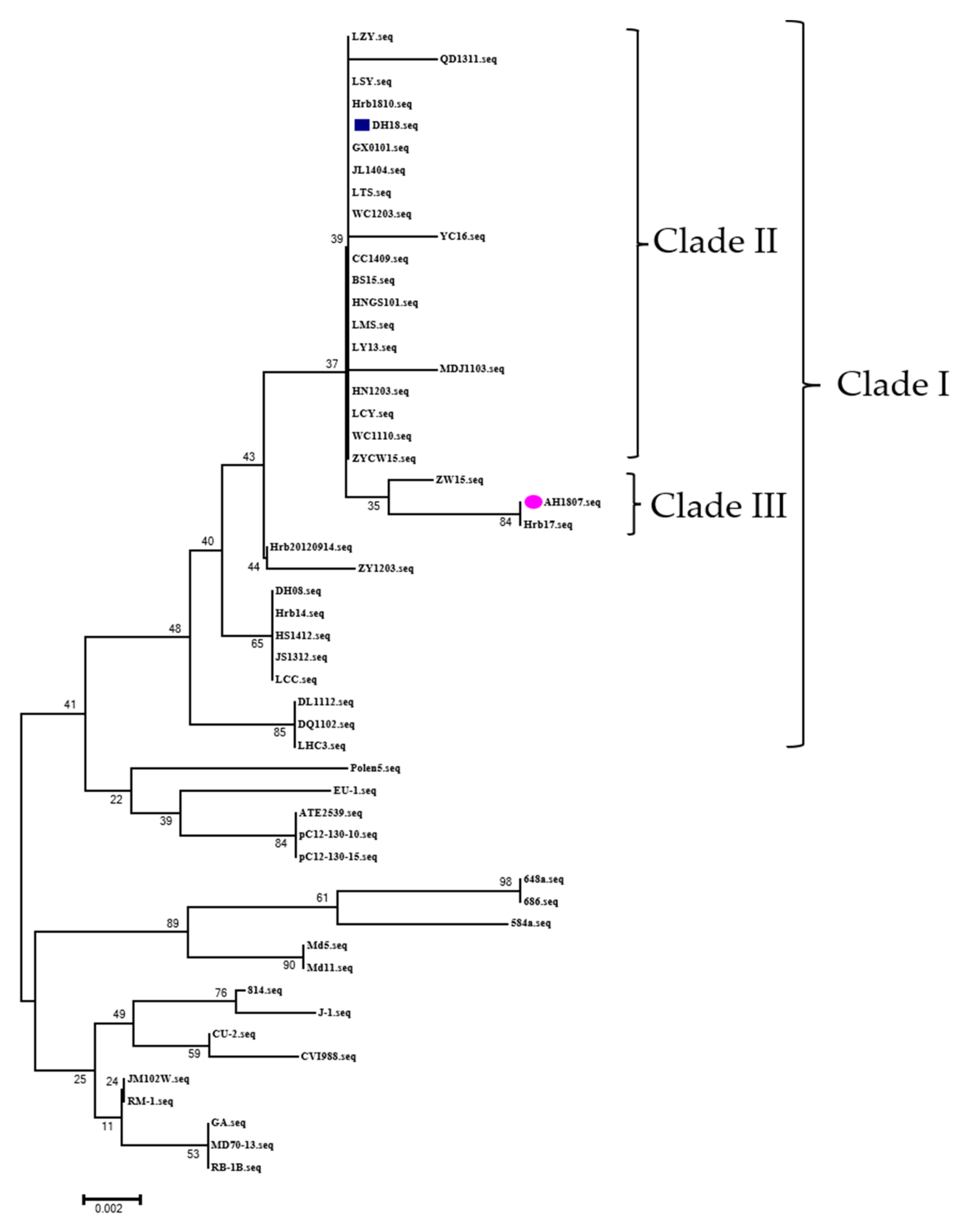
3.8. Sequence Alignment and Phylogenetic Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Gimeno, I.M.; Cortes, A.L.; Faiz, N.M.; Hernandez-Ortiz, B.A.; Guy, J.S.; Hunt, H.D.; Silva, R.F. Evaluation of the Protection Efficacy of a Serotype 1 Marek’s Disease Virus-Vectored Bivalent Vaccine Against Infectious Laryngotracheitis and Marek’s Disease. Avian Dis. 2015, 59, 255–262. [Google Scholar] [CrossRef]
- Churchill, A.E.; Biggs, P.M. Agent of Marek’s Disease in Tissue Culture. Nature 1967, 215, 528–530. [Google Scholar] [CrossRef] [PubMed]
- Nair, V. Evolution of Marek’s disease—A paradigm for incessant race between the pathogen and the host. Vet. J. 2005, 170, 175–183. [Google Scholar] [CrossRef]
- Witter, R.L.; Calnek, B.W.; Buscaglia, C.; Gimeno, I.; Schat, K.A. Classification of Marek’s disease viruses according to pathotype: Philosophy and methodology. Avian Pathol. 2005, 34, 75–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teng, M.; Zheng, L.-P.; Li, H.-Z.; Ma, S.-M.; Zhu, Z.-J.; Chai, S.-J.; Yao, Y.; Nair, V.; Zhang, G.-P.; Luo, J. Pathogenicity and Pathotype Analysis of Henan Isolates of Marek’s Disease Virus Reveal Long-Term Circulation of Highly Virulent MDV Variant in China. Viruses 2022, 14, 1651. [Google Scholar] [CrossRef]
- Gimeno, I.M. Marek’s disease vaccines: A solution for today but a worry for tomorrow? Vaccine 2008, 26, C31–C41. [Google Scholar] [CrossRef]
- Morrow, C.; Fehler, F. 5-Marek’s disease: A worldwide problem. In Marek’s Disease; Davison, F., Nair, V., Eds.; Academic Press: Oxford, UK, 2004; pp. 49–61. [Google Scholar] [CrossRef]
- Davison, F.; Nair, V. Use of Marek’s disease vaccines: Could they be driving the virus to increasing virulence? Expert Rev. Vaccines 2005, 4, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, D.A.; Read, A.F. Why the evolution of vaccine resistance is less of a concern than the evolution of drug resistance. Proc. Natl. Acad. Sci. USA 2018, 115, 12878–12886. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Lan, X.; Wang, Y.; Lin, Y.; Yu, Z.; Guo, R.; Li, K.; Cui, H.; Qi, X.; Wang, Y.; et al. Emerging natural recombinant Marek’s disease virus between vaccine and virulence strains and their pathogenicity. Transbound. Emerg. Dis. 2022, 69, e1702–e1709. [Google Scholar] [CrossRef]
- Yu, Z.; Zhang, Y.; Lan, X.; Wang, Y.; Zhang, F.; Gao, Y.; Li, K.; Gao, L.; Pan, Q.; Qi, X.; et al. Natural co-infection with two virulent wild strains of Marek’s disease virus in a commercial layer flock. Vet. Microbiol. 2019, 240, 108501. [Google Scholar] [CrossRef]
- Li, K.; Yu, Z.; Lan, X.; Wang, Y.; Qi, X.; Cui, H.; Gao, L.; Wang, X.; Zhang, Y.; Gao, Y.; et al. Complete genome analysis reveals evolutionary history and temporal dynamics of Marek’s disease virus. Front. Microbiol. 2022, 13, 1046832. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Ge, Z.; Luo, Q.; Fu, Q.; Chen, R. A highly pathogenic Marek’s disease virus isolate from chickens immunized with a bivalent vaccine in China. Arch. Virol. 2022, 167, 861–870. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.-R.; Zhang, Y.-P.; Lv, H.-C.; Zhou, L.-Y.; Cui, H.-Y.; Gao, Y.-L.; Qi, X.-L.; Wang, Y.-Q.; Li, K.; Gao, L.; et al. A Chinese Variant Marek’s Disease Virus Strain with Divergence between Virulence and Vaccine Resistance. Viruses 2017, 9, 71. [Google Scholar] [CrossRef] [Green Version]
- Teng, L.-Q.; Wei, P.; Song, Z.-B.; Yang, N.-L. Evaluation of the pathogenicity of a field isolate of Marek’s disease virus integrated with retroviral long terminal repeat sequence. Bing du xue bao = Chin. J. Virol. 2009, 25, 376–381. [Google Scholar]
- Zhang, Y.-P.; Li, Z.-J.; Bao, K.-Y.; Lv, H.-C.; Gao, Y.-L.; Gao, H.-L.; Qi, X.-L.; Cui, H.-Y.; Wang, Y.-Q.; Ren, X.-G.; et al. Pathogenic characteristics of Marek’s disease virus field strains prevalent in China and the effectiveness of existing vaccines against them. Vet. Microbiol. 2015, 177, 62–68. [Google Scholar] [CrossRef]
- Lv, H.C.; Zhang, Y.P.; Sun, G.R.; Gao, Y.L.; Li, Z.J.; Zheng, H.W.; Bao, K.Y.; Wang, X.M.; Liu, C.J. Assessments of a PCR method for detection and identification of virulent Marek’s disease virus and vaccine strain for Marek’s disease diagnosis. Chin. J. Prev. Vet. Med. 2016, 38, 567–571. [Google Scholar]
- Li, H.; Wang, P.; Lin, L.; Shi, M.; Gu, Z.; Huang, T.; Mo, M.; Wei, T.; Zhang, H.; Wei, P. The emergence of the infection of subgroup J avian leucosis virus escalated the tumour incidence in commercial Yellow chickens in Southern China in recent years. Transbound. Emerg. Dis. 2018, 66, 312–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishitha, Y.; Priyanka, E.; Krishna, S.V.; Kannaki, T.R. Co-infection of Marek’s disease virus with different oncogenic immunosuppressive viruses in chicken flocks. Virusdisease 2021, 32, 804–809. [Google Scholar] [CrossRef]
- Haridy, M.; Goryo, M.; Sasaki, J.; Okada, K. Pathological and immunohistochemical study of chickens with co-infection of Marek’s disease virus and chicken anaemia virus. Avian Pathol. 2009, 38, 469–483. [Google Scholar] [CrossRef]
- Gopal, S.; Manoharan, P.; Kathaperumal, K.; Chidambaram, B.; Divya, K.C. Differential Detection of Avian Oncogenic Viruses in Poultry Layer Farms and Turkeys by Use of Multiplex PCR. J. Clin. Microbiol. 2012, 50, 2668–2673. [Google Scholar] [CrossRef] [Green Version]
- Qin, L.T.; Gao, Y.L.; Pan, W.; Deng, X.Y.; Sun, F.F.; Li, K.; Qi, X.-G.L.; Gao, H.L.; Liu, C.N.; Wang, X.M. Investigation of co-infection of ALV-J with REV, MDV, CAV in layer chicken flocks in some regions of China. Chin. J. Prev. Vet. Med. 2010, 32, 90–93. [Google Scholar]
- Witter, R.L.; Kreager, K.S. Serotype 1 viruses modified by backpassage or insertional mutagenesis: Approaching the threshold of vaccine efficacy in Marek’s disease. Avian Dis. 2004, 48, 768–782. [Google Scholar] [CrossRef]
- Baigent, S.J.; Petherbridge, L.J.; Howes, K.; Smith, L.P.; Currie, R.J.; Nair, V.K. Absolute quantitation of Marek’s disease virus genome copy number in chicken feather and lymphocyte samples using real-time PCR. J. Virol. Methods 2005, 123, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.; Zhang, L.; Wang, J.; Chen, L.; Shan, H.; Wang, Z.; Ma, H. Isolation and analysis of a very virulent Marek’s disease virus strain in China. Virol. J. 2013, 10, 155–158. [Google Scholar] [CrossRef] [Green Version]
- Sharma, J.M.; Burmester, B.R. Resistance to Marek’s disease at hatching in chickens vaccinated as embryos with the turkey herpesvirus. Avian Dis. 1982, 26, 134–149. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-P.; Liu, C.-J.; Zhang, F.; Shi, W.; Li, J. Sequence analysis of the Meq gene in the predominant Marek’s disease virus strains isolated in China during 2006–2008. Virus Genes 2011, 43, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Deng, Q.; Shi, M.; Li, Q.; Wang, P.; Li, M.; Wang, W.; Gao, Y.; Li, H.; Lin, L.; Huang, T.; et al. Analysis of the evolution and transmission dynamics of the field MDV in China during the years 1995–2020, indicating the emergence of a unique cluster with the molecular characteristics of vv+ MDV that has become endemic in southern China. Transbound. Emerg. Dis. 2020, 68, 3574–3587. [Google Scholar] [CrossRef]
- Li, K.; Liu, Y.; Xu, Z.; Zhang, Y.; Luo, D.; Gao, Y.; Qian, Y.; Bao, C.; Liu, C.; Zhang, Y.; et al. Avian oncogenic herpesvirus antagonizes the cGAS-STING DNA-sensing pathway to mediate immune evasion. PLoS Pathog. 2019, 15, e1007999. [Google Scholar] [CrossRef] [Green Version]
- Du, X.; Zhou, D.; Zhou, J.; Xue, J.; Wang, G.; Cheng, Z. Marek’s disease virus serine/threonine kinase Us3 facilitates viral replication by targeting IRF7 to block IFN-β production. Veter-Microbiol. 2022, 266. [Google Scholar] [CrossRef]
- Couteaudier, M.; Denesvre, C. Marek’s disease virus and skin interactions. Vet. Res. 2014, 45, 36. [Google Scholar] [CrossRef] [Green Version]
- Conradie, A.M.; Bertzbach, L.D.; Trimpert, J.; Patria, J.N.; Murata, S.; Parcells, M.S.; Kaufer, B.B. Distinct polymorphisms in a single herpesvirus gene are capable of enhancing virulence and mediating vaccinal resistance. PLoS Pathog. 2020, 16, e1009104. [Google Scholar] [CrossRef]
- Spatz, S.J.; Petherbridge, L.; Zhao, Y.; Nair, V. Comparative full-length sequence analysis of oncogenic and vaccine (Rispens) strains of Marek’s disease virus. J. Gen. Virol. 2007, 88, 1080–1096. [Google Scholar] [CrossRef]
- Cui, N.; Su, S.; Sun, P.; Zhang, Y.; Han, N.; Cui, Z. Isolation and pathogenic analysis of virulent Marek’s disease virus field strain in China. Poult. Sci. 2016, 95, 1521–1528. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-P.; Lv, H.-C.; Bao, K.-Y.; Gao, Y.-L.; Gao, H.-L.; Qi, X.-L.; Cui, H.-Y.; Wang, Y.-Q.; Li, K.; Gao, L.; et al. Molecular and pathogenicity characterization of Gallid herpesvirus 2 newly isolated in China from 2009 to 2013. Virus Genes 2015, 52, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Beachboard, D.C.; Horner, S.M. Innate immune evasion strategies of DNA and RNA viruses. Curr. Opin. Microbiol. 2016, 32, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.J.; Ashkar, A.A. The Dual Nature of Type I and Type II Interferons. Front. Immunol. 2018, 9, 2061. [Google Scholar] [CrossRef] [Green Version]
- Bertzbach, L.D.; Harlin, O.; Härtle, S.; Fehler, F.; Vychodil, T.; Kaufer, B.B.; Kaspers, B. IFNα and IFNγ Impede Marek’s Disease Progression. Viruses 2019, 11, 1103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Gao, L.; Xu, Z.; Luo, D.; Zhang, Y.; Gao, Y.; Liu, C.; Zhang, Y.; Qi, X.; Cui, H.; et al. Marek’s Disease Virus RLORF4 Inhibits Type I Interferon Production by Antagonizing NF-κB Activation. J. Virol. 2019, 93, e01037-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, G.-R.; Zhou, L.-Y.; Zhang, Y.-P.; Zhang, F.; Yu, Z.-H.; Pan, Q.; Gao, L.; Li, K.; Wang, Y.-Q.; Cui, H.-Y.; et al. Differential expression of type I interferon mRNA and protein levels induced by virulent Marek’s disease virus infection in chickens. Vet. Immunol. Immunopathol. 2019, 212, 15–22. [Google Scholar] [CrossRef]
- Gimeno, I.M.; Glaize, A.; Cortes, A.L. Effect of Marek’s disease vaccines on interferon and toll like receptors when administered in ovo. Vet. Immunol. Immunopathol. 2018, 201, 62–66. [Google Scholar] [CrossRef]
- Jin, H.; Kong, Z.; Mehboob, A.; Jiang, B.; Xu, J.; Cai, Y.; Liu, W.; Hong, J.; Li, Y. Transcriptional Profiles Associated with Marek’s Disease Virus in Bursa and Spleen Lymphocytes Reveal Contrasting Immune Responses during Early Cytolytic Infection. Viruses 2020, 12, 354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kano, R.; Konnai, S.; Onuma, M.; Ohashi, K. Cytokine profiles in chickens infected with virulent and avirulent Marek’s disease viruses: Interferon-gamma is a key factor in the protection of Marek’s disease by vaccination. Microbiol. Immunol. 2009, 53, 224–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boodhoo, N.; Behboudi, S. Differential Virus-Specific IFN-Gamma Producing T Cell Responses to Marek’s Disease Virus in Chickens With B19 and B21 MHC Haplotypes. Front. Immunol. 2022, 12. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | No. of Chickens | Vaccine | Challenge |
|---|---|---|---|
| 1 | 36 | - | - |
| 2 | 18 | - | AH/1807 |
| 3 | 18 | - | AH/1807 |
| 4 | 18 | CVI988 | AH/1807 |
| 5 | 18 | CVI988 | AH/1807 |
| 6 | 18 | - | DH/18 |
| 7 | 18 | - | DH/18 |
| 8 | 18 | CVI988 | DH/18 |
| 9 | 18 | CVI988 | DH/18 |
| Gene | Type | Sequence (5′–3′) | Amplicon Size (bp) |
|---|---|---|---|
| meq | Forward Reverse Probe | GGAGCCGGAGAGGCTTTATG ATCTGGCCCGAATACAAGGAA CGTCTTACCGAGGATCCCGAACAGG | 69 |
| ovo | Forward Reverse Probe | CACTGCCACTGGGCTCTGT GCAATGGCAATAAACCTCCAA AGTCTGGAGAAGTCTGTGCAGCCTCCA | 71 |
| IFN-γ | Forward Reverse Probe | TACTGAGCCAGATTGTTTCGAT TCACCTTCTTCACGCCAT AAGTCAAAGCCGCACATCAAACAC | 132 |
| IFN-β | Forward Reverse Probe | CACAACAAGACGTGACTTTTCCATTT AAGCATGTTGAAGAGGTGTTGGAG AGGACAAGAAGCAAGCAGCCATCACCACC | 110 |
| 28S | Forward Reverse Probe | GGCGAAGCCAGAGGAAACT GACGACCGATTTGCACGTC AGGACCGCTACGGACCTCCACCA | 62 |
| Group | Vaccine | Challenge | MD Incidence | PI | Mortality | Tumor Incidence | Time (dpc) |
|---|---|---|---|---|---|---|---|
| 1 | - | - | - | - | - | - | |
| 3 | - | AH/1807 | 18/18(100%) | - | 14/18(77.8%) | 9/18(50%) | 72 |
| 5 | CVI988 | AH/1807 | 1/17(5.9%) | 94.1 | 1/17(5.9%) | 0% | 72 |
| 7 | - | DH/18 | 18/18(100%) | - | 9/18(50%) | 6/18(33.3%) | 72 |
| 9 | CVI988 | DH/18 | 7/18(38.9%) | 61.1 | 0/18(0%) | 0% | 72 |
| Strains | 77 | 80 | 88 | 93 | 115 | 139 | 176 | 217 |
|---|---|---|---|---|---|---|---|---|
| GA | K | D | A | Q | V | T | P | P |
| AH/1807 | E | Y | T | R | A | A | R | A |
| DH/18 | E | Y | A | Q | A | A | R | A |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Z.-H.; Zhang, Y.-P.; Lan, X.-G.; Wang, Y.-N.; Guo, R.-R.; Li, K.; Gao, L.; Qi, X.-L.; Cui, H.-Y.; Wang, X.-M.; et al. Differences in Pathogenicity and Vaccine Resistance Discovered between Two Epidemic Strains of Marek’s Disease Virus in China. Viruses 2023, 15, 945. https://doi.org/10.3390/v15040945
Yu Z-H, Zhang Y-P, Lan X-G, Wang Y-N, Guo R-R, Li K, Gao L, Qi X-L, Cui H-Y, Wang X-M, et al. Differences in Pathogenicity and Vaccine Resistance Discovered between Two Epidemic Strains of Marek’s Disease Virus in China. Viruses. 2023; 15(4):945. https://doi.org/10.3390/v15040945
Chicago/Turabian StyleYu, Zheng-Hao, Yan-Ping Zhang, Xing-Ge Lan, Ya-Nan Wang, Rong-Rong Guo, Kai Li, Li Gao, Xiao-Le Qi, Hong-Yu Cui, Xiao-Mei Wang, and et al. 2023. "Differences in Pathogenicity and Vaccine Resistance Discovered between Two Epidemic Strains of Marek’s Disease Virus in China" Viruses 15, no. 4: 945. https://doi.org/10.3390/v15040945