
The Divergent Pattern of SARS-CoV-2 Variant Predominance and Transmission Dynamics in the Brazilian Island of Ilhabela

 , , , , , , , ,
, , , , , , , ,  , , , , and
, , , , and 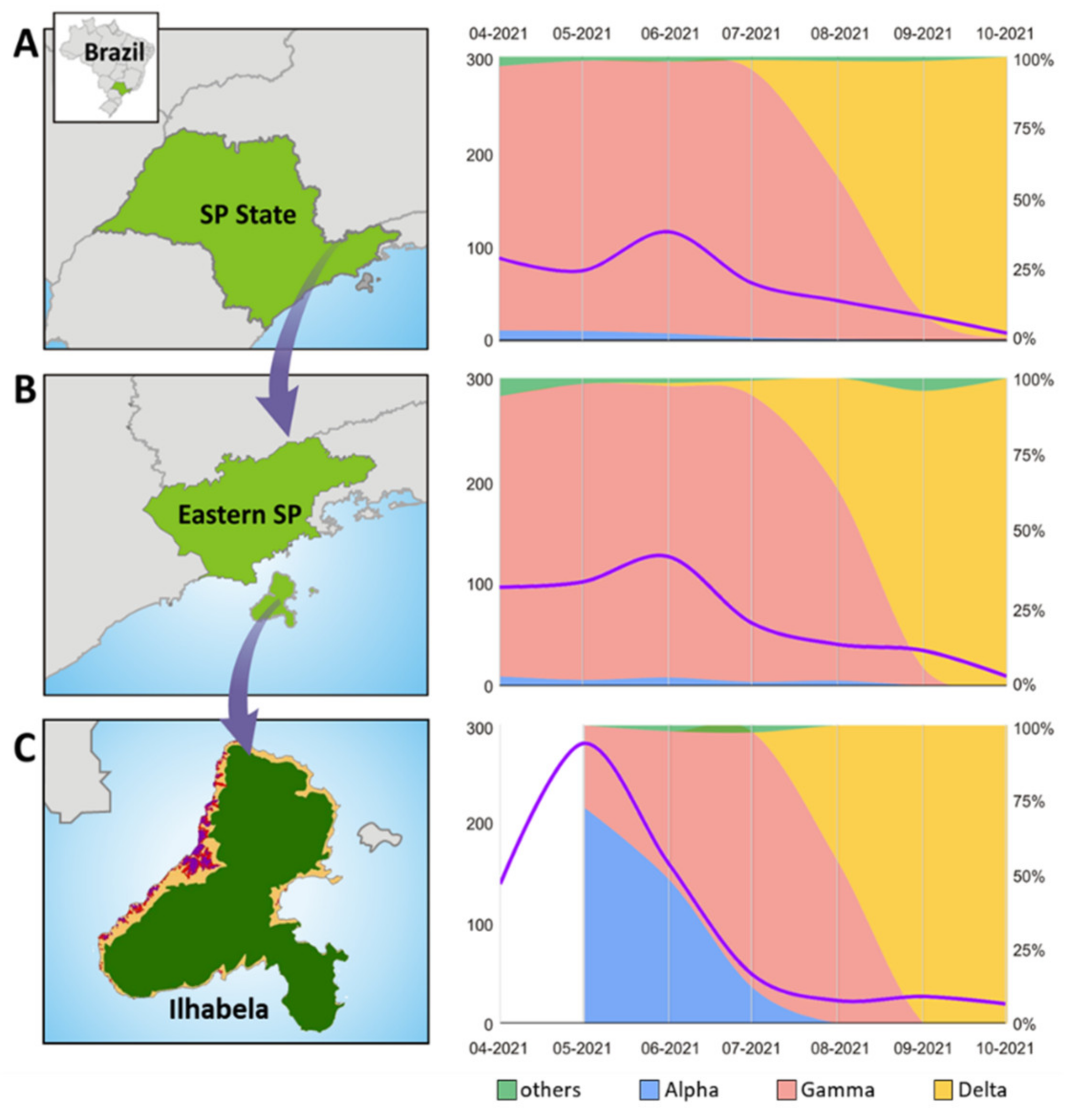{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling, Sample Preparation and Next-Generation Sequencing
2.2. SARS-CoV-2 Whole-Genome Assembly and Lineage Identification
2.3. Phylogenetic Analysis
2.4. COVID-19 Incidence Dataset
3. Results
3.1. SARS-CoV-2 VOC Predominance and Lineage Replacement on Ilhabela Island
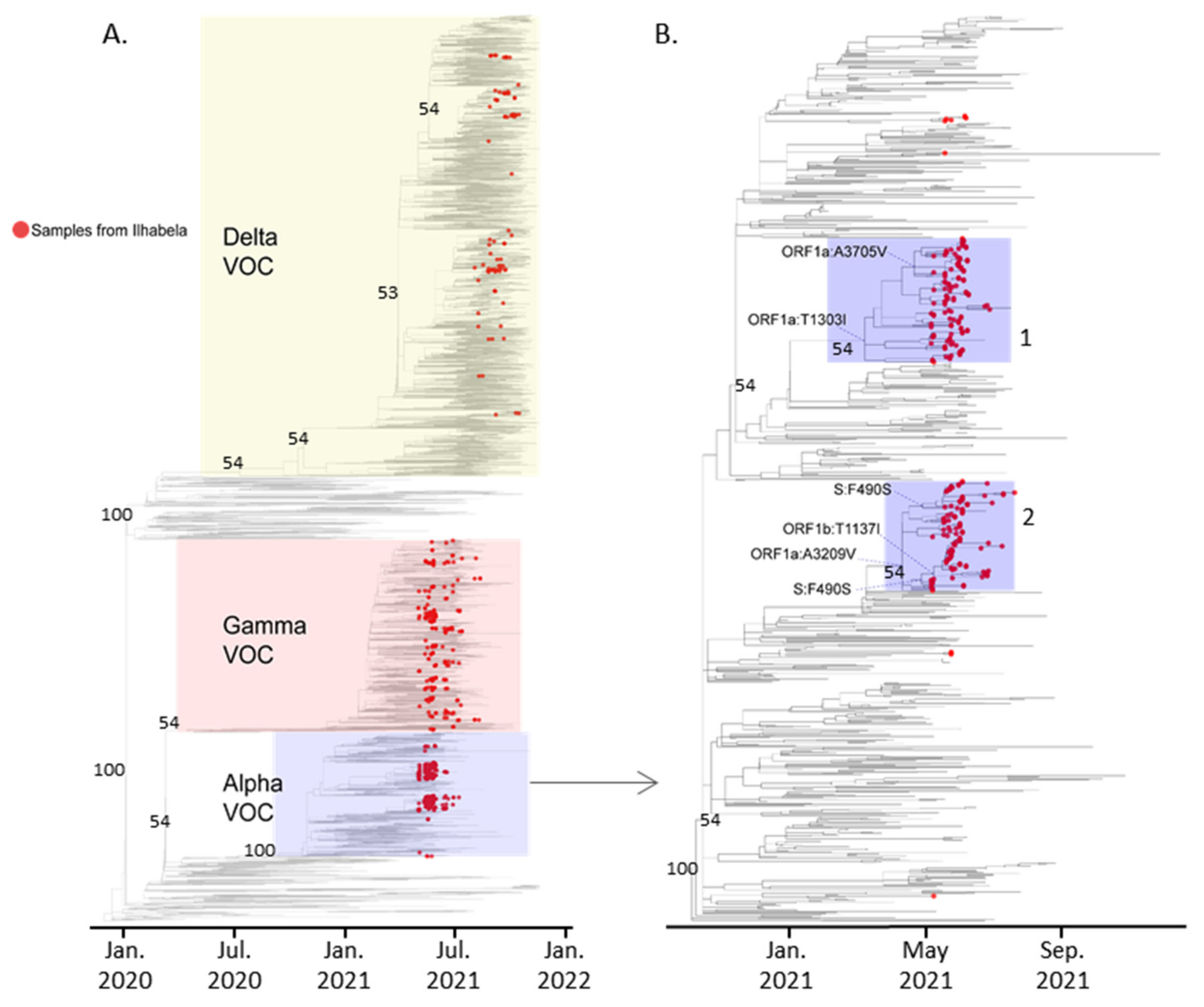
3.2. Phylogenetic Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Buss, L.F.; Prete, C.A.; Abrahim, C.M.M.; Mendrone, A.; Salomon, T.; de Almeida-Neto, C.; França, R.F.O.; Belotti, M.C.; Carvalho, M.P.S.S.; Costa, A.G.; et al. Three-Quarters Attack Rate of SARS-CoV-2 in the Brazilian Amazon during a Largely Unmitigated Epidemic. Science 2021, 371, 288–292. [Google Scholar] [CrossRef] [PubMed]
- Giovanetti, M.; Fonseca, V.; Wilkinson, E.; Tegally, H.; San, E.J.; Althaus, C.L.; Xavier, J.; Slavov, S.N.; Viala, V.L.; Lima, A.R.J.; et al. Replacement of the Gamma by the Delta Variant in Brazil: Impact of Lineage Displacement on the Ongoing Pandemic. medRxiv 2021, 2021, 21268309. [Google Scholar] [CrossRef] [PubMed]
- Naveca, F.G.; Nascimento, V.; de Souza, V.C.; de Corado, A.L.; Nascimento, F.; Silva, G.; Costa, Á.; Duarte, D.; Pessoa, K.; Mejía, M.; et al. COVID-19 in Amazonas, Brazil, Was Driven by the Persistence of Endemic Lineages and P.1 Emergence. Nat. Med. 2021, 27, 1230–1238. [Google Scholar] [CrossRef] [PubMed]
- Slavov, S.N.; Patané, J.S.L.; Bezerra, R.D.S.; Giovanetti, M.; Fonseca, V.; Martins, A.J.; Viala, V.L.; Rodrigues, E.S.; Santos, E.V.; Barros, C.R.S.; et al. Genomic Monitoring Unveil the Early Detection of the SARS-CoV-2 B.1.351 (Beta) Variant (20H/501Y.V2) in Brazil. J. Med. Virol. 2021, 93, 6782–6787. [Google Scholar] [CrossRef]
- Kashima, S.; Slavov, S.N.; Giovanetti, M.; Rodrigues, E.S.; Patané, J.S.L.; Viala, V.L.; Santos, E.V.; Evaristo, M.; Lima, L.P.O.; Martins, A.J.; et al. Introduction of SARS-CoV-2 C.37 (WHO VOI Lambda) in the Sao Paulo State, Southeast Brazil. J. Med. Virol. 2022, 94, 1206–1211. [Google Scholar] [CrossRef]
- Patané, J.; Viala, V.; Lima, L.; Martins, A.; Barros, C.; Marqueze, E.; Bernardino, J.; Moretti, D.; Slavov, S.; Bezerra, R.; et al. SARS-CoV-2 Delta Variant of Concern in Brazil—Multiple Introductions, Communitary Transmission, and Early Signs of Local Evolution. medRxiv 2021, 2021, 21262846. [Google Scholar] [CrossRef]
- Tyson, J.R.; James, P.; Stoddart, D.; Sparks, N.; Wickenhagen, A.; Hall, G.; Choi, J.H.; Lapointe, H.; Kamelian, K.; Smith, A.D.; et al. Improvements to the ARTIC Multiplex PCR Method for SARS-CoV-2 Genome Sequencing Using Nanopore. bioRxiv 2020, 3, 283077. [Google Scholar] [CrossRef]
- Andrews, S. Babraham Bioinformatics—FastQC A Quality Control Tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 25 January 2022).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Li, H. Aligning Sequence Reads, Clone Sequences and Assembly Contigs with BWA-MEM. arXiv 2013, arXiv:1303.3997. [Google Scholar]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map Format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; et al. Pilon: An Integrated Tool for Comprehensive Microbial Variant Detection and Genome Assembly Improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef] [PubMed]
- Narasimhan, V.; Danecek, P.; Scally, A.; Xue, Y.; Tyler-Smith, C.; Durbin, R. BCFtools/RoH: A Hidden Markov Model Approach for Detecting Autozygosity from next-Generation Sequencing Data. Bioinformatics 2016, 32, 1749–1751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- GitHub—Lh3/Seqtk: Toolkit for Processing Sequences in FASTA/Q Formats. Available online: https://github.com/lh3/seqtk (accessed on 11 January 2022).
- O’Toole, Á.; Kraemer, M.U.G.; Hill, V.; Pybus, O.G.; Watts, A.; Bogoch, I.I.; Khan, K.; Messina, J.P.; Tegally, H.; Lessells, R.R.; et al. Tracking the International Spread of SARS-CoV-2 Lineages B.1.1.7 and B.1.351/501Y-V2. Wellcome Open Res. 2021, 6, 121. [Google Scholar] [CrossRef]
- Shu, Y.; McCauley, J. GISAID: Global Initiative on Sharing All Influenza Data—From Vision to Reality. Eurosurveillance 2017, 22, 30494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aksamentov, I.; Roemer, C.; Hodcroft, E.B.; Neher, R.A. Nextclade: Clade Assignment, Mutation Calling and Quality Control for Viral Genomes. J. Open Source Softw. 2021, 6, 3773. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Larsson, A. AliView: A Fast and Lightweight Alignment Viewer and Editor for Large Datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Cordeiro, T.L.; Pereira, C.S.; Novaes, R.A., Jr. Análise Da Expansão Urbana e Das Variáveis Climatológicas Do Município de Ilhabela—SP. 2010. Available online: http://mtc-m16d.sid.inpe.br/col/sid.inpe.br/mtc-m19/2010/09.22.18.40/doc/Thiago%20Lob%E3o%20Cordeiro.pdf (accessed on 10 December 2021).
- Slavov, S.N.; dos Bezerra, R.S.; Rodrigues, E.S.; Santos, E.V.; Borges, J.S.; de la Roque, D.G.L.; Patané, J.S.L.; Lima, A.R.J.; Ribeiro, G.; Viala, V.L.; et al. Genomic monitoring of the SARS-CoV-2 b.1.1.7 (who voc alpha) in the sao paulo state, Brazil. Virus Res. 2021, 308, 198643. [Google Scholar] [CrossRef]
- Chrysostomou, A.C.; Vrancken, B.; Koumbaris, G.; Themistokleous, G.; Aristokleous, A.; Masia, C.; Eleftheriou, C.; Ioannou, C.; Stylianou, D.C.; Ioannides, M.; et al. A Comprehensive Molecular Epidemiological Analysis of SARS-CoV-2 Infection in Cyprus from April 2020 to January 2021: Evidence of a Highly Polyphyletic and Evolving Epidemic. Viruses 2021, 13, 1098. [Google Scholar] [CrossRef]
- Richter, J.; Fanis, P.; Tryfonos, C.; Koptides, D.; Krashias, G.; Bashiardes, S.; Hadjisavvas, A.; Loizidou, M.; Oulas, A.; Alexandrou, D.; et al. Molecular Epidemiology of SARS-CoV-2 in Cyprus. PLoS ONE 2021, 16, e0248792. [Google Scholar] [CrossRef] [PubMed]
- Cazelles, B.; Nguyen-Van-Yen, B.; Champagne, C.; Comiskey, C. Dynamics of the COVID-19 Epidemic in Ireland under Mitigation. BMC Infect. Dis. 2021, 21, 735. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Viala, V.L.; Slavov, S.N.; de Lima, L.P.O.; Lima, A.R.J.; Ribeiro, G.; Martins, A.J.; Petry, B.; Banho, C.A.; Barros, C.R.d.S.; Moncau, C.T.; et al. The Divergent Pattern of SARS-CoV-2 Variant Predominance and Transmission Dynamics in the Brazilian Island of Ilhabela. Viruses 2022, 14, 1481. https://doi.org/10.3390/v14071481
Viala VL, Slavov SN, de Lima LPO, Lima ARJ, Ribeiro G, Martins AJ, Petry B, Banho CA, Barros CRdS, Moncau CT, et al. The Divergent Pattern of SARS-CoV-2 Variant Predominance and Transmission Dynamics in the Brazilian Island of Ilhabela. Viruses. 2022; 14(7):1481. https://doi.org/10.3390/v14071481
Chicago/Turabian StyleViala, Vincent Louis, Svetoslav Nanev Slavov, Loyze Paola Oliveira de Lima, Alex Ranieri Jeronimo Lima, Gabriela Ribeiro, Antonio Jorge Martins, Bruna Petry, Cecilia Artico Banho, Claudia Renata dos Santos Barros, Cristina Tschorny Moncau, and et al. 2022. "The Divergent Pattern of SARS-CoV-2 Variant Predominance and Transmission Dynamics in the Brazilian Island of Ilhabela" Viruses 14, no. 7: 1481. https://doi.org/10.3390/v14071481