
Interferon-Induced Protein 6-16 (IFI6-16) from Litopenaeus vannamei Regulate Antiviral Immunity via Apoptosis-Related Genes

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cloning of Full Length of LvIFI6-16 cDNA
2.2. Sequence and Phylogenetic Analysis of LvIFI6-16
2.3. Confocal Laser Scanning Microscopy
2.4. The Quantitative RT-PCR Analysis of LvIFI6-16 Expression
2.5. Detection of Expression Levels of LvIFI6-16 and Apoptosis-Associated Genes in LvIFI6-16 Silenced L. vannamei
2.6. WSSV and PBS Challenge Experiments in LvIFI6-16-Knockdown Shrimp
3. Results
3.1. Sequence Features of LvIFI6-16
3.2. Phylogenetic Analysis
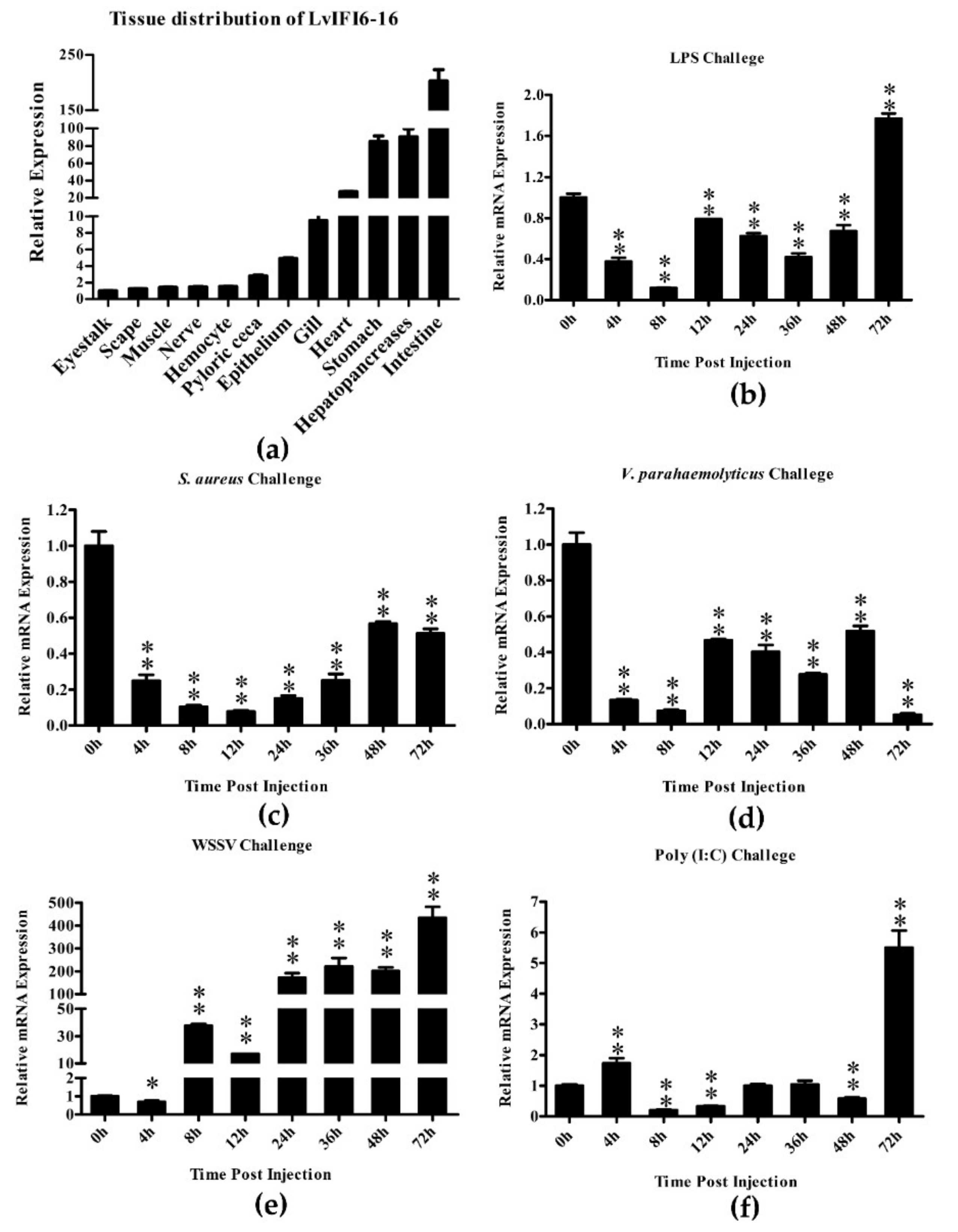
3.3. Expression Levels of LvIFI6-16 mRNA in Healthy and Immune-Challenged Shrimps
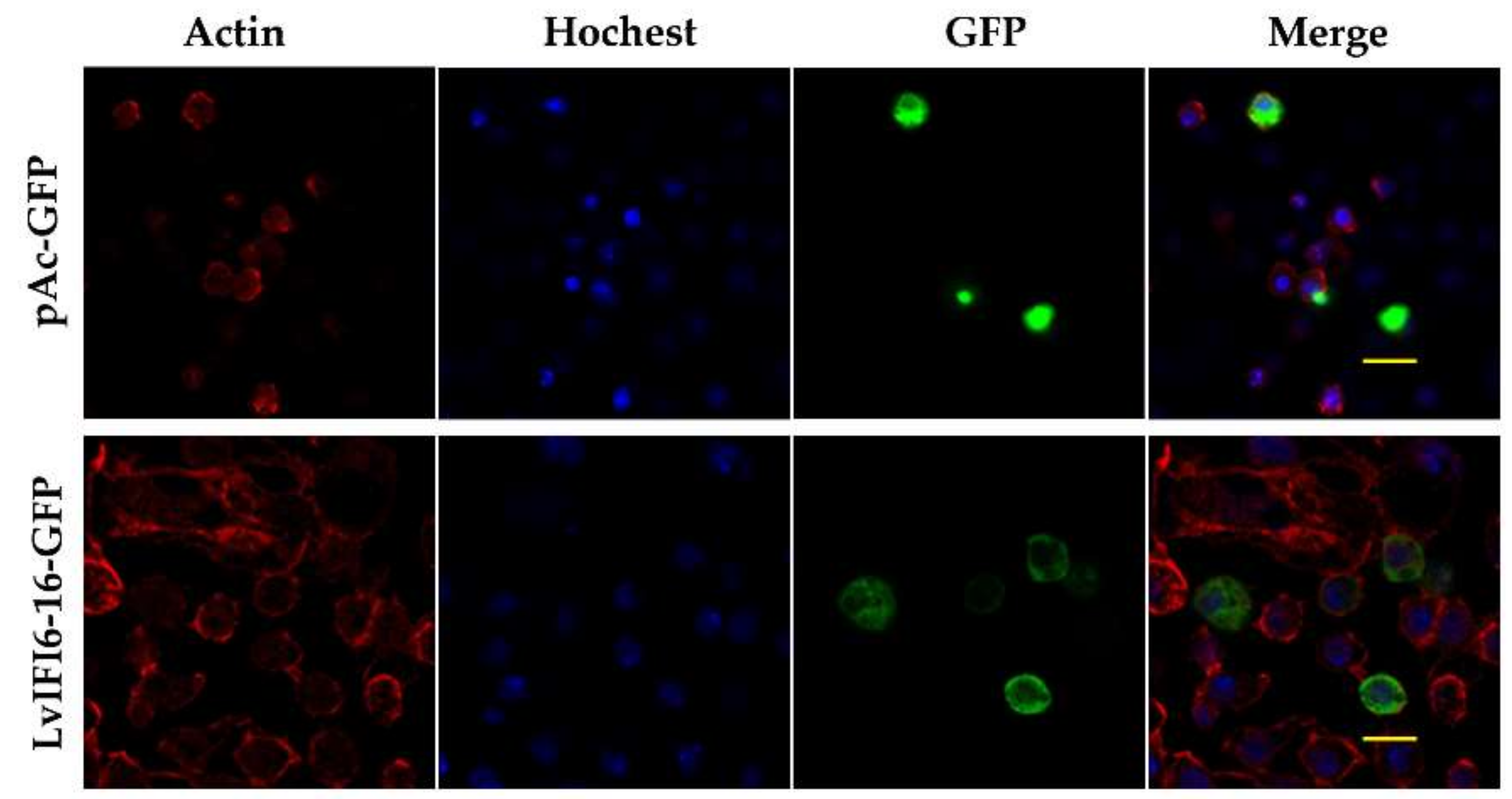
3.4. Subcellular Localization of LvIFI6-16
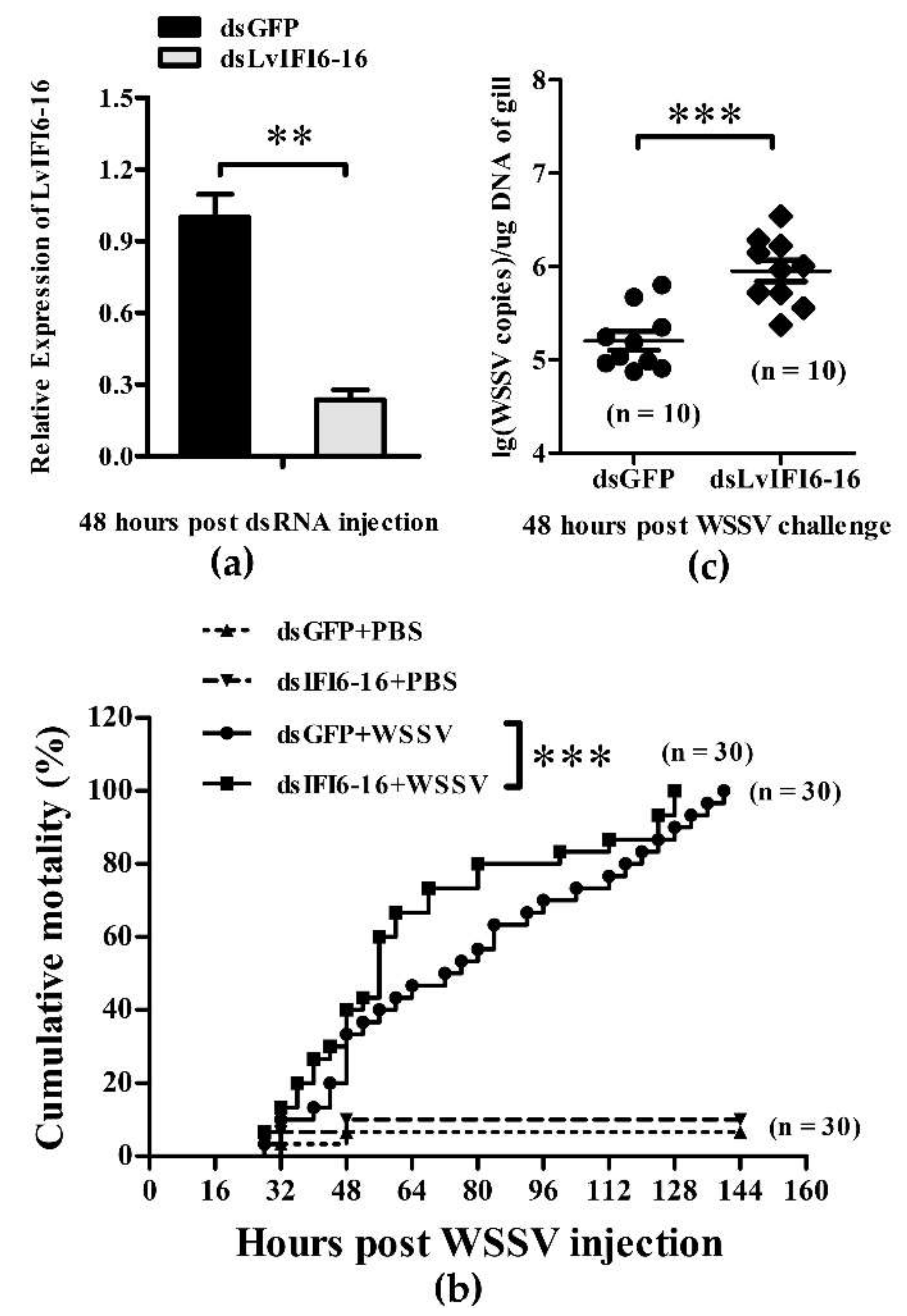
3.5. LvIFI6-16 Played a Key Role in Defense against WSSV
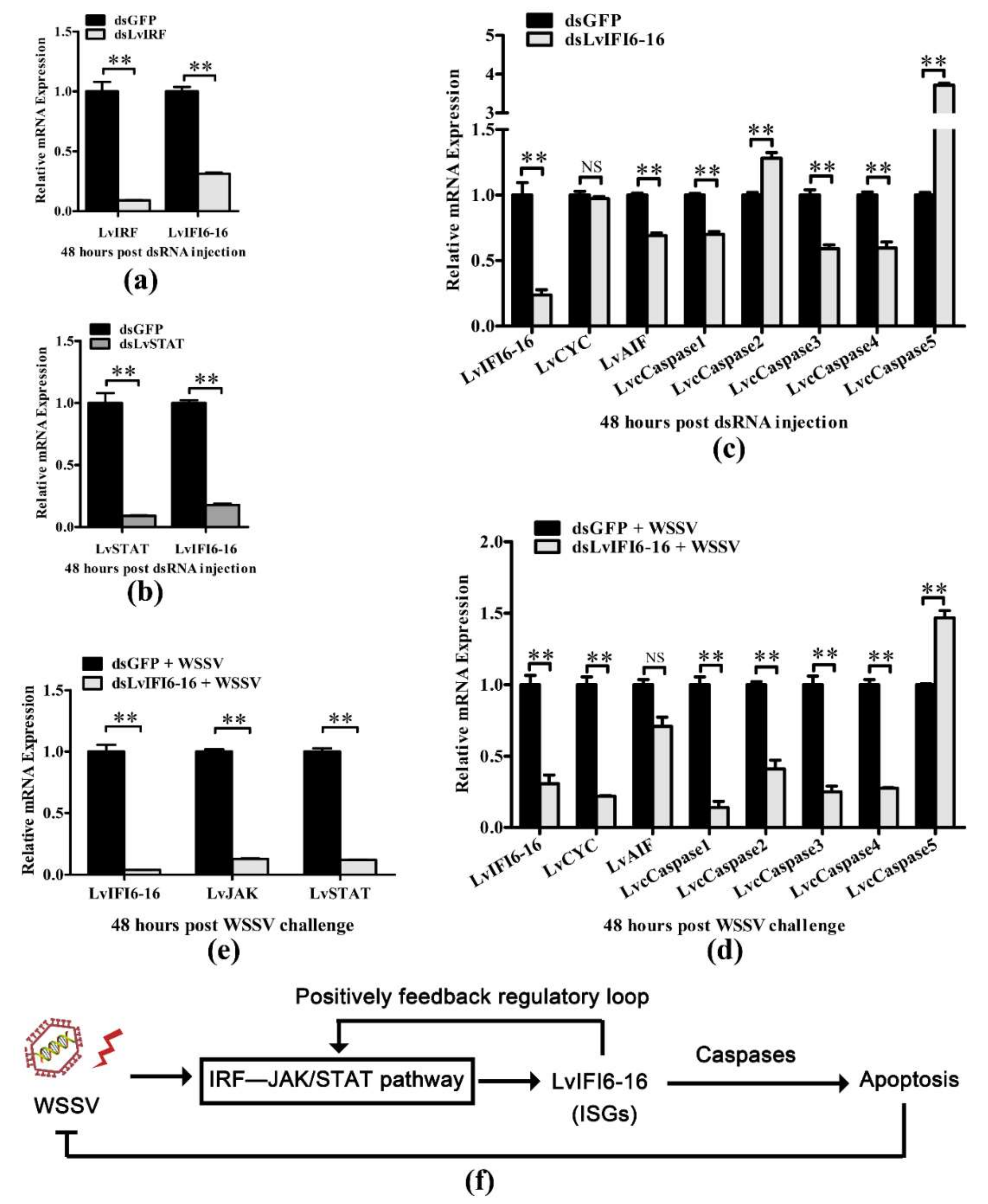
3.6. LvIFI6-16 Regulated Apoptosis-Related Genes with or without WSSV Challenge
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schneider, W.M.; Chevillotte, M.D.; Rice, C.M. Interferon-Stimulated Genes: A Complex Web of Host Defenses. Annu. Rev. Immunol. 2014, 32, 513–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stark, G.R.; Darnell, J.E., Jr. The JAK-STAT Pathway at Twenty. Immunity 2012, 36, 503–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Navajas, J.M.; Lee, J.; David, M.; Raz, E. Immunomodulatory functions of type I interferons. Nat. Rev. Immunol. 2012, 12, 125–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, J.; Porter, A.; Chernajovsky, Y.; Gilbert, C.; Stark, G.; Kerr, I. Characterization of a human gene inducible by alpha- and beta-interferons and its expression in mouse cells. EMBO J. 1986, 5, 1601–1606. [Google Scholar] [CrossRef]
- Cheriyath, V.; Leaman, D.W.; Borden, E.C. Emerging roles of FAM14 family members (G1P3/ISG 6-16 and ISG12/IFI27) in innate immunity and cancer. J. Interferon Cytokine Res. 2011, 31, 173–181. [Google Scholar] [CrossRef]
- Cheriyath, V.; Kaur, J.; Davenport, A.; Khalel, A.; Chowdhury, N.; Gaddipati, L. G1P3 (IFI6), a mitochondrial localised antiapoptotic protein, promotes metastatic potential of breast cancer cells through mtROS. Br. J. Cancer 2018, 119, 52–64. [Google Scholar] [CrossRef]
- Qi, Y.; Li, Y.; Zhang, Y.; Zhang, L.; Wang, Z.; Zhang, X.; Gui, L.; Huang, J. IFI6 Inhibits Apoptosis via Mitochondrial-Dependent Pathway in Dengue Virus 2 Infected Vascular Endothelial Cells. PLoS ONE 2015, 10, e0132743. [Google Scholar]
- Pang, X.; Li, X.-H.; Mo, Z.; Huang, J.; Deng, H.; Lei, Z.; Zheng, X.; Feng, Z.; Xie, D.; Gao, Z. IFI16 is involved in HBV-associated acute-on-chronic liver failure inflammation. BMC Gastroenterol. 2018, 18, 61. [Google Scholar] [CrossRef]
- Chen, S.; Li, S.; Chen, L. Interferon-inducible Protein 6-16 (IFI-6-16, ISG16) promotes Hepatitis C virus replication in vitro. J. Med. Virol. 2015, 88, 109–114. [Google Scholar] [CrossRef]
- Li, C.; Weng, S.; He, J. WSSV–host interaction: Host response and immune evasion. Fish Shellfish Immunol. 2018, 84, 558–571. [Google Scholar] [CrossRef]
- Liao, X.; He, J.; Li, C. Decapod iridescent virus 1: An emerging viral pathogen in aquaculture. Rev. Aquac. 2022. [Google Scholar] [CrossRef]
- Li, C.; Wang, S.; He, J. The Two NF-κB Pathways Regulating Bacterial and WSSV Infection of Shrimp. Front. Immunol. 2019, 10, 1785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, D.; Liu, W.; Alvarez, A.; Huang, T. Cellular immune responses against viral pathogens in shrimp. Dev. Comp. Immunol. 2014, 47, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Li, H.; Chen, Y.; Chen, Y.; Wang, S.; Weng, S.-P.; Xu, X.; He, J. Activation of Vago by interferon regulatory factor (IRF) suggests an interferon system-like antiviral mechanism in shrimp. Sci. Rep. 2015, 5, 15078. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Zhong, D.; Li, G. The role of microglia in viral encephalitis: A review. J. Neuroinflamm. 2019, 16, 76. [Google Scholar] [CrossRef]
- Li, C.; Weng, S.; Chen, Y.; Yu, X.; Lü, L.; Zhang, H.; He, J.; Xu, X. Analysis of Litopenaeus vannamei Transcriptome Using the Next-Generation DNA Sequencing Technique. PLoS ONE 2012, 7, e47442. [Google Scholar] [CrossRef]
- Letunic, I.; Doerks, T.; Bork, P. SMART: Recent updates, new developments and status in 2015. Nucleic Acids Res. 2014, 43, D257–D260. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Qiu, W.; Zhang, S.; Chen, Y.-G.; Wang, P.-H.; Xu, X.-P.; Li, C.-Z.; Chen, Y.-H.; Fan, W.-Z.; Yan, H.; Weng, S.-P.; et al. Litopenaeus vannamei NF-κB is required for WSSV replication. Dev. Comp. Immunol. 2014, 45, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Xu, L.; Su, J.; Peppelenbosch, M.P.; Pan, Q. Transcriptional Regulation of Antiviral Interferon-Stimulated Genes. Trends Microbiol. 2017, 25, 573–584. [Google Scholar] [CrossRef] [PubMed]
- Qiao, X.; Zong, Y.; Liu, Z.; Wu, Z.; Li, Y.; Wang, L.; Song, L. The cGAS/STING–TBK1–IRF Regulatory Axis Orchestrates a Primitive Interferon-Like Antiviral Mechanism in Oyster. Front. Immunol. 2021, 12, 689783. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Tang, X.; Guo, Y.; Hu, J. Endoplasmic reticulum composition and form: Proteins in and out. Curr. Opin. Cell Biol. 2021, 71, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Sajid, M.; Ullah, H.; Yan, K.; He, M.; Feng, J.; Shereen, M.A.; Hao, R.; Li, Q.; Guo, D.; Chen, Y.; et al. The Functional and Antiviral Activity of Interferon Alpha-Inducible IFI6 Against Hepatitis B Virus Replication and Gene Expression. Front. Immunol. 2021, 12, 634937. [Google Scholar] [CrossRef] [PubMed]
- Meyer, K.; Kwon, Y.-C.; Liu, S.; Hagedorn, C.H.; Ray, R.B.; Ray, R. Interferon-α inducible protein 6 impairs EGFR activation by CD81 and inhibits hepatitis C virus infection. Sci. Rep. 2015, 5, 9012. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.-H.; Wan, D.-H.; Chen, Y.-G.; Weng, S.-P.; Yu, X.-Q.; He, J.-G. Characterization of four novel caspases from Litopenaeus vannamei (Lvcaspase2-5) and their role in WSSV infection through dsRNA-mediated gene silencing. PLoS ONE 2013, 8, e80418. [Google Scholar]
- Rijiravanich, A.; Browdy, C.L.; Withyachumnarnkul, B. Knocking down caspase-3 by RNAi reduces mortality in Pacific white shrimp Penaeus (Litopenaeus) vannamei challenged with a low dose of white-spot syndrome virus. Fish Shellfish Immunol. 2008, 24, 308–313. [Google Scholar] [CrossRef]
- Hu, W.-Y.; Yao, C.-L. Molecular and immune response characterizations of a novel AIF and cytochrome c in Litopenaeus vannamei defending against WSSV infection. Fish Shellfish Immunol. 2016, 56, 84–95. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5’-3’) |
|---|---|
| RACE | |
| 5’RACE-1 | TGAGTGCGTTCCGCTTCAT |
| 5’RACE-2 | TCCTTGCTGTCCTTCCGTGTG |
| 3’RACE-1 | AGCTGCGGGAGTTGGAGCTGCT |
| 3’RACE-2 | AATTAAGAACACATTAATGAATGTA |
| Quantitative RT-PCR | |
| qPCR-LvEF-1α-F | TATGCTCCTTTTGGACGTTTTGC |
| qPCR-LvEF-1α-R | CCTTTTCTGCGGCCTTGGTAG |
| qPCR-LvAIF-F | TCTCTGGTGAGGGTGAAGCTCCCTA |
| qPCR-LvAIF-R | CTCCTTTCTTTCCCGTTCCATTGTT |
| qPCR-LvCYC-F | GAACGCGTCCCGGGGTTTC |
| qPCR-LvCYC-R | CTCGGTCTGCACATTCGGTCT |
| qPCR-Lvcasp1-F | CCGGGGCAAGAGGGCGGAGGAATAT |
| qPCR-Lvcasp1-R | CGGCACTGGGTCGCGGTTTGAGAGC |
| qPCR-Lvcasp2-F | ATGGCTCGTGGTTCATTCAG |
| qPCR-Lvcasp2-R | CATCAGGGTTGAGACAATACAGG |
| qPCR-Lvcasp3-F | AGTTAGTACAAACAGATTGGAGCG |
| qPCR-Lvcasp3-R | TTGTGGACAGACAGTATGAGGC |
| qPCR-Lvcasp4-F | CATGCTTGACATACCCGATG |
| qPCR-Lvcasp4-R | TGTCCGGCATTGTTGAGTAG |
| qPCR-Lvcasp5-F | GAAGGAGTGAAGCTAAACGAGAC |
| qPCR-Lvcasp5-R | CAGTAGACCAGCAGATAAGGAAGT |
| qPCR-LvIF6-16-F | ACTCATTCTCATTTTGCTGCTTTTCTC |
| qPCR-LvIF6-16-R | CCACATTCATCGGTGCTGTATTCT |
| qPCR-LvSTAT-F | CACAGAAGGTGTCAGGGCTATT |
| qPCR-LvSTAT-R | GATGCGCTGCTGAAGACTATTT |
| qPCR-LvJAK-F | TTTTGTAGGATGCTTGAATGGGTA |
| qPCR-LvJAK-R | GATAGAGAAGAGAAGGCGTTGAT |
| Protein expression | |
| LvIFI6-16-F | GGGGTACCATCAAAATGAAGCGGAACGCACTCATTC |
| LvIFI6-16-R | CGGAATTCATTGCCCTTTTCCCCACCAAAG |
| GFP-F | GGTTCGAAATCAAAATGGTGAGCAAGGGCGAGGAG |
| GFP-R | TTGTTTAAACTTACTTGTACAGCTCGTCCATGC |
| dsRNA template amplification | |
| GFP-T7-F | GGATCCTAATACGACTCACTATAGGGTGGTCCCAGTTCTTGTT |
| GFP-R | TTCTTTGGTTTGTCTCCC |
| GFP-F | GTGGTCCCAGTTCTTGTT |
| GFP-T7-R | GGATCCTAATACGACTCACTATAGGTTCTTTGGTTTGTCTCCC |
| LvIfi6-16-T7-F | GGATCCTAATACGACTCACTATAGGGAAGCGGAACGCACTCAT |
| LvIfi6-16-R | AATTGCCCTTTTCCCCAC |
| LvIfi6-16-F | GAAGCGGAACGCACTCAT |
| LvIfi6-16-T7-R | GGATCCTAATACGACTCACTATAGGAATTGCCCTTTTCCCCAC |
| LvSTAT-T7-F | GGATCCTAATACGACTCACTATAGGTCAGTATGCCCAGTCCTT |
| LvSTAT-R | CCTAACTCTTTCCGTCTCC |
| LvSTAT-F | TCAGTATGCCCAGTCCTT |
| LvSTAT-T7-R | GGATCCTAATACGACTCACTATAGGCCTAACTCTTTCCGTCTCC |
| LvIRF-T7-F | GGATCCTAATACGACTCACTATAGGGCCGCCATCTTTCACCAA |
| LvIRF-R | TGTCGTAGGAATGCGAGGAG |
| LvIRF-F | GCCGCCATCTTTCACCAA |
| LvIRF-T7-R | GGATCCTAATACGACTCACTATAGGTGTCGTAGGAATGCGAGGAG |
| Absolute quantitative PCR | |
| WSSV32678-F | TGTTTTCTGTATGTAATGCGTGTAGGT |
| WSSV32753-R | CCCACTCCATGGCCTTCA |
| TaqMan probe WSSV32706 | CAAGTACCCAGGCCCAGTGTCATACGTT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lǚ, K.; Li, H.; Wang, S.; Li, A.; Weng, S.; He, J.; Li, C. Interferon-Induced Protein 6-16 (IFI6-16) from Litopenaeus vannamei Regulate Antiviral Immunity via Apoptosis-Related Genes. Viruses 2022, 14, 1062. https://doi.org/10.3390/v14051062
Lǚ K, Li H, Wang S, Li A, Weng S, He J, Li C. Interferon-Induced Protein 6-16 (IFI6-16) from Litopenaeus vannamei Regulate Antiviral Immunity via Apoptosis-Related Genes. Viruses. 2022; 14(5):1062. https://doi.org/10.3390/v14051062
Chicago/Turabian StyleLǚ, Kai, Haoyang Li, Sheng Wang, Anxing Li, Shaoping Weng, Jianguo He, and Chaozheng Li. 2022. "Interferon-Induced Protein 6-16 (IFI6-16) from Litopenaeus vannamei Regulate Antiviral Immunity via Apoptosis-Related Genes" Viruses 14, no. 5: 1062. https://doi.org/10.3390/v14051062