
Detection of Chronic Wasting Disease Prions in Fetal Tissues of Free-Ranging White-Tailed Deer

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. White-Tailed Deer Tissues: Origin, Handling, and Fetal Prnp Genotyping
2.2. Gestational Age
2.3. Serial Protein Misfolding Cyclic Amplification (sPMCA)
2.4. Western Blotting
2.5. Real Time Quaking Induced Conversion (RT-QuIC)
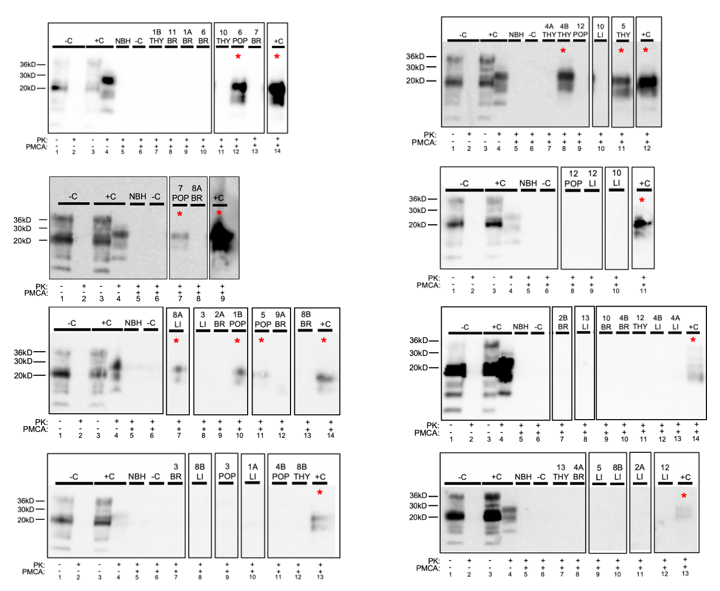
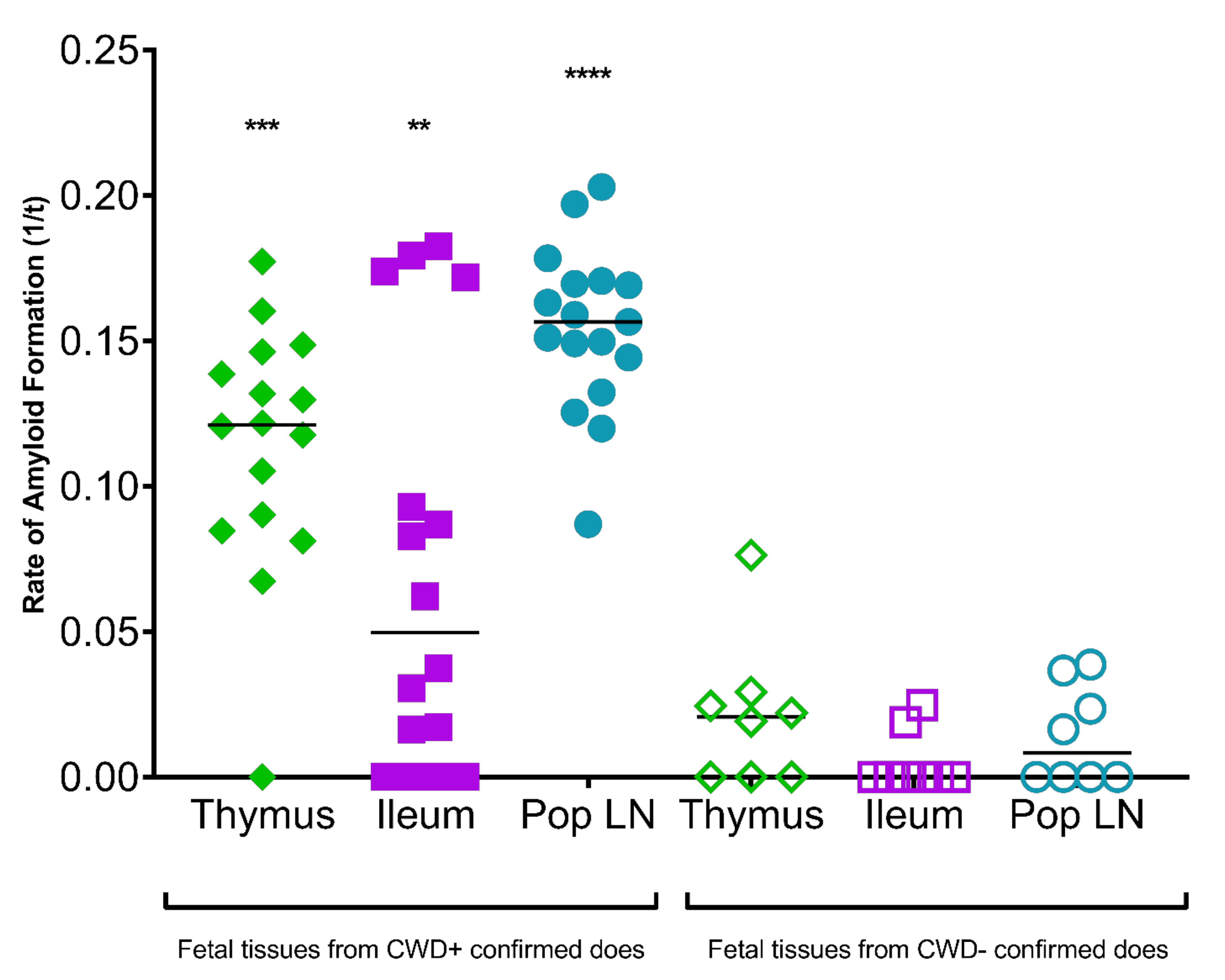
3. Results
3.1. Gestational Aging
3.2. White-Tailed Deer Fetal Tissue

{kind=link}
{kind=link}
| Fetuses | Does | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Fetus ID | Sex | Weight (g) | Length (mm) | Gestation Length (Days) | Doe ID | Date Collected | Age (Years) | Weight (kg) | CWD IHC Result | |
| RPLN | Obex | |||||||||
| 1A | M | 790 | 289 | 127–129 | 1 | March 2011 | 2.8 | 47 | Pos | Pos |
| 1B | F | 727 | 283 | |||||||
| 2A | M | 820 | 289 | 119–129 | 2 | March 2011 | 2.8 | 48 | Pos | Pos |
| 2B | F | 510 | 257 | |||||||
| 3 | M | 721 | 277 | 125 | 3 | March 2011 | 3.8 | 46 | Pos | Pos |
| 4A | M | 1115 | 332 | 139–143 | 4 | April 2011 | 6.8 | 51 | Pos | Pos |
| 4B | F | 948 | 318 | |||||||
| 5 | M | 1579 | 374 | 157 | 5 | April 2011 | 2.8 | 49 | Pos | Pos |
| 6 | M | 1647 | 378 | 158 | 6 | April 2011 | 2.8 | 45 | Pos | ND |
| 7 | M | 1195 | 336 | 144 | 7 | April 2011 | 1.8 | 41 | Pos | Pos |
| 8A | F | 418 | 238 | 113–115 | 8 | March 2012 | 3.8 | 55 | Pos | Pos |
| 8B | F | 403 | 243 | |||||||
| 9A | F | 870 | 315 | 138 | 9 | April 2012 | 2.8 | 40 | Pos | Pos |
| 9B | F | 829 | 316 | 138 | ||||||
| Fetus ID | sPMCA Run # | Thymus | Brain | Ileum | Popliteal LN | Liver |
|---|---|---|---|---|---|---|
| 1A | 1 * | Neg | Neg | |||
| 2 | Neg | Neg | Neg | Neg | ||
| 3 | Neg | Neg | Neg | Neg | ||
| 1B | 1 * | Neg | Pos | |||
| 2 | Neg | Neg | Neg | Neg | ||
| 3 | Neg | Neg | ||||
| 2A | 1 * | Neg | Neg | |||
| 2 | Neg | Neg | Neg | Neg | ||
| 3 | Neg | Neg | Neg | |||
| 2B | 1 * | Neg | ||||
| 2 | Neg | Neg | Neg | Neg | ||
| 3 | Neg | Neg | Neg | |||
| 3 | 1 * | Neg | Neg | |||
| 2 | Neg | Neg | Pos | |||
| 3 | Neg | Neg | Pos | |||
| 4A | 1 * | Neg | Neg | Neg | ||
| 2 | Neg | Neg | Neg | Neg | ||
| 3 | Neg | Neg | Neg | Neg | ||
| 4B | 1 * | Pos | Neg | Neg | Neg | |
| 2 | Neg | Neg | Neg | |||
| 3 | Neg | |||||
| 5 | 1 * | Pos | Pos | Neg | ||
| 2 | Neg | Neg | Neg | Neg | ||
| 3 | Neg | Neg | Neg | |||
| 6 | 1 * | Neg | Pos | |||
| 2 | Neg | Pos | Neg | |||
| 3 | Neg | Neg | Neg | |||
| 7 | 1 * | Neg | Pos | |||
| 2 | Pos | Neg | Neg | Neg | ||
| 3 | Neg | Neg | Neg | |||
| 8A | 1 * | Neg | Pos | |||
| 2 | Neg | Pos | ||||
| 3 | Neg | |||||
| 8B | 1 * | Neg | Neg | Neg | ||
| 2 | Neg | Neg | Neg | |||
| 3 | Neg | Neg | Neg | |||
| 9A | 1 * | Neg | ||||
| 2 | Neg | Neg | ||||
| 3 | Neg | Neg | Neg | |||
| 9B | 1 * | |||||
| 2 | Neg | Neg | Neg | |||
| 3 | Neg | Neg | Neg |
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mathiason, C.K.; Powers, J.G.; Dahmes, S.J.; Osborn, D.A.; Miller, K.V.; Warren, R.J.; Mason, G.L.; Hays, S.A.; Hayes-Klug, J.; Seelig, D.M.; et al. Infectious prions in the saliva and blood of deer with chronic wasting disease. Science 2006, 314, 133–136. [Google Scholar] [CrossRef] [Green Version]
- Tamgüney, G.; Miller, M.W.; Wolfe, L.L.; Sirochman, T.M.; Glidden, D.; Palmer, C.; Lemus, A.; DeArmond, S.J.; Prusiner, S.B. Asymptomatic deer excrete infectious prions in faeces. Nature 2009, 461, 529–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safar, J.G.; Lessard, P.; Tamgüney, G.; Freyman, Y.; Deering, C.; Letessier, F.; DeArmond, S.J.; Prusiner, S.B. Transmission and detection of prions in feces. J. Infect. Dis. 2008, 198, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Haley, N.J.; Seelig, D.M.; Zabel, M.D.; Telling, G.C.; Hoover, E.A. Detection of CWD prions in urine and saliva of deer by transgenic mouse bioassay. PLoS ONE 2009, 4, e4848. [Google Scholar] [CrossRef] [PubMed]
- Hoinville, L.J.; Tongue, S.C.; Wilesmith, J.W. Evidence for maternal transmission of scrapie in naturally affected flocks. Prev. Vet. Med. 2010, 93, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, A.G.; Young, G.B.; Stamp, J.T.; Renwick, C.C. An analysis of natural scrapie in Suffolk sheep. Heredity 1965, 20, 485–503. [Google Scholar] [CrossRef] [Green Version]
- Dickinson, A.G.S.J.; Renwich, C.C. Maternal and lateral transmission of scrapie in sheep. J. Comp. Pathol. 1974, 84, 19–25. [Google Scholar] [CrossRef]
- Andreoletti, O.L.C.; Chabert, A.; Monnereau, L.; Tabouret, G.; Lantier, F.; Berthon, P.; Eychenne, F.; Lafond-Benestad, S.; Elsen, J.M.; Schelcher, F. PrPSc accumulation in placentas of ewes exposed to natural scrapie: Influence of foetal PrP genotype and effect on ewe-to- lamb transmission. J. Gen. Virol. 2002, 83, 2607–2616. [Google Scholar] [CrossRef] [PubMed]
- O’Rourke, K.I.; Zhuang, D.; Truscott, T.C.; Yan, H.; Schneider, D.A. Sparse PrP(Sc) accumulation in the placentas of goats with naturally acquired scrapie. BMC Vet. Res. 2011, 7, 7. [Google Scholar]
- Tuo, W.; Zhuang, D.; Knowles, D.P.; Cheevers, W.P.; Sy, M.S.; O’Rourke, K.I. Prp-c and Prp-Sc at the fetal-maternal interface. J. Biol. Chem. 2001, 276, 18229–18234. [Google Scholar] [CrossRef] [Green Version]
- Tuo, W.; O’Rourke, K.I.; Zhuang, D.; Cheevers, W.P.; Spraker, T.R.; Knowles, D.P. Pregnancy status and fetal prion genetics determine PrPSc accumulation in placentomes of scrapie-infected sheep. Proc. Natl. Acad. Sci. USA 2002, 99, 6310–6315. [Google Scholar] [CrossRef] [Green Version]
- Onodera, T.; Ikeda, T.; Muramatsu, Y.; Shinagawa, M. Isolation of scrapie agent from the placenta of sheep with natural scrapie in Japan. Microbiol. Immunol. 1993, 37, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Lacroux, C.; Corbière, F.; Tabouret, G.; Lugan, S.; Costes, P.; Mathey, J.; Delmas, J.M.; Weisbecker, J.L.; Foucras, G.; Cassard, H.; et al. Dynamics and genetics of PrPSc placental accumulation in sheep. J. Gen. Virol. 2007, 88 Pt 3, 1056–1061. [Google Scholar] [CrossRef] [PubMed]
- Foster, J.D.; Goldmann, W.; Hunter, N. Evidence in sheep for pre-natal transmission of scrapie to lambs from infected mothers. PLoS ONE 2013, 8, e79433. [Google Scholar]
- Alverson, J.; O’Rourke, K.I.; Baszler, T.V. PrPSc accumulation in fetal cotyledons of scrapie-resistant lambs is influenced by fetus location in the uterus. J. Gen. Virol. 2006, 87 Pt 4, 1035–1041. [Google Scholar] [CrossRef] [PubMed]
- Race, R.; Jenny, A.; Sutton, D. Scrapie infectivity and proteinase K-resistant prion protein in sheep placenta, brain, spleen, and lymph node: Implications for transmission and antemortem diagnosis. J. Infect. Dis. 1998, 178, 949–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, D.A.; Madsen-Bouterse, S.A.; Zhuang, D.; Truscott, T.C.; Dassanayake, R.P.; O’Rourke, K.I. The placenta shed from goats with classical scrapie is infectious to goat kids and lambs. J. Gen. Virol. 2015, 96, 2464–2469. [Google Scholar] [CrossRef]
- Spiropoulos, J.; Hawkins, S.A.; Simmons, M.M.; Bellworthy, S.J. Evidence of in utero transmission of classical scrapie in sheep. J. Virol. 2014, 88, 4591–4594. [Google Scholar] [CrossRef] [Green Version]
- Maddison, B.C.; Baker, C.A.; Rees, H.C.; Terry, L.A.; Thorne, L.; Bellworthy, S.J.; Whitelam, G.C.; Gough, K.C. Prions are secreted in milk from clinically normal scrapie-exposed sheep. J. Virol. 2009, 83, 8293–8296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konold, T.; Moore, S.J.; Bellworthy, S.J.; Simmons, H.A. Evidence of scrapie transmission via milk. BMC Vet. Res. 2008, 4, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konold, T.; Moore, S.J.; Bellworthy, S.J.; Terry, L.; Thorne, L.; Ramsay, A.; Salguero, F.J.; Simmons, M.M.; Simmons, H. Evidence of effective scrapie transmission via colostrum and milk in sheep. BMC Vet. Res. 2013, 9, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madsen-Bouterse, S.A.; Highland, M.A.; Dassanayake, R.P.; Zhuang, D.; Schneider, D.A. Low-volume goat milk transmission of classical scrapie to lambs and goat kids. PLoS ONE 2018, 13, e0204281. [Google Scholar] [CrossRef] [PubMed]
- Ligios, C.; Cancedda, M.G.; Carta, A.; Santucciu, C.; Maestrale, C.; Demontis, F.; Saba, M.; Patta, C.; DeMartini, J.C.; Aguzzi, A.; et al. Sheep with scrapie and mastitis transmit infectious prions through the milk. J. Virol. 2011, 85, 1136–1139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nalls, A.V.; McNulty, E.; Powers, J.; Seelig, D.M.; Hoover, C.; Haley, N.J.; Hayes-Klug, J.; Anderson, K.; Stewart, P.; Goldmann, W.; et al. Mother to offspring transmission of chronic wasting disease in reeves’ muntjac deer. PLoS ONE 2013, 8, e71844. [Google Scholar]
- Nalls, A.V.; McNulty, E.; Hoover, C.E.; Pulscher, L.A.; Hoover, E.A.; Mathiason, C.K. Infectious Prions in the Pregnancy Microenvironment of CWD-infected Reeves’ Muntjac Deer. J. Virol. 2017, 91, e00501-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selariu, A.; Powers, J.G.; Nalls, A.; Brandhuber, M.; Mayfield, A.; Fullaway, S.; Wyckoff, C.A.; Goldmann, W.; Zabel, M.M.; Wild, M.A.; et al. In utero transmission and tissue distribution of chronic wasting disease-associated prions in free-ranging Rocky Mountain elk. J. Gen. Virol. 2015, 96, 3444–3455. [Google Scholar] [CrossRef] [PubMed]
- Stewart, P.; Campbell, L.; Skogtvedt, S.; Griffin, K.A.; Arnemo, J.M.; Tryland, M.; Girling, S.; Miller, M.W.; Tranulis, M.A.; Goldmann, W. Genetic predictions of prion disease susceptibility in carnivore species based on variability of the prion gene coding region. PLoS ONE 2012, 7, e50623. [Google Scholar]
- Hamilton, R.J.; Tobin, M.L.; Moore, W.G. Aging fetal white-tailed deer. In Proceedings of the Annual Conference Southeastern Association of Fish and Wildlife Agencies, Lexington, KY, USA, 27–30 October 1985; pp. 389–395. [Google Scholar]
- McNulty, E.; Nalls, A.V.; Mellentine, S.; Hughes, E.; Pulscher, L.; Hoover, E.A.; Mathiason, C.K. Comparison of conventional, amplification and bio-assay detection methods for a chronic wasting disease inoculum pool. PLoS ONE 2019, 14, e0216621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saa, P.; Castilla, J.; Soto, C. Presymptomatic detection of prions in blood. Science 2006, 313, 92–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bravo-Risi, F.; Soto, P.; Eckland, T.; Dittmar, R.; Ramírez, S.; Catumbela, C.S.G.; Soto, C.; Lockwood, M.; Nichols, T.; Morales, R. Detection of CWD prions in naturally infected white-tailed deer fetuses and gestational tissues by PMCA. Sci. Rep. 2021, 11, 18385. [Google Scholar]
- Garza, M.C.; Fernández-Borges, N.; Boles, R.; Badiola, J.J.; Castilla, J.; Monleon, E. Detection of PrPres in Genetically Susceptible Fetuses from Sheep with Natural Scrapie. PLoS ONE 2011, 6, e27525. [Google Scholar] [CrossRef] [Green Version]
- Slack, J.M.W. Essential Developmental Biology; Wiley-Blackwell: Oxford, UK, 2013. [Google Scholar]
- Hunter, N.; Foster, J.; Chong, A.; McCutcheon, S.; Parnham, D.; Eaton, S.; MacKenzie, C.; Houston, F. Transmission of prion diseases by blood transfusion. J. Gen. Virol. 2002, 83 Pt 11, 2897–2905. [Google Scholar] [CrossRef] [PubMed]
- Houston, F.; Foster, J.D.; Chong, A.; Hunter, N.; Bostock, C.J. Transmission of BSE by blood transfusion in sheep. Lancet 2000, 356, 999–1000. [Google Scholar] [CrossRef]
- Mathiason, C.K.; Powers, J.G.; Dahmes, S.J. Don’t kiss that deer. Compend. Contin. Educ. For. Pract. Vet. 2006, 28, 822. [Google Scholar]
- Mathiason, C.K.; Hayes-Klug, J.; Hays, S.A.; Powers, J.; Osborn, D.A.; Dahmes, S.J.; Miller, K.V.; Warren, R.J.; Mason, G.L.; Telling, G.C.; et al. B Cells and Platelets Harbor Prion Infectivity in the Blood of Deer Infected with Chronic Wasting Disease. J. Virol. 2010, 84, 5097–5107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peden, A.H.; Head, M.W.; Ritchie, D.L.; Bell, J.E.; Ironside, J.W. Preclinical vCJD after blood transfusion in a PRNP codon 129 heterozygous patient. Lancet 2004, 364, 527–529. [Google Scholar] [CrossRef]
- Myagkaya, G.; Schellens, J.P. Final stages of erythrophagocytosis in the sheep placenta. Cell Tissue Res. 1981, 214, 501–518. [Google Scholar] [CrossRef]
- Wooding, F.B. The role of the binucleate cell in ruminant placental structure. J. Reprod. Fertil. Suppl. 1982, 31, 31–39. [Google Scholar] [PubMed]
- Wooding, F.B. Current topic: The synepitheliochorial placenta of ruminants; binculeate cell fusions and hormone production. Placenta 1992, 13, 101–113. [Google Scholar] [CrossRef]
- Wooding, F.B. Marshall’s Physiology of Reproduction, 4th ed.; Springer: London, UK, 1994. [Google Scholar]
- Wooding, F.B.; Morgan, G.; Adam, C.L. Structure and function in the rumiant synepithelialchorial placenta: Central role of the trophoblast binucleate cll in deer. Microsc. Res. Tech. 1997, 38, 88–99. [Google Scholar] [CrossRef]
- Soto, P.; Claflin, I.A.; Bursott, A.L.; Schwab-McCoy, A.D.; Bartz, J.C. Cellular prion protein gene polymorphisms linked to differential scrapie susceptibility correlate with distinct residue connectivity between secondary structure elements. J. Biomol. Struct. Dyn. 2021, 39, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Hagenaars, T.J.; Melchior, M.B.; Bossers, A.; Davidse, A.; Engel, B.; van Zijderveld, F.G. Scrapie prevalence in sheep of susceptible genotype is declining in a population subject to breeding for resistance. BMC Vet. Res. 2010, 6, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haley, N.; Donner, R.; Merrett, K.; Miller, M.; Senior, K. Selective Breeding for Disease-Resistant PRNP Variants to Manage Chronic Wasting Disease in Farmed Whitetail Deer. Genes 2021, 12, 1396. [Google Scholar] [CrossRef] [PubMed]
- Benestad, S.L.; Telling, G.C. Chronic wasting disease: An evolving prion disease of cervids. Handb. Clin. Neurol. 2018, 153, 135–151. [Google Scholar] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nalls, A.V.; McNulty, E.E.; Mayfield, A.; Crum, J.M.; Keel, M.K.; Hoover, E.A.; Ruder, M.G.; Mathiason, C.K. Detection of Chronic Wasting Disease Prions in Fetal Tissues of Free-Ranging White-Tailed Deer. Viruses 2021, 13, 2430. https://doi.org/10.3390/v13122430
Nalls AV, McNulty EE, Mayfield A, Crum JM, Keel MK, Hoover EA, Ruder MG, Mathiason CK. Detection of Chronic Wasting Disease Prions in Fetal Tissues of Free-Ranging White-Tailed Deer. Viruses. 2021; 13(12):2430. https://doi.org/10.3390/v13122430
Chicago/Turabian StyleNalls, Amy V., Erin E. McNulty, Amber Mayfield, James M. Crum, Michael K. Keel, Edward A. Hoover, Mark G. Ruder, and Candace K. Mathiason. 2021. "Detection of Chronic Wasting Disease Prions in Fetal Tissues of Free-Ranging White-Tailed Deer" Viruses 13, no. 12: 2430. https://doi.org/10.3390/v13122430