
Rocio Virus: An Updated View on an Elusive Flavivirus

 ,
,  ,
,  , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
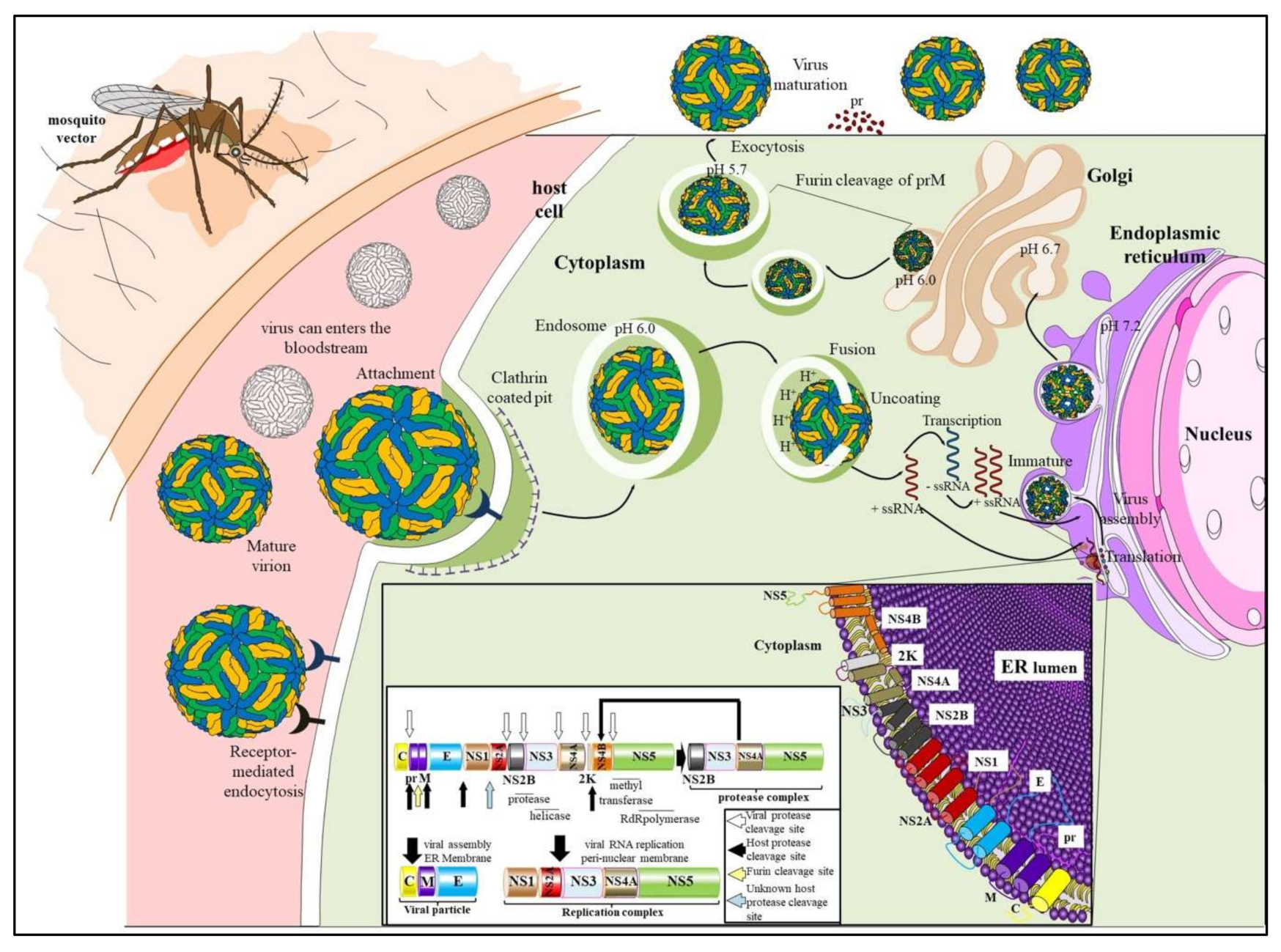
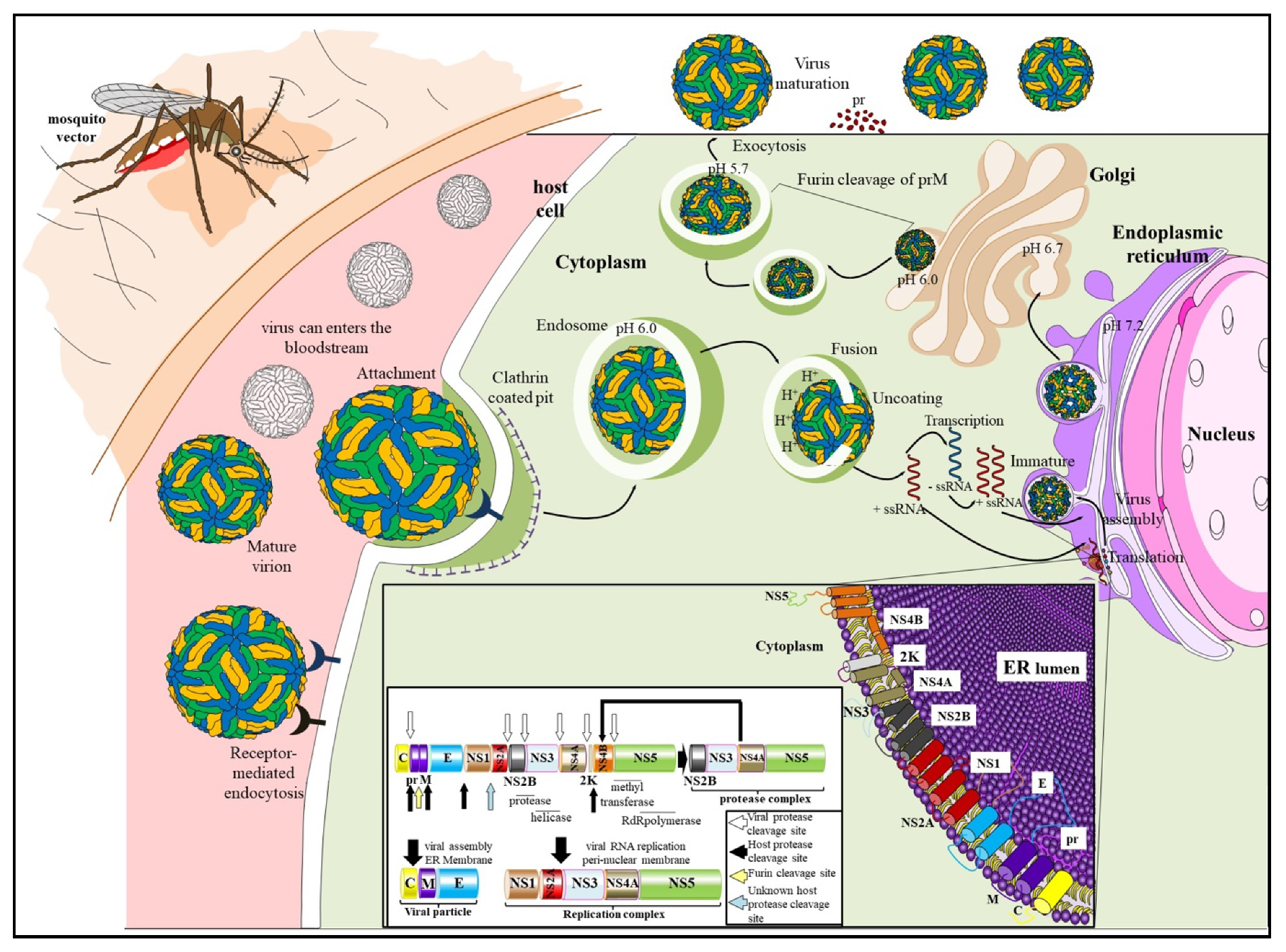
2. Genome Organization and Replication
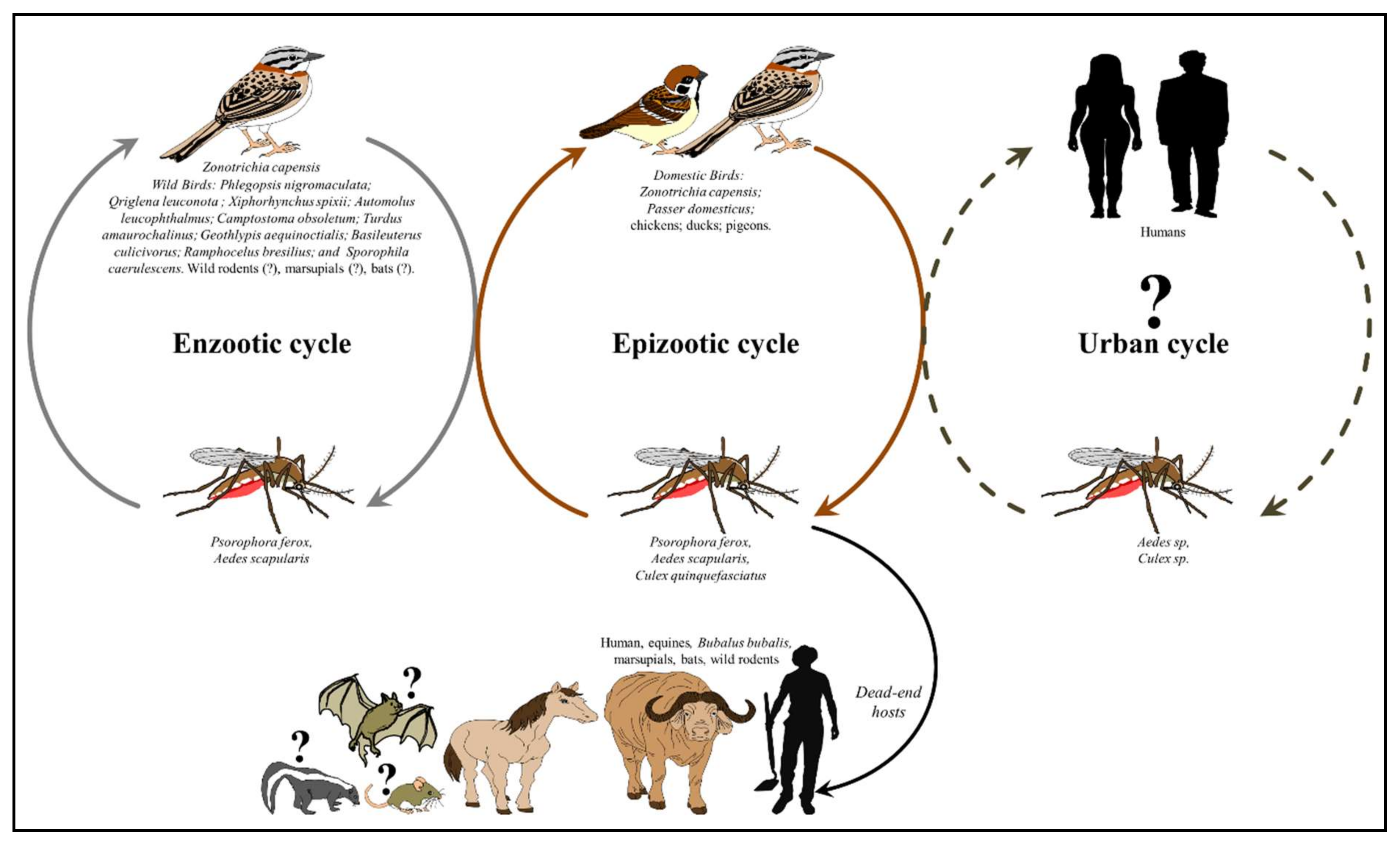
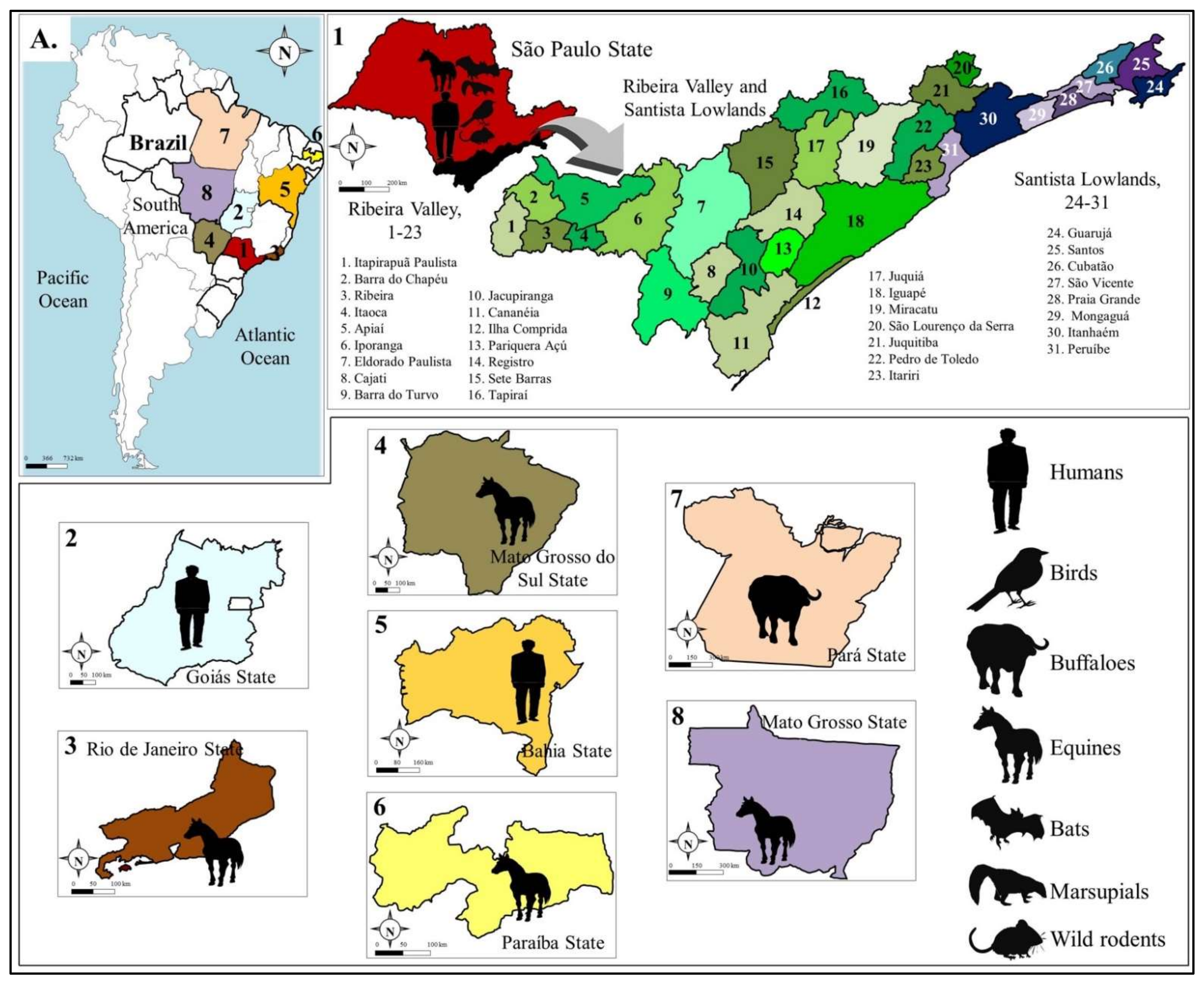
3. Transmission Cycles and Host Range
4. Human Epidemiology
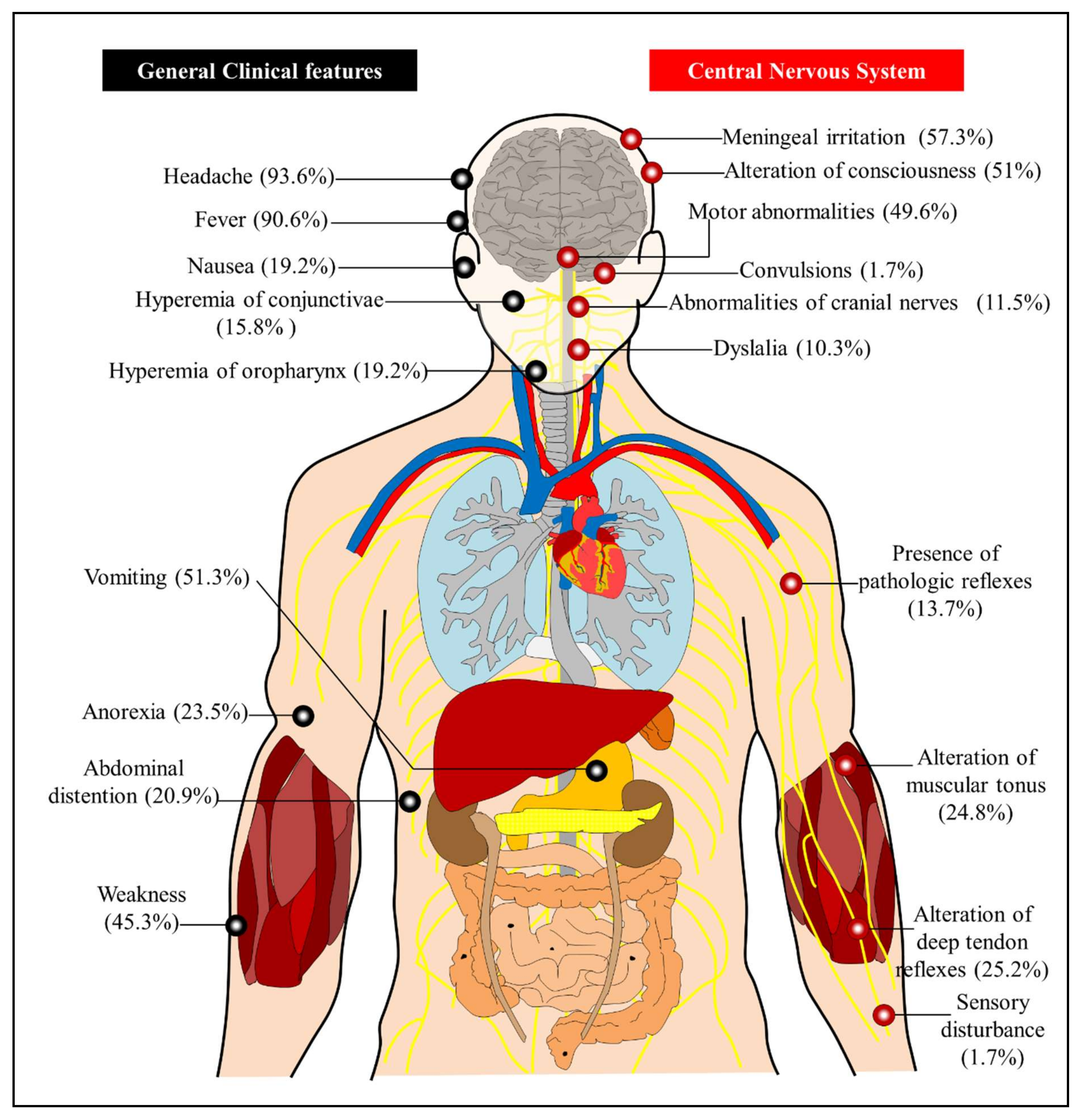
5. Clinical Presentation and Pathogenesis of ROCV Infection
6. Diagnosis, Treatment and Prevention Options
7. Conclusions and Future Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tanaka, H.; Weigl, D.R.; de Souza Lopes, O. The replication of Rocio virus in brain tissue of suckling mice. Study by electron microscopy. Arch. Virol. 1983, 78, 309–314. [Google Scholar] [CrossRef]
- De Souza Lopes, O.; Coimbra, T.L.; de Abreu Sacchetta, L.; Calisher, C.H. Emergence of a new arbovirus disease in Brazil. I. Isolation and characterization of the etiologic agent, Rocio virus. Am. J. Epidemiol. 1978, 107, 444–449. [Google Scholar] [CrossRef]
- De Souza Lopes, O.; de Abreu Sacchetta, L.; Coimbra, T.L.; Pinto, G.H.; Glasser, C.M. Emergence of a new arbovirus disease in Brazil. II. Epidemiologic studies on 1975 epidemic. Am. J. Epidemiol. 1978, 108, 394–401. [Google Scholar]
- Diaz, L.A.; Re, V.; Almiron, W.R.; Farias, A.; Vazquez, A.; Sanchez-Seco, M.P.; Aguilar, J.; Spinsanti, L.; Konigheim, B.; Visintin, A.; et al. Genotype III Saint Louis encephalitis virus outbreak, Argentina, 2005. Emerg. Infect. Dis. 2006, 12, 1752–1754. [Google Scholar] [CrossRef]
- Causey, O.R.; Causey, C.E.; Maroja, O.M.; Macedo, D.G. The isolation of arthropod-borne viruses, including members of two hitherto undescribed serological groups, in the Amazon region of Brazil. Am. J. Trop. Med. Hyg. 1961, 10, 227–249. [Google Scholar] [CrossRef] [PubMed]
- Tiriba, A.C.; Miziara, A.M.; Lorenco, R.; da Costa, R.B.; Costa, C.S.; Pinto, G.H. Primary human epidemic encephalitis induced by Arbovirus found at the sea shore south of the State of Sao Paulo. Clinical study in an emergency hospital. AMB Rev. Assoc. Med. Bras. 1976, 22, 415–420. [Google Scholar]
- Iversson, L.B. Aspects of the encephalitis epidemic due to arbovirus in the region of Vale do Ribeira, S. Paulo, Brazil from 1975–1978. Rev. Saude Publica 1980, 14, 9–35. [Google Scholar] [CrossRef] [PubMed]
- De Souza Lopes, O.; de Abreu Sacchetta, L.; Francy, D.B.; Jakob, W.L.; Calisher, C.H. Emergence of a new arbovirus disease in Brazil. III. Isolation of Rocio virus from Psorophora Ferox (Humboldt, 1819). Am. J. Epidemiol. 1981, 113, 122–125. [Google Scholar] [CrossRef]
- Saivish, M.V.; da Costa, V.G.; Rodrigues, R.L.; Feres, V.C.R.; Montoya-Diaz, E.; Moreli, M.L. Detection of Rocio Virus SPH 34675 during Dengue Epidemics, Brazil, 2011–2013. Emerg. Infect. Dis. 2020, 26, 797–799. [Google Scholar] [CrossRef] [PubMed]
- Degallier, N.; Travassos da Rosa, A.P.A.; da Silva, J.M.C.; Rodrigues, S.G.; Vasconcelos, P.F.C.; Travassos da Rosa, J.F.S.; da Silva, G.P.; da Silva, R.P. Birds as Arbovirus Hosts in Brazilian Amazonia; Centers for Disease Control: Belém, Brazil, 1991; pp. 66–67. [Google Scholar]
- Ferreira, I.B.; Pereira, L.E.; Rocco, I.M.; Marti, A.T.; de Souza, L.T.; Iversson, L.B. Surveillance of arbovirus infections in the Atlantic Forest Region, State of Sao Paulo, Brazil. I. Detection of hemagglutination-inhibiting antibodies in wild birds between 1978 and 1990. Rev. Inst. de Med. Trop. Sao Paulo 1994, 36, 265–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, J.R.; Romeiro, M.F.; Souza, W.M.; Munhoz, T.D.; Borges, G.P.; Soares, O.A.; Campos, C.H.; Machado, R.Z.; Silva, M.L.; Faria, J.L.; et al. A Saint Louis encephalitis and Rocio virus serosurvey in Brazilian horses. Rev. Soc. Bras. Med. Trop. 2014, 47, 414–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casseb, A.R.; Cruz, A.V.; Jesus, I.S.; Chiang, J.O.; Martins, L.C.; Silva, S.P.; Henriques, D.F.; Casseb, L.M.; Vasconcelos, P.F. Seroprevalence of flaviviruses antibodies in water buffaloes (Bubalus bubalis) in Brazilian Amazon. J. Venom. Anim. Toxins Incl. Trop. Dis. 2014, 20, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Straatmann, A.; Santos-Torres, S.; Vasconcelos, P.F.; da Rosa, A.P.; Rodrigues, S.G.; Tavares-Neto, J. Serological evidence of the circulation of the Rocio arbovirus (Flaviviridae) in Bahia. Rev. Soc. Bras. Med. Trop. 1997, 30, 511–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iversson, L.B.; Travassos da Rosa, A.P.; Rosa, M.D. Recent occurrence of human infection by Rocio arbovirus in the Valley of Ribeira region. Rev. Inst. Med. Trop. Sao Paulo 1989, 31, 28–31. [Google Scholar] [CrossRef]
- Iversson, L.B. Rocio encephalitis. In The Arboviruses: Epidemiology and Ecology; Monath, T.P., Ed.; CRC Press: Boca Raton, FL, USA, 1986; Volume IV, pp. 77–92. [Google Scholar]
- Medeiros, D.B.A.; Nunes, M.R.T.; Vasconcelos, P.F.C.; Chang, G.J.; Kuno, G. Complete genome characterization of Rocio virus (Flavivirus: Flaviviridae), a Brazilian flavivirus isolated from a fatal case of encephalitis during an epidemic in Sao Paulo state. J. Gen. Virol. 2007, 88, 2237–2246. [Google Scholar] [CrossRef] [PubMed]
- Simmonds, P.; Becher, P.; Collett, M.S.; Could, E.A.; Heinz, F.X.; Meyers, G.; Monath, T.P.; Plentev, A.; Rice, C.M.; Stiasny, K.; et al. Flaviviridae. In Virus Taxonomy—Classification and Nomenclature of Viruses. Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier Academic Press: New York, NY, USA, 2012; pp. 1003–1020. [Google Scholar]
- Rothan, H.A.; Kumar, M. Role of Endoplasmic Reticulum-Associated Proteins in Flavivirus Replication and Assembly Complexes. Pathogens 2019, 8, 148. [Google Scholar] [CrossRef] [Green Version]
- Roby, J.A.; Setoh, Y.X.; Hall, R.A.; Khromykh, A.A. Post-translational regulation and modifications of flavivirus structural proteins. J. Gen. Virol. 2015, 96, 1551–1569. [Google Scholar] [CrossRef]
- Boldescu, V.; Behnam, M.A.M.; Vasilakis, N.; Klein, C.D. Broad-spectrum agents for flaviviral infections: Dengue, Zika and beyond. Nat. Rev. Drug. Discov. 2017, 16, 565–586. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Fulcher, A.; Hossain, T.; Davidson, C.; Beier, J.C.; Xue, R.D. Body size, blood feeding activity, and fecundity of Psorophora howardii, Psorophora ciliata, and Psorophora ferox (Diptera: Culicidae). J. Med. Entomol. 2014, 51, 382–386. [Google Scholar] [CrossRef] [PubMed]
- Lane, J. Tribe Culicini, Deinocerites, Uranotaenia, Mansonia, Orthopodomyia, Aedomyia, Aedes, Psorophora, Haemagogus. In Neotropical Culicidae; University of Sao Paulo: Sao Paulo, Brazil, 1953; Volume II, pp. 553–1112. [Google Scholar]
- Causey, O.R.; Kumm, H.W.; Laemmert, H.W., Jr. Dispersion of forest mosquitoes in Brazil; further studies. Am. J. Trop. Med. Hyg. 1950, 30, 301–312. [Google Scholar] [CrossRef]
- Hammon, W.M.; Reeves, W.C. Laboratory Transmission of St. Louis Encephalitis Virus by Three Genera of Mosquitoes. J. Exp. Med. 1943, 78, 241–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turell, M.J.; O’Guinn, M.L.; Jones, J.W.; Sardelis, M.R.; Dohm, D.J.; Watts, D.M.; Fernandez, R.; Travassos da Rosa, A.; Guzman, H.; Tesh, R.; et al. Isolation of viruses from mosquitoes (Diptera: Culicidae) collected in the Amazon Basin region of Peru. J. Med. Entomol. 2005, 42, 891–898. [Google Scholar] [CrossRef] [PubMed]
- Chamberlain, R.W.; Sikes, R.K.; Nelson, D.B. Infection of Mansonia perturbans and Psorophora ferox mosquitoes with Venezuelan equine encephalomyelitis virus. Proc. Soc. Exp. Biol. Med. 1956, 91, 215–216. [Google Scholar] [CrossRef]
- Edman, J.D. Host-feeding patterns of Florida mosquitoes. I. Aedes, Anopheles, Coquillettidia, Mansonia and Psorophora. J. Med Entomol. 1971, 8, 687–695. [Google Scholar] [CrossRef]
- Davis, D.E. A comparison of mosquitoes captured with human bait and avian bait. Proc. Entomol. Soc. Wash. 1945, 47, 252–256. [Google Scholar]
- Forattini, O.P.; Gomes Ade, C.; Santos, J.L.; Galati, E.A.; Rabello, E.X.; Natal, D. Activity of Culicidae mosquitoes in residual forests in the Ribeira Valley, Sao Paulo, Brazil. Rev. Saude Publica 1981, 15, 557–586. [Google Scholar] [CrossRef] [PubMed]
- Forattini, O.P.; de Castro Gomes, A.; Galati, E.A.; Rabello, E.X.; Iversson, L.B. Ecologic studies on Culicidae mosquitoes in the Serra do Mar system, Brazil. 1. Observations in the extradomiciliary environment. Rev. Saude Publica 1978, 12, 297–325. [Google Scholar] [CrossRef] [PubMed]
- Forattini, O.P.; de Castro Gomes, A.; Galati, E.A.; Rabello, E.X.; Iversson, L.B. Ecological studies on Culicidae mosquitoes in the Serra do Mar system, Brazil. 2. Observations in domiciliary environments. Rev. Saude Publica 1978, 12, 476–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, C.J.; Forattini, O.P. Experimental transmission of Rocio encephalitis virus by Aedes scapularis (Diptera: Culicidae) from the epidemic zone in Brazil. J. Med. Entomol. 1984, 21, 34–37. [Google Scholar] [CrossRef]
- Mitchell, C.J.; Forattini, O.P.; Miller, B.R. Vector competence experiments with Rocio virus and three mosquito species from the epidemic zone in Brazil. Rev. Saude Publica 1986, 20, 171–177. [Google Scholar] [CrossRef]
- Reeves, L.E.; Medina, J.; Miqueli, E.; Sloyer, K.E.; Petrie, W.; Vasquez, C.; Burkett-Cadena, N.D. Establishment of Aedes (Ochlerotatus) scapularis (Diptera: Culicidae) in Mainland Florida, With Notes on the Ochlerotatus Group in the United States. J. Med. Entomol. 2021, 58, 717–729. [Google Scholar] [CrossRef]
- Arnell, J. Mosquito studies (Diptera, Culicidae) XXXIII. A revision of the scapularis group of Aedes. Contrib. Am. Entomol. Inst. 1976, 13, 1–144. [Google Scholar]
- Cunha, M.S.; Faria, N.R.; Caleiro, G.S.; Candido, D.S.; Hill, S.C.; Claro, I.M.; da Costa, A.C.; Nogueira, J.S.; Maeda, A.Y.; da Silva, F.G.; et al. Genomic evidence of yellow fever virus in Aedes scapularis, southeastern Brazil, 2016. Acta Trop. 2020, 205, 105390. [Google Scholar] [CrossRef]
- Shannon, R.C.; Whitman, L.; Franca, M. Yellow Fever Virus in Jungle Mosquitoes. Science 1938, 88, 110–111. [Google Scholar] [CrossRef]
- Aitken, T.H.; Anderson, C.R. Virus transmission studies with Trinidadian mosquitoes II. Further observations. Am. J. Trop. Med. Hyg. 1959, 8, 41–45. [Google Scholar] [CrossRef]
- Sellers, R.F.; Bergold, G.H.; Suarez, O.M.; Morales, A. Investigations during Venezuelan Equine Encephalitis Outbreaks in Venezuela--1962–1964. Am. J. Trop. Med. Hyg. 1965, 14, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Scherer, W.F.; Dickerman, R.W.; Diaz-Najera, A.; Ward, B.A.; Miller, M.H.; Schaffer, P.A. Ecologic studies of Venezuelan encephalitis virus in southeastern Mexico. 3. Infection of mosquitoes. Am. J. Trop. Med. Hyg. 1971, 20, 969–979. [Google Scholar] [CrossRef]
- Forattini, O.P.; Gomes Ade, C.; Natal, D.; Kakitani, I.; Marucci, D. Food preferences and domiciliation of Culicidae mosquitoes in the Ribeira Valley, Sao Paulo, Brazil, with special reference to Aedes scapularis and Culex (Melanoconion). Rev. Saude Publica 1989, 23, 9–19. [Google Scholar] [CrossRef]
- Lorosa, E.S.; Faria, M.S.; de Oliveira, L.C.; Alencar, J.; Marcondes, C.B. Blood meal identification of selected mosquitoes in Rio de Janeiro, Brazil. J. Am. Mosq. Control Assoc. 2010, 26, 18–23. [Google Scholar] [CrossRef]
- De Carvalho, G.C.; Malafronte Rdos, S.; Miti Izumisawa, C.; Souza Teixeira, R.; Natal, L.; Marrelli, M.T. Blood meal sources of mosquitoes captured in municipal parks in Sao Paulo, Brazil. J. Vector. Ecol. 2014, 39, 146–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, C.S.; Pie, M.R.; da Rocha, T.C.; Navarro-Silva, M.A. Molecular identification of blood meals in mosquitoes (Diptera, Culicidae) in urban and forested habitats in southern Brazil. PLoS ONE 2019, 14, e0212517. [Google Scholar] [CrossRef]
- Forattini, O.P.; Kakitani, I.; Massad, E.; Marucci, D. Studies on mosquitoes (Diptera: Culicidae) and anthropic environment. 9-Synanthropy and epidemiological vector role of Aedes scapularis in south-eastern Brazil. Rev. Saude Publica 1995, 29, 199–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapman, F.M. The post-glacial history of Zonotrichia capensis. In Bulletin of the American Museum of Natural History; American Museum of Natural History: New York, MY, USA, 1940; Volume 77, pp. 381–438. [Google Scholar]
- Ridgely, R.S.; Tudor, G. The Birds of South America; University of Texas Press: Austin, TX, USA, 1989; Volume 1. [Google Scholar]
- Lougheed, S.C.; Campagna, L.; Davila, J.A.; Tubaro, P.L.; Lijtmaer, D.A.; Handford, P. Continental phylogeography of an ecologically and morphologically diverse Neotropical songbird, Zonotrichia capensis. BMC Evol. Biol. 2013, 13, 58. [Google Scholar] [CrossRef] [Green Version]
- Monath, T.P.; Kemp, G.E.; Cropp, C.B.; Bowen, G.S. Experimental infection of house sparrows (Passer domesticus) with Rocio virus. Am. J. Trop. Med. Hyg. 1978, 27, 1251–1254. [Google Scholar] [CrossRef] [PubMed]
- Casals, J. Relationships among Arthropod-Borne Animal Viruses Determined by Cross-Challenge Tests. Am. J. Trop. Med. Hyg. 1963, 12, 587–596. [Google Scholar] [CrossRef]
- Rodriguez-Barraquer, I.; Costa, F.; Nascimento, E.J.M.; Nery, N.J.; Castanha, P.M.S.; Sacramento, G.A.; Cruz, J.; Carvalho, M.; De Olivera, D.; Hagan, J.E.; et al. Impact of preexisting dengue immunity on Zika virus emergence in a dengue endemic region. Science 2019, 363, 607–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niederman, J.C.; Henderson, J.R.; Opton, E.M.; Black, F.L.; Skvrnova, K. A nationwide serum survey of Brazilian military recruits, 1964. II. Antibody patterns with arboviruses, polioviruses, measles and mumps. Am. J. Epidemiol. 1967, 86, 319–329. [Google Scholar] [CrossRef] [PubMed]
- De Paula Pinheiro, F.; Schatzmayr, H.; De Andrade Travassos Da Rosa, A.P.; Homma, A.; Bensabath, G. Arbovirus antibodies in children of rural Guanabara, Brazil. Intervirology 1975, 5, 93–96. [Google Scholar] [CrossRef] [Green Version]
- Iversson, L.B.; da Rosa, A.P.; da Rosa, J.T.; Costa Cda, S. Serological studies for research on arbovirus antibodies in a human population of the Vale do Ribeira region. III. Survey in inhabitants with cases of Flavivirus Rocio encephalitis. Rev. Saude Publica 1982, 16, 160–170. [Google Scholar] [CrossRef]
- Iversson, L.B.; Travassos da Rosa, A.P.; Travassos da Rosa, J.F.; Pinto, G.H.; Macedo, O. Serological studies in research on arbovirus antibodies in a human population of the Ribeira Valley region. IV—Survey of school children living in Iguape County, SP (Brazil). Rev. Saude Publica 1983, 17, 423–435. [Google Scholar] [CrossRef]
- Iversson, L.B.; da Rosa, A.P.; de Rosa, J.T. Serological studies in research on arbovirus antibodies in the human population of the Ribeira Valley region. II—Survey of patients of Pariquera-Acu Regional Hospital, 1980. Rev. Saude Publica 1981, 15, 587–602. [Google Scholar] [CrossRef] [Green Version]
- Iversson, L.B.; Coimbra, T.L. Encephalitis in the Valley of Ribeira region, Sao Paulo, Brazil, in the post-endemic period from 1978 to 1983: Status of the etiological diagnosis and epidemiological characteristics. Rev. Saude Publica 1984, 18, 323–332. [Google Scholar] [CrossRef] [Green Version]
- Romano-Lieber, N.S.; Iversson, L.B. Serological survey on arbovirus infection in residents of an ecological reserve. Rev. Saude Publica 2000, 34, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Tavares-Neto, J.; Travassos da Rosa, A.P.; Vasconcelos, P.F.; Costa, J.M.; Travassos da Rosa, J.F.; Marsden, P.D. Research on antibodies to arbovirus in the serum of residents of the village of Corte de Pedra, Valencia, Bahia. Mem. Inst. Oswaldo Cruz 1986, 81, 351–358. [Google Scholar] [CrossRef] [Green Version]
- Rosemberg, S. Neuropathology of S. Paulo south coast epidemic encephalitis (Rocio flavivurus). J. Neurol. Sci. 1980, 45, 1–12. [Google Scholar] [CrossRef]
- Harrison, A.K.; Murphy, F.A.; Gardner, J.J.; Bauer, S.P. Myocardial and pancreatic necrosis induced by Rocio virus, a new flavivirus. Exp. Mol. Pathol. 1980, 32, 102–113. [Google Scholar] [CrossRef]
- De Barros, V.E.; Saggioro, F.P.; Neder, L.; de Oliveira Franca, R.F.; Mariguela, V.; Chavez, J.H.; Penharvel, S.; Forjaz, J.; da Fonseca, B.A.; Figueiredo, L.T. An experimental model of meningoencephalomyelitis by Rocio flavivirus in BALB/c mice: Inflammatory response, cytokine production, and histopathology. Am. J. Trop. Med. Hyg. 2011, 85, 363–373. [Google Scholar] [CrossRef] [Green Version]
- Chavez, J.H.; Franca, R.F.; Oliveira, C.J.; de Aquino, M.T.; Farias, K.J.; Machado, P.R.; de Oliveira, T.F.; Yokosawa, J.; Soares, E.G.; da Silva, J.S.; et al. Influence of the CCR-5/MIP-1 alpha axis in the pathogenesis of Rocio virus encephalitis in a mouse model. Am. J. Trop. Med. Hyg. 2013, 89, 1013–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henriques, D.F.; Quaresma, J.A.; Fuzii, H.T.; Nunes, M.R.; Silva, E.V.; Carvalho, V.L.; Martins, L.C.; Casseb, S.M.; Chiang, J.O.; Vasconcelos, P.F. Persistence of experimental Rocio virus infection in the golden hamster (Mesocricetus auratus). Mem. Inst. Oswaldo Cruz 2012, 107, 630–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amarilla, A.A.; Santos-Junior, N.N.; Figueiredo, M.L.; Luiz, J.P.M.; Fumagalli, M.J.; Colon, D.F.; Lippi, V.; Alfonso, H.L.; Lima-Junior, D.S.; Trabuco, A.C.; et al. CCR2 Plays a Protective Role in Rocio Virus-Induced Encephalitis by Promoting Macrophage Infiltration Into the Brain. J. Infect. Dis. 2019, 219, 2015–2025. [Google Scholar] [CrossRef]
- Calberg-Bacq, C.M.; Rentier-Delrue, F.; Osterrieth, P.M.; Duchesne, P.Y. Electron microscopy studies on Banzi virus particle and its development in the suckling mice brains. J. Ultrastruct. Res. 1975, 53, 193–203. [Google Scholar] [CrossRef]
- Oyanagi, S.; Ikuta, F.; Ross, E.R. Electron microscopic observations in mice infected with Japanese encephalitis. Acta Neuropathol. 1969, 13, 169–191. [Google Scholar] [CrossRef] [PubMed]
- Murphy, F.A.; Harrison, A.K.; Gary, G.W., Jr.; Whitfield, S.G.; Forrester, F.T. St. Louis encephalitis virus infection in mice. Electron microscopic studies of central nervous system. Lab Invest. 1968, 19, 652–662. [Google Scholar]
- De Morais Bronzoni, R.V.; Baleotti, F.G.; Ribeiro Nogueira, R.M.; Nunes, M.; Moraes Figueiredo, L.T. Duplex reverse transcription-PCR followed by nested PCR assays for detection and identification of Brazilian alphaviruses and flaviviruses. J. Clin. Microbiol. 2005, 43, 696–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romeiro, M.F.; Souza, W.M.; Tolardo, A.L.; Vieira, L.C.; Colombo, T.E.; Aquino, V.H.; Nogueira, M.L.; Figueiredo, L.T. Evaluation and optimization of SYBR Green real-time reverse transcription polymerase chain reaction as a tool for diagnosis of the Flavivirus genus in Brazil. Rev. Soc. Bras. Med. Trop. 2016, 49, 279–285. [Google Scholar] [CrossRef]
- De Puig, H.; Bosch, I.; Collins, J.J.; Gehrke, L. Point-of-Care Devices to Detect Zika and Other Emerging Viruses. Annu. Rev. Biomed. Eng. 2020, 22, 371–386. [Google Scholar] [CrossRef] [PubMed]
- Bosch, I.; de Puig, H.; Hiley, M.; Carre-Camps, M.; Perdomo-Celis, F.; Narvaez, C.F.; Salgado, D.M.; Senthoor, D.; O’Grady, M.; Phillips, E.; et al. Rapid antigen tests for dengue virus serotypes and Zika virus in patient serum. Sci. Transl. Med. 2017, 9, 1589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopes Ode, S.; Sacchetta Lde, A.; Nassar Eda, S.; de Oliveira, M.I.; Bisordi, I.; Suzuki, A.; Kimura, E.K. Serological evaluation of vaccine against human encephalitis caused by Rocio virus. Rev. Inst. Med. Trop. Sao Paulo 1980, 22, 108–113. [Google Scholar] [PubMed]
- Sacchetto, L.; Drumond, B.P.; Han, B.A.; Nogueira, M.L.; Vasilakis, N. Re-emergence of yellow fever in the neotropics quo vadis? Emerg. Top. Life Sci. 2020, 4, 399–410. [Google Scholar] [CrossRef] [PubMed]
- Iversson, L.B.; Silva, R.A.; da Rosa, A.P.; Barros, V.L. Circulation of eastern equine encephalitis, western equine encephalitis, Ilheus, Maguari and Tacaiuma viruses in equines of the Brazilian Pantanal, South America. Rev. Inst. Med. Trop. Sao Paulo 1993, 35, 355–359. [Google Scholar] [CrossRef]
- Han, B.A.; Majumdar, S.; Calmon, F.P.; Glicksberg, B.S.; Horesh, R.; Kumar, A.; Perer, A.; von Marschall, E.B.; Wei, D.; Mojsilovic, A.; et al. Confronting data sparsity to identify potential sources of Zika virus spillover infection among primates. Epidemics 2019, 27, 59–65. [Google Scholar] [CrossRef]
- Evans, M.V.; Dallas, T.A.; Han, B.A.; Murdock, C.C.; Drake, J.M. Data-driven identification of potential Zika virus vectors. Elife 2017, 6, e22053. [Google Scholar] [CrossRef]
- Han, B.A.; O’Regan, S.M.; Paul Schmidt, J.; Drake, J.M. Integrating data mining and transmission theory in the ecology of infectious diseases. Ecol. Lett. 2020, 23, 1178–1188. [Google Scholar] [CrossRef] [PubMed]
- Kaul, R.B.; Evans, M.V.; Murdock, C.C.; Drake, J.M. Spatio-temporal spillover risk of yellow fever in Brazil. Parasites Vectors 2018, 11, 488. [Google Scholar] [CrossRef] [PubMed]
- Childs, J.E.; Gordon, E.R. Surveillance and control of zoonotic agents prior to disease detection in humans. Mt. Sinai J. Med. 2009, 76, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Soper, F.L.; Wilson, D.B.; Lima, S.; Antunes, W.S. The Organization of Permanent Nationwide Anti-Aedes Aegypti Measures in Brazil; The Rockefeller Foundation: New York, NY, USA, 1943. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | State | # of Cases | Diagnostic Tests Performed | Reference |
|---|---|---|---|---|
| 1974–1977 | São Paulo | >1000 | Blood and serology (HI, CF) | [2,3,7] |
| 1978–1983 | São Paulo | 4 | Blood and serology (HI) | [58] |
| 1984 | Bahia | 1 | Blood and serology (neutralization test, HI) | [60] |
| 1987 | São Paulo | 2 | Blood and serology (neutralization test, MAC-ELISA) | [15] |
| 1990 | São Paulo | 6 | Blood and serology (neutralization test, HI) | [59] |
| 1995 | Bahia | 8 | Blood and serology (neutralization test, HI, MAC-ELISA) | [14] |
| 2012–2013 | Goiás | 2 | Blood, molecular (RT-PCR) | [9] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saivish, M.V.; Gomes da Costa, V.; de Lima Menezes, G.; Alves da Silva, R.; Dutra da Silva, G.C.; Moreli, M.L.; Sacchetto, L.; Pacca, C.C.; Vasilakis, N.; Nogueira, M.L. Rocio Virus: An Updated View on an Elusive Flavivirus. Viruses 2021, 13, 2293. https://doi.org/10.3390/v13112293
Saivish MV, Gomes da Costa V, de Lima Menezes G, Alves da Silva R, Dutra da Silva GC, Moreli ML, Sacchetto L, Pacca CC, Vasilakis N, Nogueira ML. Rocio Virus: An Updated View on an Elusive Flavivirus. Viruses. 2021; 13(11):2293. https://doi.org/10.3390/v13112293
Chicago/Turabian StyleSaivish, Marielena Vogel, Vivaldo Gomes da Costa, Gabriela de Lima Menezes, Roosevelt Alves da Silva, Gislaine Celestino Dutra da Silva, Marcos Lázaro Moreli, Livia Sacchetto, Carolina Colombelli Pacca, Nikos Vasilakis, and Maurício Lacerda Nogueira. 2021. "Rocio Virus: An Updated View on an Elusive Flavivirus" Viruses 13, no. 11: 2293. https://doi.org/10.3390/v13112293