
CRISPR Screening: Molecular Tools for Studying Virus–Host Interactions

 , , , and
, , , and
Abstract
:1. Introduction
2. CRISPR/Cas-Based Molecular Tools Used for CRISPR Screens
3. Types of CRISPR Screens
3.1. Pooled and Arrayed CRISPR Screens
3.2. Loss-of-Function and Gain-of-Function CRISPR Screens
3.3. Classical and Combinatorial CRISPR Screening
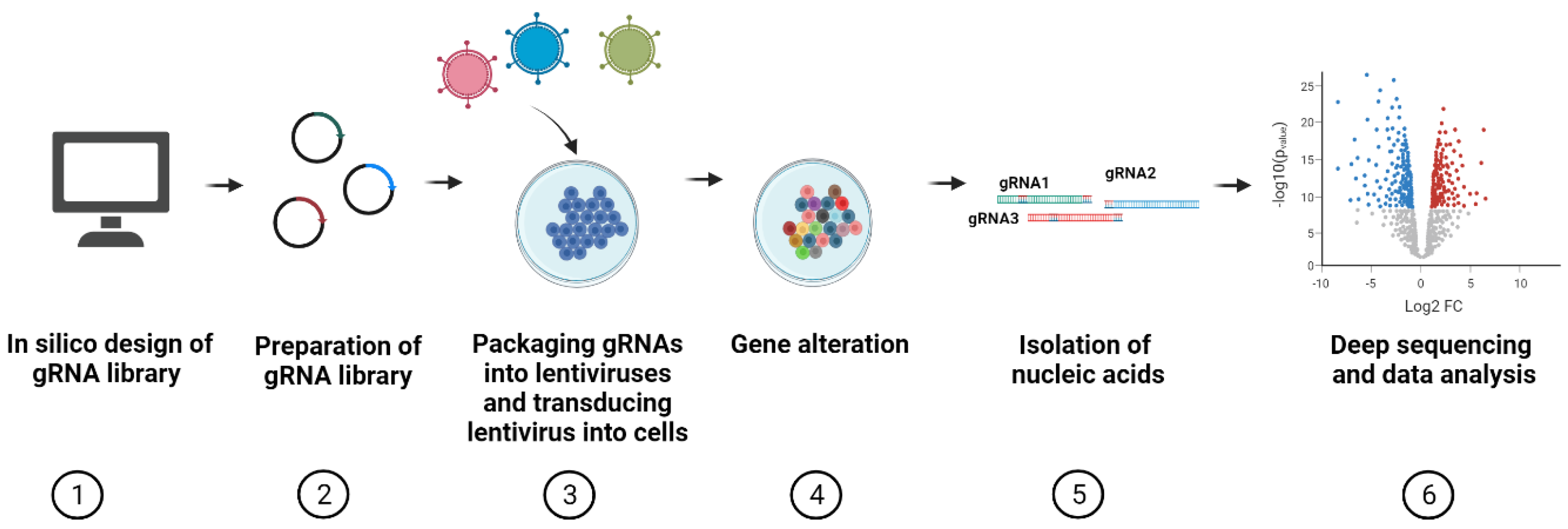
4. Workflow of CRISPR Pooled Screening
4.1. Design of gRNAs for CRISPR Screens
4.2. Available sgRNA Libraries for CRISPR Screens
4.3. Construction of Cas9- and gRNA-Encoding Vectors
4.4. Lentiviral Transduction
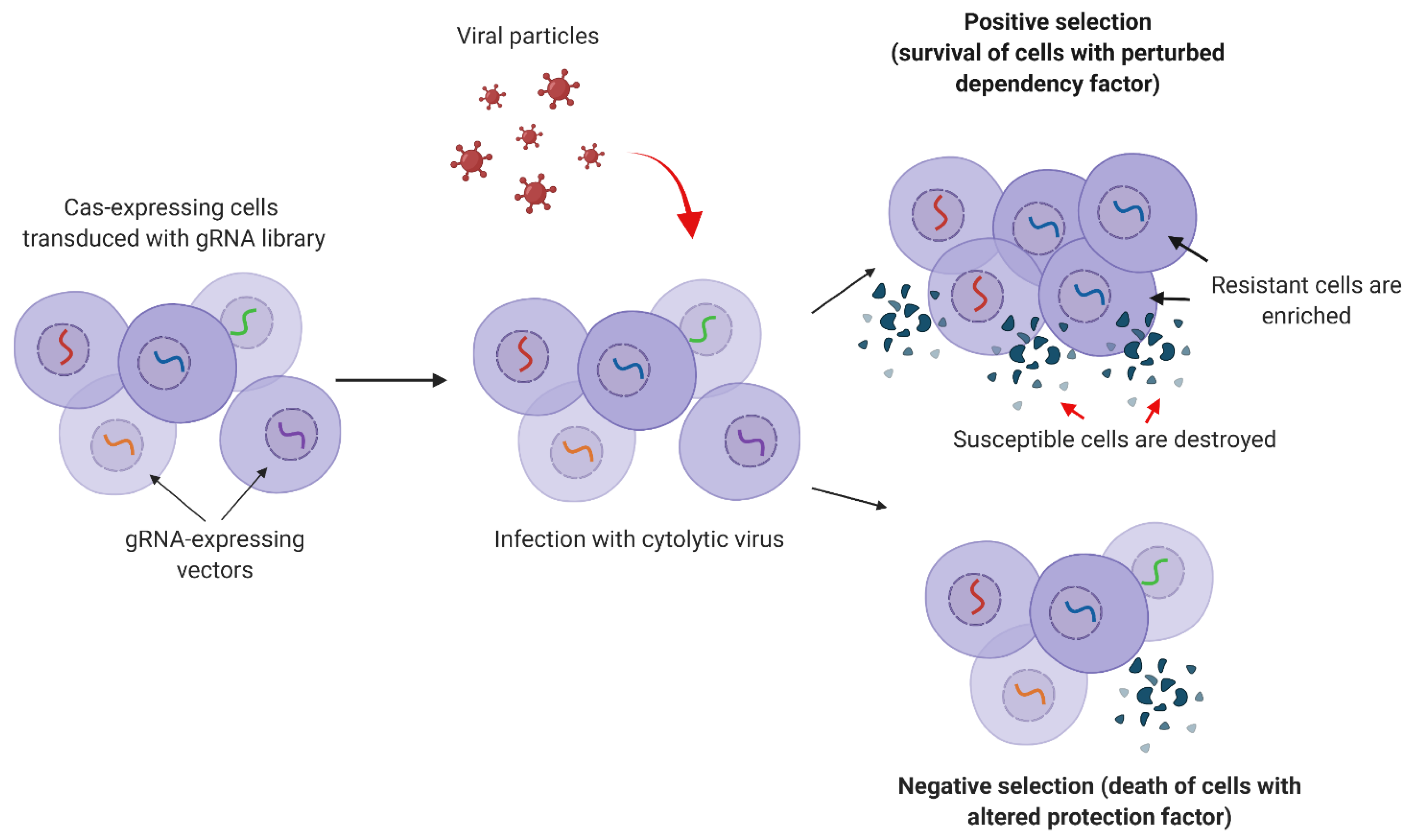
4.5. Selection
4.6. Sequencing
4.7. Data Analysis
5. CRISPR Screens for Studying Viral Infections
5.1. CRISPR Screens for Studying Virus–Host Interactions
5.1.1. Virus Entry and Transmission
5.1.2. Viral Replication
5.1.3. Viral Protein Stability
5.2. CRISPR Screens for Identifying New Antiviral Targets
5.2.1. Latency
5.2.2. Entry
5.2.3. Protein Translation
5.3. Immunity Studies
6. Conclusions
Funding
Conflicts of Interest
References
- Brezgin, S.; Kostyusheva, A.; Kostyushev, D.; Chulanov, V. Dead Cas Systems: Types, Principles, and Applications. Int. J. Mol. Sci. 2019, 20, 6041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brezgin, S.; Kostyusheva, A.; Bayurova, E.; Volchkova, E.; Gegechkori, V.; Gordeychuk, I.; Glebe, D.; Kostyushev, D.; Chulanov, V. Immunity and Viral Infections: Modulating Antiviral Response via CRISPR–Cas Systems. Viruses 2021, 13, 1373. [Google Scholar] [CrossRef] [PubMed]
- Parnas, O.; Jovanovic, M.; Eisenhaure, T.M.; Herbst, R.H.; Dixit, A.; Ye, C.J.; Przybylski, D.; Platt, R.J.; Tirosh, I.; Sanjana, E.N.; et al. A genome-wide CRISPR screen in primary immune cells to dissect regulatory networks. Cell 2015, 162, 675–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zetsche, B.; Gootenberg, J.S.; Abudayyeh, O.O.; Slaymaker, I.M.; Makarova, K.S.; Essletzbichler, P.; Volz, S.E.; Joung, J.; van der Oost, J.; Regev, A.; et al. Cpf1 Is a Single RNA-Guided Endonuclease of a Class 2 CRISPR-Cas System. Cell 2015, 163, 759–771. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Li, J.; Wang, Y.; Yang, B.; Wei, J.; Wu, J.; Wang, R.; Huang, X.; Chen, J.; Yang, L. Efficient base editing in methylated regions with a human APOBEC3A-Cas9 fusion. Nat. Biotechnol. 2018, 36, 946. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Leete, T.C.; Born, D.A.; Young, L.; Barrera, L.A.; Lee, S.-J.; Rees, H.A.; Ciaramella, G.; Gaudelli, N.M. Cytosine base editors with minimized unguided DNA and RNA off-target events and high on-target activity. Nat. Commun. 2020, 11, 1–10. [Google Scholar] [CrossRef]
- Ma, Y.; Zhang, J.; Yin, W.; Zhang, Z.; Song, Y.; Chang, X. Targeted AID-mediated mutagenesis (TAM) enables efficient genomic diversification in mammalian cells. Nat. Methods. 2016, 13, 1029. [Google Scholar] [CrossRef] [PubMed]
- Friedman, J.R.; Fredericks, W.J.; Jensen, D.E.; Speicher, D.W.; Huang, X.-P.; Neilson, E.G.; Rauscher, F.J., 3rd. KAP-1, a novel corepressor for the highly conserved KRAB repression domain. Genes. Dev. 1996, 10, 2067–2078. [Google Scholar] [CrossRef] [Green Version]
- Parsi, K.M.; Hennessy, E.; Kearns, N.; Maehr, R. Using an inducible CRISPR-dCas9-KRAB effector system to dissect transcriptional regulation in human embryonic stem cells. Methods Mol. Biol. 2017, 1507, 221–233. [Google Scholar] [CrossRef]
- Chen, X.; Wei, M.; Liu, X.; Song, S.; Wang, L.; Yang, X.; Song, Y. Construction and validation of the CRISPR/dCas9-EZH2 system for targeted H3K27Me3 modification. Biochem. Biophys. Res. Commun. 2019, 511, 246–252. [Google Scholar] [CrossRef]
- Yeo, N.C.; Chavez, A.; Lance-Byrne, A.; Chan, Y.; Menn, D.; Milanova, D.; Kuo, C.C.; Guo, X.; Sharma, S.; Tung, A.; et al. An enhanced CRISPR repressor for targeted mammalian gene regulation. Nat. Methods 2018, 15, 611. [Google Scholar] [CrossRef]
- Huang, Y.-H.; Su, J.; Lei, Y.; Brunetti, L.; Gundry, M.C.; Zhang, X.; Jeong, M.; Li, W.; Goodell, M.A. DNA epigenome editing using CRISPR-Cas SunTag-directed DNMT3A. Genome Biol. 2017, 18, 176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stepper, P.; Kungulovski, G.; Jurkowska, R.Z.; Chandra, T.; Krueger, F.; Reinhardt, R.; Reik, W.; Jeltsch, A.; Jurkowski, T.P. Efficient targeted DNA methylation with chimeric dCas9—Dnmt3a–Dnmt3L Methyltransferase. Nucleic. Acids Res. 2018, 45, 1703–1713. [Google Scholar] [CrossRef] [PubMed]
- Chedin, F.; Lieber, M.R.; Hsieh, C.-L. The DNA methyltransferase-like protein DNMT3L stimulates de novo methylation by Dnmt3a. Proc. Natl. Acad. Sci. USA 2002, 99, 16916–16921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.U.; Su, Y.; Zhong, C.; Ming, G.; Song, H. Hydroxylation of 5-Methylcytosine by TET1 Promotes Active DNA Demethylation in the Adult Brain. Cell 2011, 145, 423–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chavez, A.; Scheiman, J.; Vora, S.; Pruitt, B.W.; Tuttle, M.; Iyer, E.P.R.; Lin, S.; Kiani, S.; Guzman, C.D.; Wiegand, D.J.; et al. Highly efficient Cas9-mediated transcriptional programming. Nat. Methods 2015, 12, 326. [Google Scholar] [CrossRef] [Green Version]
- Fang, L.; Hung, S.S.C.; Yek, J.; El Wazan, L.; Nguyen, T.; Khan, S.; Lim, S.Y.; Hewitt, A.W.; Wong, R.C.B. A Simple Cloning-free Method to Efficiently Induce Gene Expression Using CRISPR/Cas9. Mol. Ther. Nucleic Acids 2019, 14, 184–191. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yin, C.; Zhang, T.; Li, F.; Yang, W.; Kaminski, R.; Fagan, P.R.; Putatunda, R.; Young, W.B.; Khalili, K.; et al. CRISPR/gRNA-directed synergistic activation mediator (SAM) induces specific, persistent and robust reactivation of the HIV-1 latent reservoirs. Sci. Rep. 2015, 5, 16277. [Google Scholar] [CrossRef]
- Ji, H.; Jiang, Z.; Lu, P.; Ma, L.; Li, C.; Pan, H.; Fu, Z.; Qu, X.; Wang, P.; Deng, J.; et al. Specific reactivation of latent HIV-1 by dCas9-SunTag-VP64-mediated guide RNA targeting the HIV-1 promoter. Mol. Ther. 2016, 24, 508–521. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.B.; Komor, A.C.; Levy, J.M.; Packer, M.S.; Zhao, K.T.; Liu, D.R. Increasing the genome-targeting scope and precision of base editing with engineered Cas9-cytidine deaminase fusions. Nat. Biotechnol. 2017, 35, 371. [Google Scholar] [CrossRef]
- Rees, H.A.; Liu, D.R. Base editing: Precision chemistry on the genome and transcriptome of living cells. Nat. Rev. Genet. 2018, 19, 770–788. [Google Scholar] [CrossRef] [PubMed]
- Thakore, P.I.; Ippolito, A.M.D.; Song, L.; Safi, A.; Shivakumar, N.K.; Kabadi, A.M.; Reddy, T.E.; Crawford, G.E.; Gersbach, C.A. Highly specific epigenome editing by CRISPR-Cas9 repressors for silencing of distal regulatory elements. Nat. Methods 2015, 12, 1143–1149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuñez, J.K.; Chen, J.; Pommier, G.C.; Cogan, J.Z.; Replogle, J.M.; Adriaens, C.; Ramadoss, G.N.; Shi, Q.; Hung, K.L.; Samelson, A.J.; et al. Genome-wide programmable transcriptional memory by CRISPR-based epigenome editing. Cell 2021, 184, 2503–2519. [Google Scholar] [CrossRef] [PubMed]
- Brocken, D.J.W.; Tark-Dame, M.; Dame, R.T. dCas9: A Versatile Tool for Epigenome Editing. Curr. Issues Mol. Biol. 2018, 26, 15–32. [Google Scholar] [CrossRef] [Green Version]
- Adli, M. The CRISPR tool kit for genome editing and beyond. Nat. Commun. 2018, 9, 1911. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Qi, L.S. A CRISPR–dCas toolbox for genetic engineering and synthetic biology. J. Mol. Biol. 2019, 431, 34–47. [Google Scholar] [CrossRef]
- Tan, J.; Martin, S.E. Validation of synthetic CRISPR reagents as a tool for arrayed functional genomic screening. PLoS ONE 2016, 11, e0168968. [Google Scholar] [CrossRef] [Green Version]
- Billon, P.; Bryant, E.E.; Joseph, S.A.; Nambiar, T.S.; Hayward, S.B.; Rothstein, R.; Ciccia, A. CRISPR-Mediated Base Editing Enables Efficient Disruption of Eukaryotic Genes through Induction of STOP Codons. Mol. Cell 2017, 67, 1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuscu, C.; Parlak, M.; Tufan, T.; Yang, J.; Szlachta, K.; Wei, X.; Mammadov, R.; Adli, M. CRISPR-STOP: Gene silencing through base-editing-induced nonsense mutations. Nat. Methods 2017, 14, 710. [Google Scholar] [CrossRef]
- López-Ferreras, L.; Martínez-García, N.; Maeso-Alonso, L.; Martín-López, M.; Díez-Matilla, Á.; Villoch-Fernandez, J.; Alonso-Olivares, H.; Marques, M.M.; Marin, M.C. Deciphering the nature of Trp73 isoforms in mouse embryonic stem cell models: Generation of isoform-specific deficient cell lines using the CRISPR/Cas9 gene editing system. Cancers 2021, 13, 3182. [Google Scholar] [CrossRef]
- Cuella-Martin, R.; Hayward, S.B.; Fan, X.; Chen, X.; Huang, J.-W.; Taglialatela, A.; Leuzzi, G.; Zhao, J.; Rabadan, R.; Lu, C.; et al. Functional interrogation of DNA damage response variants with base editing screens. Cell 2021, 184, 1081–1097. [Google Scholar] [CrossRef]
- Lin, J.; Wu, S.; Shen, Q.; Liu, J.; Huang, S.; Peng, G.; Qiao, Y. Base editing-mediated perturbation of endogenous PKM1/2 splicing facilitates isoform-specific functional analysis in vitro and in vivo. Cell Prolif. 2021, 54, e13096. [Google Scholar] [CrossRef]
- Anzalone, A.V.; Koblan, L.W.; Liu, D.R. Genome editing with CRISPR–Cas nucleases, base editors, transposases and prime editors. Nat. Biotechnol. 2020, 38, 824–844. [Google Scholar] [CrossRef] [PubMed]
- Park, R.J.; Wang, T.; Koundakjian, D.; Hultquist, J.F.; Lamothe-Molina, P.; Monel, B.; Schumann, K.; Yu, H.; Krupzcak, K.M.; Garcia-Beltran, W.; et al. A genome-wide CRISPR screen identifies a restricted set of HIV host dependency factors. Nat. Genet. 2017, 49, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Krey, K.; Babnis, A.W.; Pichlmair, A. System-Based Approaches to Delineate the Antiviral Innate Immune Landscape. Viruses 2020, 12, 1196. [Google Scholar] [CrossRef] [PubMed]
- Tanenbaum, M.E.; Gilbert, L.A.; Qi, L.S.; Weissman, J.S.; Vale, R.D. A Protein-Tagging System for Signal Amplification in Gene Expression and Fluorescence Imaging. Cell 2014, 159, 635–646. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, L.A.; Horlbeck, M.A.; Adamson, B.; Villalta, J.E.; Chen, Y.; Whitehead, E.H.; Guimaraes, C.; Panning, B.; Ploegh, H.L.; Bassik, M.C.; et al. Genome-scale CRISPR-mediated control of gene repression and activation. Cell 2014, 159, 647–661. [Google Scholar] [CrossRef] [Green Version]
- Fortin, J.-P.; Tan, J.; Gascoigne, K.E.; Haverty, P.M.; Forrest, W.F.; Costa, M.R.; Martin, S.E. Multiple-gene targeting and mismatch tolerance can confound analysis of genome-wide pooled CRISPR screens. Genome. Biol. 2019, 20, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Bodapati, S.; Daley, T.P.; Lin, X.; Zou, J.; Qi, L.S. A benchmark of algorithms for the analysis of pooled CRISPR screens. Genome. Biol. 2020, 21, 1–13. [Google Scholar] [CrossRef] [Green Version]
- DeWeirdt, P.C.; Sanson, K.R.; Sangree, A.K.; Hegde, M.; Hanna, R.E.; Feeley, M.N.; Griffith, A.L.; Teng, T.; Borys, S.M.; Strand, C.; et al. Optimization of AsCas12a for combinatorial genetic screens in human cells. Nat. Biotechnol. 2021, 39, 94–104. [Google Scholar] [CrossRef]
- Campa, C.C.; Weisbach, N.R.; Santinha, A.J.; Incarnato, D.; Platt, R.J. Multiplexed genome engineering by Cas12a and CRISPR arrays encoded on single transcripts. Nat. Methods. 2019, 16, 887–893. [Google Scholar] [CrossRef]
- Zhou, P.; Chan, B.K.C.; Wan, Y.K.; Wong, K.H.; Chan, H.Y.E.; Wong, A.S.L. Article A Three-Way Combinatorial CRISPR Screen for Analyzing Interactions among Druggable Targets ll A Three-Way Combinatorial CRISPR Screen for Analyzing Interactions among Druggable Targets. CellReports 2020, 32, 108020. [Google Scholar] [CrossRef]
- Hartenian, E.; Doench, J.G. Genetic screens and functional genomics using CRISPR/Cas9 technology. FEBS J 2015, 282, 1383–1393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.; Xiao, T.; Chen, C.-H.; Li, W.; Meyer, C.A.; Wu, Q.; Cong, L.; Zhang, F.; Liu, J.S. Sequence determinants of improved CRISPR sgRNA design. Genome. Res. 2015, 25, 1147–1157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doench, J.G.; Hartenian, E.; Graham, D.B.; Tothova, Z.; Hegde, M.; Smith, I.; Sullender, M.; Ebert, B.L.; Xavier, R.J.; Root, D.E. Rational design of highly active sgRNAs for CRISPR-Cas9–mediated gene inactivation. Nat. Biotechnol. 2014, 32, 1262–1267. [Google Scholar] [CrossRef] [Green Version]
- Doench, J.G.; Fusi, N.; Sullender, M.; Hegde, M.; Vaimberg, E.W.; Donovan, K.F.; Smith, I.; Tothova, Z.; Wilen, C.; Orchard, R.; et al. Optimized sgRNA design to maximize activity and minimize off-target effects of CRISPR-Cas9. Nat. Biotechnol. 2016, 34, 184. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Mateos, M.A.; Vejnar, C.E.; Beaudoin, J.; Fernandez, J.P.; Mis, E.K.; Khokha, M.K.; Giraldex, A.J. CRISPRscan: Designing highly efficient sgRNAs for CRISPR-Cas9 targeting in vivo. Nat. Methods 2015, 12, 982–988. [Google Scholar] [CrossRef] [Green Version]
- Labuhn, M.; Adams, F.F.; Ng, M.; Knoess, S.; Schambach, A.; Charpentier, E.M.; Schwarzer, A.; Mateo, J.L.; Klusmann, J.H.; Heckl, D. Refined sgRNA efficacy prediction improves large-and small-scale CRISPR–Cas9 applications. Nucleic Acids Res. 2018, 46, 1375–1385. [Google Scholar] [CrossRef] [Green Version]
- Labun, K.; Montague, T.G.; Krause, M.; Torres Cleuren, Y.N.; Tjeldnes, H.; Valen, E. CHOPCHOP v3: Expanding the CRISPR web toolbox beyond genome editing. Nucleic Acids Res. 2019, 47, W171–W174. [Google Scholar] [CrossRef] [Green Version]
- Heigwer, F.; Kerr, G.; Boutros, M. E-CRISP: Fast CRISPR target site identification. Nat Methods 2014, 11, 122–123. [Google Scholar] [CrossRef]
- Haeussler, M.; Schönig, K.; Eckert, H.; Eschstruth, A.; Mianné, J.; Renaud, J.-B.; Schneider-Maunoury, S.; Shkumatava, A.; Teboul, L.; Kent, J.; et al. Evaluation of off-target and on-target scoring algorithms and integration into the guide RNA selection tool CRISPOR. Genome. Biol. 2016, 17, 1–12. [Google Scholar] [CrossRef]
- Stemmer, M.; Thumberger, T.; del Sol Keyer, M.; Wittbrodt, J.; Mateo, J.L. CCTop: An intuitive, flexible and reliable CRISPR/Cas9 target prediction tool. PLoS ONE 2015, 10, e0124633. [Google Scholar]
- Oliveros, J.C.; Franch, M.; Tabas-Madrid, D.; San-León, D.; Montoliu, L.; Cubas, P.; Pazos, F. Breaking-Cas—interactive design of guide RNAs for CRISPR-Cas experiments for ENSEMBL genomes. Nucleic Acids Res. 2016, 44, W267–W271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez, A.R.; Pritykin, Y.; Vidigal, J.A.; Chhangawala, S.; Zamparo, L.; Leslie, C.S.; Ventura, A. GuideScan software for improved single and paired CRISPR guide RNA design. Nat. Biotechnol. 2017, 35, 347–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Wang, S.; Luo, F.; Wu, F.-X.; Wang, J. MultiGuideScan: A multi-processing tool for designing CRISPR guide RNA libraries. Bioinformatics 2020, 36, 920–921. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Köster, J.; Qin, Q.; Hu, S.; Li, W.; Chen, C.; Cao, Q.; Wang, J.; Mei, S.; Liu, Q.; et al. CRISPR-DO for genome-wide CRISPR design and optimization. Bioinformatics 2016, 32, 3336–3338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, N.; Liu, W.; Wang, X. WU-CRISPR: Characteristics of functional guide RNAs for the CRISPR/Cas9 system. Genome Biol. 2015, 16, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.-L.; Rodiger, J.; Chung, V.; Viswanatha, R.; Mohr, S.E.; Hu, Y.; Perrimon, N. SNP-CRISPR: A web tool for SNP-specific genome editing. G3 Genes Genomes Genet. 2020, 10, 489–494. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.; Xu, J.; Cheng, M.; Liao, X.; Peng, S. Review of CRISPR/Cas9 sgRNA design tools. Interdiscip. Sci. Comput. Life Sci. 2018, 10, 455–465. [Google Scholar] [CrossRef]
- Liu, G.; Zhang, Y.; Zhang, T. Computational approaches for effective CRISPR guide RNA design and evaluation. Comput. Struct Biotechnol. J. 2020, 18, 35–44. [Google Scholar] [CrossRef]
- Horlbeck, M.A.; Gilbert, L.A.; Villalta, J.E.; Adamson, B.; Pak, R.A.; Chen, Y.; Fields, A.P.; Park, C.Y.; Corn, J.E.; Kampmann, M.; et al. Compact and highly active next-generation libraries for CRISPR-mediated gene repression and activation. Elife 2016, 5, e19760. [Google Scholar] [CrossRef]
- Kweon, J.; Kim, Y. High-throughput genetic screens using CRISPR–Cas9 system. Arch. Pharm. Res. 2018, 41, 875–884. [Google Scholar] [CrossRef] [PubMed]
- Sanson, K.R.; Hanna, R.E.; Hegde, M.; Donovan, K.F.; Strand, C.; Sullender, M.E.; Vaimberg, E.W.; Goodale, A.; Root, D.E.; Piccioni, F.; et al. Optimized libraries for CRISPR-Cas9 genetic screens with multiple modalities. Nat. Commun. 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, M.K.; Rahman, M.S. CRISPRpred: A flexible and efficient tool for sgRNAs on-target activity prediction in CRISPR/Cas9 systems. PLoS ONE 2017, 12, e0181943. [Google Scholar]
- Peng, H.; Zheng, Y.; Blumenstein, M.; Tao, D.; Li, J. CRISPR/Cas9 cleavage efficiency regression through boosting algorithms and Markov sequence profiling. Bioinformatics 2018, 34, 3069–3077. [Google Scholar] [CrossRef]
- Zhang, D.; Hurst, T.; Duan, D.; Chen, S.-J. Unified energetics analysis unravels SpCas9 cleavage activity for optimal gRNA design. Proc. Natl. Acad. Sci. USA 2019, 116, 8693–8698. [Google Scholar] [CrossRef] [Green Version]
- Hwang, G.-H.; Park, J.; Lim, K.; Kim, S.; Yu, J.; Yu, E.; Kim, S.-T.; Eils, R.; Kim, J.-S.; Bae, S. Web-based design and analysis tools for CRISPR base editing. BMC Bioinform. 2018, 19, 542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clement, K.; Hsu, J.Y.; Canver, M.C.; Joung, J.K.; Pinello, L. Technologies and Computational Analysis Strategies for CRISPR Applications. Mol. Cell 2020, 79, 11–29. [Google Scholar] [CrossRef]
- Shalem, O.; Sanjana, N.E.; Hartenian, E.; Shi, X.; Scott, D.A.; Mikkelsen, T.S.; Heckl, D.; Ebert, B.L.; Root, D.E.; Doench, J.G.; et al. Genome-scale CRISPR-Cas9 knockout screening in human cells. Science 2014, 343, 84–87. [Google Scholar] [CrossRef] [Green Version]
- Sanjana, N.E.; Shalem, O.; Zhang, F. Improved vectors and genome-wide libraries for CRISPR screening. Nat. Methods 2014, 11, 783. [Google Scholar] [CrossRef] [Green Version]
- Hart, T.; Tong, A.H.Y.; Chan, K.; Van Leeuwen, J.; Seetharaman, A.; Aregger, M.; Chandrashekhar, M.; Hustedt, N.; Seth, S.; Noonan, A.; et al. Evaluation and design of genome-wide CRISPR/SpCas9 knockout screens. G3 Genes Genomes Genet. 2017, 7, 2719–2727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Srinivasan, S.; Li, C.-Y.; Ho, I.-L.; Rose, J.; Shaheen, M.; Wang, G.; Yao, W.; Deem, A.; Bristow, C.; et al. Pooled library screening with multiplexed Cpf1 library. Nat. Commun. 2019, 10, 3144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeWeirdt, P.C.; Sangree, A.K.; Hanna, R.E.; Sanson, K.R.; Hegde, M.; Strand, C.; Persky, N.S.; Doench, J.G. Genetic screens in isogenic mammalian cell lines without single cell cloning. Nat. Commun. 2020, 11, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, P.; Liu, Z.; Liu, Y.; Ma, H.; Xu, Y.; Bao, Y.; Zhu, S.; Cao, Z.; Wu, Z.; Zhou, Z.; et al. Genome-wide interrogation of gene functions through base editor screens empowered by barcoded sgRNAs. Nat. Biotechnol. 2021, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Roesch, F.; OhAinle, M.; Emerman, M. A CRISPR screen for factors regulating SAMHD1 degradation identifies IFITMs as potent inhibitors of lentiviral particle delivery. Retrovirology 2018, 15, 26. [Google Scholar] [CrossRef] [Green Version]
- Chong, Z.-S.; Ohnishi, S.; Yusa, K.; Wright, G.J. Pooled extracellular receptor-ligand interaction screening using CRISPR activation. Genome Biol. 2018, 19, 1–16. [Google Scholar] [CrossRef]
- Słabicki, M.; Kozicka, Z.; Petzold, G.; Li, Y.-D.; Manojkumar, M.; Bunker, R.D.; Donovan, K.A.; Sievers, Q.L.; Koeppel, J.; Suchyta, D.; et al. The CDK inhibitor CR8 acts as a molecular glue degrader that depletes cyclin K. Nature 2020, 585, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.; Tong, A.H.Y.; Brown, K.R.; Mero, P.; Moffat, J. Pooled CRISPR-Based Genetic Screens in Mammalian Cells. J. Vis. Exp. 2019, 151, e59780. [Google Scholar] [CrossRef]
- Wang, T.; Lander, E.S.; Sabatini, D.M. Single guide RNA library design and construction. Cold Spring Harb. Protoc. 2016, 2016, pdb-prot090803. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Wang, M.; Zhang, W.; Xiao, T.; Chen, C.-H.; Wu, A.; Wu, F.; Traugh, N.; Wang, X.; Li, Z.; et al. Integrative analysis of pooled CRISPR genetic screens using MAGeCKFlute. Nat. Protoc. 2019, 14, 756–780. [Google Scholar] [CrossRef]
- Li, W.; Xu, H.; Xiao, T.; Cong, L.; Love, M.I.; Zhang, F.; Irizarry, R.A.; Liu, J.S.; Brown, M.; Liu, X.S. MAGeCK enables robust identification of essential genes from genome-scale CRISPR/Cas9 knockout screens. Genome. Biol. 2014, 15, 554. [Google Scholar] [CrossRef]
- Hart, T.; Moffat, J. BAGEL: A computational framework for identifying essential genes from pooled library screens. BMC Bioinform. 2016, 17, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyers, R.M.; Bryan, J.G.; McFarland, J.M.; Weir, B.A.; Sizemore, A.E.; Xu, H. Computational correction of copy number effect improves specificity of CRISPR-Cas9 essentiality screens in cancer cells. Nat. Genet. 2017, 49, 1779–1784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daley, T.P.; Lin, Z.; Lin, X.; Liu, Y.; Wong, W.H.; Qi, L.S. CRISPhieRmix: A hierarchical mixture model for CRISPR pooled screens. Genome Biol. 2018, 19, 159. [Google Scholar] [CrossRef] [Green Version]
- Allen, F.; Behan, F.; Khodak, A.; Iorio, F.; Yusa, K.; Garnett, M.; Parts, L. JACKS: Joint analysis of CRISPR/Cas9 knockout screens. Genome Res. 2019, 29, 464–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Köster, J.; Xu, H.; Chen, C.-H.; Xiao, T.; Liu, J.S.; Brown, M.; Liu, X.S. Quality control, modeling, and visualization of CRISPR screens with MAGeCK-VISPR. Genome Biol. 2015, 16, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joung, J.; Konermann, S.; Gootenberg, J.S.; Abudayyeh, O.O.; Platt, R.J.; Brigham, M.D.; Sanjana, N.E.; Zhang, F. Genome-scale CRISPR-Cas9 knockout and transcriptional activation screening. Nat. Protoc. 2017, 12, 828–863. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Hajian, C.; Greene, W.C. Identification of unrecognized host factors promoting HIV-1 latency. PLoS Pathog. 2020, 16, e1009055. [Google Scholar] [CrossRef]
- Yang, X.; Wang, Y.; Lu, P.; Shen, Y.; Zhao, X.; Zhu, Y.; Jiang, Z.; Yang, H.; Pan, H.; Zhao, L.; et al. PEBP 1 suppresses HIV transcription and induces latency by inactivating MAPK/NF-κB signaling. EMBO Rep. 2020, 21, e49305. [Google Scholar] [CrossRef]
- Krasnopolsky, S.; Kuzmina, A.; Taube, R. Genome-wide CRISPR knockout screen identifies ZNF304 as a silencer of HIV transcription that promotes viral latency. PLoS Pathog. 2020, 16, e1008834. [Google Scholar] [CrossRef]
- Rathore, A.; Iketani, S.; Wang, P.; Jia, M.; Sahi, V.; Ho, D.D. CRISPR-based gene knockout screens reveal deubiquitinases involved in HIV-1 latency in two Jurkat cell models. Sci. Rep. 2020, 10, 1–14. [Google Scholar]
- Jin, S.; Liao, Q.; Chen, J.; Zhang, L.; He, Q.; Zhu, H.; Zhang, X.; Xu, J. TSC1 and DEPDC5 regulate HIV-1 latency through the mTOR signaling pathway. Emerg. Microbes. Infect. 2018, 7, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, W.M.; Luna, J.M.; Hoffmann, H.-H.; Sánchez-Rivera, F.J.; Leal, A.A.; Ashbrook, A.W. Genome-scale identification of SARS-CoV-2 and pan-coronavirus host factor networks. Cell 2021, 184, 120–132. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Alfajaro, M.M.; DeWeirdt, P.C.; Hanna, R.E.; Lu-Culligan, W.J.; Cai, W.L. Genome-wide CRISPR screens reveal host factors critical for SARS-CoV-2 infection. Cell 2021, 184, 76–91.e13. [Google Scholar] [CrossRef] [PubMed]
- Daniloski, Z.; Jordan, T.X.; Wessels, H.-H.; Hoagland, D.A.; Kasela, S.; Legut, M.; Maniatis, S.; Mimitou, E.P.; Lu, L.; Geller, E.; et al. Identification of required host factors for SARS-CoV-2 infection in human cells. Cell 2020. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Simoneau, C.R.; Kulsuptrakul, J.; Bouhaddou, M.; Travisano, K.A.; Hayashi, J.M.; Carlson-Stevermer, J.; Zengel, J.R.; Richards, C.M.; Fozouni, P.; et al. Genetic screens identify host factors for SARS-CoV-2 and common cold coronaviruses. Cell 2020, 184, 106–119. [Google Scholar] [CrossRef]
- Labeau, A.; Simon-Loriere, E.; Hafirassou, M.-L.; Bonnet-Madin, L.; Tessier, S.; Zamborlini, A.; Dupré, T.; Seta, N.; Schwartz, O.; Chaix, M.-L.; et al. A genome-wide CRISPR-Cas9 screen identifies the dolichol-phosphate mannose synthase complex as a host dependency factor for dengue virus infection. J. Virol. 2020, 94, e01751-19. [Google Scholar] [CrossRef]
- Lin, D.L.; Cherepanova, N.A.; Bozzacco, L.; MacDonald, M.R.; Gilmore, R.; Tai, A.W. Dengue virus hijacks a noncanonical oxidoreductase function of a cellular oligosaccharyltransferase complex. mBio 2017, 8, e00939-17. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, H.-H.; Schneider, W.M.; Rozen-Gagnon, K.; Miles, L.A.; Schuster, F.; Razooky, B.; Jacobson, E.; Wu, X.; Yi, S.; Rudin, C.M.; et al. TMEM41B is a pan-flavivirus host factor. Cell 2020, 184, 133–148. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, Q.; Tiwari, S.K.; Lichinchi, G.; Yau, E.H.; Hui, H.; Li, W.; Furnari, F.; Rana, T.M. Integrin αvβ5 internalizes Zika virus during neural stem cells infection and provides a promising target for antiviral therapy. Cell Rep. 2020, 30, 969–983. [Google Scholar] [CrossRef] [Green Version]
- Dukhovny, A.; Lamkiewicz, K.; Chen, Q.; Fricke, M.; Jabrane-Ferrat, N.; Marz, M.; Lamkiewicz, K.; Chen, Q.; Fricke, M.; Jabrane-Ferrat, N.; et al. A CRISPR Activation Screen Identifies Genes That Protect against Zika Virus Infection. J. Virol. 2019, 93, e00211-19. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Muffat, J.; Javed, A.O.; Keys, H.R.; Lungjangwa, T.; Bosch, I.; Khan, M.; Virgilio, M.C.; Gehrke, L.; Sabatini, D.M.; et al. Genome-wide CRISPR screen for Zika virus resistance in human neural cells. Proc. Natl. Acad. Sci. USA 2019, 116, 9527–9532. [Google Scholar] [CrossRef] [Green Version]
- Das, A.; Barrientos, R.; Shiota, T.; Madigan, V.; Misumi, I.; McKnight, K.L.; Sun, L.; Li, Z.; Meganck, R.M.; Li, Y.; et al. Gangliosides are essential endosomal receptors for quasi-enveloped and naked hepatitis A virus. Nat. Microbiol. 2020, 5, 1069–1078. [Google Scholar] [CrossRef] [PubMed]
- Hyrina, A.; Jones, C.; Chen, D.; Clarkson, S.; Cochran, N.; Feucht, P.; Hoffman, G.; Lindeman, A.; Russ, C.; Sigoillot, F.; et al. A genome-wide CRISPR screen identifies ZCCHC14 as a host factor required for hepatitis B surface antigen production. Cell Rep. 2019, 29, 2970–2978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, R.; Jiang, C.; Zhang, Y.; Govande, A.; Trudeau, S.J.; Chen, F.; Fry, C.J.; Puri, R.; Wolinsky, E. MYC controls the Epstein-Barr virus lytic switch. Mol. Cell 2020, 78, 653–669. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Jiang, S.; Zhang, L.; Li, D.; Liang, J.; Narita, Y.; Hou, I.; Zhong, Q.; Gewurz, B.E.; Teng, M.; et al. TAF family proteins and MEF2C are essential for Epstein-Barr virus super-enhancer activity. J. Virol. 2019, 93, e00513-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Walsh, M.J.; Bernhardt, K.; Ashbaugh, C.W.; Trudeau, S.J.; Ashbaugh, I.Y.; Jiang, S.; Jiang, C.; Zhao, B.; Root, D.E.; et al. CRISPR/Cas9 screens reveal Epstein-Barr virus-transformed B cell Shost dependency factors. Cell Host Microbe. 2017, 21, 580–591. [Google Scholar] [CrossRef] [Green Version]
- Xiaofei, E.; Meraner, P.; Lu, P.; Perreira, J.M.; Aker, A.M.; McDougall, W.M.; Zhuge, R.; Chan, G.C.; Gerstein, R.M.; Caposio, P.; et al. OR14I1 is a receptor for the human cytomegalovirus pentameric complex and defines viral epithelial cell tropism. Proc. Natl. Acad. Sci. USA 2019, 116, 7043–7052. [Google Scholar]
- Wu, K.; Oberstein, A.; Wang, W.; Shenk, T. Role of PDGF receptor-α during human cytomegalovirus entry into fibroblasts. Proc. Natl. Acad. Sci. USA 2018, 115, E9889–E9898. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Clohisey, S.M.; Chia, B.S.; Wang, B.; Cui, A.; Eisenhaure, T.; Schweitzer, L.D.; Hoover, P.; Parkinson, N.J.; Nachshon, A.; et al. Genome-wide CRISPR screen identifies host dependency factors for influenza A virus infection. Nat. Commun. 2020, 11, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Perez, J.T.; Chen, C.; Li, Y.; Benitez, A.; Kandasamy, M.; Lee, Y.; Andrade, J.; tenOever, B.; Manicassamy, B. Genome-wide CRISPR/Cas9 Screen Identifies Host Factors Essential for Influenza Virus Replication. Cell Rep. 2018, 23, 596–607. [Google Scholar] [CrossRef] [Green Version]
- Dudek, A.M.; Zabaleta, N.; Zinn, E.; Pillay, S.; Zengel, J.; Porter, C.; Franceshini, J.S.; Estelien, R.; Carette, J.E.; Zhou, G.L.; et al. GPR108 is a highly conserved AAV entry factor. Mol. Ther. 2020, 28, 367–381. [Google Scholar] [CrossRef] [PubMed]
- Madigan, V.J.; Tyson, T.O.; Yuziuk, J.A.; Pillai, M.; Moller-Tank, S.; Asokan, A. A CRISPR screen identifies the cell polarity determinant crumbs 3 as an adeno-associated virus restriction factor in hepatocytes. J. Virol. 2019, 93, e00943-19. [Google Scholar] [CrossRef] [PubMed]
- Flint, M.; Chatterjee, P.; Lin, D.L.; McMullan, L.K.; Shrivastava-Ranjan, P.; Bergeron, É. A genome-wide CRISPR screen identifies N-acetylglucosamine-1-phosphate transferase as a potential antiviral target for Ebola virus. Nat. Commun. 2019, 10, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diep, J.; Ooi, Y.S.; Wilkinson, A.W.; Peters, C.E.; Foy, E.; Johnson, J.R.; Zengel, J.; Ding, S.; Weng, K.-F.; Laufman, O.; et al. Enterovirus pathogenesis requires the host methyltransferase SETD3. Nat. Microbiol. 2019, 4, 2523–2537. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Li, J.; Liu, Y.; Qu, Y.; Li, A.; Li, C.; Zhang, Q.; Wu, W.; Li, J.; Liu, Y.; et al. SNX11 identified as an essential host factor for SFTS virus infection by CRISPR knockout screening. Virol. Sin. 2019, 34, 508–520. [Google Scholar] [CrossRef] [PubMed]
- Orchard, R.C.; Sullender, M.E.; Dunlap, B.F.; Balce, D.R.; Doench, J.G.; Virgin, H.W. Identification of antinorovirus genes in human cells using Genome-Wide CRISPR activation screening. J. Virol. 2019, 93, e01324-18. [Google Scholar] [CrossRef] [Green Version]
- Richardson, R.B.; Ohlson, M.B.; Eitson, J.L.; Kumar, A.; McDougal, M.B.; Boys, I.N. A CRISPR screen identifies IFI6 as an ER-resident interferon effector that blocks flavivirus replication. Nat. Publ. Group 2018, 3, 1214–1223. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Miner, J.J.; Gorman, M.J.; Rausch, K.; Ramage, H.; White, J.P.; Zuiani, A.; Zhang, P.; Fernandez, E.; Zhang, Q.; et al. A CRISPR screen defines a signal peptide processing pathway required by flaviviruses. Nature 2016, 535, 164–168. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.; Dang, Y.; Wu, Y.; Jia, G.; Anaya, E.; Zhang, J.; Abraham, S.; Choi, J.-G.; Shi, G.; Qi, L.; et al. A CRISPR-based screen identifies genes essential for West-Nile-virus-induced cell death. Cell Rep. 2015, 12, 673–683. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Kim, A.S.; Fox, J.M.; Nair, S.; Basore, K.; Klimstra, W.B.; Rimkunas, R.; Fong, R.H.; Lin, H.; Poddar, S.; et al. Mxra8 is a receptor for multiple arthritogenic alphaviruses. Nature 2018, 557, 570–574. [Google Scholar] [CrossRef]
- Ding, S.; Diep, J.; Feng, N.; Ren, L.; Li, B.; Ooi, Y.S.; Wang, X.; Brulois, K.F.; Yasukawa, L.L.; Li, X.; et al. STAG2 deficiency induces interferon responses via cGAS-STING pathway and restricts virus infection. Nat. Commun. 2018, 9, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Gordon, D.E.; Jang, G.M.; Bouhaddou, M.; Xu, J.; Obernier, K.; White, K.M.; O’Meara, M.J.; Rezelj, V.V.; Guo, J.Z.; Swaney, D.L.; et al. A SARS-CoV-2 protein interaction map reveals targets for drug repurposing. Nature 2020, 583, 459–468. [Google Scholar] [CrossRef]
- Hoffmann, H.-H.; Sánchez-Rivera, F.J.; Schneider, W.M.; Luna, J.M.; Soto-Feliciano, Y.M.; Ashbrook, A.W. Functional interrogation of a SARS-CoV-2 host protein interactome identifies unique and shared coronavirus host factors. Cell Host Microbe. 2021, 29, 267–280. [Google Scholar] [CrossRef]
- Yang, S.; Zeng, W.; Zhang, J.; Lu, F.; Chang, J.; Guo, J.-T. Restoration of a functional antiviral immune response to chronic HBV infection by reducing viral antigen load: If not sufficient, is it necessary? Emerg. Microbes. Infect. 2021, 10, 1545–1554. [Google Scholar] [CrossRef]
- Matharu, N.; Rattanasopha, S.; Tamura, S.; Maliskova, L.; Wang, Y.; Bernard, A.; Hardin, A.; Echalbar, W.L.; Vaisse, C. CRISPR-mediated activation of a promoter or enhancer rescues obesity caused by haploinsufficiency. Science 2019, 363, 246. [Google Scholar] [CrossRef] [PubMed]
- Saayman, S.M.; Lazar, D.C.; Scott, T.A.; Hart, J.R.; Takahashi, M.; Burnett, J.C. Potent and Targeted Activation of Latent HIV-1 Using the CRISPR/dCas9 Activator Complex. Mol. Ther. 2016, 24, 488–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ophinni, Y.; Inoue, M.; Kotaki, T.; Kameoka, M. CRISPR/Cas9 system targeting regulatory genes of HIV-1 inhibits viral replication in infected T-cell cultures. Sci. Rep. 2018, 8, 7784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- OhAinle, M.; Helms, L.; Vermeire, J.; Roesch, F.; Humes, D.; Basom, R.; Delrow, J.J.; Overbaugh, J.; Emerman, M. A virus-packageable CRISPR screen identifies host factors mediating interferon inhibition of HIV. Elife 2018, 7, e39823. [Google Scholar] [CrossRef]
- Chia, B.S.; Li, B.; Cui, A.; Eisenhaure, T.; Raychowdhury, R.; Lieb, D.; Hacohen, N. Loss of the nuclear protein RTF2 enhances influenza virus replication. J. Virol. 2020, 94, e00319-20. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Type of Screen | Library Name | AddGene Catalog Number | Genes Targeted | gRNAs Per Gene | Total gRNAs |
|---|---|---|---|---|---|
| Genome-Wide Libraries | |||||
| Ko | GeCKO v2 | 1000000048 and 1000000049 | 19,052 | 6 | 123,411 |
| Ko | Toronto v3 | 90294 and 125517 | 18,053 | 4 | 70,948 |
| Ko | Brunello | 73179 and 73178 | 19,114 | 4 | 76,441 |
| Ko | Gattinara | 136986 | 19,993 | 2 | 40,964 |
| Ko | Mini-Human AsCas12a-Based Library | 130630 | 16,997 | 3–4 | 17,032 arrays (3–4 gRNAs per array) |
| Ko | BARBEKO (for screening with a base editor) | 174163 | 17,501 | 3 | 53,502 |
| CRISPRi | Dolcetto | 1000000114 | 18,901 (Set A); 18,899 (Set B) | 3–6 | 57,050 (Set A); 57,011 (Set B) |
| CRISPRi | CRISPRi-v2 | 83969 1000000090 | 18,905 | 5–10 | 104,535 209,070 |
| CRISPRa | Calabrese | 1000000111 | 18,885 (Set A); 18,843 (Set B) | 3–6 | 56,762 (Set A); 56,476 (Set B) |
| CRISPRa | CRISPRa-v2 | 83978 1000000091 | 18,915 | 5 or 10 | 104,540 or 209,080 |
| CRISPRa | SAM Library (3-plasmid system) | 1000000057 and 1000000074 | 23,430 | 3 | 70,290 |
| Pathway-Specific Libraries | |||||
| Ko | Human Interferon-Stimulated Gene CRISPR Knockout Library | 125753 | 1902 | 8 | 15,416 |
| Ko | Bison sgRNA Library (ubiquitination and deubiquitination genes) | 169942 | 713 | 4 | 2852 |
| Ko | Li Human UBDUB CRISPR Knockout Library (ubiquitination and deubiquitination genes) | 171531 | 1500 | ~6 | 9274 |
| CRISPRa | Wright Human Membrane Protein Activation Library (surface proteins) | 113345 | 6213 | 7–14 | 58,570 |
| Virus | Screen Type | gRNA Library | Cells | Top Screening Hits | Ref. |
|---|---|---|---|---|---|
| HIV | CRISPR-ko | Custom library of 187,536 gRNAs targeting 18,543 genes | GXR cell line | TPST2, SLC35B2, ALCAM, CCR5, CD4 | [34] |
| CRISPRi | CRISPRi-v2 | Jurkat | TMEM178A, FTSJ3, INTS2, INTS5, INTS8, NICN1 | [88] | |
| CRISPR-ko | GeCKO | C11 cell line | PEPB1, BRD2, BRD4 | [89] | |
| CRISPR-ko | GeCKO | Jurkat | ZNF304 | [90] | |
| CRISPR-ko | GeCKO v2 | Jurkat (J-Lat 10.6 cell line) | 52 genes including IWS1, POLE3, POLR1B, PSMD1, TGM2 | [91] | |
| CRISPR-ko | GeCKO v2 | Jurkat (HIV-1 latent infection cell line [C11]) | TCS1, DEPDC5, SUV39H1, SPATA6L, NFKB2, and other genes | [92] | |
| Coronaviruses (SARS-CoV-2, MERS) | CRISPR-ko | Brunello | Huh-7.5 | 146 (37 °C) and 171 (33 °C) genes, including TMEM41B for all of investigated viruses | [93] |
| CRISPR-ko | Custom library of 83,963 gRNAs | Vero-E6 | Genes of SWI/SNF complex, ACE2, DPP4, CTSL, PCBD1, KMT2D, SMAD3, HMGB1, and others | [94] | |
| CRISPR-ko | GeCKO v2 | A549ACE2 | RAB7A, CCDC22, VPS35, ACE2, CTSL, and others | [95] | |
| CRISPR-ko | GeCKO v2 | Huh7.5.1 | TMEM106B and other genes | [96] | |
| Dengue virus (DENV) | CRISPR-ko | GeCKO v2 | Haploid HAP1 cells | 17 genes including DPM1 and DPM3 | [97] |
| CRISPR-ko | GeCKOv2 | Huh7.5.1 | STT3A, STT3B, DC2, MAGT, RPN2, OST4 | [98] | |
| Zika virus (ZIKV), yellow fever virus | CRISPR-ko | GeCKO | HAP1 | TMEM41B and VMP1 (overlap between the two viruses) | [99] |
| ZIKV | CRISPR-ko | Brunello | TS576 | CENPH, ITGB5, MYLPH, HOMER1, BAALC, GABBR2, EPHA10, PTNP2, GCNT7, TRAM1, TMEM41B | [100] |
| CRISPRa | LentiSAMv2 | Huh7 | IFI6, IFNL2, ISG20, HELZ2 | [101] | |
| CRISPR-ko | LentiCRISPRv1; custom library | Human neural progenitors cells | TM9SF2, ATP6V1C1, ATP6V1F, SSR2, SSR3, EMC2, EMC6, C3orf58, ISG15, SOCS3, STAT3 | [102] | |
| Hepatitis A virus (HAV) | CRISPR-ko | Brunello | HeLa | 39 genes including UGCG, GALE, and SLC35A2 | [103] |
| Hepatis B virus (HBV) | CRISPR-ko | Custom library of 19,050 genes, with 5 gRNAs/gene | HepG2 | 22 pro-HBsAg genes, including ZCCHC14, NXT1, and ENY2; 38 anti-HBsAg genes, including DCAF7, UBE2J1, RNF139, and UBE2J2 | [104] |
| Epstein-Barr virus | CRISPR-ko | AVANA | P3HR-1 | MYC, EP300, STAGA, FACT, cohesin subunits | [105] |
| CRISPR-ko | Brunello | Lymphoblastoid cell lines (LCL) | TAF family proteins, MEF2C | [106] | |
| CRISPR-ko | Avana | P3HR-1, GM12878 | 57 genes for P3HR1 and 87 genes for GM12878 | [107] | |
| Human cytomegalovirus (HCMV) | CRISPR-ko | GeCKO v2 | ARPE-19; HEL fibroblasts | OR14I1, PDGFRA | [108] |
| CRISPR-ko | GeCKO v2 | Human foreskin fibroblasts (HFF) | PDGFRA | [109] | |
| Influenza A virus (IAV) | CRISPR-ko | AVANA-4 | A549 | WDR7, CCDC115, TMEM199, CMTR1 | [110] |
| CRISPR-ko | GeCKO | A549 | SLC35A1 and other genes | [111] | |
| Adeno-associated virus | CRISPR-ko | GeCKO | Huh7 | GPR108, NEU1, GCNT4, CTSA | [112] |
| CRISPR-ko | GeCKO v2 | Huh7 | Crb3, CLDN15 | [113] | |
| Ebola virus (EBOV) | CRISPR-ko | GeCKO v2 | Huh7.5.1 | GNPTAB, NPC1, SPNS1, SLC30A1, HOPS complex, UVRAG | [114] |
| Enteroviruses (RV-C15 and non-polio EV-D68) | CRISPR-ko | GeCKO v2 | H1-HeLa cells | SETD3, CSDE1, PLA2G16 | [115] |
| Severe fever with thrombocytopenia syndrome virus | CRISPR-ko | GeCKO v2 | HeLa | SNX11 | [116] |
| Norovirus restriction factors | CRISPRa | Calabrese | Hela | TRIM7, PITX1, HOXC11, DDX60, MX1, PLSCR1 | [117] |
| Flaviviruses | CRISPR-ko | Brunello | Huh7.5 | IFI6, STAT2, IRF9 | [118] |
| CRISPR-ko | GeCKO v2 | 293T | STT3A, SEC63, SPCS1, SPCS3 | [119] | |
| West Nile virus | CRISPR-ko | Custom library of 77,406 gRNAs covering 20,121 genes | 293FT | EMC2, EMC3, SEL1L, DERL2, UBE2G2, UBE2J1, HRD1 | [120] |
| Arthritogenic alphaviruses (chikungunya, Ross River, Mayaro, O’nyong nyong) | CRISPR-ko | GeCKO v2 | 3T3 | MXRA8 | [121] |
| Rotavirus | CRISPR-ko | GeCKO | H1-Hela | SLC35A1, GNE, CMAS, UGCG, FA2H, LATS2, STAG2 | [122] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chulanov, V.; Kostyusheva, A.; Brezgin, S.; Ponomareva, N.; Gegechkori, V.; Volchkova, E.; Pimenov, N.; Kostyushev, D. CRISPR Screening: Molecular Tools for Studying Virus–Host Interactions. Viruses 2021, 13, 2258. https://doi.org/10.3390/v13112258
Chulanov V, Kostyusheva A, Brezgin S, Ponomareva N, Gegechkori V, Volchkova E, Pimenov N, Kostyushev D. CRISPR Screening: Molecular Tools for Studying Virus–Host Interactions. Viruses. 2021; 13(11):2258. https://doi.org/10.3390/v13112258
Chicago/Turabian StyleChulanov, Vladimir, Anastasiya Kostyusheva, Sergey Brezgin, Natalia Ponomareva, Vladimir Gegechkori, Elena Volchkova, Nikolay Pimenov, and Dmitry Kostyushev. 2021. "CRISPR Screening: Molecular Tools for Studying Virus–Host Interactions" Viruses 13, no. 11: 2258. https://doi.org/10.3390/v13112258