
Identification of a Chlorovirus PBCV-1 Protein Involved in Degrading the Host Cell Wall during Virus Infection


 , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses and Host Strains
2.2. Recombinant A561L and A561LD4
2.3. vLysin Preparation and Activity Assay
2.4. Cell Wall Degrading Activity Assayed by Chlorophyll Release
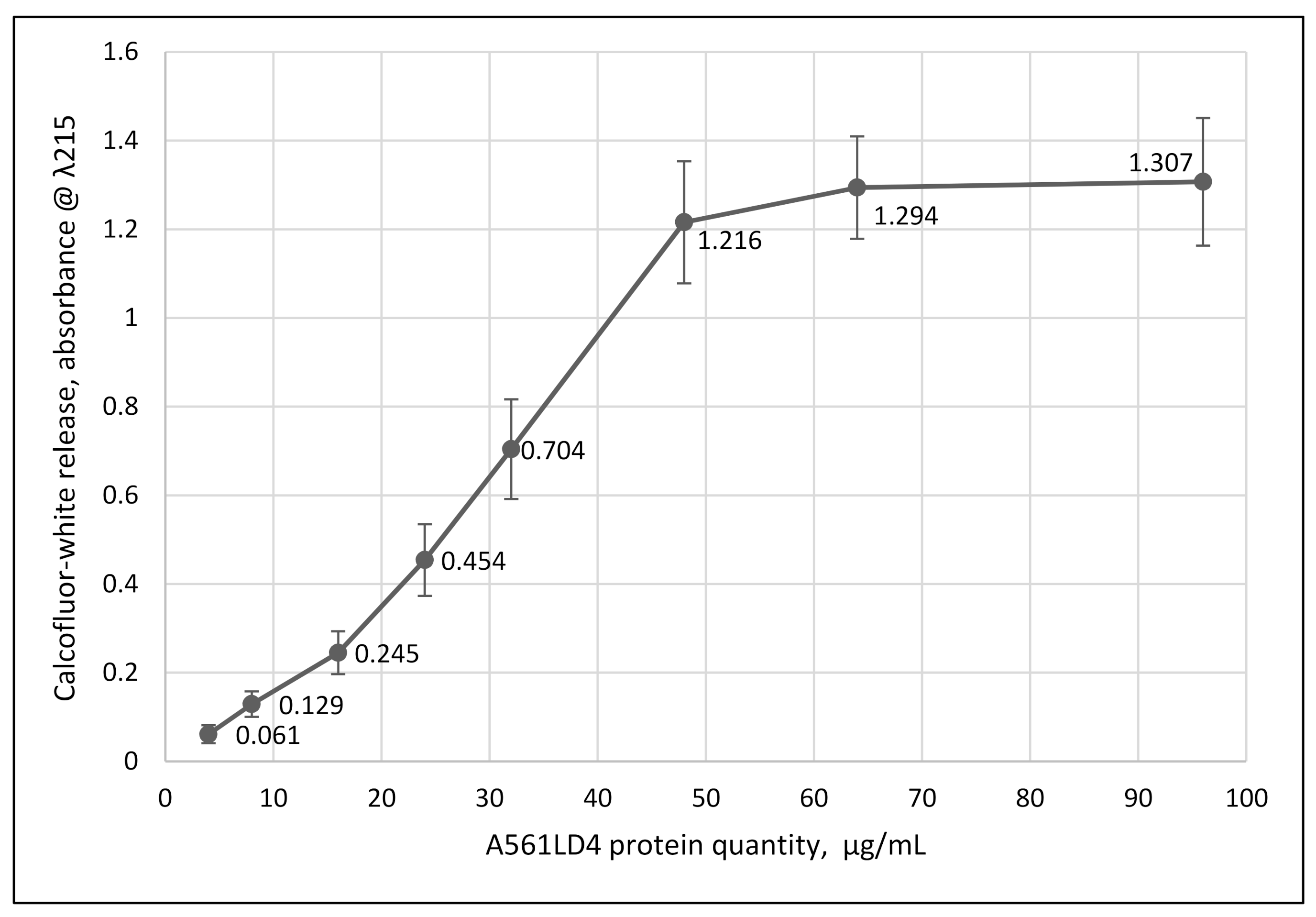
2.5. Protein Activity Assayed by Calcofluor-White (CFW) Release
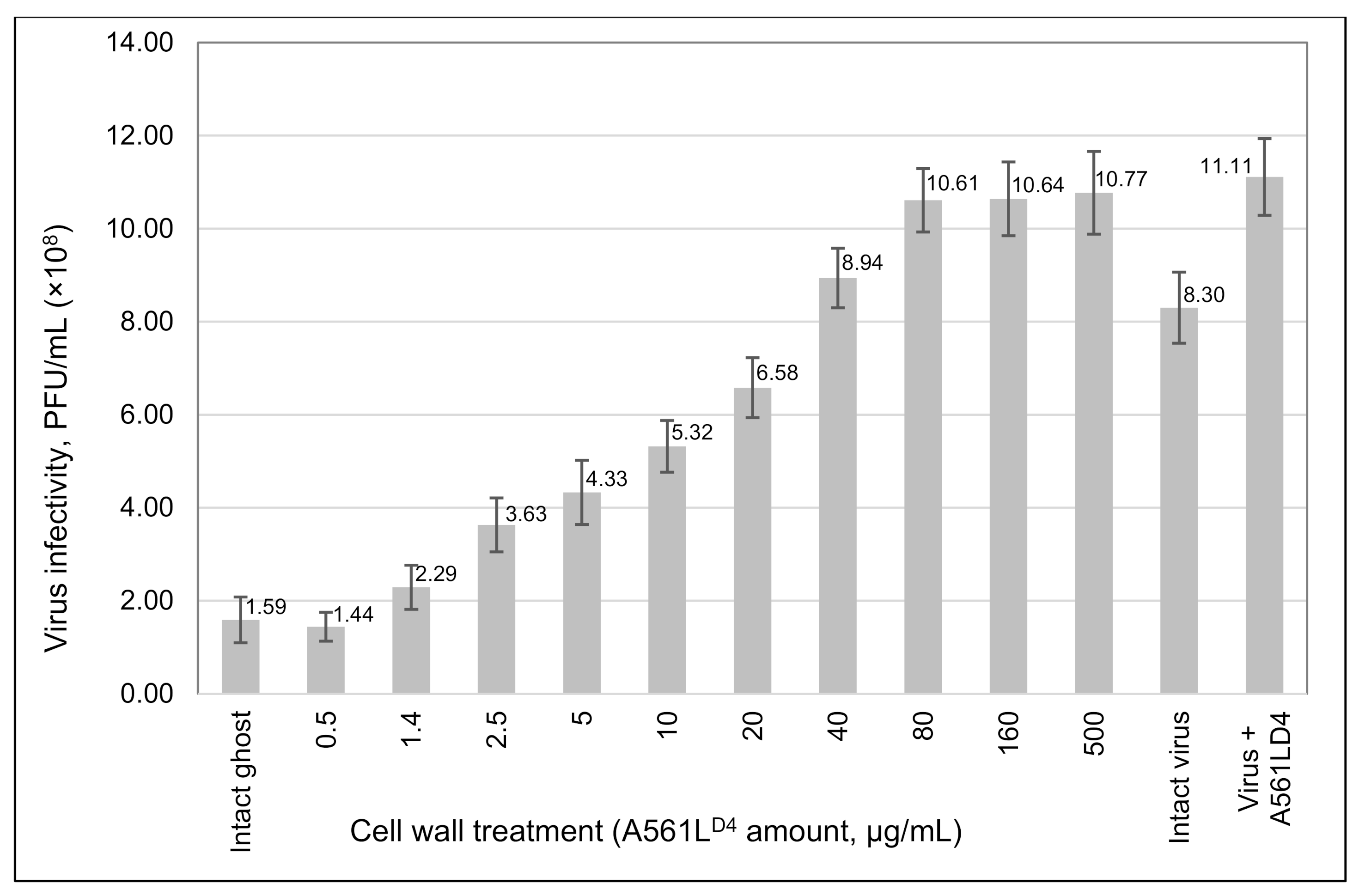
2.6. Effect of A561LD4 Protein Treatment of C. variabilis Ghost Cells on PBCV-1 Virus Attachment
2.7. Effect of Soluble Products (Possibly Receptors) Resulting from A561LD4 Treatment of C. variabilis NC64A Ghost Cells on PBCV-1 Virus Infectivity
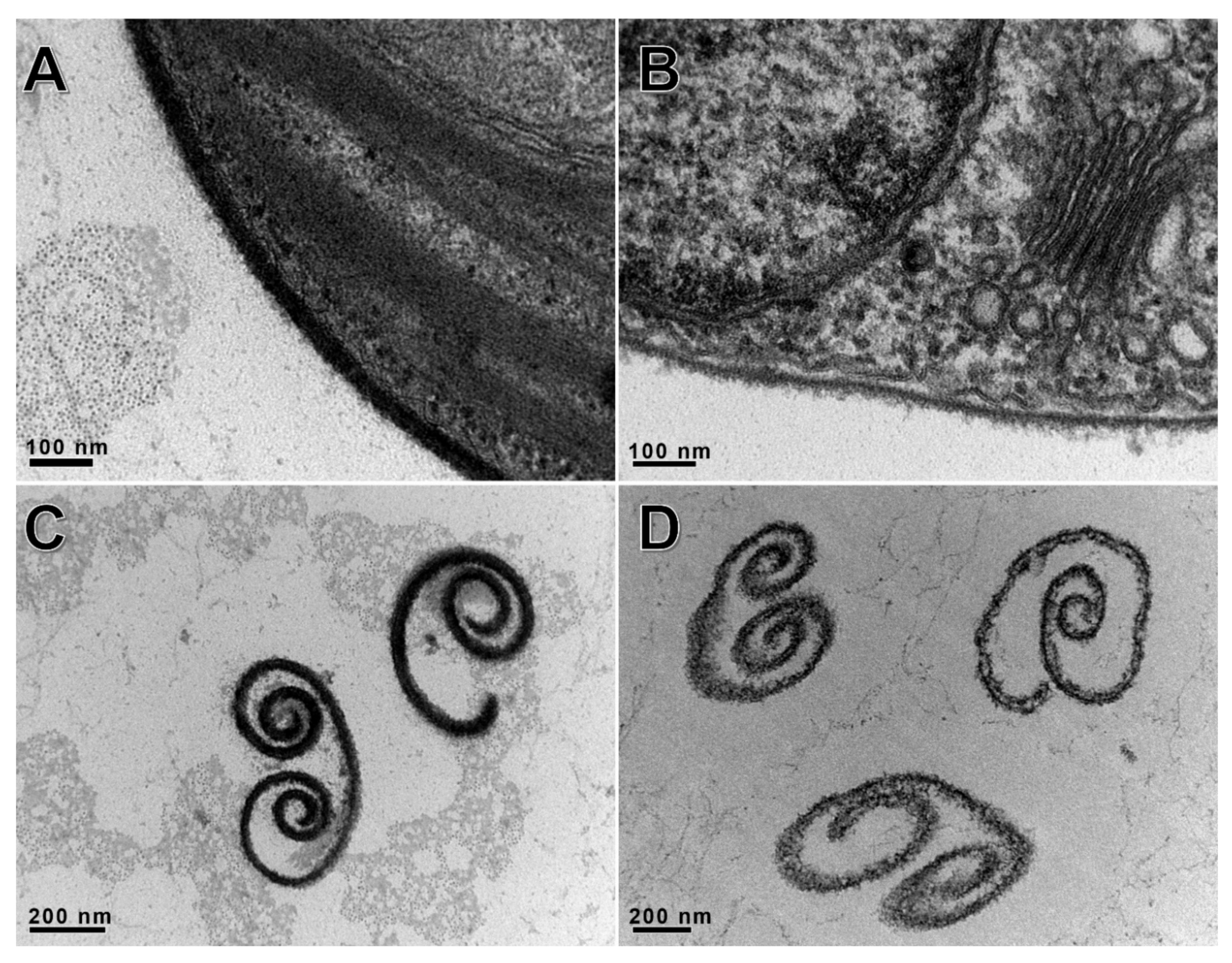
2.8. Electron Microscopy
2.9. SDS-PAGE and Western Blots
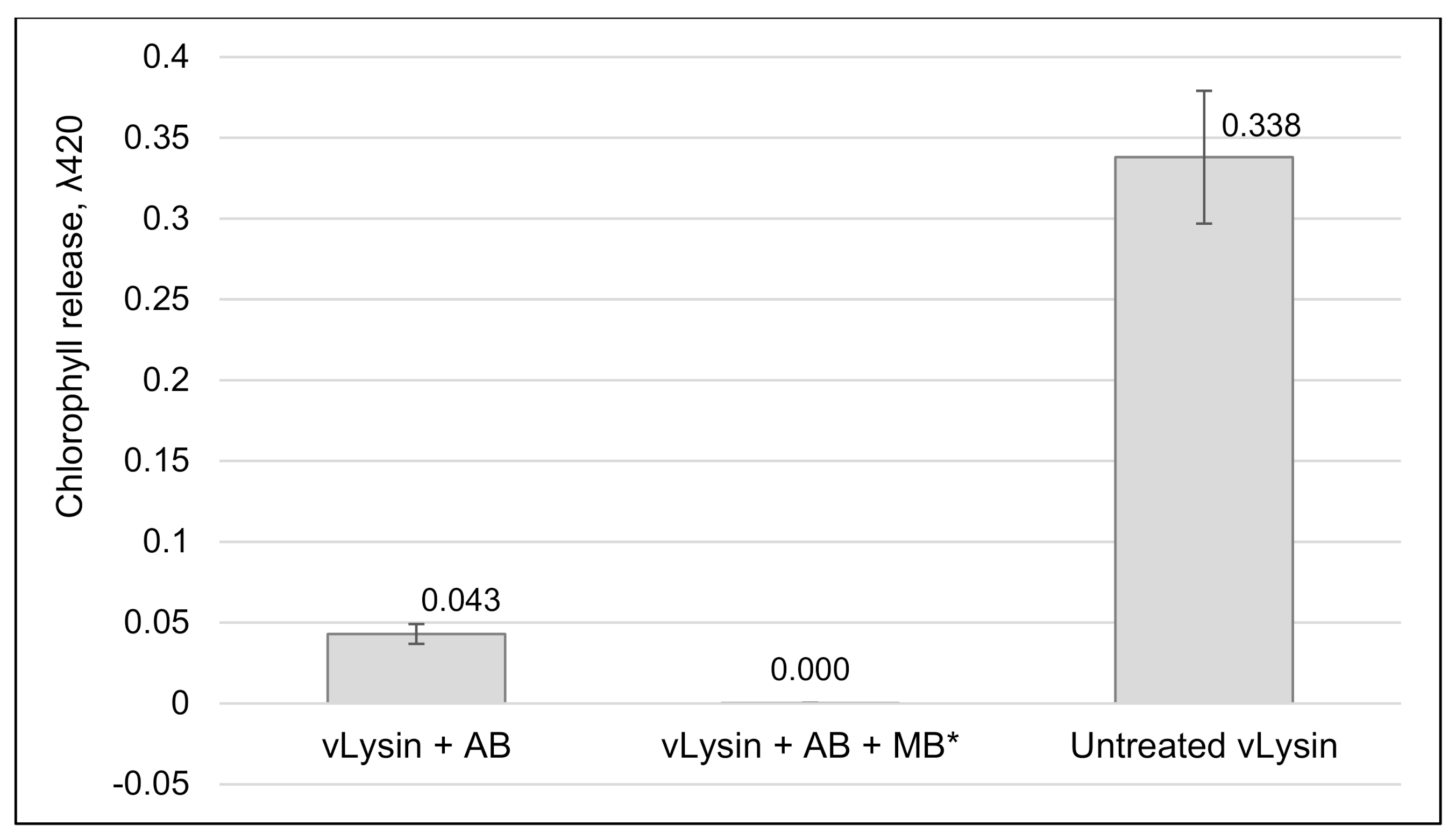
2.10. Effect of Anti A561LD4 Antibody on vLysin Activity
2.11. Phylogenetic Analyses
3. Results and Discussion
3.1. PBCV-1 A561L Is a Candidate for Degrading the Host Cell Wall
3.2. Development of Assays for Chlorella Cell Wall Degrading Activity
3.3. Expression and Purification of A561LD4
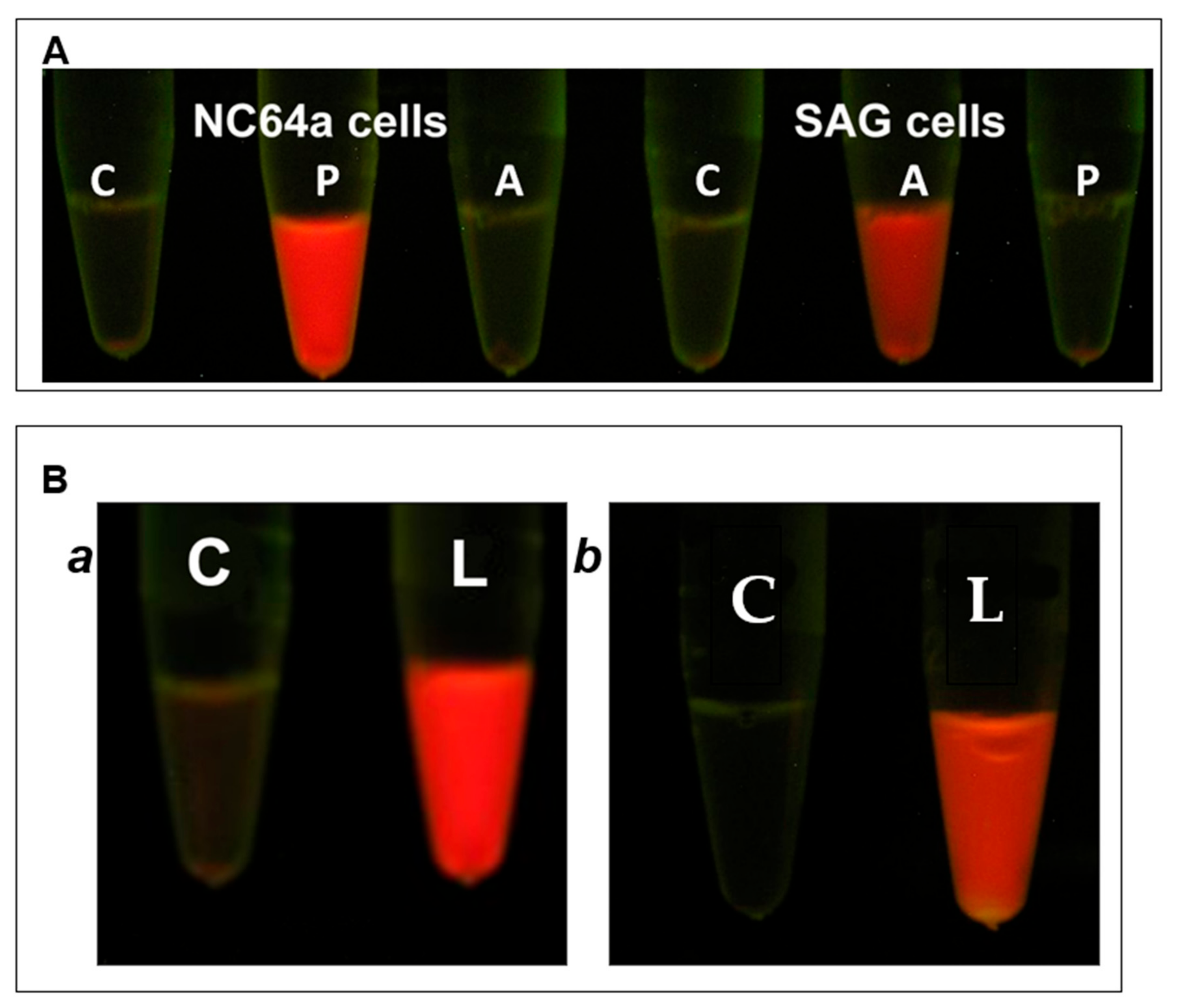
3.4. Wall Degradation by A561LD4
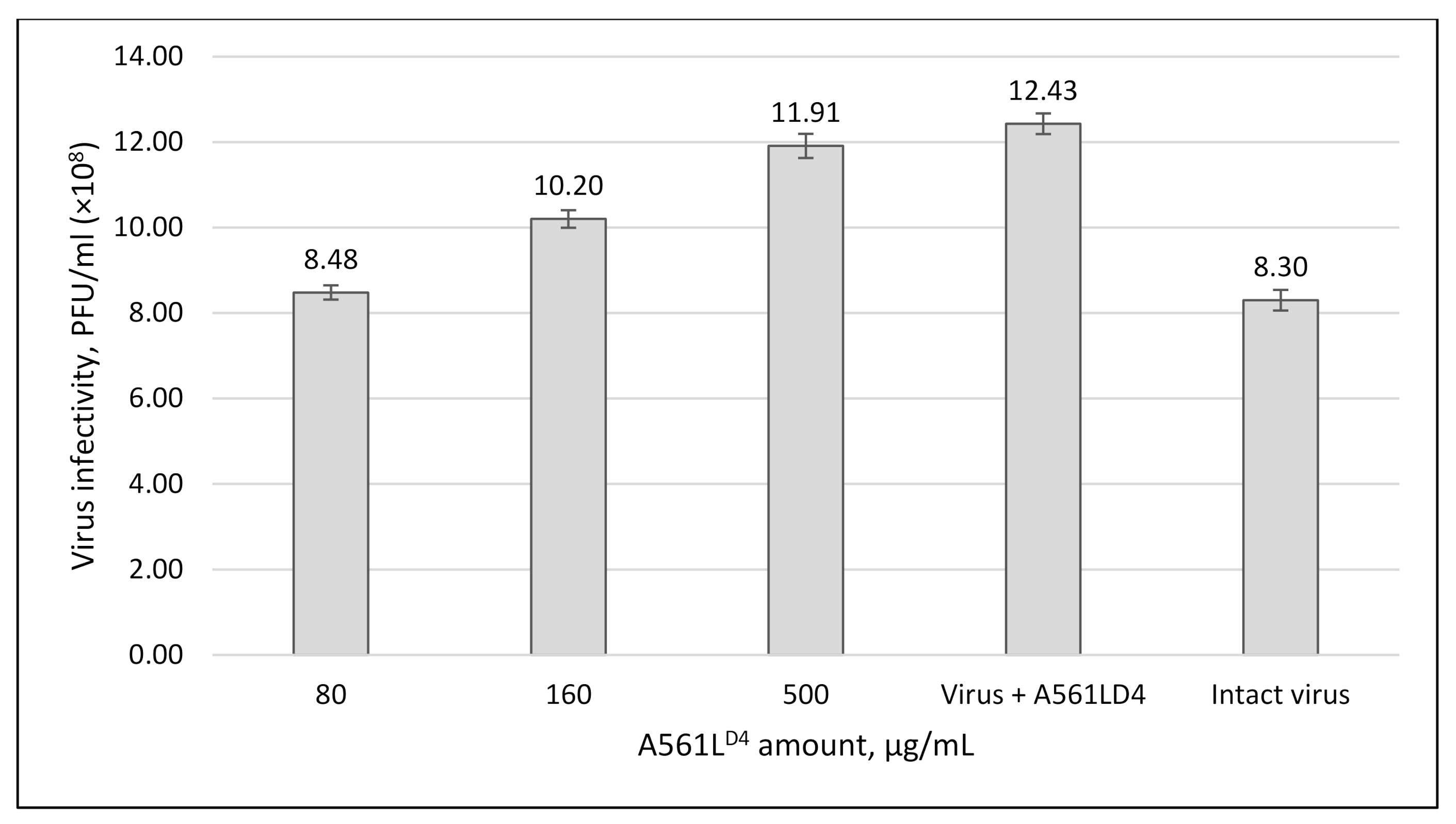
3.5. A561LD4 Treatment Increased PBCV-1 Infectivity
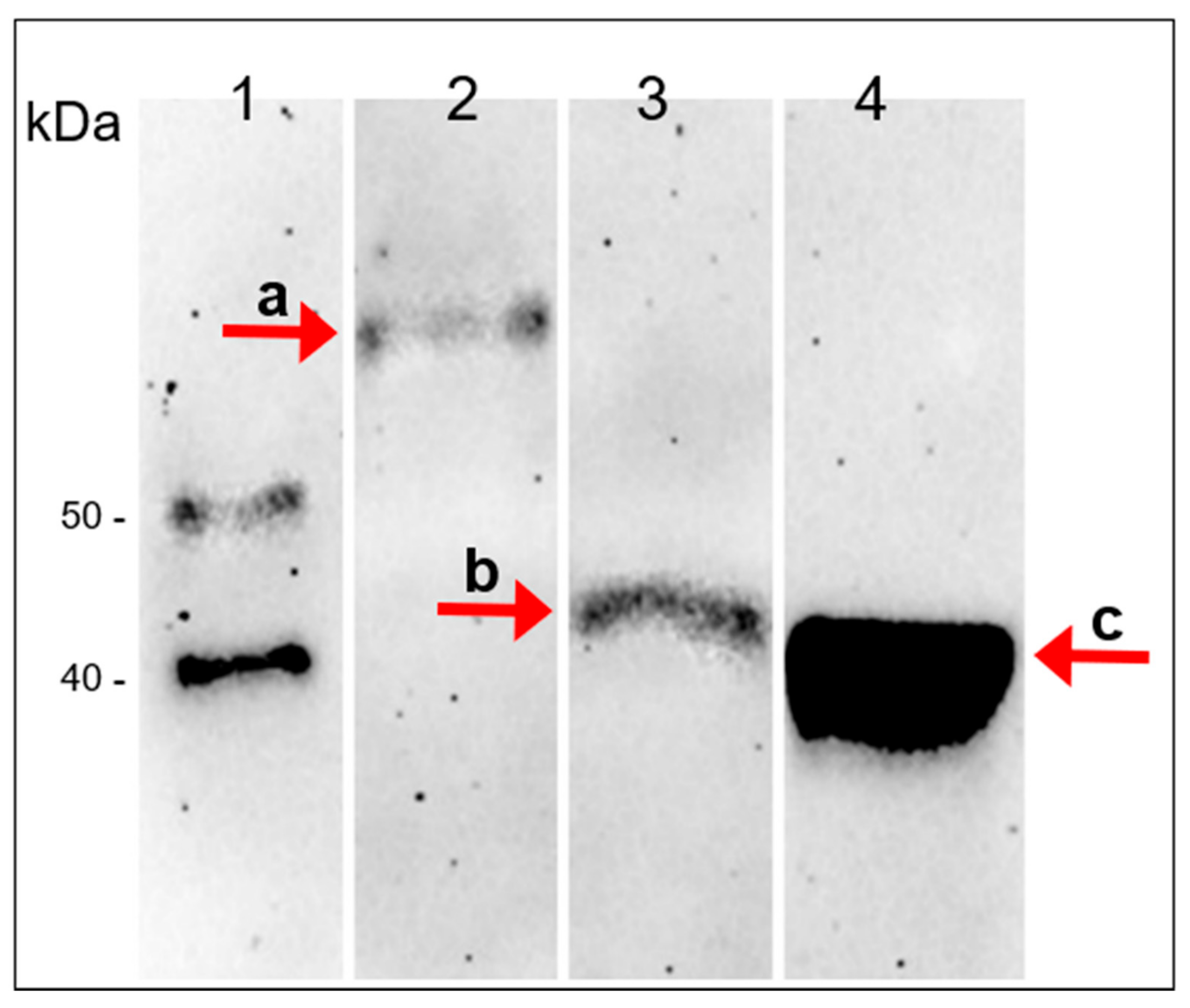
3.6. Immunological Detection of the A561L Protein in the Virion
3.7. A561L Is Located Inside the PBCV-1 Virion
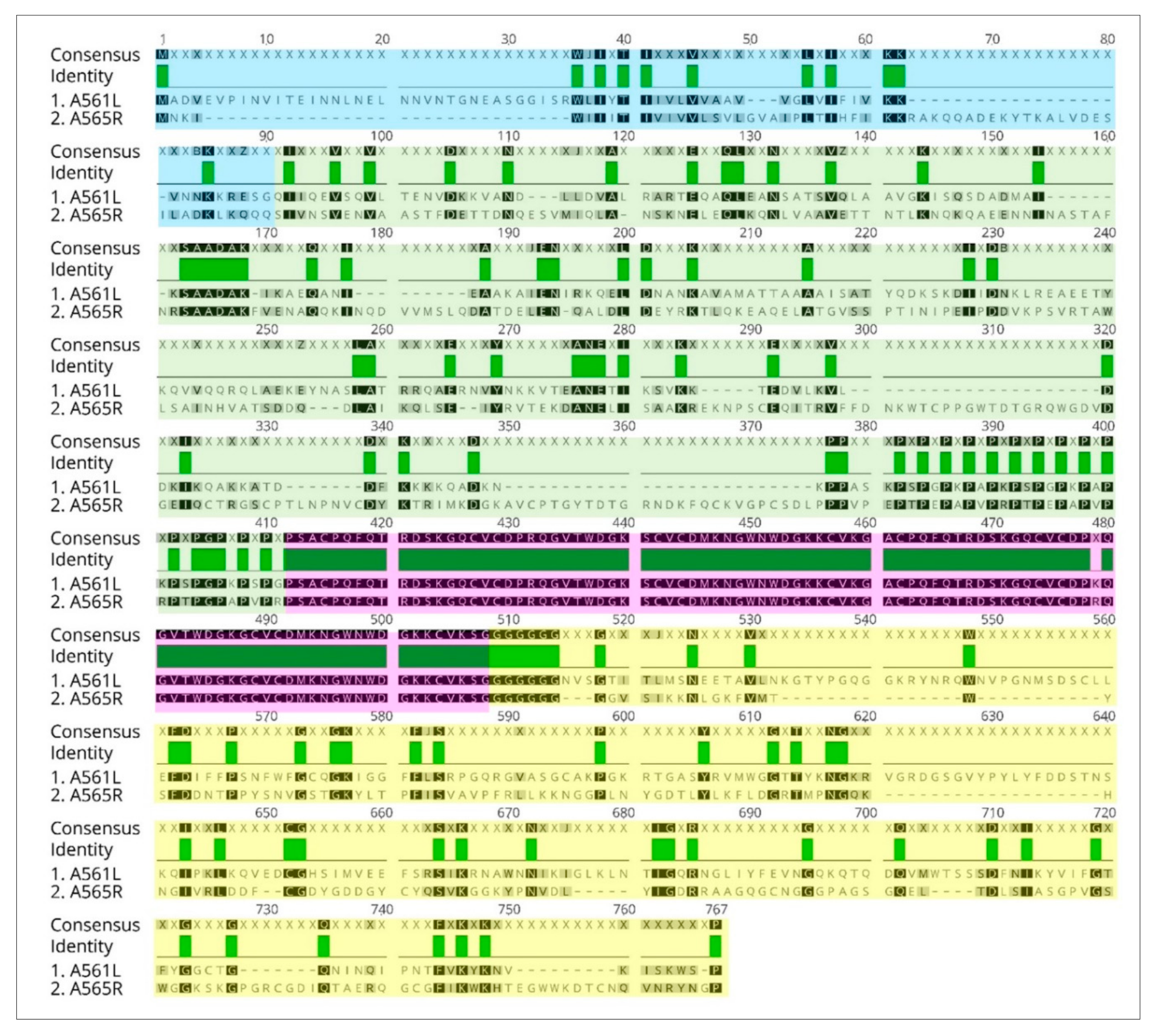
3.8. A561LD4 Homologs Are Common among the Chloroviruses
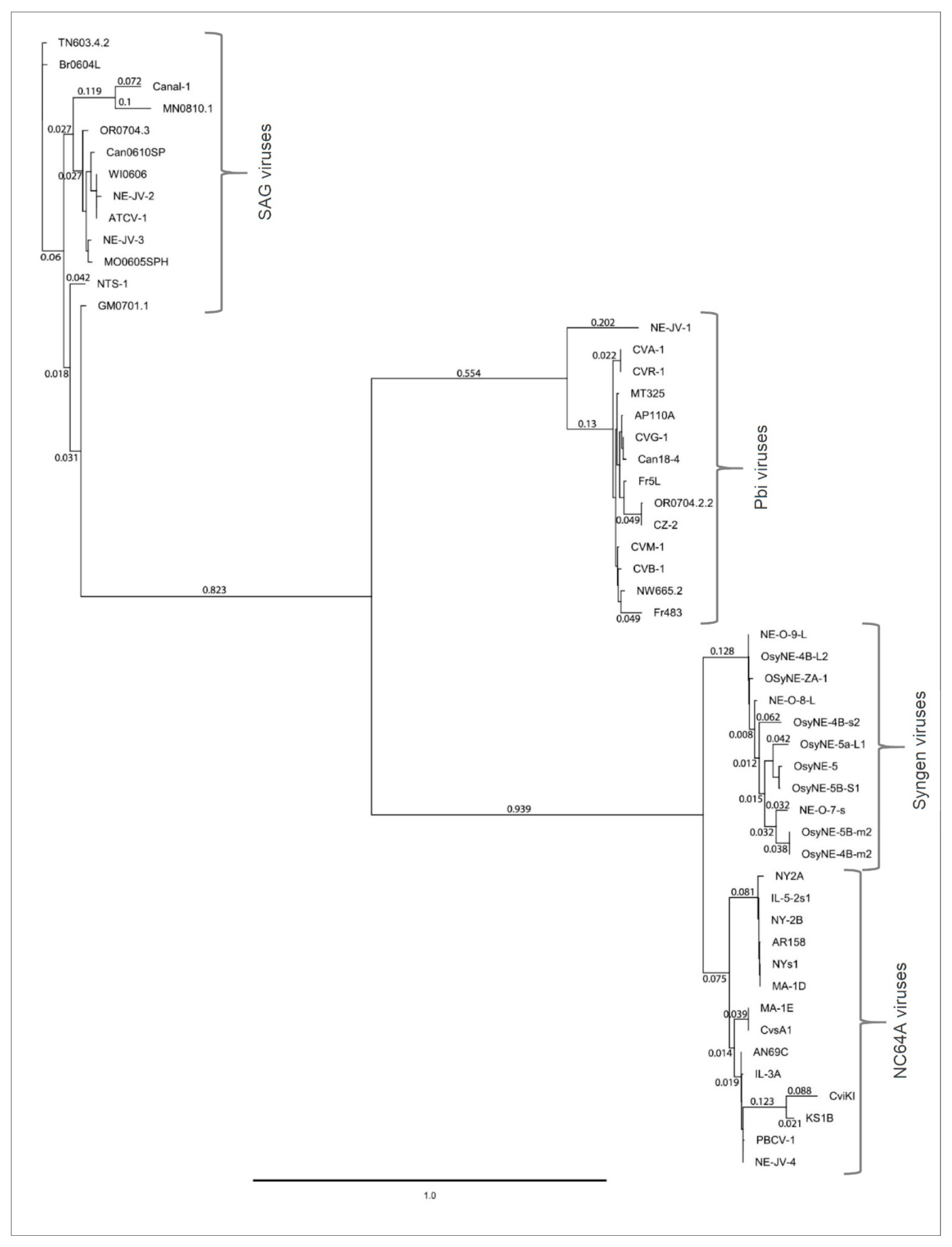
3.9. Phylogenetic Analysis of A561L
3.10. A561LD4 Degrades the Cell Wall of Several Chlorella Species
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Template, PDB | Template Information | Confidence, % | % i.d. | % of Coverage |
|---|---|---|---|---|
| c5gmtB | crystal structure of the marine pl-14 alginate lyase from Aplysia kurodai | 100 | 23 | 85 |
| c6kcvB | structure of alginate lyase from Chitinophaga sp. MD30 aly36b mutant k143a/y185a in complex with alginate tetrasaccharide | 100 | 23 | 87 |
| c3im0A | crystal structure of chlorella virus Val-1 | 100 | 30 | 87 |
| c2zzjA | crystal structure of endo-beta-1,4-glucuronan lyase from fungus Trichoderma reesei | 99.7 | 16 | 79 |


References
- Nelson, D.C.; Schmelcher, M.; Rodriguez-Rubio, L.; Klumpp, J.; Pritchard, D.G.; Dong, S.; Donovan, D.M. Endolysins as antimicrobials. Adv. Virus Res. 2012, 83, 299–365. [Google Scholar]
- Carter, J.B.; Saunders, V.A. Virology: Principles and Applications, 2nd ed.; John Wiley & Sons: Hoboken, NJ, USA, 2014; p. 400. [Google Scholar]
- Van Etten, J.L.; Agarkova, I.V.; Dunigan, D.D. Chloroviruses. Viruses 2020, 12, 20. [Google Scholar] [CrossRef] [Green Version]
- Proschold, T.; Darienko, T.; Silva, P.C.; Reisser, W.; Krienitz, L. The systematics of zoochlorella revisited employing an integrative approach. Environ. Microbiol. 2011, 13, 350–364. [Google Scholar] [CrossRef]
- Hoshina, R.; Iwataki, M.; Imamura, N. Chlorella variabilis and Micractinium reisseri sp. nov. (Chlorellaceae, Trebouxiophyceae): Redescription of the endosymbiotic green algae of Paramecium bursaria (Peniculia, Oligohymenophorea) in the 120th year. Psychol. Res. 2010, 58, 188–201. [Google Scholar]
- Quispe, C.F.; Esmael, A.; Sonderman, O.; McQuinn, M.; Agarkova, I.; Battah, M.; Duncan, G.A.; Dunigan, D.D.; Smith, T.P.L.; De Castro, C.; et al. Characterization of a new chlorovirus type with permissive and non-permissive features on phylogenetically related algal strains. Virology 2017, 500, 103–113. [Google Scholar] [CrossRef]
- Arriola, M.B.; Velmurugan, N.; Zhang, Y.; Plunkett, M.H.; Hondzo, H.; Barney, B.M. Genome sequences of Chlorella sorokiniana UTEX 1602 and Micractinium conductrix SAG 241.80: Implications to maltose excretion by a green alga. Plant J. 2018, 93, 566–586. [Google Scholar] [CrossRef] [Green Version]
- Cherrier, M.V.; Kostyuchenko, V.A.; Xiao, C.; Bowman, V.D.; Battisti, A.J.; Yan, X.; Chipman, P.R.; Baker, T.S.; Van Etten, J.L.; Rossmann, M.G. An icosahedral algal virus has a complex unique vertex decorated by a spike. Proc. Natl. Acad. Sci. USA 2009, 106, 11085–11089. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Xiang, Y.; Dunigan, D.D.; Klose, T.; Chipman, P.R.; Van Etten, J.L.; Rossmann, M.G. Three-dimensional structure and function of the Paramecium bursaria chlorella virus capsid. Proc. Natl. Acad. Sci. USA 2011, 108, 14837–14842. [Google Scholar] [CrossRef] [Green Version]
- Meints, R.H.; Lee, K.; Burbank, D.E.; Van Etten, J.L. Infection of a chlorella-like alga with the virus, PBCV-1: Ultrastructural studies. Virology 1984, 138, 341–346. [Google Scholar] [CrossRef]
- Milrot, E.; Shimoni, E.; Dadosh, T.; Rechav, K.; Unger, T.; Van Etten, J.L.; Minsky, A. Structural studies demonstrating a bacteriophage-like replication cycle of the eukaryote-infecting Paramecium bursaria chlorella virus-1. PLoS Pathog. 2017, 13, e1006562. [Google Scholar] [CrossRef] [Green Version]
- Frohns, F.; Käsmann, A.; Kramer, D.; Schäfer, B.; Mehmel, M.; Kang, M.; Van Etten, J.L.; Gazzarrini, S.; Moroni, A.; Thiel, G. Potassium ion channels of chlorella viruses cause rapid depolarization of host cells during infection. J. Virol. 2006, 80, 2437–2444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neupärtl, M.; Meyer, C.; Woll, I.; Frohns, F.; Kang, M.; Van Etten, J.L.; Kramer, D.; Hertel, B.; Moroni, A.; Thiel, G. Chlorella viruses evoke a rapid release of K+ from host cells during early phase of infection. Virology 2008, 372, 340–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thiel, G.; Moroni, A.; Dunigan, D.; Van Etten, J.L. Initial events associated with virus PBCV-1 infection of Chlorella NC64A. In Progress in Botany; Lüttge, U., Beyschlag, W., Büdel, B., Eds.; Springer: Berlin, Germany, 2010; pp. 169–183. [Google Scholar]
- Meints, R.H.; Lee, K.; Van Etten, J.L. Assembly site of the virus PBCV-1 in a Chlorella-like green alga: Ultrastructural studies. Virology 1986, 154, 240–245. [Google Scholar] [CrossRef]
- Dunigan, D.D.; Cerny, R.L.; Bauman, A.T.; Roach, J.C.; Lane, L.C.; Agarkova, I.V.; Wulser, K.; Yanai-Balser, G.M.; Gurnon, J.R.; Vitek, J.C.; et al. Paramecium bursaria Chlorella Virus 1 proteome reveals novel architectural and regulatory features of a giant virus. J. Virol. 2012, 86, 8821–8834. [Google Scholar] [CrossRef] [Green Version]
- Hiramatsu, S.; Ishihara, M.; Fujie, M.; Usami, S.; Yamada, T. Expression of a chitinase gene and lysis of the host cell wall during Chlorella virus CVK2 infection. Virology 1999, 260, 308–315. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Adams, B.; Gurnon, J.; Ye, Y.; Van Etten, J.L. Characterization of two chitinase genes and one chitosanase gene encoded by chlorella virus PBCV-1. Virology 1999, 263, 376–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, T.; Hiramatsu, S.; Songsri, P.; Fujie, M. Alternative expression of chitosanase gene produces two different proteins in cells infected with Chlorella virus CVK2. Virology 1997, 230, 361–368. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Gurnon, J.R.; Adams, B.J.; Graves, M.V.; Van Etten, J.L. Characterization of a β-1,3-glucanase encoded by chlorella virus PBCV-1. Virology 2000, 276, 27–36. [Google Scholar] [CrossRef] [Green Version]
- Ogura, K.; Yamasaki, M.; Yamada, T.; Mikami, B.; Hahimoto, W.; Murata, K. Crystal structure of family 14 polysaccharide lyase with pH-dependent modes of action. J. Biol. Chem. 2009, 284, 35572–35579. [Google Scholar] [CrossRef] [Green Version]
- Sugimoto, I.; Onimatsu, H.; Fujie, M.; Usami, S.; Yamada, T. vAL-1, a novel polysaccharide lyase encoded by chlorovirus CVK2. FEBS Lett. 2004, 559, 51–56. [Google Scholar] [CrossRef] [Green Version]
- Van Etten, J.L.; Agarkova, I.; Dunigan, D.D.; Tonetti, M.; De Castro, C.; Duncan, G.A. Chloroviruses have a sweet tooth. Viruses 2017, 9, 88. [Google Scholar] [CrossRef]
- Meints, R.H.; Burbank, D.E.; Van Etten, J.L.; Lamport, D.T. Properties of the chlorella receptor for the virus PBCV-1. Virology 1988, 164, 15–21. [Google Scholar] [CrossRef]
- Agarkova, I.; Hertel, B.; Zhang, X.; Lane, L.; Tchourbanov, A.; Dunigan, D.D.; Thiel, G.; Rossmann, M.G.; Van Etten, J.L. Dynamic attachment of Chlorovirus PBCV-1 to Chlorella variabilis. Virology 2014, 466–467, 95–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Etten, J.L.; Burbank, D.E.; Kuczmarski, D.; Meints, R.H. Virus infection of culturable chlorella-like algae and dlevelopment of a plaque assay. Science 1983, 219, 994–996. [Google Scholar] [CrossRef] [PubMed]
- Reisser, W.; Becker, B.; Klein, T. Studies on ultrastructure and host range of a Chlorella attacking virus. Protoplasma 1986, 135, 162–165. [Google Scholar] [CrossRef]
- Agarkova, I.V.; Dunigan, D.D.; Van Etten, J.L. Virion-associated restriction endonucleases of chloroviruses. J. Virol. 2006, 80, 8114–8123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Etten, J.L.; Burbank, D.E.; Xia, Y.; Meints, R.H. Growth cycle of a virus, PBCV- 1, that infects chlorella-like algae. Virology 1983, 126, 117–125. [Google Scholar] [CrossRef]
- Kennedy, J.E. Purification and characterization of a lytic enzyme from viral infection of Chlorella-like green algae. Masters’s Thesis, University of Nebraska, Lincoln, Nebraska, 1987. [Google Scholar]
- Milrot, E.; Mutsafi, Y.; Fridmann-Sirkis, Y.; Shimoni, E.; Rechav, K.; Gurnon, J.R.; Van Etten, J.L.; Minsky, A. Virus–host interactions: Insights from the replication cycle of the large Paramecium bursaria chlorella virus. Cell. Microbiol. 2016, 18, 3–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skrdla, M.P.; Burbank, D.E.; Xia, Y.; Meints, R.H.; Van Etten, J.L. Structural proteins and lipids in a virus, PBCV-1, which replicates in a Chlorella-like alga. Virology 1984, 135, 308–315. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate Maximum-Likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- UniProt, C. UniProt: A worldwide hub of protein knowledge. Nucleic Acids Res. 2019, 47, D506–D515. [Google Scholar]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Sugimoto, I.; Hiramatsu, S.; Murakami, D.; Fujie, M.; Usami, S.; Yamada, T. Algal-lytic activities encoded by chlorella virus CVK2. Virology 2000, 277, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, A.; Chang, H.-Y.; Daugherty, L.; Fraser, M.; Hunter, S.; Lopez, R.; McAnulla, C.; McMenamin, C.; Nuka, G.; Pesseat, S.; et al. The InterPro protein families database: The classification resource after 15 years. Nucleic Acids Res. 2015, 43, D213–D221. [Google Scholar] [CrossRef]
- Imam, S.H.; Buchanan, M.J.; Shin, H.C.; Snell, W.J. The Chlamydomonas cell wall: Characterization of the wall framework. J. Cell Biol. 1985, 101, 1599–1607. [Google Scholar] [CrossRef] [Green Version]
- Yamada, T.; Sakaguchi, K. Comparative studies on Chlorella cell walls: Induction of protoplast formation. Arch. Microbiol. 1982, 132, 10–13. [Google Scholar] [CrossRef]










| Species | Strain Designation | Chlorophyll Release, λ = 420 # |
|---|---|---|
| Auxenochlorella protothecoides | stock 29 | 0.840 |
| Chlorella heliozoae | SAG 3.83 | 0.570 |
| Chlorella miniata | UTEX 490 | 0.198 |
| Chlorella sorokiniana | UTEX 246 | 0.335 |
| Chlorella sorokiniana | UTEX 1810 | 0.294 |
| Chlorella sorokiniana | UTEX 1230 | 0 |
| Chlorella sorokiniana | CS01 China | 0 |
| Chlorella variabilis | NC64A | 0.267 |
| Chlorella variabilis | NIES 2540 | 0.236 |
| Chlorella variabilis | NIES 2541 | 0.368 |
| Chlorella variabilis | Syngen 2-3 | 0.229 |
| Chlorella vulgaris | UTEX 395 | 0.687 |
| Chlorella sp. | UTEX BSN 069 | 0.681 |
| Chlorella sp. | UTEX BSN069 | 0.507 |
| Chloroidium saccharophilum | UTEX 247 | 0 |
| Coccomyxa subellipsoidea | C169 | 0 |
| Micractinium conductrix | Pbi | 0.376 |
| Parachlorella kessleri | UTEX 2228 | 0 |
| Scenedesmus dimorphus | UTEX 417 | 0.392 |
| Scenedesmus dimorphus | UTEX 1237 | 0 |
| Scenedesmus obliquus | UTEX B2630 | 0 |
| Scenedesmus obliquus | UTEX 393 | 0 |
| Scenedesmus obliquus | UTEX 1450 | 0 |
| Scenedesmus parisiensis | UTEX 1585 | 0 |
| Scenedesmus sp | UTEX 1590 | 0 |
| Scenedesmus sp | UTEX 1589 | 0 |
| Scenedesmus sp | UTEX 2193 | 0 |
| Scenedesmus sp | UTEX 1591 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agarkova, I.V.; Lane, L.C.; Dunigan, D.D.; Quispe, C.F.; Duncan, G.A.; Milrot, E.; Minsky, A.; Esmael, A.; Ghosh, J.S.; Van Etten, J.L. Identification of a Chlorovirus PBCV-1 Protein Involved in Degrading the Host Cell Wall during Virus Infection. Viruses 2021, 13, 782. https://doi.org/10.3390/v13050782
Agarkova IV, Lane LC, Dunigan DD, Quispe CF, Duncan GA, Milrot E, Minsky A, Esmael A, Ghosh JS, Van Etten JL. Identification of a Chlorovirus PBCV-1 Protein Involved in Degrading the Host Cell Wall during Virus Infection. Viruses. 2021; 13(5):782. https://doi.org/10.3390/v13050782
Chicago/Turabian StyleAgarkova, Irina V., Leslie C. Lane, David D. Dunigan, Cristian F. Quispe, Garry A. Duncan, Elad Milrot, Abraham Minsky, Ahmed Esmael, Jayadri S. Ghosh, and James L. Van Etten. 2021. "Identification of a Chlorovirus PBCV-1 Protein Involved in Degrading the Host Cell Wall during Virus Infection" Viruses 13, no. 5: 782. https://doi.org/10.3390/v13050782